Effects of Maternal Resveratrol on Maternal High-Fat Diet/Obesity with or without Postnatal High-Fat Diet


 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Maternal HFD/Obesity Affects Placenta and the Effects of Maternal Resveratrol
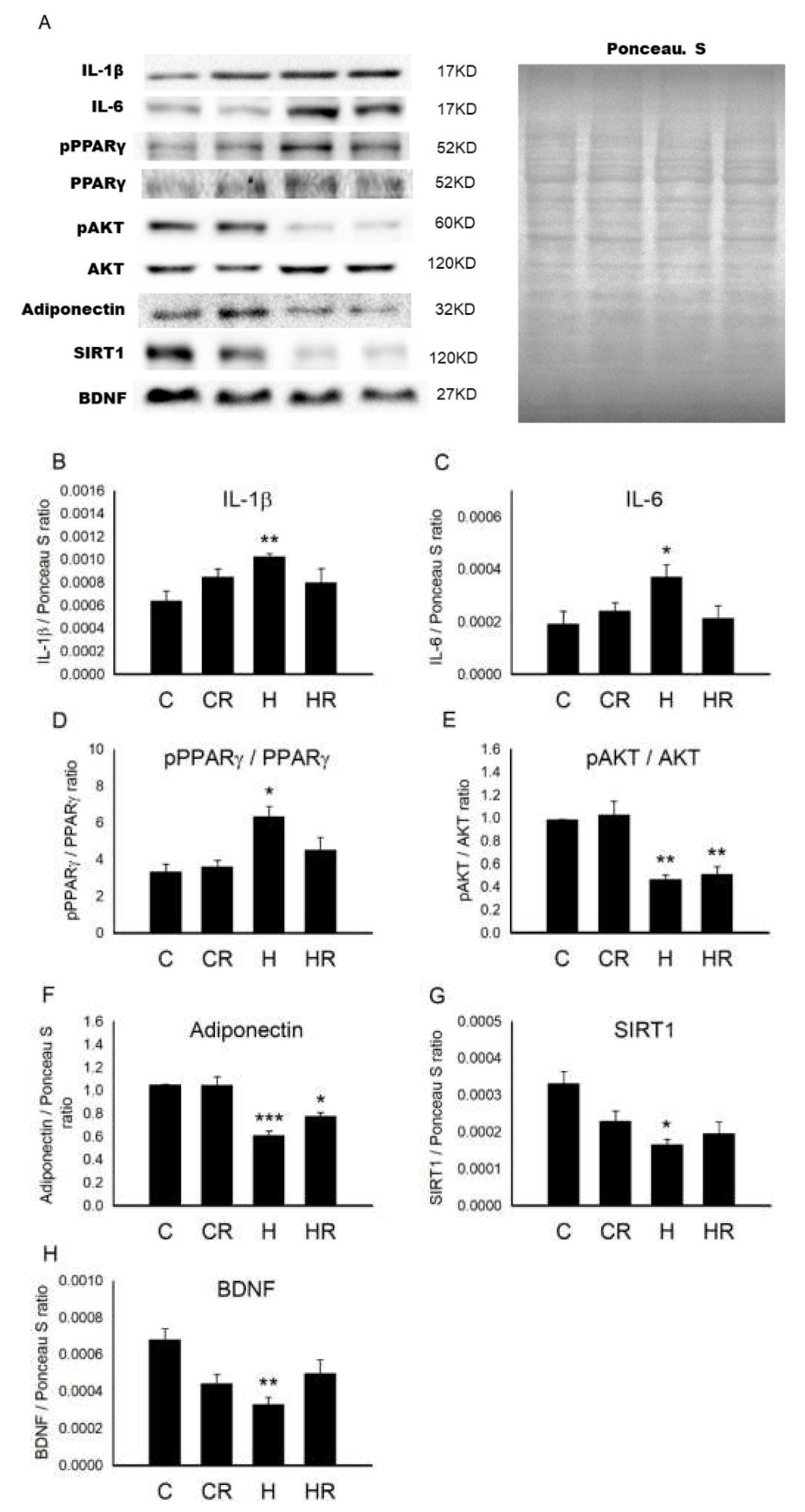
2.2. Maternal HFD/Obesity Affects Fetal Brain and the Effects of Maternal Resveratrol
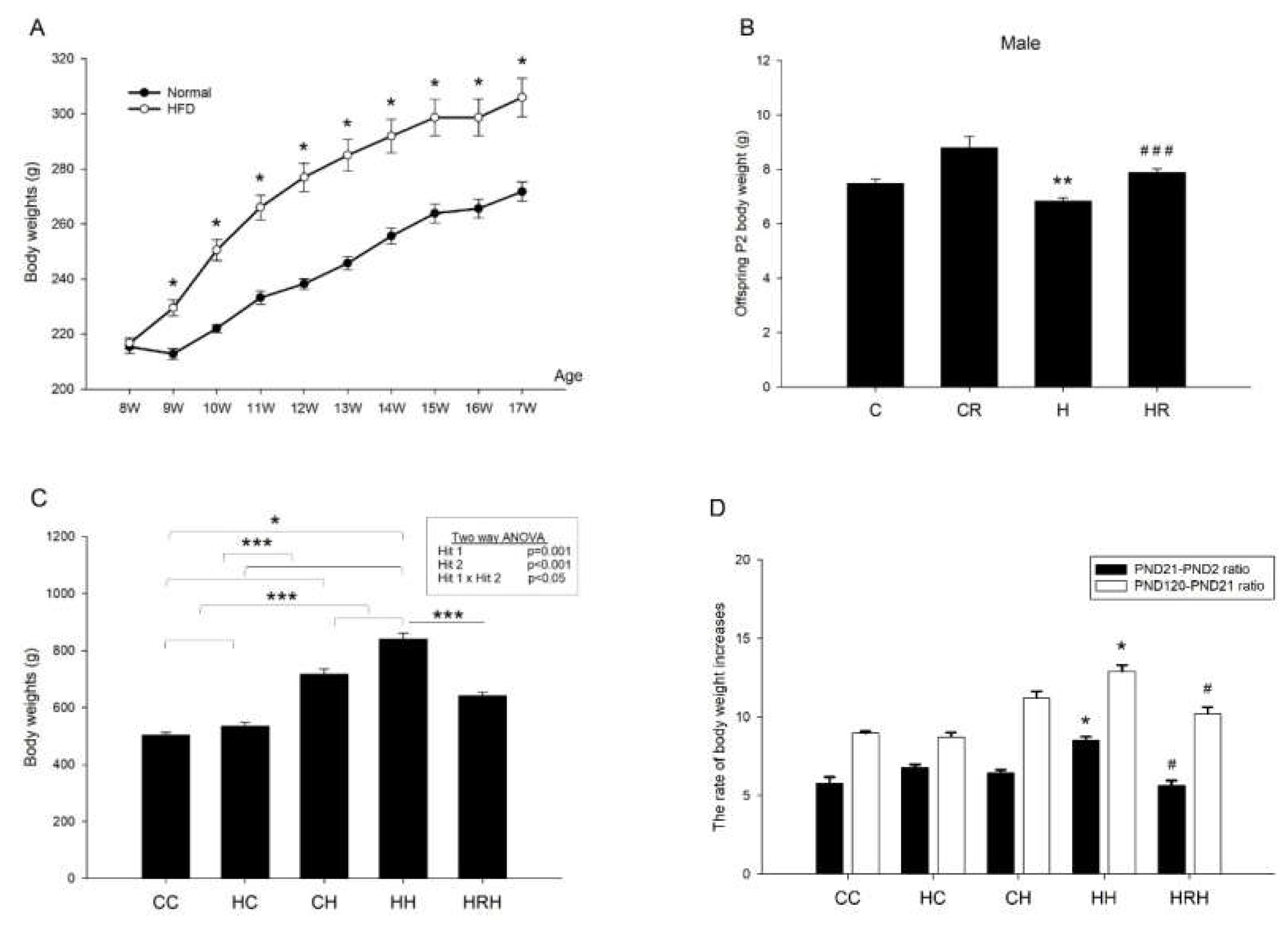
2.3. Body Weights after Birth
2.4. Blood Pressure
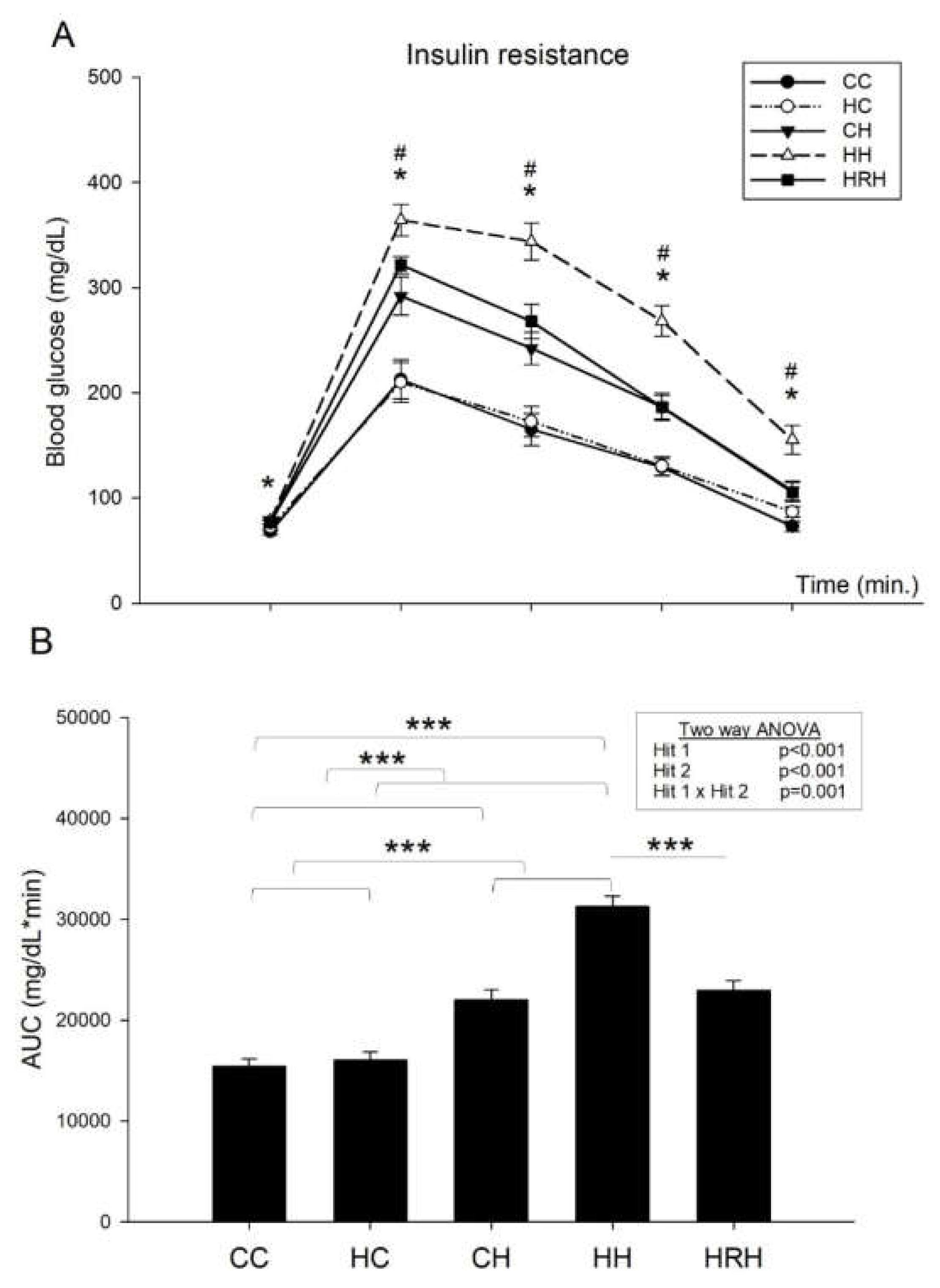
2.5. Intraperitoneal Glucose Tolerance Test (IPGTT)
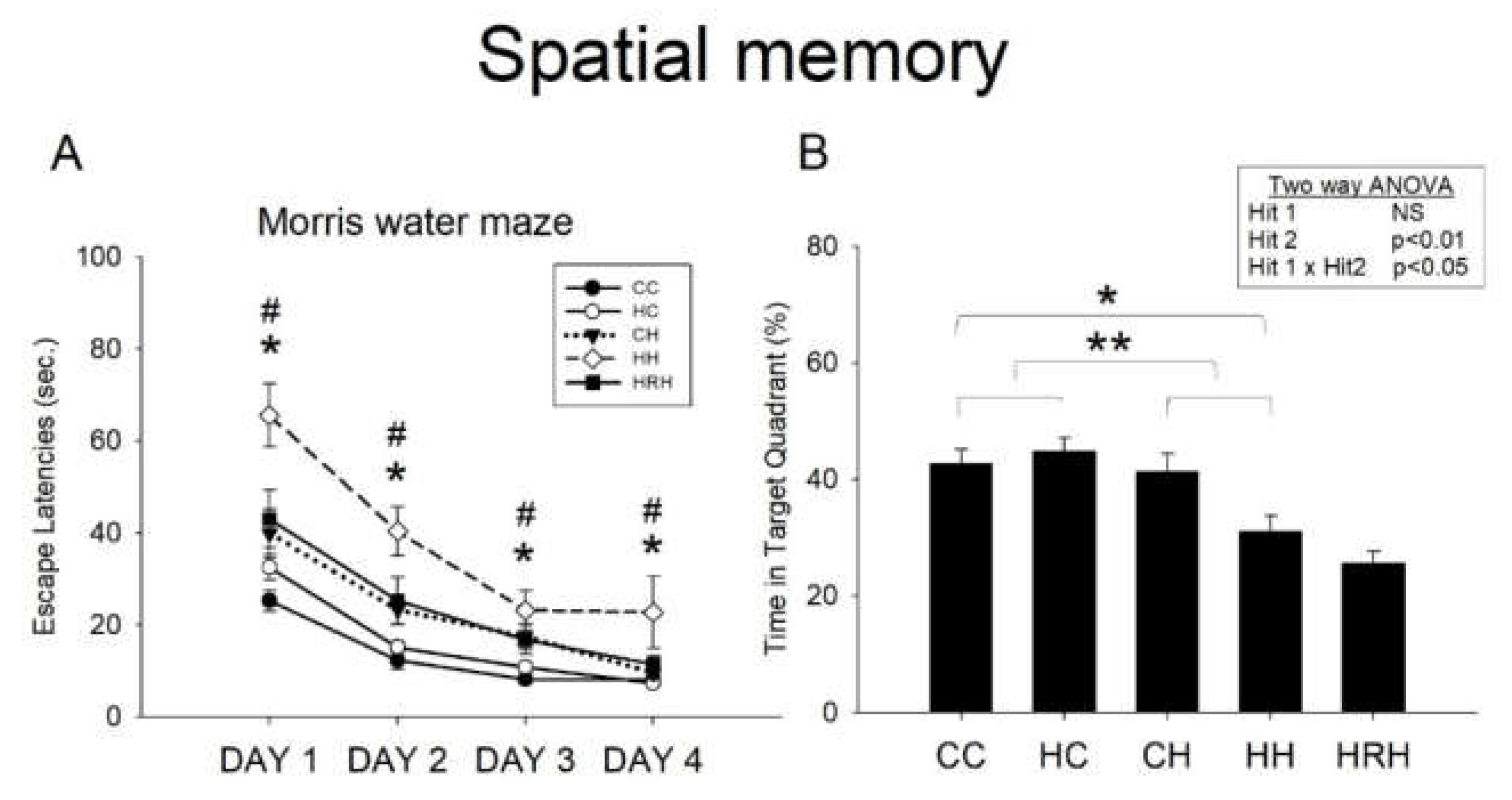
2.6. Morris Water Maze
2.7. Biochemistry Parameters
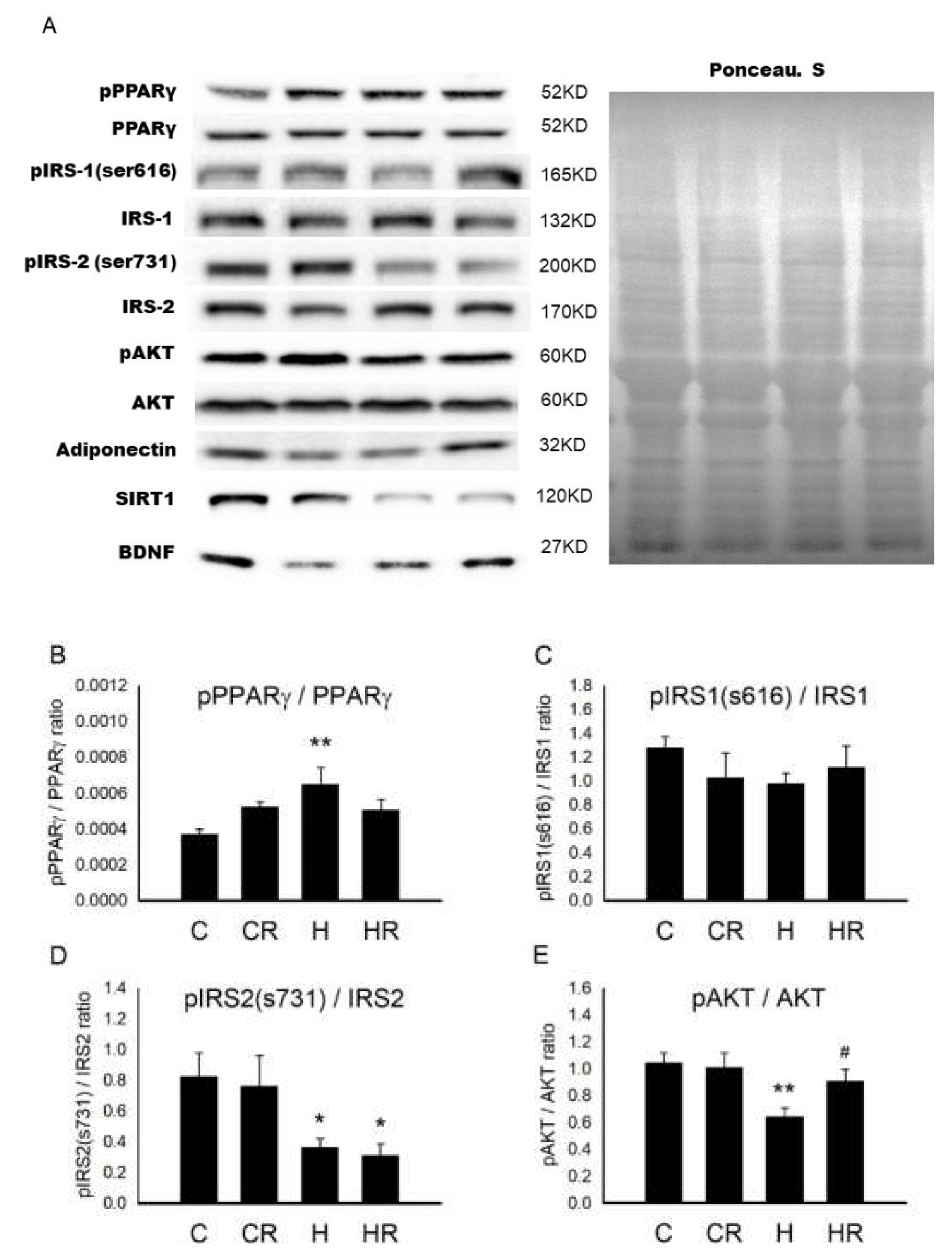
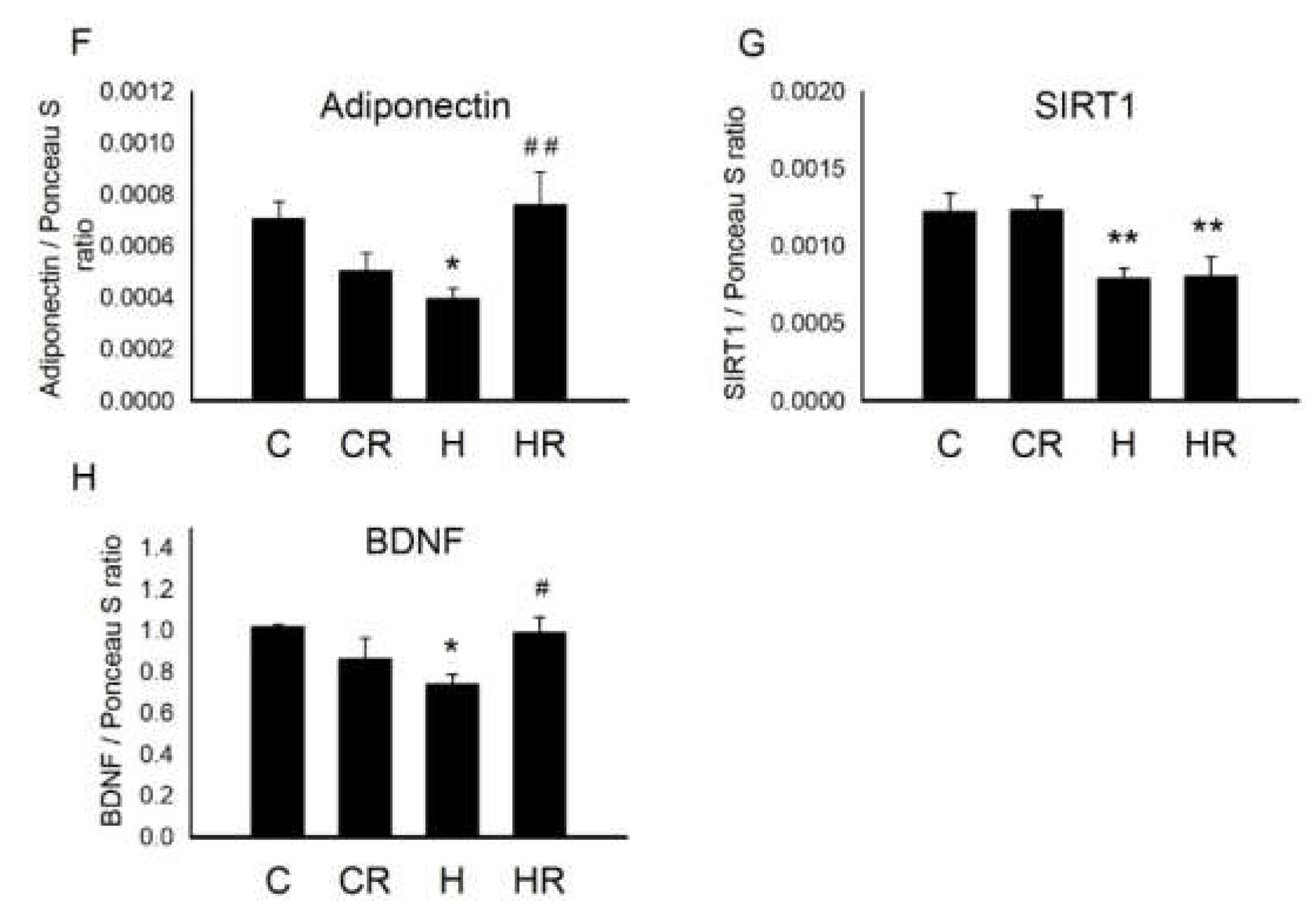
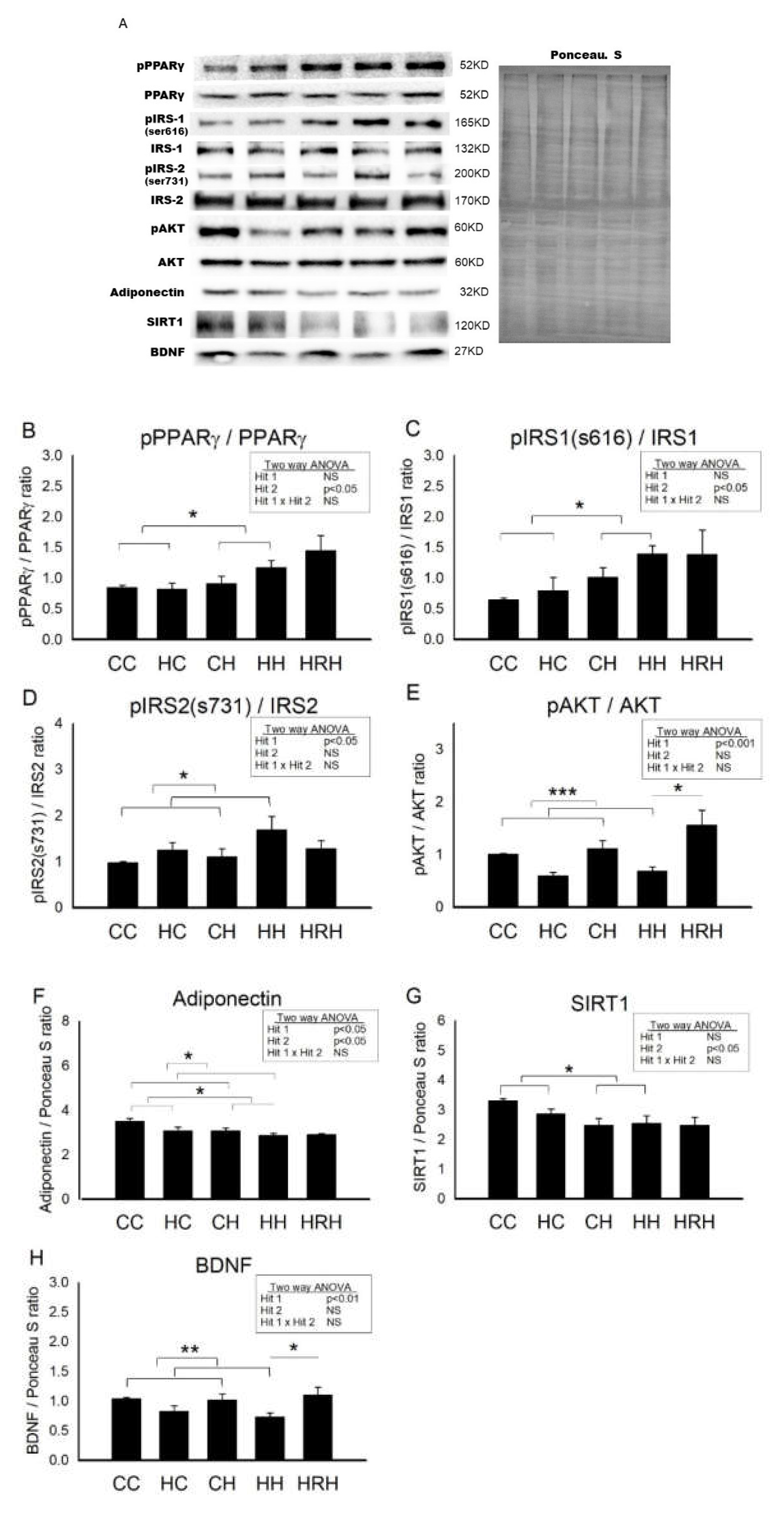
2.8. Protein Levels Involved in Cognition and Insulin Signaling in Dorsal Hippocampus in 4-Month-Old Male Offspring
3. Discussion
4. Materials and Methods
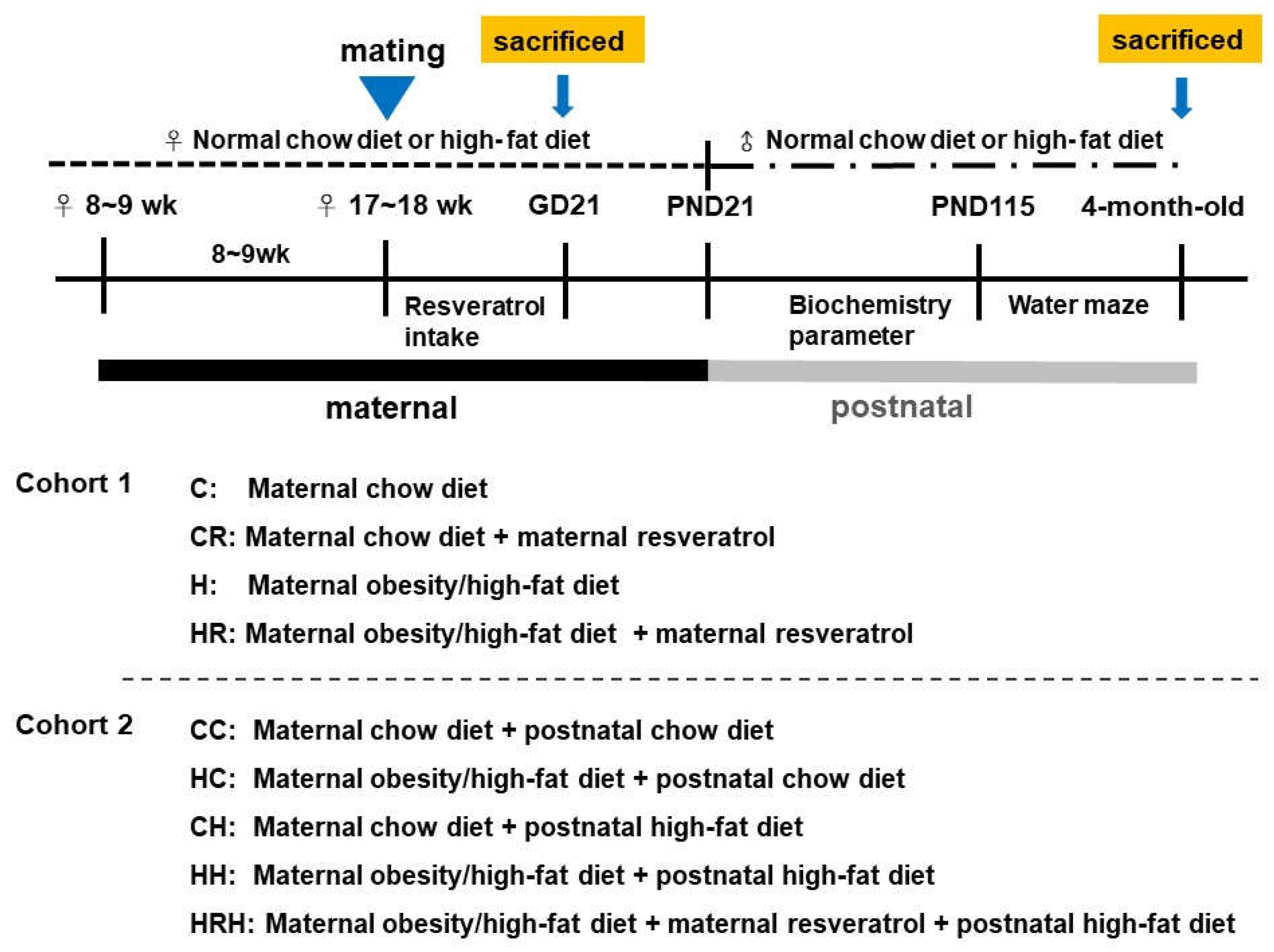
4.1. Animals and Treatment
4.2. Tissue Collection and Blood Sampling
4.3. Blood Pressure
4.4. Intraperitoneally Injected Glucose Tolerance Test (IPGTT)
4.5. Morris Water Maze: Spatial Memory
4.6. Biochemistry
4.7. Western Blotting
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Flegal, K.M. Metabolically healthy overweight and obesity. Ann. Intern. Med. 2014, 160, 515–516. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.C.; Mahmood, T. Obesity in pregnancy. Best Pract. Res. Clin. Obstet. Gynaecol. 2015, 29, 309–319. [Google Scholar] [CrossRef]
- Trasande, L.; Lee, M.; Liu, Y.; Weitzman, M.; Savitz, D. Incremental charges, costs, and length of stay associated with obesity as a secondary diagnosis among pregnant women. Med. Care 2009, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- Fusco, S.; Spinelli, M.; Cocco, S.; Ripoli, C.; Mastrodonato, A.; Natale, F.; Rinaudo, M.; Livrizzi, G.; Grassi, C. Maternal insulin resistance multigenerationally impairs synaptic plasticity and memory via gametic mechanisms. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Twinn, D.S.; Hjort, L.; Novakovic, B.; Ozanne, S.E.; Saffery, R. Intrauterine programming of obesity and type 2 diabetes. Diabetologia 2019, 62, 1789–1801. [Google Scholar] [CrossRef] [Green Version]
- Wankhade, U.D.; Thakali, K.M.; Shankar, K. Persistent influence of maternal obesity on offspring health: Mechanisms from animal models and clinical studies. Mol. Cell. Endocrinol. 2016, 435, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.; Jellyman, J.K.; Han, G.; Beall, M.; Lane, R.H.; Ross, M.G. Maternal obesity and high-fat diet program offspring metabolic syndrome. Am. J. Obstet. Gynecol. 2014, 211, 237.e1–237.e13. [Google Scholar] [CrossRef] [Green Version]
- Ito, J.; Ishii, N.; Akihara, R.; Lee, J.; Kurahashi, T.; Homma, T.; Kawasaki, R.; Fujii, J. A high-fat diet temporarily renders Sod1-deficient mice resistant to an oxidative insult. J. Nutr. Biochem. 2017, 40, 44–52. [Google Scholar] [CrossRef]
- Li, S.-W.; Yu, H.-R.; Sheen, J.-M.; Tiao, M.-M.; Tain, Y.-L.; Lin, I.-C.; Lin, Y.-J.; Chang, K.-A.; Tsai, C.-C.; Huang, L.-T. A maternal high-fat diet during pregnancy and lactation, in addition to a postnatal high-fat diet, leads to metabolic syndrome with spatial learning and memory deficits: Beneficial effects of resveratrol. Oncotarget 2017, 8, 111998. [Google Scholar] [CrossRef] [Green Version]
- Parente, L.B.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. Deleterious effects of high-fat diet on perinatal and postweaning periods in adult rat offspring. Clin. Nutr. 2008, 27, 623–634. [Google Scholar] [CrossRef]
- Zheng, S.; Feng, Q.; Cheng, J.; Zheng, J. Maternal resveratrol consumption and its programming effects on metabolic health in offspring mechanisms and potential implications. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darby, J.R.; Dollah, M.H.M.; Regnault, T.R.; Williams, M.T.; Morrison, J.L. Systematic review: Impact of resveratrol exposure during pregnancy on maternal and fetal outcomes in animal models of human pregnancy complications—Are we ready for the clinic? Pharmacol. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhu, J.; Zhang, X.; Wang, J.; Xiao, W.; Li, B.; Jin, L.; Lian, J.; Zhou, L.; Liu, J. Inhibition of cardiomyocytes hypertrophy by resveratrol is associated with amelioration of endoplasmic reticulum stress. Cell. Physiol. Biochem. 2016, 39, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Nie, P.; Hu, W.; Zhang, T.; Yang, Y.; Hou, B.; Zou, Z. Synergistic induction of erlotinib-mediated apoptosis by resveratrol in human non-Small-Cell lung cancer cells by down-regulating survivin and up-regulating PUMA. Cell. Physiol. Biochem. 2015, 35, 2255–2271. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Zhao, L.; Feng, D.; Ma, W.; Liu, Y.; Wang, Y.; Liang, J.; Yang, F.; Bi, C.; Chen, H. Resveratrol attenuated low ambient temperature-Induced myocardial hypertrophy via inhibiting cardiomyocyte apoptosis. Cell. Physiol. Biochem. 2015, 35, 2451–2462. [Google Scholar] [CrossRef]
- Diaz-Gerevini, G.T.; Repossi, G.; Dain, A.; Tarres, M.C.; Das, U.N.; Eynard, A.R. Beneficial action of resveratrol: How and why? Nutrition 2016, 32, 174–178. [Google Scholar] [CrossRef]
- Jeon, B.T.; Jeong, E.A.; Shin, H.J.; Lee, Y.; Lee, D.H.; Kim, H.J.; Kang, S.S.; Cho, G.J.; Choi, W.S.; Roh, G.S. Resveratrol attenuates obesity-Associated peripheral and central inflammation and improves memory deficit in mice fed a high-fat diet. Diabetes 2012, 61, 1444–1454. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Jin, Y.; Choi, Y.; Park, T. Resveratrol exerts anti-Obesity effects via mechanisms involving down-regulation of adipogenic and inflammatory processes in mice. Biochem. Pharmacol. 2011, 81, 1343–1351. [Google Scholar] [CrossRef]
- Hsu, M.-H.; Chen, Y.-C.; Sheen, J.-M.; Huang, L.-T. Maternal Obesity Programs Offspring Development and Resveratrol Potentially Reprograms the Effects of Maternal Obesity. Int. J. Environ. Res. Public Health 2020, 17, 1610. [Google Scholar] [CrossRef] [Green Version]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; van de Weijer, T.; Goossens, G.H.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab. 2011, 14, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.-L.; Huang, L.-T.; Hsu, C.-N. Developmental programming of adult disease: Reprogramming by melatonin? Int. J. Mol. Sci. 2017, 18, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tain, Y.-L.; Hsu, C.-N. Developmental programming of the metabolic syndrome: Can we reprogram with resveratrol? Int. J. Mol. Sci. 2018, 19, 2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lendvai, Á.; Deutsch, M.J.; Plösch, T.; Ensenauer, R. The peroxisome proliferator-activated receptors under epigenetic control in placental metabolism and fetal development. Am. J. Physiol.-Endocrinol. Metab. 2016, 310, E797–E810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, Y.; Qi, G.; Fan, R.; Qiao, Q.; Sun, Y.; Gao, Y.; Liu, X.J.T.F.J. EGCG ameliorates high-Fat–And high-Fructose–Induced cognitive defects by regulating the IRS/AKT and ERK/CREB/BDNF signaling pathways in the CNS. FASEB J. 2017, 31, 4998–5011. [Google Scholar] [CrossRef] [Green Version]
- Aye, I.L.; Rosario, F.J.; Powell, T.L.; Jansson, T. Reply to Carbillon: Fetal/placental weight ratio and placental function. Proc Natl Acad Sci USA 2016, 113, E261. [Google Scholar] [CrossRef] [Green Version]
- Qiao, L.; Guo, Z.; Bosco, C.; Guidotti, S.; Wang, Y.; Wang, M.; Parast, M.; Schaack, J.; Hay, W.W.; Moore, T.R.J.D. Maternal high-fat feeding increases placental lipoprotein lipase activity by reducing SIRT1 expression in mice. Diabetes. 2015, 64, 3111–3120. [Google Scholar] [CrossRef] [Green Version]
- Dhobale, M. Neurotrophins: Role in adverse pregnancy outcome. Int. J. Dev. Neurosci. 2014, 37, 8–14. [Google Scholar] [CrossRef]
- Copps, K.; White, M. Regulation of insulin sensitivity by serine/threonine phosphorylation of insulin receptor substrate proteins IRS1 and IRS2. Diabetologia 2012, 55, 2565–2582. [Google Scholar] [CrossRef] [Green Version]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Kiliaan, A.J.; Arnoldussen, I.A.; Gustafson, D.R. Adipokines: A link between obesity and dementia? Lancet Neurol. 2014, 13, 913–923. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, A.L.; Diniz, B.S.; Campos, A.C.; Miranda, A.S.; Rocha, N.P.; Talib, L.L.; Gattaz, W.F.; Forlenza, O.V. Decreased levels of circulating adiponectin in mild cognitive impairment and Alzheimer’s disease. Neuromolecular Med. 2013, 15, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Villalobos-Labra, R.; Silva, L.; Subiabre, M.; Araos, J.; Salsoso, R.; Fuenzalida, B.; Sáez, T.; Toledo, F.; González, M.; Quezada, C. Akt/mTOR role in human foetoplacental vascular insulin resistance in diseases of pregnancy. J. Diabetes Res. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.M.; Seo, H.W.; Kwon, M.S.; Han, A.R.; Lee, S.K. SIRT1 negatively regulates invasive and angiogenic activities of the extravillous trophoblast. Am. J. Reprod. Immunol. 2019. [Google Scholar] [CrossRef]
- Briana, D.D.; Malamitsi-Puchner, A. The role of adipocytokines in fetal growth. Ann. N.Y. Acad. Sci. 2010, 1205, 82–87. [Google Scholar] [CrossRef]
- Briana, D.D.; Malamitsi-Puchner, A. Developmental origins of adult health and disease: The metabolic role of BDNF from early life to adulthood. Metabolism 2018, 81, 45–51. [Google Scholar] [CrossRef]
- Guzzardi, M.A.; Sanguinetti, E.; Bartoli, A.; Kemeny, A.; Panetta, D.; Salvadori, P.A.; Burchielli, S.; Iozzo, P. Elevated glycemia and brain glucose utilization predict BDNF lowering since early life. J. Cereb. Blood Flow Metab. 2018, 38, 447–455. [Google Scholar] [CrossRef]
- Lewis, R.M.; Desoye, G. Placental lipid and fatty acid transfer in maternal overnutrition. Ann. Nutr. Metab. 2017, 70, 228–231. [Google Scholar] [CrossRef]
- Malti, N.; Merzouk, H.; Merzouk, S.; Loukidi, B.; Karaouzene, N.; Malti, A.; Narce, M. Oxidative stress and maternal obesity: Feto-Placental unit interaction. Placenta 2014, 35, 411–416. [Google Scholar] [CrossRef]
- Nogues, P.; Dos Santos, E.; Jammes, H.; Berveiller, P.; Arnould, L.; Vialard, F.; Dieudonné, M.-N. Maternal obesity influences expression and DNA methylation of the adiponectin and leptin systems in human third-Trimester placenta. Clin. Epigenetics 2019, 11, 20. [Google Scholar] [CrossRef] [Green Version]
- Rosario, F.J.; Powell, T.L.; Jansson, T. Activation of placental insulin and mTOR signaling in a mouse model of maternal obesity associated with fetal overgrowth. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2015, 310, R87–R93. [Google Scholar] [CrossRef] [Green Version]
- Prince, C.S.; Maloyan, A.; Myatt, L. Maternal obesity alters brain derived neurotrophic factor (BDNF) signaling in the placenta in a sexually dimorphic manner. Placenta 2017, 49, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edlow, A.; Hui, L.; Wick, H.; Fried, I.; Bianchi, D. Assessing the fetal effects of maternal obesity via transcriptomic analysis of cord blood: A prospective case–Control study. BJOG Int. J. Obstet. Gynaecol. 2016, 123, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stachowiak, E.K.; Srinivasan, M.; Stachowiak, M.K.; Patel, M.S. Maternal obesity induced by a high fat diet causes altered cellular development in fetal brains suggestive of a predisposition of offspring to neurological disorders in later life. Metab. Brain Dis. 2013, 28, 721–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niculescu, M.D.; Lupu, D.S. High fat diet-Induced maternal obesity alters fetal hippocampal development. Int. J. Dev. Neurosci. 2009, 27, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Bourque, S.; Dolinsky, V.; Dyck, J.; Davidge, S. Maternal resveratrol treatment during pregnancy improves adverse fetal outcomes in a rat model of severe hypoxia. Placenta 2012, 33, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.; Hafner, H.; Varghese, M.; Griffin, C.; Clemente, J.; Islam, M.; Carlson, Z.; Zhu, A.; Hak, L.; Abrishami, S. Programming effects of maternal and gestational obesity on offspring metabolism and metabolic inflammation. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Roberts, V.H.; Pound, L.D.; Thorn, S.R.; Gillingham, M.B.; Thornburg, K.L.; Friedman, J.E.; Frias, A.E.; Grove, K.L. Beneficial and cautionary outcomes of resveratrol supplementation in pregnant nonhuman primates. FASEB J. 2014, 28, 2466–2477. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.; Jain, S.; Bhattacharjee, J. Correlation of adiponectin and leptin with insulin resistance: A pilot study in healthy north Indian population. Indian J. Clin. Biochem. 2011, 26, 193–196. [Google Scholar] [CrossRef] [Green Version]
- Pancani, T.; Anderson, K.L.; Brewer, L.D.; Kadish, I.; DeMoll, C.; Landfield, P.W.; Blalock, E.M.; Porter, N.M.; Thibault, O. Effect of high-Fat diet on metabolic indices, cognition, and neuronal physiology in aging F344 rats. Neurobiol. Aging 2013, 34, 1977–1987. [Google Scholar] [CrossRef] [Green Version]
- Heyward, F.D.; Gilliam, D.; Coleman, M.A.; Gavin, C.F.; Wang, J.; Kaas, G.; Trieu, R.; Lewis, J.; Moulden, J.; Sweatt, J.D. Obesity weighs down memory through a mechanism involving the neuroepigenetic dysregulation of Sirt1. J. Neurosci. 2016, 36, 1324–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Liu, J.; Wang, J.; Liu, Q. The controversial links among calorie restriction, SIRT1, and resveratrol. Free Radic. Biol. Med. 2011, 51, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Boyuk, B.; Degirmencioglu, S.; Atalay, H.; Guzel, S.; Acar, A.; Celebi, A.; Ekizoglu, I.; Simsek, C. Relationship between levels of brain-Derived neurotrophic factor and metabolic parameters in patients with type 2 diabetes mellitus. J. Diabetes Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and synaptic plasticity, cognitive function, and dysfunction. In Neurotrophic Factors; Springer: Berlin/Heidelberg, Germany, 2014; pp. 223–250. [Google Scholar]
- Molteni, R.; Barnard, R.; Ying, Z.; Roberts, C.; Gomez-Pinilla, F. A high-Fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience 2002, 112, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Stranahan, A.M.; Norman, E.D.; Lee, K.; Cutler, R.G.; Telljohann, R.S.; Egan, J.M.; Mattson, M.P. Diet-Induced insulin resistance impairs hippocampal synaptic plasticity and cognition in middle-Aged rats. Hippocampus 2008, 18, 1085–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tozuka, Y.; Kumon, M.; Wada, E.; Onodera, M.; Mochizuki, H.; Wada, K. Maternal obesity impairs hippocampal BDNF production and spatial learning performance in young mouse offspring. Neurochem. Int. 2010, 57, 235–247. [Google Scholar] [CrossRef]
- Hurley, L.L.; Akinfiresoye, L.; Kalejaiye, O.; Tizabi, Y. Antidepressant effects of resveratrol in an animal model of depression. Behav. Brain Res. 2014, 268, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Cholerton, B.; Baker, L.D.; Craft, S. Insulin, cognition, and dementia. Eur. J. Pharmacol. 2013, 719, 170–179. [Google Scholar] [CrossRef] [Green Version]
- Boocock, D.J.; Faust, G.E.; Patel, K.R.; Schinas, A.M.; Brown, V.A.; Ducharme, M.P.; Booth, T.D.; Crowell, J.A.; Perloff, M.; Gescher, A.J. Phase I dose escalation pharmacokinetic study in healthy volunteers of resveratrol, a potential cancer chemopreventive agent. Cancer Epidemiol. Prev. Biomark. 2007, 16, 1246–1252. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.D.; Burdock, G.A.; Edwards, J.A.; Beck, M.; Bausch, J. Safety studies conducted on high-Purity trans-resveratrol in experimental animals. Food Chem. Toxicol. 2009, 47, 2170–2182. [Google Scholar] [CrossRef]
- Cordner, Z.A.; Khambadkone, S.G.; Boersma, G.J.; Song, L.; Summers, T.N.; Moran, T.H.; Tamashiro, K.L. Maternal high-Fat diet results in cognitive impairment and hippocampal gene expression changes in rat offspring. Exp. Neurol. 2019, 318, 92–100. [Google Scholar] [CrossRef]
- Chen, H.-E.; Lin, Y.-J.; Lin, I.-C.; Yu, H.-R.; Sheen, J.-M.; Tsai, C.-C.; Huang, L.-T.; Tain, Y.-L. Resveratrol prevents combined prenatal NG-nitro-L-arginine-methyl ester (L-NAME) treatment plus postnatal high-Fat diet induced programmed hypertension in adult rat offspring: Interplay between nutrient-Sensing signals, oxidative stress and gut microbiota. J. Nutr. Biochem. 2019, 70, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.-L.; Lin, Y.-J.; Sheen, J.-M.; Yu, H.-R.; Tiao, M.-M.; Chen, C.-C.; Tsai, C.-C.; Huang, L.-T.; Hsu, C.-N. High fat diets sex-Specifically affect the renal transcriptome and program obesity, kidney injury, and hypertension in the offspring. Nutrients 2017, 9, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tain, Y.-L.; Sheen, J.-M.; Yu, H.-R.; Chen, C.-C.; Tiao, M.-M.; Hsu, C.-N.; Lin, Y.-J.; Kuo, K.-C.; Huang, L.-T. Maternal melatonin therapy rescues prenatal dexamethasone and postnatal high-Fat diet induced programmed hypertension in male rat offspring. Front. Physiol. 2015, 6, 377. [Google Scholar] [CrossRef] [Green Version]
- Sheen, J.-M.; Hsieh, C.-S.; Tain, Y.-L.; Li, S.-W.; Yu, H.-R.; Chen, C.-C.; Tiao, M.-M.; Chen, Y.-C.; Huang, L.-T. Programming effects of prenatal glucocorticoid exposure with a postnatal high-fat diet in diabetes mellitus. Int. J. Mol. Sci. 2016, 17, 533. [Google Scholar] [CrossRef] [Green Version]
- Sheen, J.-M.; Chen, Y.-C.; Hsu, M.-H.; Tain, Y.-L.; Yu, H.-R.; Huang, L.-T. Combined intraperitoneal and intrathecal etanercept reduce increased brain tumor necrosis factor-alpha and asymmetric dimethylarginine levels and rescues spatial deficits in young rats after bile duct ligation. Front. Cell. Neurosci. 2016, 10, 167. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-W.; Chen, Y.-C.; Sheen, J.-M.; Hsu, M.-H.; Tain, Y.-L.; Chang, K.-A.; Huang, L.-T. Minocycline restores cognitive-relative altered proteins in young bile duct-ligated rat prefrontal cortex. Life Sci. 2017, 180, 75–82. [Google Scholar] [CrossRef]
- Parkhurst, C.N.; Yang, G.; Ninan, I.; Savas, J.N.; Yates III, J.R.; Lafaille, J.J.; Hempstead, B.L.; Littman, D.R.; Gan, W.-B. Microglia promote learning-dependent synapse formation through brain-derived neurotrophic factor. Cell 2013, 155, 1596–1609. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Shen, Y. An old method facing a new challenge: Re-Visiting housekeeping proteins as internal reference control for neuroscience research. Life Sci. 2013, 92, 747–751. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Hsieh, C.S.; Chen, C.C.; Sheen, J.M.; Lee, C.T.; Huang, L.T. Melatonin prevents increased asymmetric dimethylarginine in young rats with bile duct ligation. J. Pineal Res. 2010, 48, 212–221. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | CC | HC | CH | HH | HRH | Two-Way ANOVA (p Value) | ||
|---|---|---|---|---|---|---|---|---|
| Rat numbers | n = 14 | n = 13 | n = 16 | n = 14 | n = 14 | Hit 1 | Hit 2 | Hit 1 × Hit 2 |
| AST (U/L) | 99.43 ± 10.24 | 98.23 ± 7.6 | 226.88 ± 45.58 | 431.93 ± 80.82 | 168.50 ± 60.91### | <0.05 | <0.0001 | 0.036 |
| ALT (U/L) | 24.29 ± 1.55 | 29.77 ± 1.03 | 153.75 ± 48.57 | 259.43 ± 52.55 | 80.29 ± 32.35 ### | NS | <0.0001 | NS |
| Cholesterol (mg/dL) | 65.00 ± 3.66 | 73.00 ± 4.07 | 68.25 ± 5.78 | 92.14 ± 5.80 | 54.14 ± 2.95 ### | <0.05 | <0.05 | NS |
| Triglyceride (mg/dL) | 77.00 ± 9.87 | 79.63 ± 8.23 | 82.88 ± 10.81 | 74.63 ± 4.77 | 78.13 ± 7.48 | NS | <0.05 | NS |
| Target | Company | Catalog | Dilution Factor |
|---|---|---|---|
| IL-6 (interleukin 6) | Abcam | ab9324 | 1:1000 |
| IL-1β (interleukin 1 beta) | Abcam | ab9722 | 1:1000 |
| Adiponectin | Abcam | ab62551 | 1:1000 |
| BDNF | Santa cruz | Sc-546 | 1:1000 |
| pPPARγ (phosphoryl peroxisome proliferator-activated receptors γ) | Bioss | bs-3737R | 1:1000 |
| PPARγ (peroxisome proliferator-activated receptors γ) | Santa cruz | 7196 | 1:1000 |
| pAKT (phosphoryl alpha serine/threonine-protein kinase) | Cell signaling | #9271 | 1:1000 |
| AKT (alpha serine/threonine-protein kinase) | Cell signaling | #9272 | 1:1000 |
| pIRS1 (phosphoryl insulin receptor substrate 1) | Millipore | #07-843 | 1:1000 |
| IRS1 (insulin receptor substrate 1) | Cell signaling | #2382 | 1:1000 |
| pIRS2 (phosphoryl insulin receptor substrate 2) | abcam | ab3690 | 1:1000 |
| IRS2 (insulin receptor substrate 2) | Abclonal | A7945 | 1:1000 |
| SIRT1 (sirtuin 1) | Abcam | ab104833 | 1:1000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, M.-H.; Sheen, J.-M.; Lin, I.-C.; Yu, H.-R.; Tiao, M.-M.; Tain, Y.-L.; Huang, L.-T. Effects of Maternal Resveratrol on Maternal High-Fat Diet/Obesity with or without Postnatal High-Fat Diet. Int. J. Mol. Sci. 2020, 21, 3428. https://doi.org/10.3390/ijms21103428
Hsu M-H, Sheen J-M, Lin I-C, Yu H-R, Tiao M-M, Tain Y-L, Huang L-T. Effects of Maternal Resveratrol on Maternal High-Fat Diet/Obesity with or without Postnatal High-Fat Diet. International Journal of Molecular Sciences. 2020; 21(10):3428. https://doi.org/10.3390/ijms21103428
Chicago/Turabian StyleHsu, Mei-Hsin, Jiunn-Ming Sheen, I-Chun Lin, Hong-Ren Yu, Mao-Meng Tiao, You-Lin Tain, and Li-Tung Huang. 2020. "Effects of Maternal Resveratrol on Maternal High-Fat Diet/Obesity with or without Postnatal High-Fat Diet" International Journal of Molecular Sciences 21, no. 10: 3428. https://doi.org/10.3390/ijms21103428
APA StyleHsu, M. -H., Sheen, J. -M., Lin, I. -C., Yu, H. -R., Tiao, M. -M., Tain, Y. -L., & Huang, L. -T. (2020). Effects of Maternal Resveratrol on Maternal High-Fat Diet/Obesity with or without Postnatal High-Fat Diet. International Journal of Molecular Sciences, 21(10), 3428. https://doi.org/10.3390/ijms21103428