Super-Resolution Microscopy Reveals Diversity of Plant Centromere Architecture

 , , , , ,
, , , , , 

Abstract
:1. Introduction
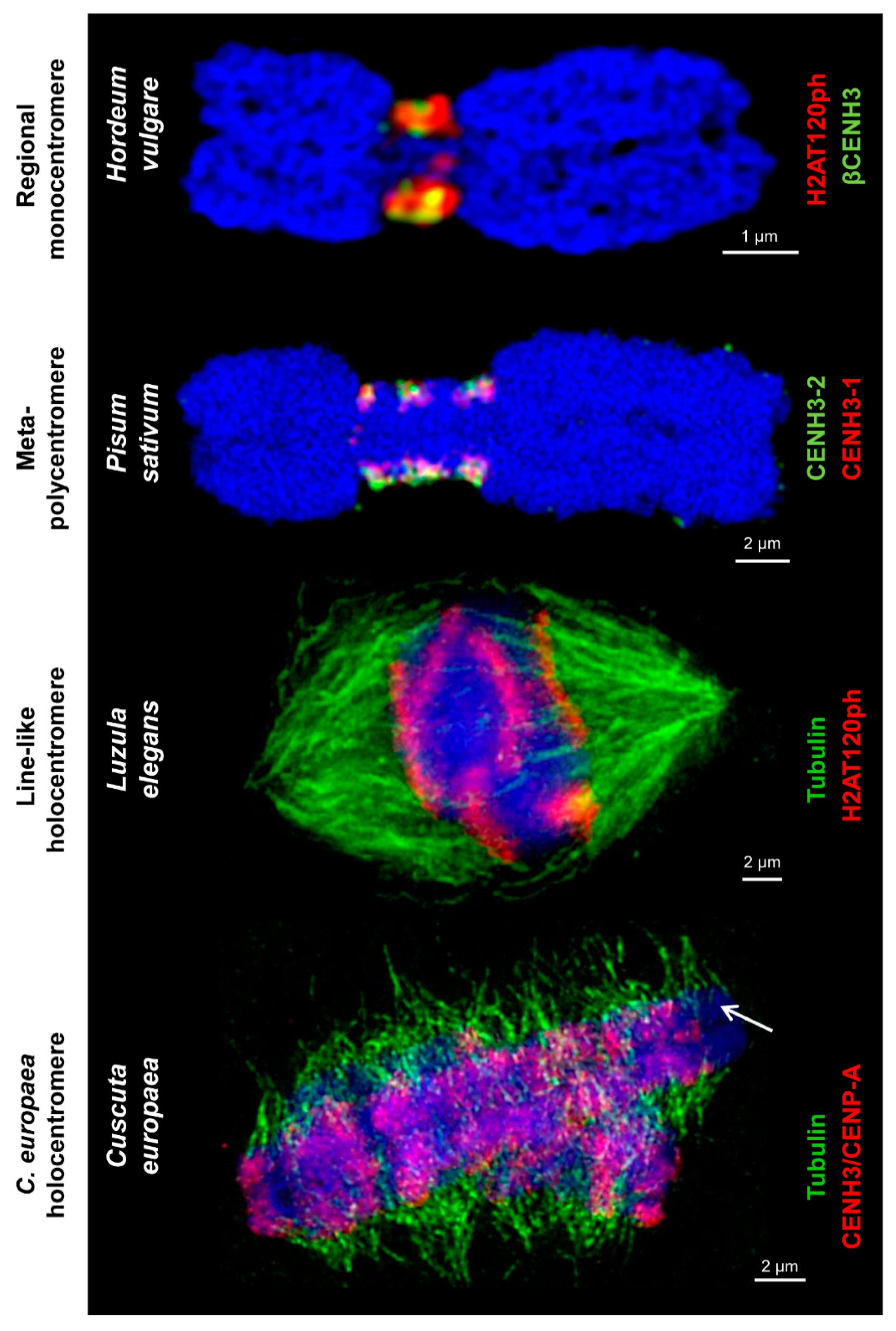
2. Centromere Diversity in Plants
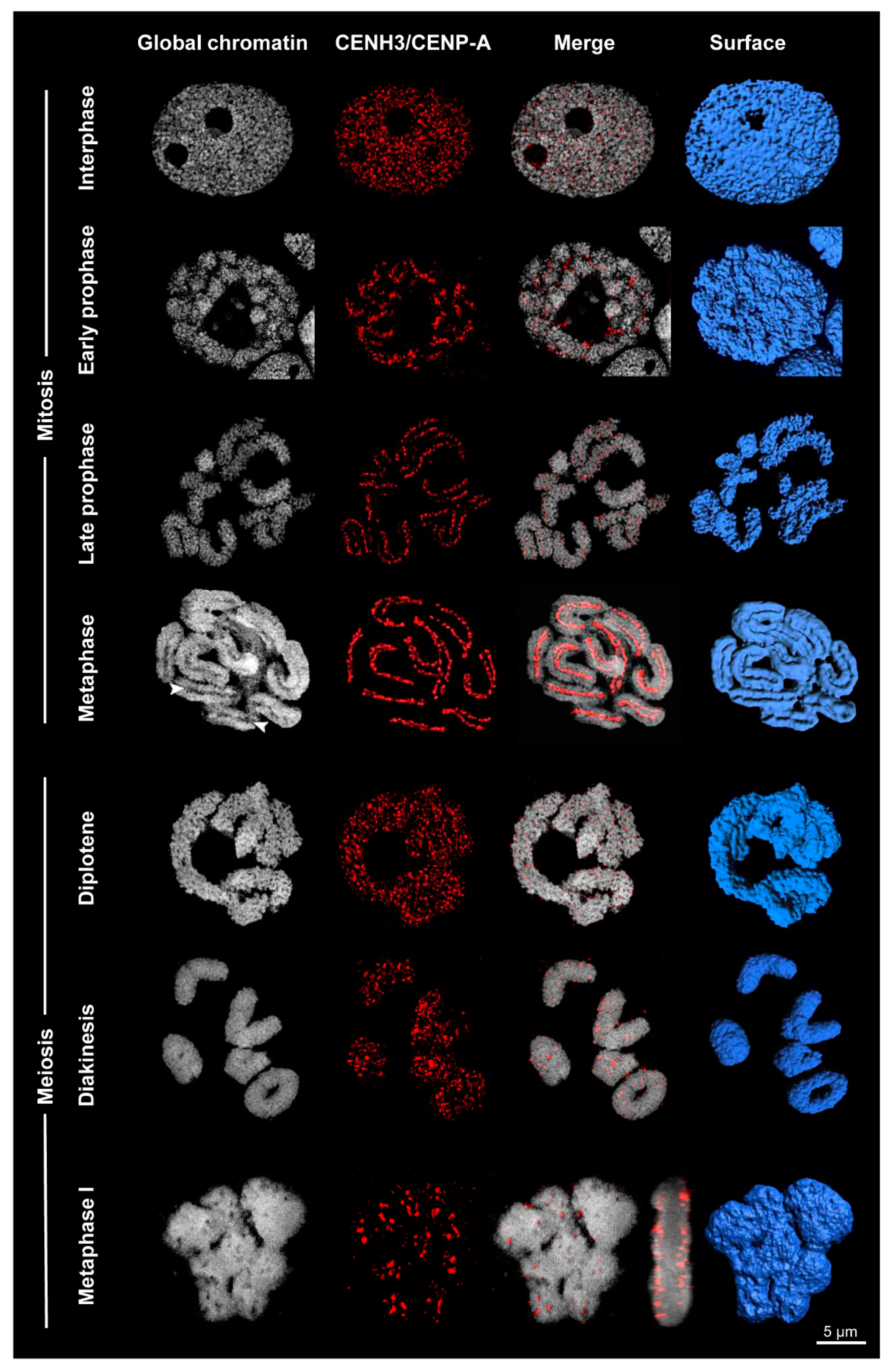
3. Variation at Different Stages of the Cell Cycle
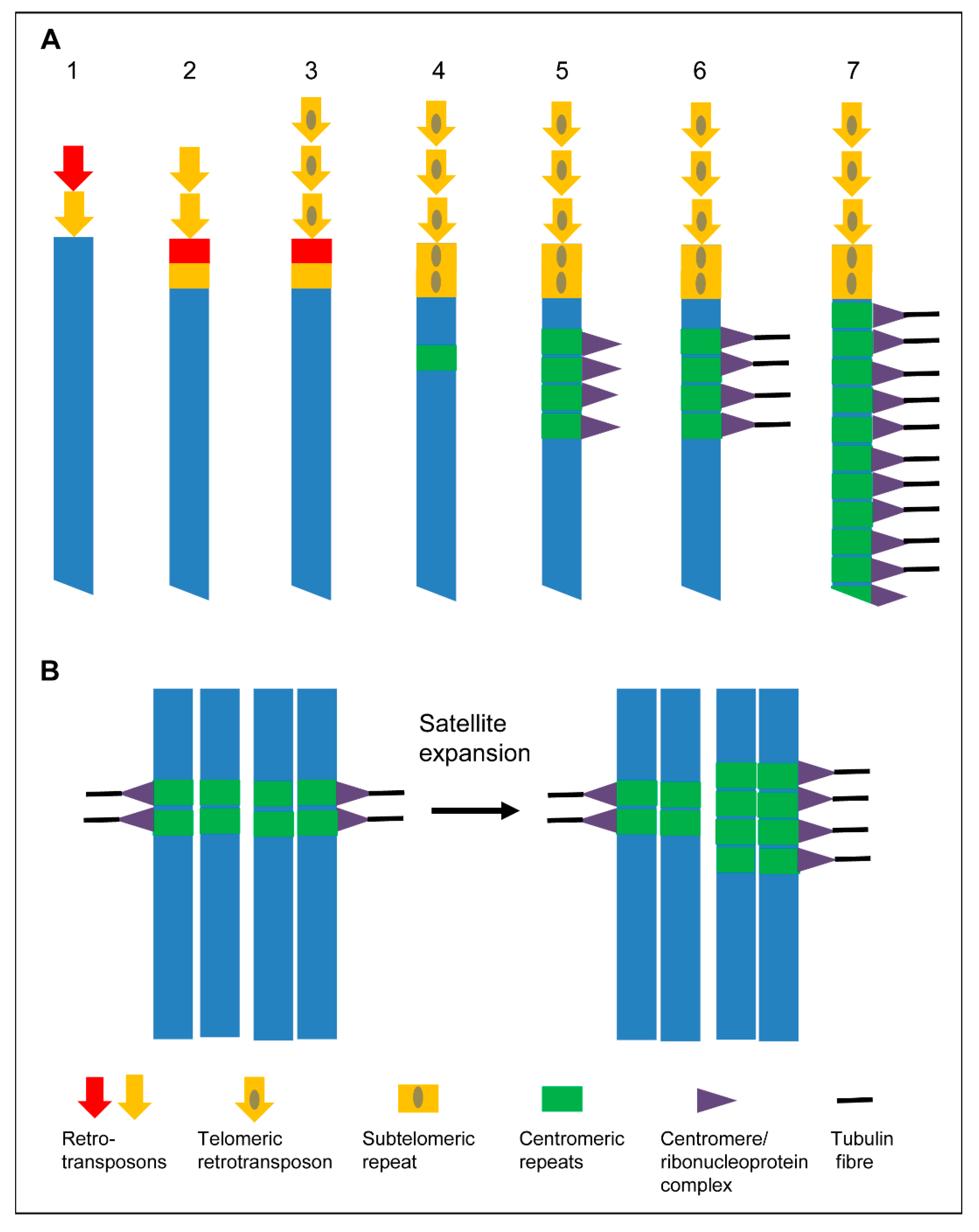
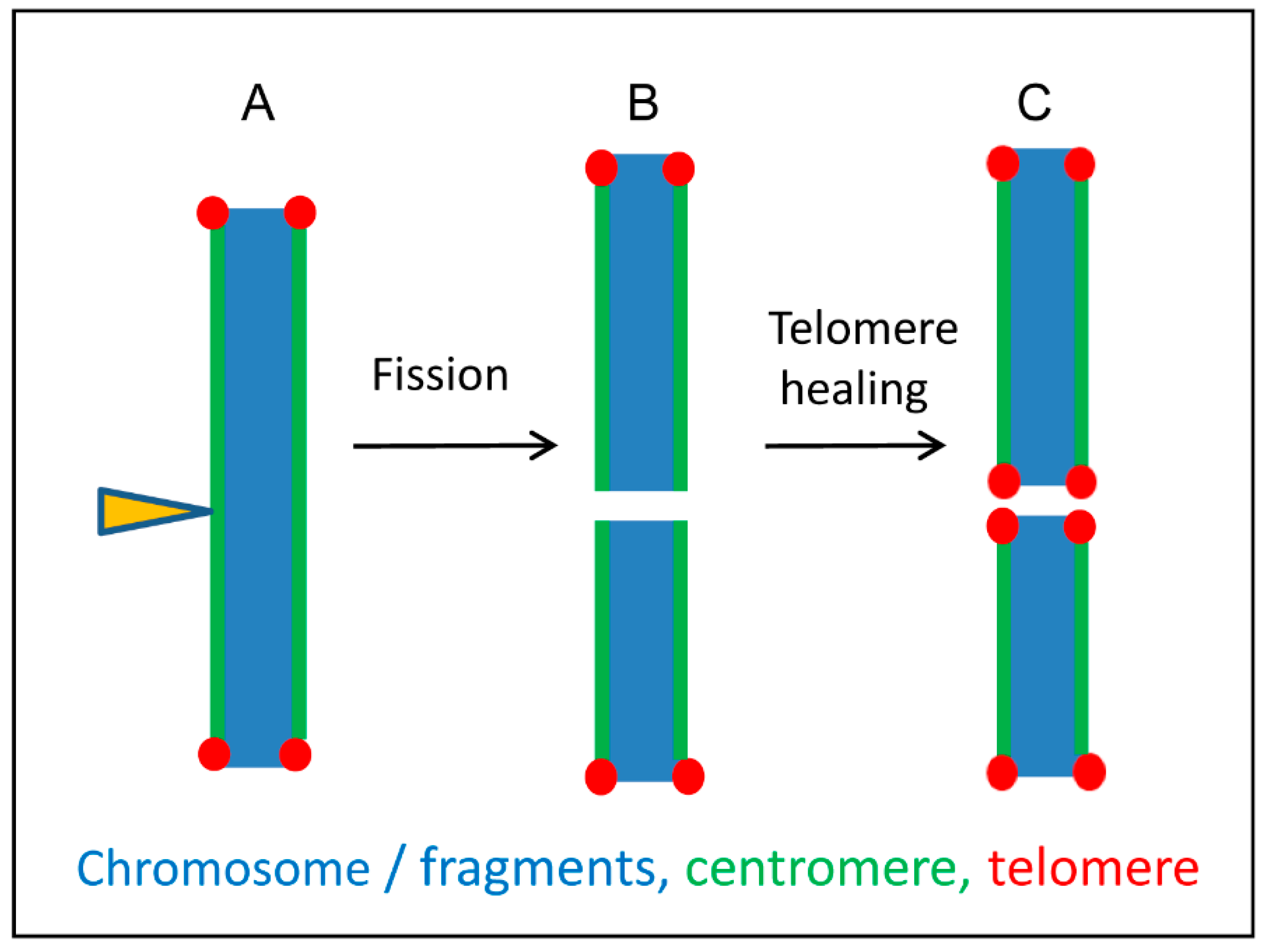
4. Centromere Evolution
4.1. Is Holocentricity Original or Derived?
4.2. Is Holocentricity Related to CENH3/CENP-A Loss?
5. Advantages and Challenges of Holocentricity
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Musacchio, A.; Desai, A. A molecular view of kinetochore assembly and function. Biology (Basel) 2017, 6, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henikoff, S.; Ahmad, K.; Malik, H.S. The centromere paradox: Stable inheritance with rapidly evolving DNA. Science 2001, 293, 1098–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleveland, D.W.; Mao, Y.; Sullivan, K.F. Centromeres and kinetochores: From epigenetics to mitotic checkpoint signaling. Cell 2003, 112, 407–421. [Google Scholar] [CrossRef] [Green Version]
- Plohl, M.; Meštrović, N.; Mravinac, B. Centromere identity from the DNA point of view. Chromosoma 2014, 123, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Hoang, P.T.N.; Schubert, V.; Meister, A.; Fuchs, J.; Schubert, I. Variation in genome size, cell and nucleus volume, chromosome number and rDNA loci among duckweeds. Sci. Rep. 2019, 9, 3234. [Google Scholar] [CrossRef]
- Tran, T.D.; Cao, H.X.; Jovtchev, G.; Novák, P.; Vu, G.T.; Macas, J.; Schubert, I.; Fuchs, J. Chromatin organization and cytological features of carnivorous Genlisea species with large genome size differences. Front. Plant Sci. 2015, 6, 613. [Google Scholar] [CrossRef] [Green Version]
- Vu, G.T.H.; Schmutzer, T.; Bull, F.; Cao, H.X.; Fuchs, J.; Tran, D.T.; Jovtchev, G.; Pistrick, K.; Stein, N.; Pecinka, A.; et al. Comparative genome analysis reveals divergent genome size evolution in a carnivorous plant genus. Plant Genome 2015, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Fonsêca, A.; Ferreira, J.; dos Santos, T.R.; Mosiolek, M.; Bellucci, E.; Kami, J.; Gepts, P.; Geffroy, V.; Schweizer, D.; dos Santos, K.G.; et al. Cytogenetic map of common bean (Phaseolus vulgaris L.). Chromosome Res. 2010, 18, 487–502. [Google Scholar] [CrossRef] [Green Version]
- Da Costa Silva, S.; Marques, A.; dos Santos Soares Filho, W.; Mirkov, T.E.; Andrea Pedrosa-Harand, A.; Guerra, M. The cytogenetic map of the Poncirus trifoliata (L.) Raf.—a nomenclature system for chromosomes of all citric species. Trop. Plant Biol. 2011, 4, 99–105. [Google Scholar] [CrossRef]
- Heckmann, S.; Houben, A. Holokinetic centromeres. In Plant Centromere Biology; Jiang, J., Birchler, J.A., Eds.; Wiley-Blackwell: Ames, IA, USA, 2013; Volume 1, pp. 83–94. [Google Scholar]
- Neumann, P.; Navrátilová, A.; Schroeder-Reiter, E.; Koblížková, A.; Steinbauerová, V.; Chocholová, E.; Novák, P.; Wanner, G.; Macas, J. Stretching the rules: Monocentric chromosomes with multiple centromere domains. PLoS Genet. 2012, 8, e1002777. [Google Scholar] [CrossRef] [Green Version]
- Neumann, P.; Pavlíková, Z.; Koblížková, A.; Fuková, I.; Jedličková, V.; Novák, P.; Macas, J. Centromeres off the hook: Massive changes in centromere size and structure following duplication of CenH3 gene in Fabeae species. Mol. Biol. Evol. 2015, 32, 1862–1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, P.; Schubert, V.; Fuková, I.; Manning, J.E.; Houben, A.; Macas, J. Epigenetic histone marks of extended meta-polycentric centromeres of Lathyrus and Pisum chromosomes. Front. Plant Sci. 2016, 7, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.C.; Lee, C.C.; Kao, C.Y.; Chang, N.C.; Lin, C.C.; Shoemaker, D.; Wang, J. Evolution of long centromeres in fire ants. BMC Evol. Biol. 2016, 16, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkley, B.R.; Valdivia, M.M.; Tousson, A.; Brenner, S.L. Compound kinetochores of the Indian muntjac. Evolution by linear fusion of unit kinetochores. Chromosoma 1984, 91, 1–11. [Google Scholar] [CrossRef]
- Metcalfe, C.J.; Bulazel, K.V.; Ferreri, G.C.; Schroeder-Reiter, E.; Wanner, G.; Rens, W.; Obergfell, C.; Eldridge, M.D.; O’Neill, R.J. Genomic instability within centromeres of interspecific marsupial hybrids. Genetics 2007, 177, 2507–2517. [Google Scholar] [CrossRef] [Green Version]
- Paweletz, N.; Vig, B.K.; Finze, E.M. Evolution of compound centromeres. A new phenomenon. Cancer Genet. Cytogenet. 1989, 42, 75–86. [Google Scholar] [CrossRef]
- Clarke, L.; Carbon, J. Isolation of a yeast centromere and construction of functional small circular chromosomes. Nature 1980, 287, 504–509. [Google Scholar] [CrossRef]
- Earnshaw, W.C.; Allshire, R.C.; Black, B.E.; Bloom, K.; Brinkley, B.R.; Brown, W.; Cheeseman, I.M.; Choo, K.H.; Copenhaver, G.P.; Deluca, J.G.; et al. Esperanto for histones: CENP-A, not CenH3, is the centromeric histone H3 variant. Chromosome Res. 2013, 21, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Drinnenberg, I.A.; deYoung, D.; Henikoff, S.; Malik, H.S. Recurrent loss of CenH3 is associated with independent transitions to holocentricity in insects. eLife 2014, 3, e03676. [Google Scholar] [CrossRef]
- Akiyoshi, B.; Gull, K. Discovery of unconventional kinetochores in kinetoplastids. Cell 2014, 156, 1247–1258. [Google Scholar] [CrossRef] [Green Version]
- Akiyoshi, B. Evolution: A mosaic-type centromere in an early-diverging fungus. Curr. Biol. 2019, 29, R1184–R1186. [Google Scholar] [CrossRef]
- Navarro-Mendoza, M.I.; Pérez-Arques, C.; Panchal, S.; Nicolás, F.E.; Mondo, S.J.; Ganguly, P.; Pangilinan, J.; Grigoriev, I.V.; Heitman, J.; Sanyal, K.; et al. Early diverging fungus Mucor circinelloides lacks centromeric histone CENP-A and displays a mosaic of point and regional centromeres. Curr. Biol. 2019, 29, 3791–3802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hooff, J.J.; Tromer, E.; van Wijk, L.M.; Snel, B.; Kops, G.J. Evolutionary dynamics of the kinetochore network in eukaryotes as revealed by comparative genomics. EMBO Rep. 2017, 18, 1559–1571. [Google Scholar] [CrossRef] [PubMed]
- Schermelleh, L.; Heintzmann, R.; Leonhardt, H. A guide to super-resolution fluorescence microscopy. J. Cell Biol. 2010, 190, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, U.; Reilly, D.T.; Schroeder, C.M. Zooming in on biological processes with fluorescence nanoscopy. Curr. Opin. Biotechnol. 2013, 24, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.R.; Ross, S.T.; Davidson, M.W. Structured illumination microscopy for superresolution. ChemPhysChem 2014, 15, 566–576. [Google Scholar] [CrossRef]
- Komis, G.; Šamajová, O.; Ovečka, M.; Šamaj, J. Super-resolution microscopy in plant cell imaging. Trends Plant Sci. 2015, 20, 834–843. [Google Scholar] [CrossRef]
- Nienhaus, K.; Nienhaus, G.U. Where do we stand with super-resolution optical microscopy? J. Mol. Biol. 2016, 428, 308–322. [Google Scholar] [CrossRef]
- Baroux, C.; Schubert, V. Technical Review: Microscopy and image processing tools to analyse plant chromatin—Practical considerations. In Plant Chromatin Dynamics: Methods and Protocols; Bemer, M., Baroux, C., Eds.; Humana Press: New York, NY, USA, 2018; Volume 1675, pp. 537–589. [Google Scholar]
- Schermelleh, L.; Ferrand, A.; Huser, T.; Eggeling, C.; Sauer, M.; Biehlmaier, O.; Drummen, G.P.C. Super-resolution microscopy demystified. Nat. Cell Biol. 2019, 21, 72–84. [Google Scholar] [CrossRef]
- Rouquette, J.; Cremer, C.; Cremer, T.; Fakan, S. Functional nuclear architecture studied by microscopy: Present and future. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2010; Volume 282, pp. 1–90. [Google Scholar]
- Han, R.; Li, Z.; Fan, Y.; Jiang, Y. Recent advances in super-resolution fluorescence imaging and its applications in biology. J. Genet. Genomics 2013, 40, 583–595. [Google Scholar] [CrossRef]
- Fornasiero, E.F.; Opazo, F. Super-resolution imaging for cell biologists: Concepts, applications, current challenges and developments. Bioessays 2015, 37, 436–451. [Google Scholar] [CrossRef] [PubMed]
- Kraus, F.; Miron, E.; Demmerle, J.; Chitiashvili, T.; Budco, A.; Alle, Q.; Matsuda, A.; Leonhardt, H.; Schermelleh, L.; Markaki, Y. Quantitative 3D structured illumination microscopy of nuclear structures. Nat. Protoc. 2017, 12, 1011–1028. [Google Scholar] [CrossRef] [PubMed]
- Schubert, V. Super-resolution microscopy – applications in plant cell research. Front. Plant Sci. 2017, 8, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demidov, D.; Schubert, V.; Kumke, K.; Weiss, O.; Karimi-Ashtiyani, R.; Buttlar, J.; Heckmann, S.; Wanner, G.; Dong, Q.; Han, F.; et al. Anti-phosphorylated histone H2AThr120: A universal microscopic marker for centromeric chromatin of mono- and holocentric plant species. Cytogenet. Genome Res. 2014, 143, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Houben, A.; Wako, T.; Furushima-Shimogawara, R.; Presting, G.; Künzel, G.; Schubert, I.; Fukui, K. The cell cycle dependent phosphorylation of histone H3 is correlated with the condensation of plant mitotic chromosomes. Plant J. 1999, 18, 675–679. [Google Scholar] [CrossRef]
- Gernand, D.; Demidov, D.; Houben, A. The temporal and spatial pattern of histone H3 phosphorylation at serine 28 and serine 10 is similar in plants but differs between mono- and polycentric chromosomes. Cytogenet. Genome Res. 2003, 101, 172–176. [Google Scholar] [CrossRef]
- Weisshart, K.; Fuchs, J.; Schubert, V. Structured Illumination Microscopy (SIM) and Photoactivated Localization Microscopy (PALM) to analyze the abundance and distribution of RNA polymerase II molecules on flow-sorted Arabidopsis nuclei. Bio-Protocol 2016, 6, e1725. [Google Scholar] [CrossRef] [Green Version]
- Wanner, G.; Schroeder-Reiter, E.; Ma, W.; Houben, A.; Schubert, V. The ultrastructure of mono- and holocentric plant centromeres: An immunological investigation by structured illumination microscopy and scanning electron microscopy. Chromosoma 2015, 124, 503–517. [Google Scholar] [CrossRef]
- Furuyama, S.; Biggins, S. Centromere identity is specified by a single centromeric nucleosome in budding yeast. Proc. Natl. Acad. Sci. USA 2007, 104, 14706–14711. [Google Scholar] [CrossRef] [Green Version]
- Pluta, A.F.; Mackay, A.M.; Ainsztein, A.M.; Goldberg, I.G.; Earnshaw, W.C. The centromere: Hub of chromosomal activities. Science 1995, 270, 1591–1594. [Google Scholar] [CrossRef]
- Ishii, T.; Karimi-Ashtiyani, R.; Banaei-Moghaddam, A.M.; Schubert, V.; Fuchs, J.; Houben, A. The differential loading of two barley CENH3 variants into distinct centromeric substructures is cell type- and development-specific. Chromosome Res. 2015, 23, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Banaei-Moghaddam, A.M.; Schubert, V.; Kumke, K.; Weibeta, O.; Klemme, S.; Nagaki, K.; Macas, J.; González-Sánchez, M.; Heredia, V.; Gómez-Revilla, D.; et al. Nondisjunction in favor of a chromosome: The mechanism of rye B chromosome drive during pollen mitosis. Plant Cell 2012, 24, 4124–4134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, L.; Neumann, P.; Jang, T.-S.; Klemme, S.; Schubert, V.; Koblížková, A.; Houben, A.; Macas, J. Mitotic spindle attachment to the holocentric chromosomes of Cuscuta europaea does not correlate with the distribution of CENH3 chromatin. Front. Plant Sci. 2020, 10, 1799. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Schubert, V.; Houben, A.; Pedrosa-Harand, A. Restructuring of holocentric centromeres during meiosis in the plant Rhynchospora pubera. Genetics 2016, 204, 555–568. [Google Scholar] [CrossRef] [Green Version]
- Heckmann, S.; Schroeder-Reiter, E.; Kumke, K.; Ma, L.; Nagaki, K.; Murata, M.; Wanner, G.; Houben, A. Holocentric chromosomes of Luzula elegans are characterized by a longitudinal centromere groove, chromosome bending, and a terminal nucleolus organizer region. Cytogenet. Genome Res. 2011, 134, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Heckmann, S.; Jankowska, M.; Schubert, V.; Kumke, K.; Ma, W.; Houben, A. Alternative meiotic chromatid segregation in the holocentric plant Luzula elegans. Nat. Commun. 2014, 5, 4979. [Google Scholar] [CrossRef] [Green Version]
- Heckmann, S.; Schubert, V.; Houben, A. Holocentric plant meiosis: First sisters, then homologues. Cell Cycle 2014, 13, 3623–3624. [Google Scholar] [CrossRef] [Green Version]
- Marques, A.; Ribeiro, T.; Neumann, P.; Macas, J.; Novák, P.; Schubert, V.; Pellino, M.; Fuchs, J.; Ma, W.; Kuhlmann, M.; et al. Holocentromeres in Rhynchospora are associated with genome-wide centromere-specific repeat arrays interspersed among euchromatin. Proc. Natl. Acad. Sci. USA 2015, 112, 13633–13638. [Google Scholar] [CrossRef] [Green Version]
- Lawrimore, J.; Bloom, K. The regulation of chromosome segregation via centromere loops. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 352–370. [Google Scholar] [CrossRef]
- Nagaki, K.; Kashihara, K.; Murata, M. Visualization of diffuse centromeres with centromere-specific histone H3 in the holocentric plant Luzula nivea. Plant Cell 2005, 17, 1886–1893. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, T.; Buddenhagen, C.E.; Thomas, W.W.; Souza, G.; Pedrosa-Harand, A. Are holocentrics doomed to change? Limited chromosome number variation in Rhynchospora Vahl (Cyperaceae). Protoplasma 2018, 255, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Cabral, G.; Marques, A.; Schubert, V.; Pedrosa-Harand, A.; Schlögelhofer, P. Chiasmatic and achiasmatic inverted meiosis of plants with holocentric chromosomes. Nat. Commun. 2014, 5, 5070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, M.; Cabral, G.; Cuacos, M.; González-García, M.; González-Sánchez, M.; Vega, J.; Puertas, M.J. Neocentrics and holokinetics (holocentrics): Chromosomes out of the centromeric rules. Cytogenet. Genome Res. 2010, 129, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Cuacos, M.; Franklin, F.C.H.; Heckmann, S. Atypical centromeres in plants-what they can tell us. Front. Plant Sci. 2015, 6, 913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, D.M.; Marques, A.; Andrade, C.G.; Guyot, R.; Chaluvadi, S.R.; Pedrosa-Harand, A.; Houben, A.; Bennetzen, J.L.; Vanzela, A.L. Developmental programmed cell death during asymmetric microsporogenesis in holocentric species of Rhynchospora (Cyperaceae). J. Exp. Bot. 2016, 67, 5391–5401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Schubert, V.; Martis, M.M.; Hause, G.; Liu, Z.; Shen, Y.; Conrad, U.; Shi, W.; Scholz, U.; Taudien, S.; et al. The distribution of α-kleisin during meiosis in the holocentromeric plant Luzula elegans. Chromosome Res. 2016, 24, 393–405. [Google Scholar] [CrossRef]
- Melters, D.P.; Paliulis, L.V.; Korf, I.F.; Chan, S.W. Holocentric chromosomes: Convergent evolution, meiotic adaptations, and genomic analysis. Chromosome Res. 2012, 20, 579–593. [Google Scholar] [CrossRef] [Green Version]
- Dernburg, A.F. Here, there, and everywhere: Kinetochore function on holocentric chromosomes. J. Cell Biol. 2001, 153, F33–F38. [Google Scholar] [CrossRef] [Green Version]
- Guerra, M.; Ribeiro, T.; Felix, L.P. Monocentric chromosomes in Juncus (Juncaceae) and implications for the chromosome evolution of the family. Bot. J. Linn. Soc. 2019, 191, 475–483. [Google Scholar] [CrossRef]
- Pimpinelli, S.; Goday, C. Unusual kinetochores and chromatin diminution in Parascaris. Trends Genet. 1989, 5, 310–315. [Google Scholar] [CrossRef]
- Wrensch, D.I.; Kethley, J.B.; Noton, R.A. Cytogenetics of holokinetic chromosomes and and inverted meiosis: Keys to evolutionary succes of mites, with generalization on eukaryotes. In Mites: Ecological and Evolutionary Analyses of Life History Patterns; Houck, M.A., Ed.; Springer: Boston, MA, USA, 1994; pp. 282–343. [Google Scholar]
- Mola, L.M.; Papeschi, A.G. Holokinetic chromosomes at a glance. J. Basic Appl. Genet. 2006, 17, 17–33. [Google Scholar]
- Lima de Faria, A. Genetics, origin and evolution of kinetochores. Hereditas 1949, 35, 422–444. [Google Scholar] [CrossRef]
- Câmara, A. Posição actual do problemado centromero. Genética Ibérica 1953, 5, 1–33. [Google Scholar]
- Castro, D. The structure of the centromere and its functioning. Genética Ibérica 1950, 2, 201–209. [Google Scholar]
- Moore, G.; Aragón-Alcaide, L.; Roberts, M.; Reader, S.; Miller, T.; Foote, T. Are rice chromosomes components of a holocentric chromosome ancestor? Plant Mol. Biol. 1997, 35, 17–23. [Google Scholar] [CrossRef]
- Schrader, F. The role of the kinetochore in the chromosomal evolution of the Heteroptera and Homoptera. Evolution 1947, 1, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Sybenga, J. Specialization in the behavior of chromosomes on the meiotic spindle. Genetica 1981, 57, 143–151. [Google Scholar] [CrossRef]
- Sybenga, J. Cytogenetics in Plant Breeding, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1992; p. 469. [Google Scholar]
- Vaarama, A. Cytological observation on Pleurozium schreberi, with special reference to centromere evolution. Ann. Bot. Soc. Zool. Bot. Fenn. Vanamo 1954, 28, 1–59. [Google Scholar]
- Swanson, C.P. Cytology and Cytogenetics, 1st ed.; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1957; p. 596. [Google Scholar]
- Greilhuber, J. Chromosomes of the monocotyledons (general aspects). In Monocotyledons: Systematics and Evolution; Rudall, P.J., Cribb, P.J., Cutler, D.F., Humphries, C.J., Eds.; Royal Botanic Gardens: Kew, UK, 1995; Volume 2, pp. 379–414. [Google Scholar]
- Villasante, A.; Abad, J.P.; Méndez-Lago, M. Centromeres were derived from telomeres during the evolution of the eukaryotic chromosome. Proc. Natl. Acad. Sci. USA 2007, 104, 10542–10547. [Google Scholar] [CrossRef] [Green Version]
- Villasante, A.; Méndez-Lago, M.; Abad, J.P.; Montejo de Garcíni, E. The birth of the centromere. Cell Cycle 2007, 6, 2872–2876. [Google Scholar] [CrossRef] [Green Version]
- Malik, H.S.; Henikoff, S. Conflict begets complexity: The evolution of centromeres. Curr. Opin. Genet. Dev. 2002, 12, 711–718. [Google Scholar] [CrossRef]
- Zedek, F.; Bureš, P. Holocentric chromosomes: From tolerance to fragmentation to colonization of the land. Ann. Bot. 2018, 121, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Král, J.; Forman, M.; Korinkova, T.; Lerma, A.C.R.; Haddad, C.R.; Musilova, J.; Rezac, M.; Herrera, I.M.A.; Thakur, S.; Dippenaar-Schoeman, A.S.; et al. Insights into the karyotype and genome evolution of haplogyne spiders indicate a polyploid origin of lineage with holokinetic chromosomes. Sci. Rep. 2019, 9, 3001. [Google Scholar] [CrossRef] [PubMed]
- Stimpson, K.M.; Matheny, J.E.; Sullivan, B.A. Dicentric chromosomes: Unique models to study centromere function and inactivation. Chromosome Res. 2012, 20, 595–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drinnenberg, I.A.; Akiyoshi, B. Evolutionary lessons from species with unique kinetochores. Prog. Mol. Subcell. Biol. 2017, 56, 111–138. [Google Scholar] [PubMed]
- Zedek, F.; Bureš, P. Pest arthropods with holocentric chromosomes are more resistant to sterilizing ionizing radiation. Radiat. Res. 2019, 191, 255–261. [Google Scholar] [CrossRef]
- Hughes-Schrader, S.; Ris, H. The diffuse spindle attachment of coccids, verified by the mitotic behavior of induced chromosome fragments. J. Exp. Zool. 1941, 87, 429–456. [Google Scholar] [CrossRef]
- Schrader, F. Notes an the mitotic behavior of long chromosomes. Cytologia 1935, 6, 422–430. [Google Scholar] [CrossRef] [Green Version]
- Jankowska, M.; Fuchs, J.; Klocke, E.; Fojtová, M.; Polanská, P.; Fajkus, J.; Schubert, V.; Houben, A. Holokinetic centromeres and efficient telomere healing enable rapid karyotype evolution. Chromosoma 2015, 124, 519–528. [Google Scholar] [CrossRef]
- Bureš, P.; Zedek, F.; Marková, M. Holocentric chromosomes. In Plant Genome Diversity, Physical Structure, Behaviour and Evolution of Plant Genomes; Greilhuber, J., Dolezel, J., Wendel, J.F., Eds.; Springer: Vienna, Austria, 2013; Volume 2, pp. 187–208. [Google Scholar]
- Da Silva, C.R.M.; González-Elizondo, M.S.; Vanzela, A.L.L. Reduction of chromosome number in Eleocharis subarticulata (Cyperaceae) by multiple translocations. Bot. J. Linn. Soc. 2005, 149, 457–464. [Google Scholar] [CrossRef]
- Yano, O.; Hoshino, A.T. Cytological studies of aneuploidy in Eleocharis kamtschatica (Cyperaceae). Cytologia 2006, 71, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, C.R.; González-Elizondo, M.S.; Laforga Vanzela, A.L. Chromosome reduction in Eleocharis maculosa (Cyperaceae). Cytogenet. Genome Res. 2008, 122, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Márquez-Corro, J.I.; Martín-Bravo, S.; Spalink, D.; Luceño, M.; Escudero, M. Inferring hypothesis-based transitions in clade-specific models of chromosome number evolution in sedges (Cyperaceae). Mol. Phylogenet. Evol. 2019, 135, 203–209. [Google Scholar] [CrossRef]
- Kolodin, P.; Cempírková, H.; Bureš, P.; Horová, L.; Veleba, A.; Francová, J.; Adamec, L.; Zedek, F. Holocentric chromosomes may be an apomorphy of Droseraceae. Plant Syst. Evol. 2018, 304, 1289–1296. [Google Scholar] [CrossRef]
- Sheikh, S.A.; Kondo, K. Differential staining with orcein, Giemsa, CMA, and DAPI for comparative chromosome study of 12 species of Australian Drosera (Droseraceae). Am. J. Bot. 1995, 82, 1278–1286. [Google Scholar] [CrossRef]
- Sheikh, S.A.; Kondo, K.; Hoshi, Y. Study of diffused centromeric nature of Drosera chromosomes. Cytologia 1995, 60, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Veleba, A.; Šmarda, P.; Zedek, F.; Horová, L.; Šmerda, J.; Bureš, P. Evolution of genome size and genomic GC content in carnivorous holokinetics (Droseraceae). Ann. Bot. 2017, 119, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, N.; Tanaka, N. Chromosome Studies in Chionographis (Liliaceae) I. On the holokinetic nature of chromosomes in Chionographis japonica Maxim. Cytologia 1977, 42, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, N.; Tanaka, N. Chromosome studies in Chionographis (Liliaceae). II. Morphological characteristics of the somatic chromosomes of four Japanese members. Cytologia 1979, 44, 935–949. [Google Scholar] [CrossRef] [Green Version]
- Marques, A.; Pedrosa-Harand, A. Holocentromere identity: From the typical mitotic linear structure to the great plasticity of meiotic holocentromeres. Chromosoma 2016, 125, 669–681. [Google Scholar] [CrossRef]
- Marques, A.; Schubert, V.; Houben, A.; Pedrosa-Harand, A. Loss of the line-like holocentromere structure during inverted meiosis in a holocentric plant. Cytogenet. Genome Res. 2016, 148, 128. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Centromere Type | Centromere Subtype | Features | Species |
|---|---|---|---|
| Monocentromere | Point centromere * | Single CENH3/CENP-A-containing nucleosome. | Budding yeast [42] |
| Regional monocentromere * | Single CENH3/CENP-A-containing chromatin domain where mitotic spindle fibers attach. This domain is mostly located in the primary constriction if it is discernible. | Fission yeast [43], e.g., Hordeum vulgare [37,41,44], Secale cereale [45], Cuscuta japonica [46] | |
| Meta-polycentromere | Elongated primary constriction possessing 2-5 CENH3/CENP-A-containing chromatin domains where spindle fibers attach. | Pisum sativum [11,12,13], Lathyrus [12,13] | |
| Holocentromere | Cluster-like holocentromere | Many evenly dispersed CENH3/CENP-A-clusters where spindle fibers attach along the whole chromosome without a groove. | Rhynchospora pubera (meiosis) [47] |
| Line-like holocentromere | Many CENH3/CENP-A-containing chromatin domains forming a contiguous line along the whole chromosome. Spindle fibers attach at CENH3/CENP-A-positive chromatin along a groove. | Luzula elegans [37,41,48,49,50], L. luzuloides [37], R. pubera (mitosis) [47,51], R. tenuis (mitosis) | |
| Holocentromere in C. europaea | Attachment of mitotic spindle fibers along the entire chromosome length, which does not correlate with the distribution of CENH3/CENP-A. It is not yet clear which proteins constitute the centromere in this species. | C. europaea [46] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schubert, V.; Neumann, P.; Marques, A.; Heckmann, S.; Macas, J.; Pedrosa-Harand, A.; Schubert, I.; Jang, T.-S.; Houben, A. Super-Resolution Microscopy Reveals Diversity of Plant Centromere Architecture. Int. J. Mol. Sci. 2020, 21, 3488. https://doi.org/10.3390/ijms21103488
Schubert V, Neumann P, Marques A, Heckmann S, Macas J, Pedrosa-Harand A, Schubert I, Jang T-S, Houben A. Super-Resolution Microscopy Reveals Diversity of Plant Centromere Architecture. International Journal of Molecular Sciences. 2020; 21(10):3488. https://doi.org/10.3390/ijms21103488
Chicago/Turabian StyleSchubert, Veit, Pavel Neumann, André Marques, Stefan Heckmann, Jiri Macas, Andrea Pedrosa-Harand, Ingo Schubert, Tae-Soo Jang, and Andreas Houben. 2020. "Super-Resolution Microscopy Reveals Diversity of Plant Centromere Architecture" International Journal of Molecular Sciences 21, no. 10: 3488. https://doi.org/10.3390/ijms21103488
APA StyleSchubert, V., Neumann, P., Marques, A., Heckmann, S., Macas, J., Pedrosa-Harand, A., Schubert, I., Jang, T. -S., & Houben, A. (2020). Super-Resolution Microscopy Reveals Diversity of Plant Centromere Architecture. International Journal of Molecular Sciences, 21(10), 3488. https://doi.org/10.3390/ijms21103488