Synthetic Human β Defensin-3-C15 Peptide in Endodontics: Potential Therapeutic Agent in Streptococcus gordonii Lipoprotein-Stimulated Human Dental Pulp-Derived Cells

 , , ,
, , , 
Abstract
:1. Introduction
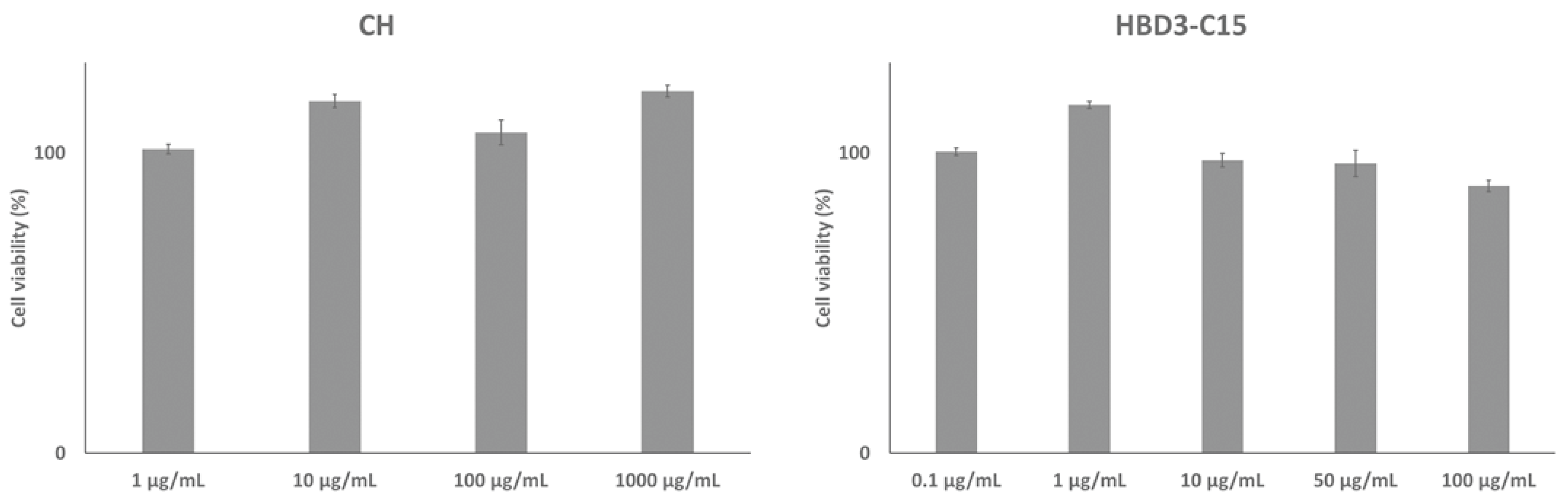
2. Results
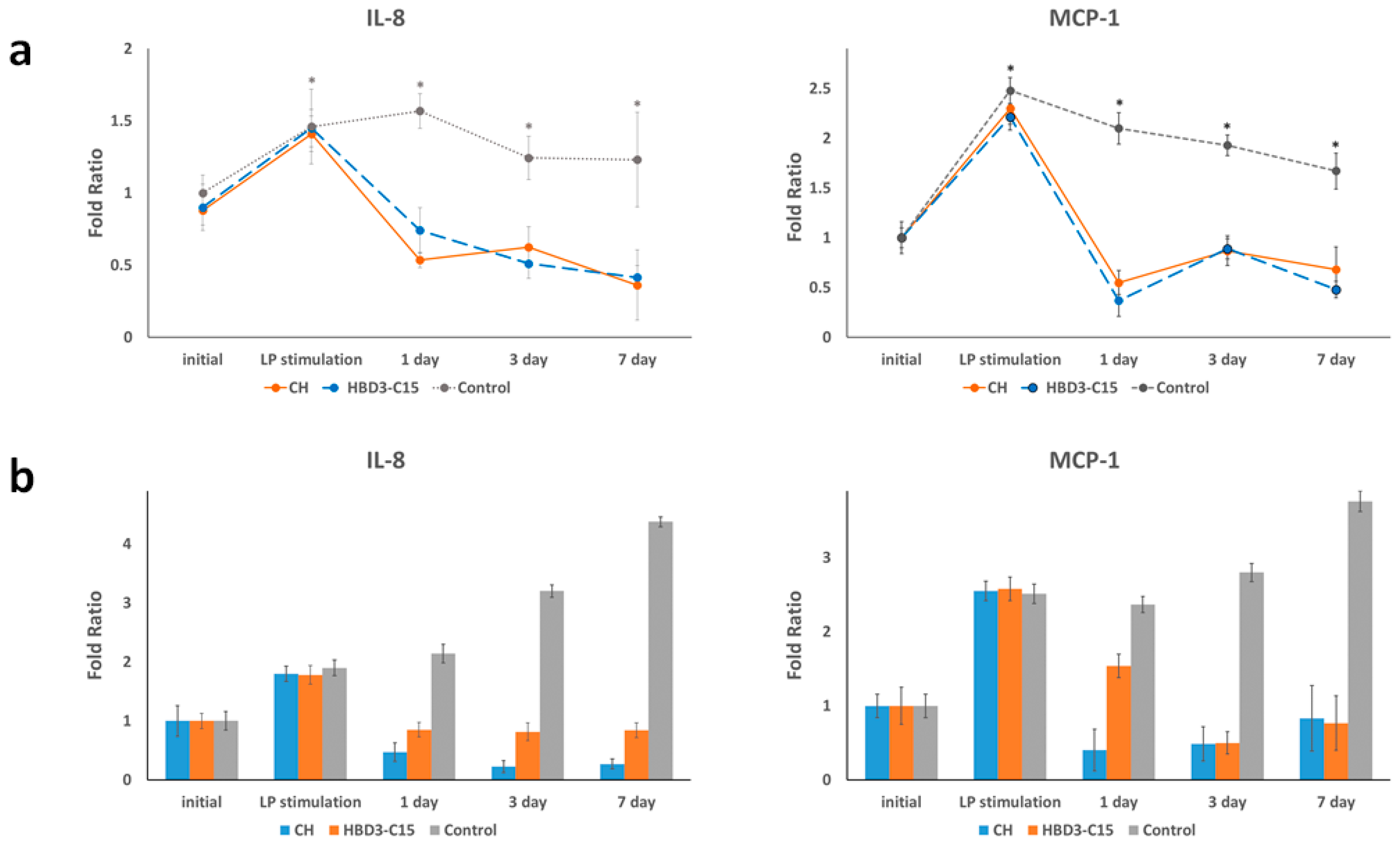
2.1. S. gordoni Lipoproteins Induced Inflammatory Mediators
2.2. HBD3-C15 and CH Suppress Inflammatory Mediators
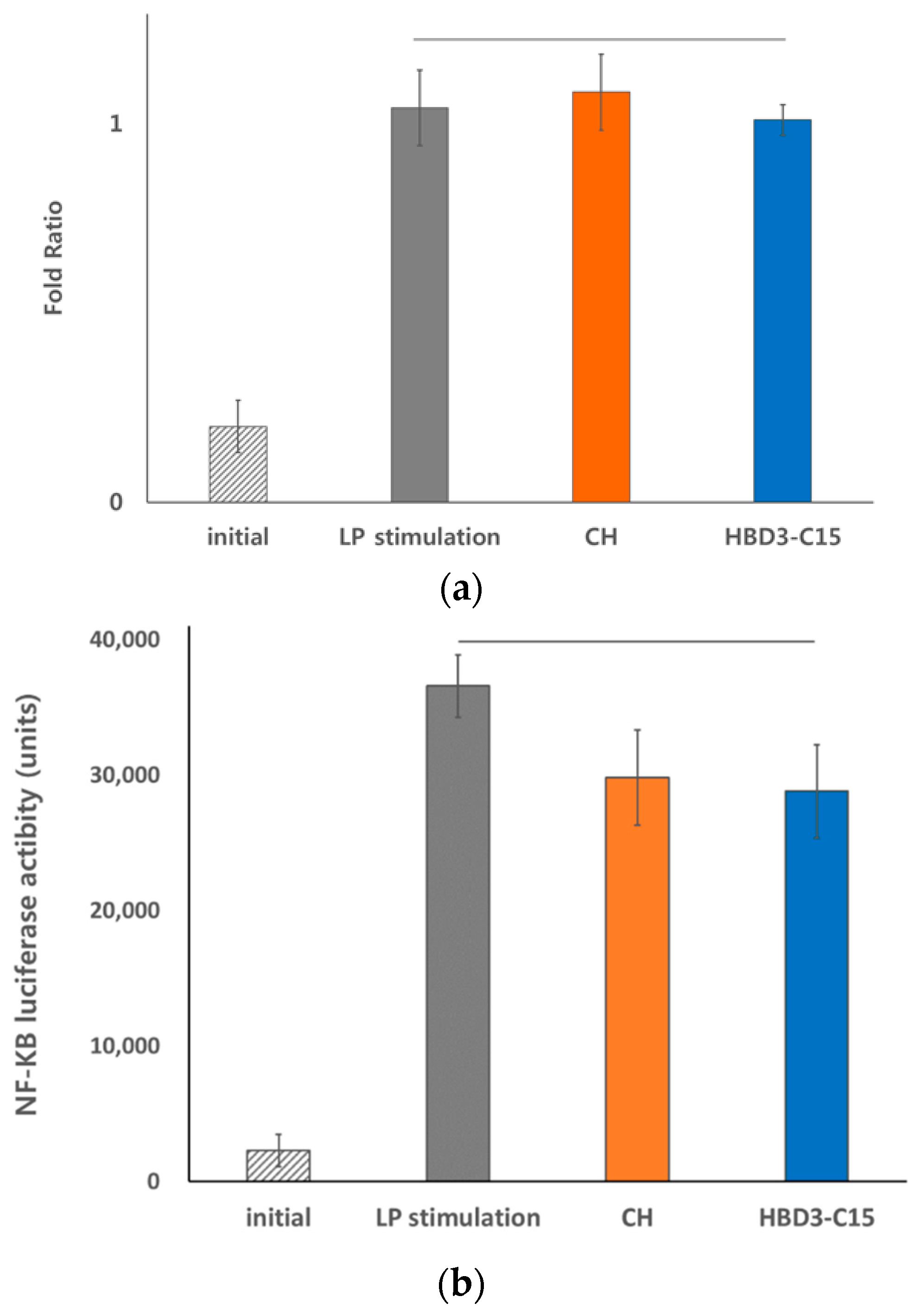
2.3. Anti-Inflammatory Effects of HBD3-C15 and CH Were not Mediated by TLR2
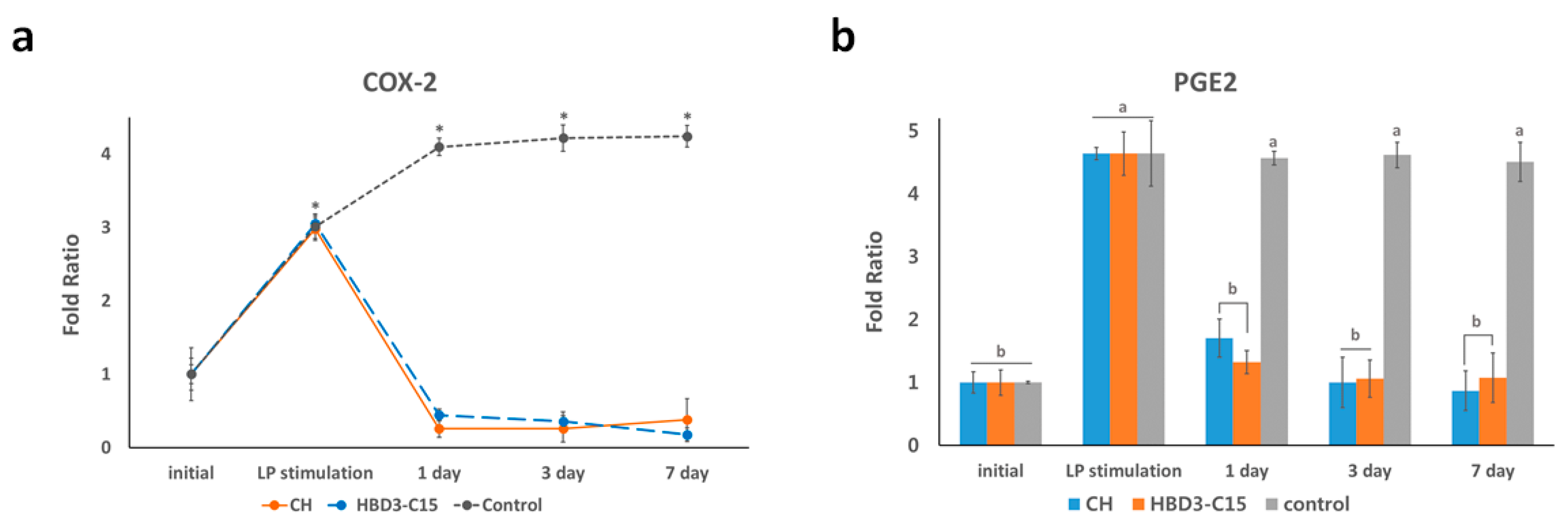
2.4. Anti-Inflammatory Effects Were Related to COX-2 Expression and PGE2 Level
3. Discussion
4. Materials and Methods
4.1. Human Dental Pulp Cell Explant Cultures
4.2. S. gordonii Lipoprotein Extracts
4.3. Cell Transfection
4.4. Peptide Preparation
4.5. Inflammatory Chemokine Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Ethical Approval
References
- Love, R.M.; Jenkinson, H.F. Invasion of dentinal tubules by oral bacteria. Crit. Rev. Oral. Biol. Med. 2002, 13, 171–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedgley, C.M.; Lee, E.H.; Martin, M.J.; Flannagan, S.E. Antibiotic resistance gene transfer between Streptococcus gordonii and Enterococcus faecalis in root canals of teeth ex vivo. J. Endod. 2008, 34, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Brightbill, H.D.; Libraty, D.H.; Krutzik, S.R.; Yang, R.B.; Belisle, J.T.; Bleharski, J.R.; Maitland, M.; Norgard, M.V.; Plevy, S.E.; Smale, S.T.; et al. Host defense mechanisms triggered by microbial lipoproteins through toll-like receptors. Science 1999, 285, 732–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, M.T.; Gotz, F. Lipoproteins of Gram-Positive Bacteria: Key Players in the Immune Response and Virulence. Microbiol. Mol. Biol. Rev. 2016, 80, 891–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.L.; Phillips, C.M.; Townsend, R.A.; Halperin, S.A.; Lee, S.F. Differential activation of dendritic cells by toll-like receptor agonists isolated from the gram-positive vaccine vector Streptococcus gordonii. Scand. J. Immunol. 2009, 69, 351–356. [Google Scholar] [CrossRef]
- Kim, A.R.; Ahn, K.B.; Kim, H.Y.; Seo, H.S.; Kum, K.Y.; Yun, C.H.; Han, S.H. Streptococcus gordonii lipoproteins induce IL-8 in human periodontal ligament cells. Mol. Immunol. 2017, 91, 218–224. [Google Scholar] [CrossRef]
- Kim, H.Y.; Baik, J.E.; Ahn, K.B.; Seo, H.S.; Yun, C.H.; Han, S.H. Streptococcus gordonii induces nitric oxide production through its lipoproteins stimulating Toll-like receptor 2 in murine macrophages. Mol. Immunol. 2017, 82, 75–83. [Google Scholar] [CrossRef]
- Siqueira, J.F., Jr.; Lopes, H.P. Mechanisms of antimicrobial activity of calcium hydroxide: A critical review. Int. Endod. J. 1999, 32, 361–369. [Google Scholar] [CrossRef]
- Safavi, K.E.; Nichols, F.C. Effect of calcium hydroxide on bacterial lipopolysaccharide. J. Endod. 1993, 19, 76–78. [Google Scholar] [CrossRef]
- Baik, J.E.; Jang, K.S.; Kang, S.S.; Yun, C.H.; Lee, K.; Kim, B.G.; Kum, K.Y.; Han, S.H. Calcium hydroxide inactivates lipoteichoic acid from Enterococcus faecalis through deacylation of the lipid moiety. J. Endod. 2011, 37, 191–196. [Google Scholar] [CrossRef]
- Baik, J.E.; Kum, K.Y.; Yun, C.H.; Lee, J.K.; Lee, K.; Kim, K.K.; Han, S.H. Calcium hydroxide inactivates lipoteichoic acid from Enterococcus faecalis. J. Endod. 2008, 34, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shen, Y.; Haapasalo, M. Antibiofilm peptides against oral biofilms. J. Oral Microbiol. 2017, 9, 1327308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, J.J.; Unholzer, A.; Schaller, M.; Schafer-Korting, M.; Korting, H.C. Human defensins. J. Mol. Med. 2005, 83, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Chang, S.W.; Perinpanayagam, H.; Lim, S.M.; Park, Y.J.; Han, S.H.; Baek, S.H.; Zhu, Q.; Bae, K.S.; Kum, K.Y. Antibacterial efficacy of a human beta-defensin-3 peptide on multispecies biofilms. J. Endod. 2013, 39, 1625–1629. [Google Scholar] [CrossRef]
- Lee, J.K.; Park, Y.J.; Kum, K.Y.; Han, S.H.; Chang, S.W.; Kaufman, B.; Jiang, J.; Zhu, Q.; Safavi, K.; Spangberg, L. Antimicrobial efficacy of a human beta-defensin-3 peptide using an Enterococcus faecalis dentine infection model. Int. Endod. J. 2013, 46, 406–412. [Google Scholar] [CrossRef]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Wu, Z.; Hoover, D.M.; Yang, D.; Boulegue, C.; Santamaria, F.; Oppenheim, J.J.; Lubkowski, J.; Lu, W. Engineering disulfide bridges to dissect antimicrobial and chemotactic activities of human beta-defensin 3. Proc. Natl. Acad. Sci. USA 2003, 100, 8880–8885. [Google Scholar] [CrossRef] [Green Version]
- Hoover, D.M.; Wu, Z.; Tucker, K.; Lu, W.; Lubkowski, J. Antimicrobial characterization of human beta-defensin 3 derivatives. Antimicrob. Agents Chemother. 2003, 47, 2804–2809. [Google Scholar] [CrossRef] [Green Version]
- Krishnakumari, V.; Rangaraj, N.; Nagaraj, R. Antifungal activities of human beta-defensins HBD-1 to HBD-3 and their C-terminal analogs Phd1 to Phd3. Antimicrob. Agents Chemother. 2009, 53, 256–260. [Google Scholar] [CrossRef] [Green Version]
- Casalinuovo, I.A.; Sorge, R.; Bonelli, G.; Di Francesco, P. Evaluation of the antifungal effect of EDTA, a metal chelator agent, on Candida albicans biofilm. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1413–1420. [Google Scholar]
- Lim, S.M.; Ahn, K.B.; Kim, C.; Kum, J.W.; Perinpanayagam, H.; Gu, Y.; Yoo, Y.J.; Chang, S.W.; Han, S.H.; Shon, W.J.; et al. Antifungal effects of synthetic human beta-defensin 3-C15 peptide. Restor. Dent. Endod. 2016, 41, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.H.; Chen, Y.J.; Huang, F.M.; Su, Y.F.; Chang, Y.C. The upregulation of matrix metalloproteinase-9 in inflamed human dental pulps. J. Endod. 2005, 31, 860–862. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T.; Potente, A.P.; Kim, J.W.; Chugal, N.; Zhang, X. Increased interleukin-8 expression in inflamed human dental pulps. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 1999, 88, 214–220. [Google Scholar] [CrossRef]
- Rechenberg, D.K.; Galicia, J.C.; Peters, O.A. Biological Markers for Pulpal Inflammation: A Systematic Review. PLoS ONE 2016, 11, e0167289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacy, P. Mechanisms of degranulation in neutrophils. Allergy Asthma Clin. Immunol. 2006, 2, 98–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omori, K.; Kida, T.; Hori, M.; Ozaki, H.; Murata, T. Multiple roles of the PGE2 -EP receptor signal in vascular permeability. Br. J. Pharmacol. 2014, 171, 4879–4889. [Google Scholar] [CrossRef] [Green Version]
- Bordier, C. Phase separation of integral membrane proteins in Triton X-114 solution. J. Biol. Chem. 1981, 256, 1604–1607. [Google Scholar]
- Chen, C.L.; Kao, C.T.; Ding, S.J.; Shie, M.Y.; Huang, T.H. Expression of the inflammatory marker cyclooxygenase-2 in dental pulp cells cultured with mineral trioxide aggregate or calcium silicate cements. J. Endod. 2010, 36, 465–468. [Google Scholar] [CrossRef]
- Park, J.; Choi, K.; Jeong, E.; Kwon, D.; Benveniste, E.N.; Choi, C. Reactive oxygen species mediate chloroquine-induced expression of chemokines by human astroglial cells. Glia 2004, 47, 9–20. [Google Scholar] [CrossRef]
- Park, S.H.; Hsiao, G.Y.; Huang, G.T. Role of substance P and calcitonin gene-related peptide in the regulation of interleukin-8 and monocyte chemotactic protein-1 expression in human dental pulp. Int. Endod. J. 2004, 37, 185–192. [Google Scholar] [CrossRef]
- Kurokawa, K.; Lee, H.; Roh, K.B.; Asanuma, M.; Kim, Y.S.; Nakayama, H.; Shiratsuchi, A.; Choi, Y.; Takeuchi, O.; Kang, H.J.; et al. The triacylated ATP binding cluster transporter substrate-binding lipoprotein of Staphylococcus aureus functions as a native ligand for toll-like receptor 2. J. Biol. Chem. 2009, 284, 8406–8411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirao, K.; Yumoto, H.; Takahashi, K.; Mukai, K.; Nakanishi, T.; Matsuo, T. Roles of TLR2, TLR4, NOD2, and NOD1 in pulp fibroblasts. J. Dent. Res. 2009, 88, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Barthel, C.R.; Levin, L.G.; Reisner, H.M.; Trope, M. TNF-alpha release in monocytes after exposure to calcium hydroxide treated Escherichia coli LPS. Int. Endod. J. 1997, 30, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Buck, R.A.; Cai, J.; Eleazer, P.D.; Staat, R.H.; Hurst, H.E. Detoxification of endotoxin by endodontic irrigants and calcium hydroxide. J. Endod. 2001, 27, 325–327. [Google Scholar] [CrossRef]
- Jiang, J.; Zuo, J.; Chen, S.H.; Holliday, L.S. Calcium hydroxide reduces lipopolysaccharide-stimulated osteoclast formation. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2003, 95, 348–354. [Google Scholar] [CrossRef]
- Jiang, J.; Yu, Y.; Kinjo, E.R.; Du, Y.; Nguyen, H.P.; Dingledine, R. Suppressing pro-inflammatory prostaglandin signaling attenuates excitotoxicity-associated neuronal inflammation and injury. Neuropharmacology 2019, 149, 149–160. [Google Scholar] [CrossRef]
- Nakanishi, T.; Shimizu, H.; Hosokawa, Y.; Matsuo, T. An immunohistological study on cyclooxygenase-2 in human dental pulp. J. Endod. 2001, 27, 385–388. [Google Scholar] [CrossRef]
- Chang, Y.C.; Yang, S.F.; Huang, F.M.; Liu, C.M.; Tai, K.W.; Hsieh, Y.S. Proinflammatory cytokines induce cyclooxygenase-2 mRNA and protein expression in human pulp cell cultures. J. Endod. 2003, 29, 201–204. [Google Scholar] [CrossRef]
- Chung, P.Y.; Khanum, R. Antimicrobial peptides as potential anti-biofilm agents against multi-drug resistant bacteria. J. Microbiol. Immunol. 2017, 50, 405–410. [Google Scholar] [CrossRef]
- Semple, F.; Webb, S.; Li, H.N.; Patel, H.B.; Perretti, M.; Jackson, I.J.; Gray, M.; Davidson, D.J.; Dorin, J.R. Human beta-defensin 3 has immunosuppressive activity in vitro and in vivo. Eur. J. Immunol. 2010, 40, 1073–1078. [Google Scholar] [CrossRef]
- Semple, F.; MacPhereson, H.; Webb, S.; Cox, S.L.; Mallin, J.L.; Tyrrell, C.; Grimes, G.R.; Semple, C.A.; Nix, M.A.; Millhouser, G.L.; et al. Human β-defensin 3 affects the activity of pro-inflammatory pathways associated with MyD88 and TRIF. Eur. J. Immunol. 2011, 41, 3291–3330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Suh, J.S.; Kim, J.M.; Kim, J.H.; Park, H.J.; Park, Y.J.; Chung, C.P. Identification of a cell-penetrating peptide domain from human beta-defensin 3 and characterization of its anti-inflammatory activity. Int. J. Nanomed. 2015, 10, 5423–5434. [Google Scholar] [CrossRef] [Green Version]
- Park, O.J.; Kim, J.; Ahn, K.B.; Lee, J.Y.; Park, Y.J.; Kum, K.Y.; Yun, C.H.; Han, S.H. A 15-amino acid C-terminal peptide of beta-defensin-3 inhibits bone resorption by inhibiting the osteoclast differentiation and disrupting podosome belt formation. J. Mol. Med. 2017, 95, 1315–1325. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Kumar, A.; Gui, J.F.; Yu, F.S. Staphylococcus aureus lipoproteins trigger human corneal epithelial innate response through toll-like receptor-2. Microb. Pathog. 2008, 44, 426–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Lee, B.H.; Lee, J.Y. Gambogic Acid Disrupts Toll-like Receptor4 Activation by Blocking Lipopolysaccharides Binding to Myeloid Differentiation Factor 2. Toxicol. Res. 2015, 31, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Human) | Primer (5′→3′) | |
|---|---|---|
| IL-8 | Forward | AGGGTTGCCAGATGCAATAC |
| Reverse | CCTTGGCCTCAATTTTGCTA | |
| MCP-1 | Forward | GCAGCAAGTGTCCCAAAGA |
| Reverse | ACAGGGTGTCTGGGGAAAG | |
| COX2 | Forward | TTCAAATGAGATTGTGGGAAAATTGCT |
| Reverse | AGATCATCTCTGCCTGAGTATCTT | |
| TLR2 | Forward | CCCATTGCTCTTTCACTGCT |
| Reverse | CTTCCTTGGAGAGGCTGATG | |
| GAPDH | Forward | GGCTGAGAACGGGAAGCTT |
| Reverse | TCCATGGTGGTGAAGACGC | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, Y.-J.; Perinpanayagam, H.; Lee, J.-Y.; Oh, S.; Gu, Y.; Kim, A.-R.; Chang, S.-W.; Baek, S.-H.; Kum, K.-Y. Synthetic Human β Defensin-3-C15 Peptide in Endodontics: Potential Therapeutic Agent in Streptococcus gordonii Lipoprotein-Stimulated Human Dental Pulp-Derived Cells. Int. J. Mol. Sci. 2020, 21, 71. https://doi.org/10.3390/ijms21010071
Yoo Y-J, Perinpanayagam H, Lee J-Y, Oh S, Gu Y, Kim A-R, Chang S-W, Baek S-H, Kum K-Y. Synthetic Human β Defensin-3-C15 Peptide in Endodontics: Potential Therapeutic Agent in Streptococcus gordonii Lipoprotein-Stimulated Human Dental Pulp-Derived Cells. International Journal of Molecular Sciences. 2020; 21(1):71. https://doi.org/10.3390/ijms21010071
Chicago/Turabian StyleYoo, Yeon-Jee, Hiran Perinpanayagam, Jue-Yeon Lee, Soram Oh, Yu Gu, A-Reum Kim, Seok-Woo Chang, Seung-Ho Baek, and Kee-Yeon Kum. 2020. "Synthetic Human β Defensin-3-C15 Peptide in Endodontics: Potential Therapeutic Agent in Streptococcus gordonii Lipoprotein-Stimulated Human Dental Pulp-Derived Cells" International Journal of Molecular Sciences 21, no. 1: 71. https://doi.org/10.3390/ijms21010071
APA StyleYoo, Y. -J., Perinpanayagam, H., Lee, J. -Y., Oh, S., Gu, Y., Kim, A. -R., Chang, S. -W., Baek, S. -H., & Kum, K. -Y. (2020). Synthetic Human β Defensin-3-C15 Peptide in Endodontics: Potential Therapeutic Agent in Streptococcus gordonii Lipoprotein-Stimulated Human Dental Pulp-Derived Cells. International Journal of Molecular Sciences, 21(1), 71. https://doi.org/10.3390/ijms21010071