Polyphenols with Anti-Amyloid β Aggregation Show Potential Risk of Toxicity Via Pro-Oxidant Properties


Abstract
:1. Amyloid β Aggregation in Alzheimer’s Disease
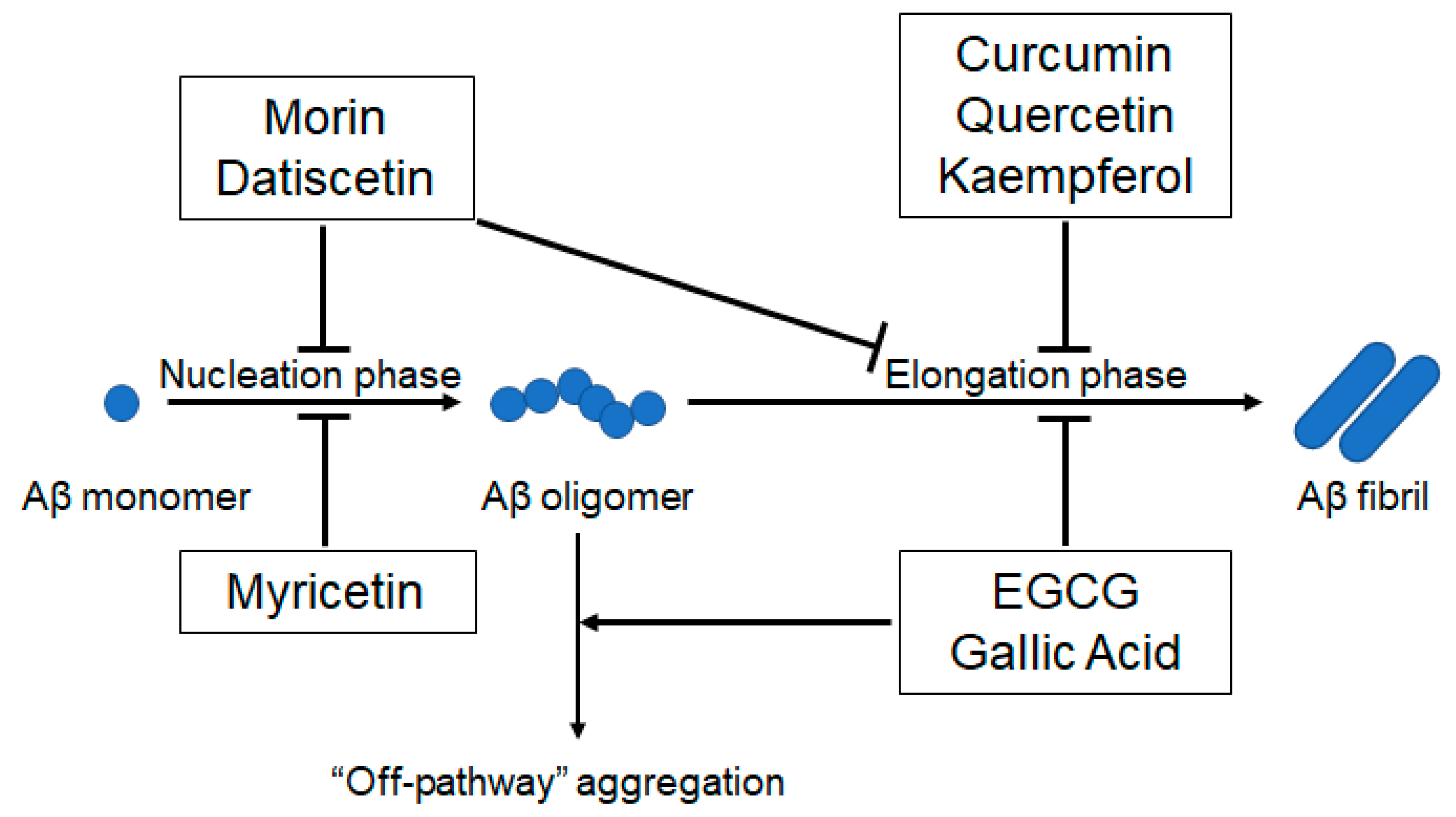
2. Beneficial Anti-Aβ Aggregation and Adverse Pro-Oxidant Effects of Polyphenols
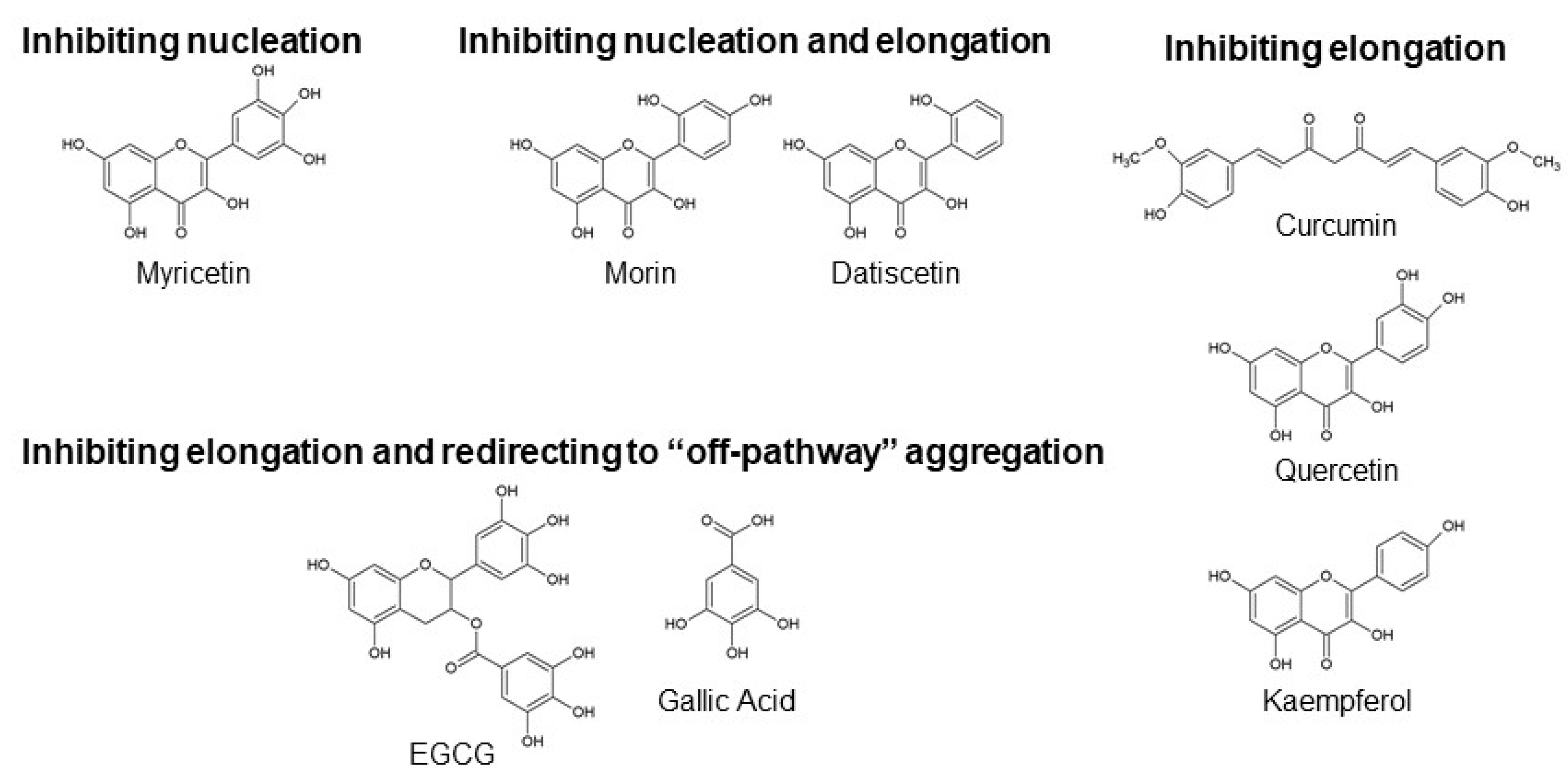
2.1. Polyphenols Involved in Inhibiting Nucleation
Myricetin
2.2. Polyphenols Involved in Inhibiting Nucleation and Elongation
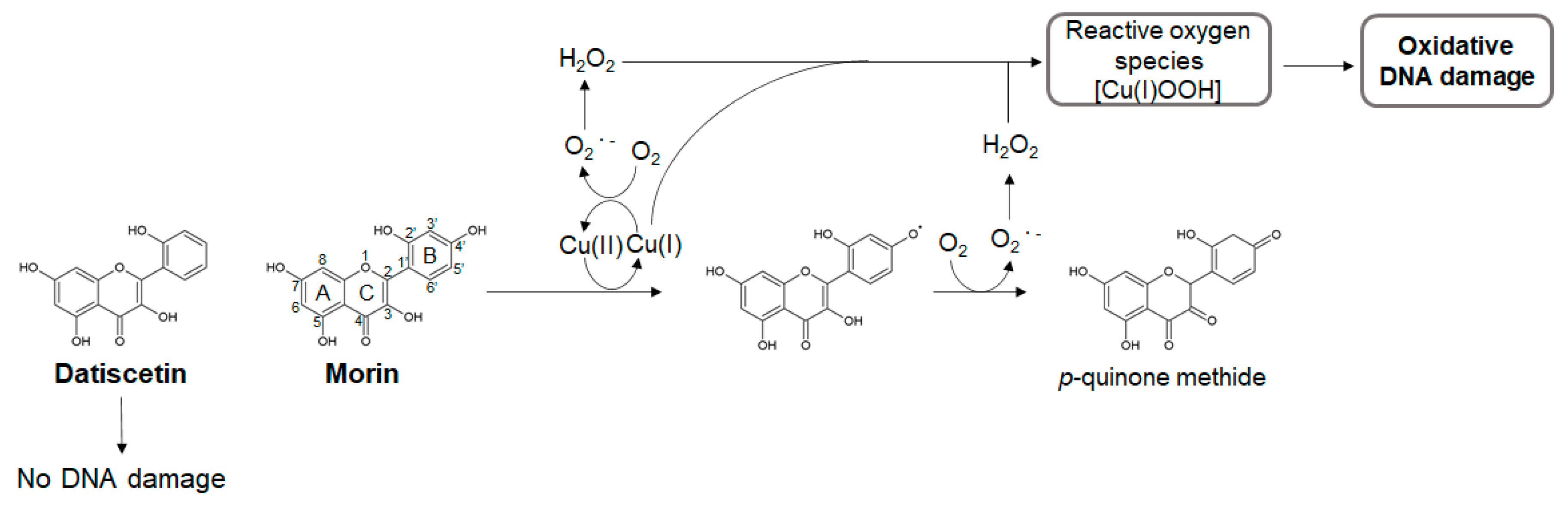
Morin and Datiscetin
2.3. Polyphenols Involved in Inhibiting Elongation
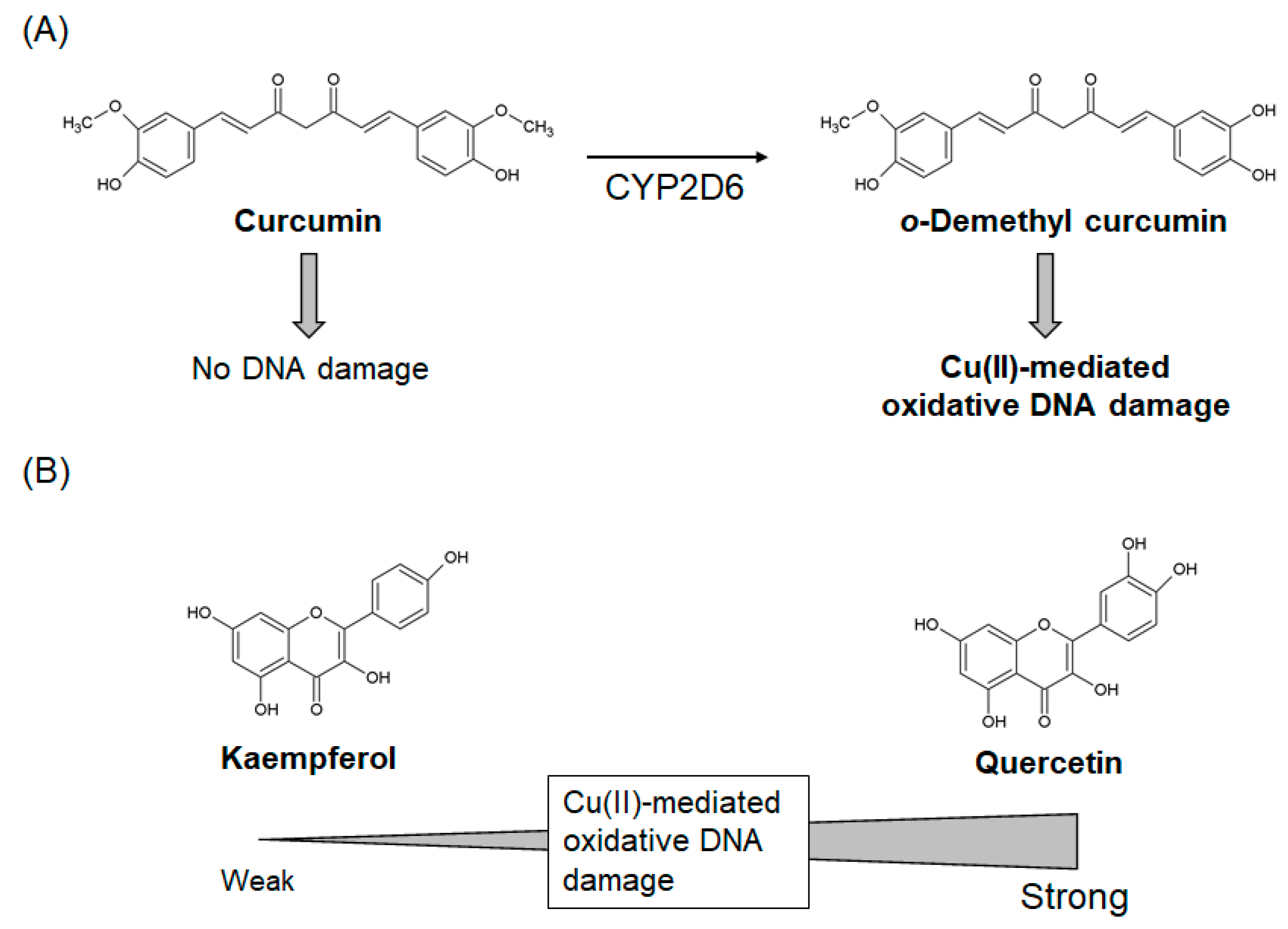
2.3.1. Curcumin
2.3.2. Quercetin and Kaempferol
2.4. Polyphenols Involved in Inhibiting Elongation and Redirecting Aβ Monomers to “Off-Pathway” Aggregation
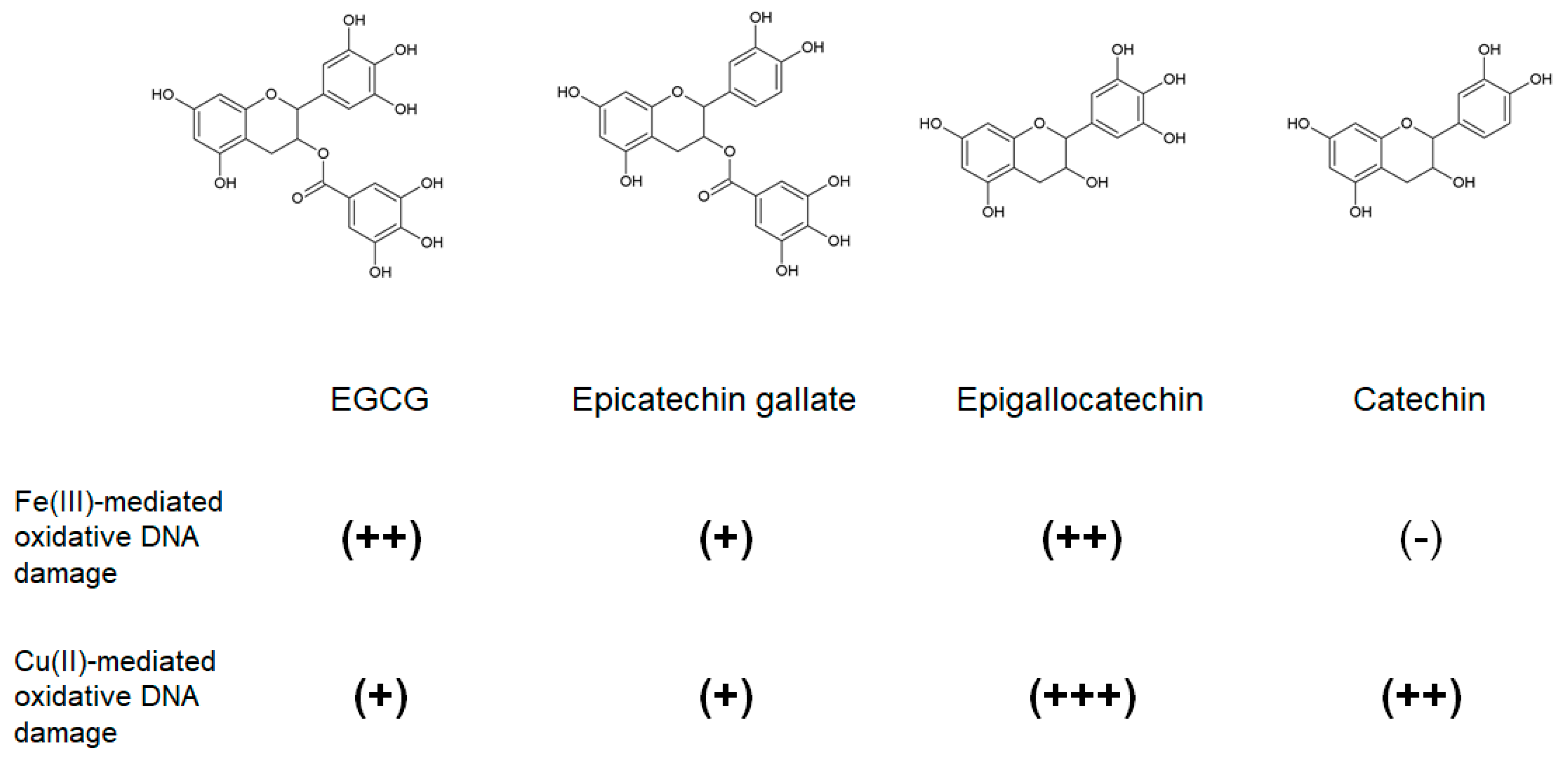
2.4.1. Epigallocatechin Gallate (EGCG) and Other Green Tea Catechins
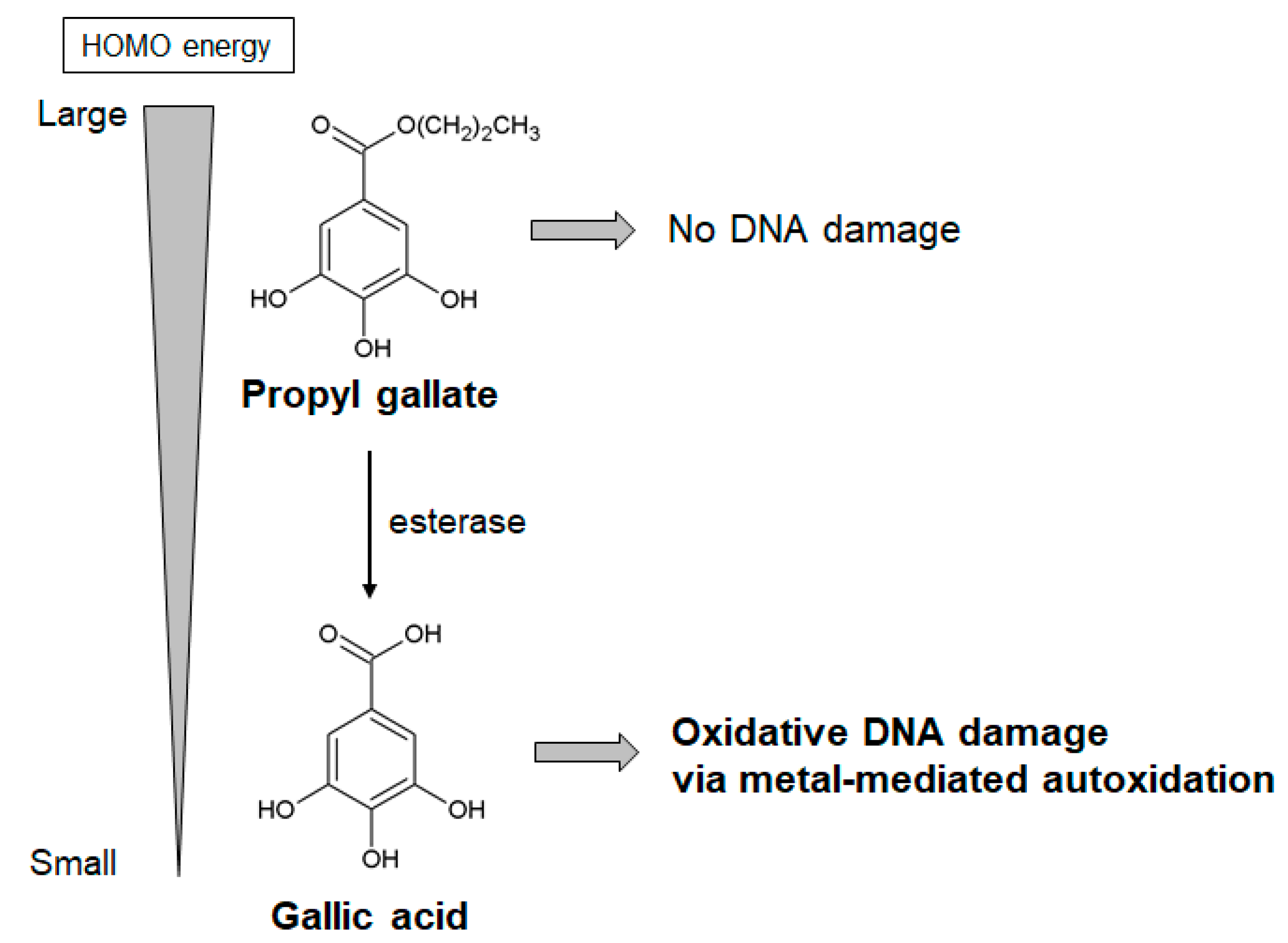
2.4.2. Propyl Gallate and Gallic Acid
3. The Role of Phenolic Hydroxyl Groups in Anti-Aβ Aggregation and Pro-Oxidant Activities of Polyphenols
4. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| Aβ | amyloid β |
| APP | amyloid precursor protein |
| ROS | reactive oxygen species |
| EGCG | epigallocatechin gallate |
| NMR | nuclear magnetic resonance |
| 8-oxodG | 8-oxo-7,8-dihydro-2′-deoxyguanosine |
| PAINS | pan-assay interference compound |
| CYP | cytochrome P450 |
| HOMO energy | highest occupied molecular orbital energy |
References
- Winblad, B.; Amouyel, P.; Andrieu, S.; Ballard, C.; Brayne, C.; Brodaty, H.; Cedazo-Minguez, A.; Dubois, B.; Edvardsson, D.; Feldman, H.; et al. Defeating Alzheimer’s disease and other dementias: A priority for European science and society. Lancet. Neurol. 2016, 15, 455–532. [Google Scholar] [CrossRef] [Green Version]
- Panza, F.; Lozupone, M.; Logroscino, G.; Imbimbo, B.P. A critical appraisal of amyloid-beta-targeting therapies for Alzheimer disease. Nat. Rev. Neurol. 2019, 15, 73–88. [Google Scholar] [CrossRef] [PubMed]
- LaFerla, F.M.; Green, K.N.; Oddo, S. Intracellular amyloid-β in Alzheimer’s disease. Nat. Rev. Neurosci. 2007, 8, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Ayala, S.; Genevaux, P.; Hureau, C.; Faller, P. (Bio)chemical Strategies To Modulate Amyloid-beta Self-Assembly. Acs Chem. Neurosci. 2019, 10, 3366–3374. [Google Scholar] [CrossRef]
- Arosio, P.; Knowles, T.P.; Linse, S. On the lag phase in amyloid fibril formation. Phys. Chem. Chem. Phys. Pccp 2015, 17, 7606–7618. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Walter, J. Phosphorylation of amyloid beta (Aβ) peptides–A trigger for formation of toxic aggregates in Alzheimer’s disease. Aging (Albany Ny) 2011, 3, 803. [Google Scholar] [CrossRef]
- Long, J.M.; Holtzman, D.M. Alzheimer Disease: An Update on Pathobiology and Treatment Strategies. Cell 2019, 179, 312–339. [Google Scholar] [CrossRef]
- Ono, K.; Li, L.; Takamura, Y.; Yoshiike, Y.; Zhu, L.; Han, F.; Mao, X.; Ikeda, T.; Takasaki, J.; Nishijo, H.; et al. Phenolic compounds prevent amyloid beta-protein oligomerization and synaptic dysfunction by site-specific binding. J. Biol. Chem. 2012, 287, 14631–14643. [Google Scholar] [CrossRef] [Green Version]
- Hanaki, M.; Murakami, K.; Akagi, K.; Irie, K. Structural insights into mechanisms for inhibiting amyloid beta42 aggregation by non-catechol-type flavonoids. Bioorganic Med. Chem. 2016, 24, 304–313. [Google Scholar] [CrossRef]
- Masuda, Y.; Fukuchi, M.; Yatagawa, T.; Tada, M.; Takeda, K.; Irie, K.; Akagi, K.-i.; Monobe, Y.; Imazawa, T.; Takegoshi, K. Solid-state NMR analysis of interaction sites of curcumin and 42-residue amyloid β-protein fibrils. Bioorganic Med. Chem. 2011, 19, 5967–5974. [Google Scholar] [CrossRef]
- Hirohata, M.; Hasegawa, K.; Tsutsumi-Yasuhara, S.; Ohhashi, Y.; Ookoshi, T.; Ono, K.; Yamada, M.; Naiki, H. The anti-amyloidogenic effect is exerted against Alzheimer’s beta-amyloid fibrils in vitro by preferential and reversible binding of flavonoids to the amyloid fibril structure. Biochemistry 2007, 46, 1888–1899. [Google Scholar] [CrossRef] [PubMed]
- Ehrnhoefer, D.E.; Bieschke, J.; Boeddrich, A.; Herbst, M.; Masino, L.; Lurz, R.; Engemann, S.; Pastore, A.; Wanker, E.E. EGCG redirects amyloidogenic polypeptides into unstructured, off-pathway oligomers. Nat. Struct. Mol. Biol. 2008, 15, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Chen, X.; Liu, J.; Ma, Q.; Zhuo, Z.; Chen, H.; Zhou, L.; Yang, S.; Zheng, L.; Ning, C.; et al. Gallic acid disruption of Abeta1-42 aggregation rescues cognitive decline of APP/PS1 double transgenic mouse. Neurobiol. Dis. 2019, 124, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Dhouafli, Z.; Cuanalo-Contreras, K.; Hayouni, E.A.; Mays, C.E.; Soto, C.; Moreno-Gonzalez, I. Inhibition of protein misfolding and aggregation by natural phenolic compounds. Cell. Mol. Life Sci. Cmls 2018, 75, 3521–3538. [Google Scholar] [CrossRef] [PubMed]
- Perez-Jimenez, J.; Neveu, V.; Vos, F.; Scalbert, A. Systematic analysis of the content of 502 polyphenols in 452 foods and beverages: An application of the phenol-explorer database. J. Agric. Food Chem. 2010, 58, 4959–4969. [Google Scholar] [CrossRef] [PubMed]
- Velander, P.; Wu, L.; Henderson, F.; Zhang, S.; Bevan, D.R.; Xu, B. Natural product-based amyloid inhibitors. Biochem. Pharmacol. 2017, 139, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Ono, K.; Hamaguchi, T.; Noguchi-Shinohara, M. Natural Phenolic Compounds as Therapeutic and Preventive Agents for Cerebral Amyloidosis. Adv. Exp. Med. Biol. 2015, 863, 79–94. [Google Scholar]
- Sen, S.; Chakraborty, R. Antioxidant Supplements: Friend or Foe? In Free Radicals in Human Health and Disease 2015, 293–322. [Google Scholar] [CrossRef]
- Martin, K.R.; Appel, C.L. Polyphenols as dietary supplements: A double-edged sword. Nutr. Diet. Suppl. 2009, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lambert, J.D.; Sang, S.; Yang, C.S. Possible controversy over dietary polyphenols: Benefits vs risks. Chem. Res. Toxicol. 2007, 20, 583–585. [Google Scholar] [CrossRef]
- Cieślik, E.; Gręda, A.; Adamus, W. Contents of polyphenols in fruit and vegetables. Food Chem. 2006, 94, 135–142. [Google Scholar] [CrossRef]
- Murata, M.; Kawanishi, S. Mechanisms of oxidative DNA damage induced by carcinogenic arylamines. Front. Biosci. (Landmark Ed.) 2011, 16, 1132–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, A.F.; Dos Santos, C.N.; Ventura, M.R. Polyphenols, their Metabolites and Derivatives as Drug Leads. Curr. Pharm. Des. 2018, 24, 2188–2207. [Google Scholar] [CrossRef] [PubMed]
- Carregosa, D.; Carecho, R.; Figueira, I.; Santos, C.N. Low-Molecular Weight Metabolites from Polyphenols as Effectors for Attenuating Neuroinflammation. J. Agric. Food Chem. 2020, 68, 1790–1807. [Google Scholar] [CrossRef] [Green Version]
- Sahu, S.C.; Flynn, T.J.; Bradlaw, J.A.; Roth, W.L.; Barton, C.N.; Yates, J.G. Pro-oxidant effects of the flavonoid myricetin on rat hepatocytes in culture. Toxicol. Methods 2001, 11, 277–283. [Google Scholar] [CrossRef]
- Park, H.; Park, S.; Bazer, F.W.; Lim, W.; Song, G. Myricetin treatment induces apoptosis in canine osteosarcoma cells by inducing DNA fragmentation, disrupting redox homeostasis, and mediating loss of mitochondrial membrane potential. J. Cell. Physiol. 2018, 233, 7457–7466. [Google Scholar] [CrossRef]
- Knickle, A.; Fernando, W.; Greenshields, A.L.; Rupasinghe, H.P.V.; Hoskin, D.W. Myricetin-induced apoptosis of triple-negative breast cancer cells is mediated by the iron-dependent generation of reactive oxygen species from hydrogen peroxide. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 118, 154–167. [Google Scholar] [CrossRef]
- Sahu, S.C.; Gray, G.C. Interactions of flavonoids, trace metals, and oxygen: Nuclear DNA damage and lipid peroxidation induced by myricetin. Cancer Lett. 1993, 70, 73–79. [Google Scholar] [CrossRef]
- Arif, H.; Sohail, A.; Farhan, M.; Rehman, A.A.; Ahmad, A.; Hadi, S.M. Flavonoids-induced redox cycling of copper ions leads to generation of reactive oxygen species: A potential role in cancer chemoprevention. Int. J. Biol. Macromol. 2018, 106, 569–578. [Google Scholar] [CrossRef]
- Jomova, K.; Hudecova, L.; Lauro, P.; Simunkova, M.; Alwasel, S.H.; Alhazza, I.M.; Valko, M. A Switch between Antioxidant and Prooxidant Properties of the Phenolic Compounds Myricetin, Morin, 3’,4’-Dihydroxyflavone, Taxifolin and 4-Hydroxy-Coumarin in the Presence of Copper(II) Ions: A Spectroscopic, Absorption Titration and DNA Damage Study. Molecules (Basel, Switz.) 2019, 24, 4335. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, M.; Haneda, M.; Naruse, M.; Murakami, K. Prooxidant activity of flavonoids: Copper-dependent strand breaks and the formation of 8-hydroxy-2’-deoxyguanosine in DNA. Mol. Genet. Metab. 1999, 68, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, C.A.; Swartz, C.; Maronpot, R.; Davis, J.; Recio, L.; Koyanagi, M.; Hayashi, S.M. Genotoxicity evaluation of the flavonoid, myricitrin, and its aglycone, myricetin. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2015, 83, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Silva, I.D.; Gaspar, J.; Rodrigues, A.; da Costa, G.G.; Laires, A.; Rueff, J. Mechanisms of myricetin mutagenicity in V79 cells: Involvement of radicalar species. Teratog. Carcinog. Mutagenesis 1996, 16, 253–268. [Google Scholar] [CrossRef]
- Yen, G.C.; Duh, P.D.; Tsai, H.L.; Huang, S.L. Pro-oxidative properties of flavonoids in human lymphocytes. Biosci. Biotechnol. Biochem. 2003, 67, 1215–1222. [Google Scholar] [CrossRef] [Green Version]
- Sahu, S.C.; Gray, G.C. Lipid peroxidation and DNA damage induced by morin and naringenin in isolated rat liver nuclei. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 1997, 35, 443–447. [Google Scholar] [CrossRef]
- Mori, Y.; Kato, S.; Fujisawa, Y.; Ohnishi, S.; Hiraku, Y.; Kawanishi, S.; Murata, M.; Oikawa, S. Mechanisms of DNA damage induced by morin, an inhibitor of amyloid beta-peptide aggregation. Free Radic. Res. 2019, 53, 115–123. [Google Scholar] [CrossRef]
- Roy, A.S.; Samanta, S.K.; Ghosh, P.; Tripathy, D.R.; Ghosh, S.K.; Dasgupta, S. Cell cytotoxicity and serum albumin binding capacity of the morin-Cu(ii) complex and its effect on deoxyribonucleic acid. Mol. Biosyst. 2016, 12, 2818–2833. [Google Scholar] [CrossRef]
- Jurado, J.; Alejandre-Duran, E.; Alonso-Moraga, A.; Pueyo, C. Study on the mutagenic activity of 13 bioflavonoids with the Salmonella Ara test. Mutagenesis 1991, 6, 289–295. [Google Scholar] [CrossRef]
- Yoshino, M.; Haneda, M.; Naruse, M.; Htay, H.H.; Tsubouchi, R.; Qiao, S.L.; Li, W.H.; Murakami, K.; Yokochi, T. Prooxidant activity of curcumin: Copper-dependent formation of 8-hydroxy-2’-deoxyguanosine in DNA and induction of apoptotic cell death. Toxicol. Vitr. Int. J. Publ. Assoc. Bibra 2004, 18, 783–789. [Google Scholar] [CrossRef]
- Woo, J.H.; Kim, Y.H.; Choi, Y.J.; Kim, D.G.; Lee, K.S.; Bae, J.H.; Min, D.S.; Chang, J.S.; Jeong, Y.J.; Lee, Y.H.; et al. Molecular mechanisms of curcumin-induced cytotoxicity: Induction of apoptosis through generation of reactive oxygen species, down-regulation of Bcl-XL and IAP, the release of cytochrome c and inhibition of Akt. Carcinogenesis 2003, 24, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Urbina-Cano, P.; Bobadilla-Morales, L.; Ramírez-Herrera, M.A.; Corona-Rivera, J.R.; Mendoza-Magaña, M.L.; Troyo-Sanromán, R.; Corona-Rivera, A. DNA damage in mouse lymphocytes exposed to curcumin and copper. J. Appl. Genet. 2006, 47, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Sakano, K.; Kawanishi, S. Metal-mediated DNA damage induced by curcumin in the presence of human cytochrome P450 isozymes. Arch. Biochem. Biophys. 2002, 405, 223–230. [Google Scholar] [CrossRef]
- National Toxicology Program. NTP Toxicology and Carcinogenesis Studies of Turmeric Oleoresin (CAS No. 8024-37-1) (Major Component 79%–85% Curcumin, CAS No. 458-37-7) in F344/N Rats and B6C3F1 Mice (Feed Studies). Natl. Toxicol. Program Tech. Rep. Ser. 1993, 427, 1–275. [Google Scholar]
- Lapidot, T.; Walker, M.D.; Kanner, J. Antioxidant and prooxidant effects of phenolics on pancreatic β-cells in vitro. J. Agric. Food Chem. 2002, 50, 7220–7225. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, N.; Kawanishi, S. Distinct mechanisms of DNA damage in apoptosis induced by quercetin and luteolin. Free Radic. Res. 2000, 33, 623–633. [Google Scholar] [CrossRef]
- Yamashita, N.; Tanemura, H.; Kawanishi, S. Mechanism of oxidative DNA damage induced by quercetin in the presence of Cu(II). Mutat. Res. 1999, 425, 107–115. [Google Scholar] [CrossRef]
- Resende, F.A.; Vilegas, W.; Dos Santos, L.C.; Varanda, E.A. Mutagenicity of flavonoids assayed by bacterial reverse mutation (Ames) test. Molecules (Basel, Switz.) 2012, 17, 5255–5268. [Google Scholar] [CrossRef]
- National Toxicology Program. Toxicology and Carcinogenesis Studies of Quercetin (CAS No. 117-39-5) in F344 Rats (Feed Studies). Natl. Toxicol. Program Tech. Rep. Ser. 1992, 409, 1–171. [Google Scholar]
- Gaspar, J.; Rodrigues, A.; Laires, A.; Silva, F.; Costa, S.; Monteiro, M.J.; Monteiro, C.; Rueff, J. On the mechanisms of genotoxicity and metabolism of quercetin. Mutagenesis 1994, 9, 445–449. [Google Scholar] [CrossRef]
- Pamukcu, A.M.; Yalciner, S.; Hatcher, J.F.; Bryan, G.T. Quercetin, a rat intestinal and bladder carcinogen present in bracken fern (Pteridium aquilinum). Cancer Res. 1980, 40, 3468–3472. [Google Scholar]
- Chan, M.M.; Soprano, K.J.; Weinstein, K.; Fong, D. Epigallocatechin-3-gallate delivers hydrogen peroxide to induce death of ovarian cancer cells and enhances their cisplatin susceptibility. J. Cell. Physiol. 2006, 207, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Hasumi, K.; Woo, J.-T.; Nagai, K.; Wachi, M. Generation of hydrogen peroxide primarily contributes to the induction of Fe (II)-dependent apoptosis in Jurkat cells by (−)-epigallocatechin gallate. Carcinogenesis 2004, 25, 1567–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.K.; Loo, G. Effects of epigallocatechin gallate and quercetin on oxidative damage to cellular DNA. Mutat. Res. 2000, 459, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, A.; Oikawa, S.; Murata, M.; Hiraku, Y.; Kawanishi, S. (-)-Epigallocatechin gallate causes oxidative damage to isolated and cellular DNA. Biochem. Pharmacol. 2003, 66, 1769–1778. [Google Scholar] [CrossRef]
- National Toxicology Program. Toxicology Studies of green tea extract in F344/NTac Rats and B6C3F1/N mice and toxicology and carcinogenesis studies of green tea extract in Wistar Han [Crl:WI(Han)] rats and B6C3F1/N Mice (gavage studies). Natl. Toxicol. Program Tech. Rep. Ser. 2016, 585, 1–192. [Google Scholar]
- Hu, J.; Webster, D.; Cao, J.; Shao, A. The safety of green tea and green tea extract consumption in adults—Results of a systematic review. Regul. Toxicol. Pharmacol. Rtp 2018, 95, 412–433. [Google Scholar] [CrossRef]
- Lee, K.W.; Hur, H.J.; Lee, H.J.; Lee, C.Y. Antiproliferative effects of dietary phenolic substances and hydrogen peroxide. J. Agric. Food Chem. 2005, 53, 1990–1995. [Google Scholar] [CrossRef]
- Isuzugawa, K.; Inoue, M.; Ogihara, Y. Catalase contents in cells determine sensitivity to the apoptosis inducer gallic acid. Biol. Pharm. Bull. 2001, 24, 1022–1026. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Takemura, G.; Koshiji, M.; Hayakawa, Y.; Kanoh, M.; Maruyama, R.; Ohno, Y.; Minatoguchi, S.; Akao, S.; Fukuda, K.; et al. Gallic acid induces vascular smooth muscle cell death via hydroxyl radical production. Heart Vessel. 2000, 15, 90–99. [Google Scholar] [CrossRef]
- Kobayashi, H.; Oikawa, S.; Hirakawa, K.; Kawanishi, S. Metal-mediated oxidative damage to cellular and isolated DNA by gallic acid, a metabolite of antioxidant propyl gallate. Mutat. Res. 2004, 558, 111–120. [Google Scholar] [CrossRef]
- Yoshino, M.; Haneda, M.; Naruse, M.; Htay, H.; Iwata, S.; Tsubouchi, R.; Murakami, K. Prooxidant action of gallic acid compounds: Copper-dependent strand breaks and the formation of 8-hydroxy-2′-deoxyguanosine in DNA. Toxicol. Vitr. 2002, 16, 705–709. [Google Scholar] [CrossRef]
- Galati, G.; Lin, A.; Sultan, A.M.; O’Brien, P.J. Cellular and in vivo hepatotoxicity caused by green tea phenolic acids and catechins. Free Radic. Biol. Med. 2006, 40, 570–580. [Google Scholar] [CrossRef]
- Abarikwu, S.O.; Durojaiye, M.; Alabi, A.; Asonye, B.; Akiri, O. Curcumin protects against gallic acid-induced oxidative stress, suppression of glutathione antioxidant defenses, hepatic and renal damage in rats. Ren. Fail. 2016, 38, 321–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, M.; Zhu, M.; Wang, L.; Yu, S. Anti-tumor effects and associated molecular mechanisms of myricetin. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 120, 109506. [Google Scholar] [CrossRef] [PubMed]
- Semwal, D.K.; Semwal, R.B.; Combrinck, S.; Viljoen, A. Myricetin: A Dietary Molecule with Diverse Biological Activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, K.; Yoshiike, Y.; Takashima, A.; Hasegawa, K.; Naiki, H.; Yamada, M. Potent anti-amyloidogenic and fibril-destabilizing effects of polyphenols in vitro: Implications for the prevention and therapeutics of Alzheimer’s disease. J. Neurochem. 2003, 87, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, T.; Ono, K.; Murase, A.; Yamada, M. Phenolic compounds prevent Alzheimer’s pathology through different effects on the amyloid-beta aggregation pathway. Am. J. Pathol. 2009, 175, 2557–2565. [Google Scholar] [CrossRef] [Green Version]
- Kataria, R.; Sobarzo-Sanchez, E.; Khatkar, A. Role of Morin in Neurodegenerative Diseases: A Review. Curr. Top. Med. Chem. 2018, 18, 901–907. [Google Scholar] [CrossRef]
- Caselli, A.; Cirri, P.; Santi, A.; Paoli, P. Morin: A Promising Natural Drug. Curr. Med. Chem. 2016, 23, 774–791. [Google Scholar] [CrossRef]
- Sinha, K.; Ghosh, J.; Sil, P.C. Morin and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 928, 453–471. [Google Scholar]
- Kim, H.; Park, B.S.; Lee, K.G.; Choi, C.Y.; Jang, S.S.; Kim, Y.H.; Lee, S.E. Effects of naturally occurring compounds on fibril formation and oxidative stress of beta-amyloid. J. Agric. Food Chem. 2005, 53, 8537–8541. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Qu, J.; Zhang, W.; Bai, M.; Zhou, Q.; Zhang, Z.; Li, Z.; Miao, J. Morin reverses neuropathological and cognitive impairments in APPswe/PS1dE9 mice by targeting multiple pathogenic mechanisms. Neuropharmacology 2016, 108, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Singh, M.; Kumar, P.; Vikram, V.; Mishra, N. Development and characterization of morin hydrate loaded microemulsion for the management of Alzheimer’s disease. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1620–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, K.; Irie, K. Three Structural Features of Functional Food Components and Herbal Medicine with Amyloid beta42 Anti-Aggregation Properties. Molecules (Basel,Switz.) 2019, 24. [Google Scholar]
- Gupta, S.C.; Sung, B.; Kim, J.H.; Prasad, S.; Li, S.; Aggarwal, B.B. Multitargeting by turmeric, the golden spice: From kitchen to clinic. Mol. Nutr. Food Res. 2013, 57, 1510–1528. [Google Scholar] [CrossRef] [PubMed]
- Kocaadam, B.; Sanlier, N. Curcumin, an active component of turmeric (Curcuma longa), and its effects on health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2889–2895. [Google Scholar] [CrossRef]
- Jin, T.R. Curcumin and dietary polyphenol research: Beyond drug discovery. Acta Pharmacol. Sin. 2018, 39, 779–786. [Google Scholar] [CrossRef] [Green Version]
- Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Ramirez-Tortosa, M. Curcumin and Health. Molecules (Basel, Switz.) 2016, 21, 264. [Google Scholar] [CrossRef]
- Schubert, D.; Currais, A.; Goldberg, J.; Finley, K.; Petrascheck, M.; Maher, P. Geroneuroprotectors: Effective Geroprotectors for the Brain. Trends Pharm. Sci 2018, 39, 1004–1007. [Google Scholar] [CrossRef]
- Ono, K.; Hasegawa, K.; Naiki, H.; Yamada, M. Curcumin has potent anti-amyloidogenic effects for Alzheimer’s beta-amyloid fibrils in vitro. J. Neurosci. Res. 2004, 75, 742–750. [Google Scholar] [CrossRef]
- Yang, F.; Lim, G.P.; Begum, A.N.; Ubeda, O.J.; Simmons, M.R.; Ambegaokar, S.S.; Chen, P.P.; Kayed, R.; Glabe, C.G.; Frautschy, S.A.; et al. Curcumin inhibits formation of amyloid beta oligomers and fibrils, binds plaques, and reduces amyloid in vivo. J. Biol. Chem. 2005, 280, 5892–5901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Alloza, M.; Borrelli, L.A.; Rozkalne, A.; Hyman, B.T.; Bacskai, B.J. Curcumin labels amyloid pathology in vivo, disrupts existing plaques, and partially restores distorted neurites in an Alzheimer mouse model. J. Neurochem. 2007, 102, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Baell, J.; Walters, M.A. Chemistry: Chemical con artists foil drug discovery. Nature 2014, 513, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef]
- Malik, P.; Mukherjee, T.K. Structure-Function Elucidation of Antioxidative and Prooxidative Activities of the Polyphenolic Compound Curcumin. Chin. J. Biol. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Alzahrani, A.M.; Rajendran, P. The Multifarious Link between Cytochrome P450s and Cancer. Oxidative Med. Cell. Longev. 2020, 2020, 3028387. [Google Scholar] [CrossRef]
- Wang, W.; Sun, C.; Mao, L.; Ma, P.; Liu, F.; Yang, J.; Gao, Y. The biological activities, chemical stability, metabolism and delivery systems of quercetin: A review. Trends Food Sci. Technol. 2016, 56, 21–38. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.J.; Wang, Y.Q.; Cui, Y.L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules (Basel, Switz.) 2019, 24, 1123. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Khan, H.; Ullah, H.; Aschner, M.; Cheang, W.S.; Akkol, E.K. Neuroprotective Effects of Quercetin in Alzheimer’s Disease. Biomolecules 2019, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Aliaga, K.; Bermejo-Bescos, P.; Benedi, J.; Martin-Aragon, S. Quercetin and rutin exhibit antiamyloidogenic and fibril-disaggregating effects in vitro and potent antioxidant activity in APPswe cells. Life Sci. 2011, 89, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-M.; Li, S.-Q.; Wu, W.-L.; Zhu, X.-Y.; Wang, Y.; Yuan, H.-Y. Effects of long-term treatment with quercetin on cognition and mitochondrial function in a mouse model of Alzheimer’s disease. Neurochem. Res. 2014, 39, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- Dajas, F.; Abin-Carriquiry, J.A.; Arredondo, F.; Blasina, F.; Echeverry, C.; Martinez, M.; Rivera, F.; Vaamonde, L. Quercetin in brain diseases: Potential and limits. Neurochem. Int. 2015, 89, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Parveen, N.; Shadab, G.G. The dual clastogenic and anti-clastogenic properties of quercetin is dose dependent. Front. Biosci. (Sch. Ed.) 2017, 9, 139–153. [Google Scholar]
- Walle, T. Methylation of dietary flavones increases their metabolic stability and chemopreventive effects. Int. J. Mol. Sci. 2009, 10, 5002–5019. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Mi, M.; Ling, W.; Zhu, J.; Zhang, Q.; Wei, N.; Zhou, Y.; Tang, Y.; Yuan, J. Structurally related cytotoxic effects of flavonoids on human cancer cells in vitro. Arch. Pharmacal Res. 2008, 31, 1137–1144. [Google Scholar] [CrossRef]
- Hara, Y.; Yang, C.S.; Isemura, M.; Tomita, I. Health benefits of green tea: An evidence-based approach; CABI: Wallingford, UK, 2017; ISBN 9781786392398. [Google Scholar]
- Islam, M.A. Cardiovascular effects of green tea catechins: Progress and promise. Recent Pat. Cardiovasc. Drug Discov. 2012, 7, 88–99. [Google Scholar] [CrossRef]
- Eng, Q.Y.; Thanikachalam, P.V.; Ramamurthy, S. Molecular understanding of Epigallocatechin gallate (EGCG) in cardiovascular and metabolic diseases. J. Ethnopharmacol. 2018, 210, 296–310. [Google Scholar] [CrossRef]
- Singh, N.A.; Mandal, A.K.; Khan, Z.A. Potential neuroprotective properties of epigallocatechin-3-gallate (EGCG). Nutr. J. 2016, 15, 60. [Google Scholar] [CrossRef] [Green Version]
- Chakrawarti, L.; Agrawal, R.; Dang, S.; Gupta, S.; Gabrani, R. Therapeutic effects of EGCG: A patent review. Expert Opin. Ther. Pat. 2016, 26, 907–916. [Google Scholar] [CrossRef]
- Ngo, S.T.; Truong, D.T.; Tam, N.M.; Nguyen, M.T. EGCG inhibits the oligomerization of amyloid beta (16-22) hexamer: Theoretical studies. J. Mol. Graph. Model. 2017, 76, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Palhano, F.L.; Lee, J.; Grimster, N.P.; Kelly, J.W. Toward the molecular mechanism(s) by which EGCG treatment remodels mature amyloid fibrils. J. Am. Chem. Soc. 2013, 135, 7503–7510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieschke, J.; Russ, J.; Friedrich, R.P.; Ehrnhoefer, D.E.; Wobst, H.; Neugebauer, K.; Wanker, E.E. EGCG remodels mature alpha-synuclein and amyloid-beta fibrils and reduces cellular toxicity. Proc. Natl. Acad. Sci. USA 2010, 107, 7710–7715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezai-Zadeh, K.; Arendash, G.W.; Hou, H.; Fernandez, F.; Jensen, M.; Runfeldt, M.; Shytle, R.D.; Tan, J. Green tea epigallocatechin-3-gallate (EGCG) reduces beta-amyloid mediated cognitive impairment and modulates tau pathology in Alzheimer transgenic mice. Brain Res. 2008, 1214, 177–187. [Google Scholar] [CrossRef]
- Rezai-Zadeh, K.; Shytle, D.; Sun, N.; Mori, T.; Hou, H.; Jeanniton, D.; Ehrhart, J.; Townsend, K.; Zeng, J.; Morgan, D.; et al. Green tea epigallocatechin-3-gallate (EGCG) modulates amyloid precursor protein cleavage and reduces cerebral amyloidosis in Alzheimer transgenic mice. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 8807–8814. [Google Scholar] [CrossRef] [Green Version]
- Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; et al. Scientific opinion on the safety of green tea catechins. Efsa J. 2018, 16. [Google Scholar]
- Dekant, W.; Fujii, K.; Shibata, E.; Morita, O.; Shimotoyodome, A. Safety assessment of green tea based beverages and dried green tea extracts as nutritional supplements. Toxicol. Lett. 2017, 277, 104–108. [Google Scholar] [CrossRef]
- Bertram, B.; Bollow, U.; Rajaee-Behbahani, N.; Burkle, A.; Schmezer, P. Induction of poly(ADP-ribosyl)ation and DNA damage in human peripheral lymphocytes after treatment with (-)-epigallocatechin-gallate. Mutat. Res. 2003, 534, 77–84. [Google Scholar] [CrossRef]
- Wang, D.; Wei, Y.; Wang, T.; Wan, X.; Yang, C.S.; Reiter, R.J.; Zhang, J. Melatonin attenuates (-)-epigallocatehin-3-gallate-triggered hepatotoxicity without compromising its downregulation of hepatic gluconeogenic and lipogenic genes in mice. J. Pineal Res. 2015, 59, 497–507. [Google Scholar] [CrossRef]
- Wang, D.; Wang, Y.; Wan, X.; Yang, C.S.; Zhang, J. Green tea polyphenol (-)-epigallocatechin-3-gallate triggered hepatotoxicity in mice: Responses of major antioxidant enzymes and the Nrf2 rescue pathway. Toxicol. Appl. Pharmacol. 2015, 283, 65–74. [Google Scholar] [CrossRef]
- Takumi-Kobayashi, A.; Ogura, R.; Morita, O.; Nishiyama, N.; Kasamatsu, T. Involvement of hydrogen peroxide in chromosomal aberrations induced by green tea catechins in vitro and implications for risk assessment. Mutat. Res. 2008, 657, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Kucera, O.; Mezera, V.; Moravcova, A.; Endlicher, R.; Lotkova, H.; Drahota, Z.; Cervinkova, Z. In vitro toxicity of epigallocatechin gallate in rat liver mitochondria and hepatocytes. Oxidative Med. Cell. Longev. 2015, 2015, 476180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isbrucker, R.A.; Bausch, J.; Edwards, J.A.; Wolz, E. Safety studies on epigallocatechin gallate (EGCG) preparations. Part 1: Genotoxicity. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2006, 44, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Rho, T.; Choi, M.S.; Jung, M.; Kil, H.W.; Hong, Y.D.; Yoon, K.D. Identification of fermented tea (Camellia sinensis) polyphenols and their inhibitory activities against amyloid-beta aggregation. Phytochemistry 2019, 160, 11–18. [Google Scholar] [CrossRef]
- Xie, H.; Wang, J.R.; Yau, L.F.; Liu, Y.; Liu, L.; Han, Q.B.; Zhao, Z.; Jiang, Z.H. Catechins and procyanidins of Ginkgo biloba show potent activities towards the inhibition of beta-amyloid peptide aggregation and destabilization of preformed fibrils. Molecules (Basel, Switz.) 2014, 19, 5119–5134. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.; Kantham, S.; Rao, V.M.; Palanivelu, M.K.; Pham, H.L.; Shaw, P.N.; McGeary, R.P.; Ross, B.P. Metal chelation, radical scavenging and inhibition of Abeta(4)(2) fibrillation by food constituents in relation to Alzheimer’s disease. Food Chem. 2016, 199, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Singh, A.; Mishra, A. Gallic acid: Molecular rival of cancer. Environ. Toxicol. Pharmacol. 2013, 35, 473–485. [Google Scholar] [CrossRef]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. Rsc Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Bastianetto, S.; Yao, Z.X.; Papadopoulos, V.; Quirion, R. Neuroprotective effects of green and black teas and their catechin gallate esters against beta-amyloid-induced toxicity. Eur. J. Neurosci. 2006, 23, 55–64. [Google Scholar] [CrossRef]
- Liu, Y.; Pukala, T.L.; Musgrave, I.F.; Williams, D.M.; Dehle, F.C.; Carver, J.A. Gallic acid is the major component of grape seed extract that inhibits amyloid fibril formation. Bioorganic Med. Chem. Lett. 2013, 23, 6336–6340. [Google Scholar] [CrossRef]
- LeVine, H.; Lampe, L.; Abdelmoti, L.; Augelli-Szafran, C.E. Dihydroxybenzoic Acid Isomers Differentially Dissociate Soluble Biotinyl-Aβ(1–42) Oligomers. Biochemistry 2012, 51, 307–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Toxicology Program. NTP Carcinogenesis Bioassay of Propyl Gallate (CAS No. 121-79-9) in F344/N Rats and B6C3F1 Mice (Feed Study). Natl. Toxicol. Program Tech. Rep. Ser. 1982, 240, 1–152. [Google Scholar]
- Locatelli, C.; Filippin-Monteiro, F.B.; Creczynski-Pasa, T.B. Alkyl esters of gallic acid as anticancer agents: A review. Eur. J. Med. Chem. 2013, 60, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, K.; Yoshida, M.; Oikawa, S.; Kawanishi, S. Base oxidation at 5’ site of GG sequence in double-stranded DNA induced by UVA in the presence of xanthone analogues: Relationship between the DNA-damaging abilities of photosensitizers and their HOMO energies. Photochem. Photobiol. 2003, 77, 349–355. [Google Scholar] [CrossRef]
- Ishii, T.; Mori, T.; Tanaka, T.; Mizuno, D.; Yamaji, R.; Kumazawa, S.; Nakayama, T.; Akagawa, M. Covalent modification of proteins by green tea polyphenol (–)-epigallocatechin-3-gallate through autoxidation. Free Radic. Biol. Med. 2008, 45, 1384–1394. [Google Scholar] [CrossRef]
- Xiao, Y.; Ma, B.; McElheny, D.; Parthasarathy, S.; Long, F.; Hoshi, M.; Nussinov, R.; Ishii, Y. Aβ (1–42) fibril structure illuminates self-recognition and replication of amyloid in Alzheimer’s disease. Nat. Struct. Mol. Biol. 2015, 22, 499–505. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Murakami, K.; Uno, M.; Nakagawa, Y.; Katayama, S.; Akagi, K.-i.; Masuda, Y.; Takegoshi, K.; Irie, K. Site-specific inhibitory mechanism for amyloid β42 aggregation by catechol-type flavonoids targeting the Lys residues. J. Biol. Chem. 2013, 288, 23212–23224. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, S.; Furukawa, A.; Asada, H.; Hirakawa, K.; Kawanishi, S. Catechins induce oxidative damage to cellular and isolated DNA through the generation of reactive oxygen species. Free Radic. Res. 2003, 37, 881–890. [Google Scholar] [CrossRef]
- Nedumpully-Govindan, P.; Kakinen, A.; Pilkington, E.H.; Davis, T.P.; Chun Ke, P.; Ding, F. Stabilizing Off-pathway Oligomers by Polyphenol Nanoassemblies for IAPP Aggregation Inhibition. Sci. Rep. 2016, 6, 19463. [Google Scholar] [CrossRef] [Green Version]
- Przygonska, K.; Pacewicz, M.; Sadowska, W.; Poznanski, J.; Bal, W.; Dadlez, M. His6, His13, and His14 residues in Abeta 1-40 peptide significantly and specifically affect oligomeric equilibria. Sci. Rep. 2019, 9, 9449. [Google Scholar] [CrossRef]
- Kong, X.; Zhao, Z.; Lei, X.; Zhang, B.; Dai, D.; Jiang, L. Interaction of metal ions with the His13-His14 sequence relevant to Alzheimer’s disease. J. Phys. Chem. A 2015, 119, 3528–3534. [Google Scholar] [CrossRef] [PubMed]
- Vaz, F.N.C.; Fermino, B.L.; Haskel, M.V.L.; Wouk, J.; de Freitas, G.B.L.; Fabbri, R.; Montagna, E.; Rocha, J.B.T.; Bonini, J.S. The Relationship Between Copper, Iron, and Selenium Levels and Alzheimer Disease. Biol. Trace Elem. Res. 2018, 181, 185–191. [Google Scholar] [CrossRef] [PubMed]
- James, S.A.; Churches, Q.I.; de Jonge, M.D.; Birchall, I.E.; Streltsov, V.; McColl, G.; Adlard, P.A.; Hare, D.J. Iron, Copper, and Zinc Concentration in Abeta Plaques in the APP/PS1 Mouse Model of Alzheimer’s Disease Correlates with Metal Levels in the Surrounding Neuropil. Acs Chem. Neurosci. 2017, 8, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Yugay, D.; Goronzy, D.P.; Kawakami, L.M.; Claridge, S.A.; Song, T.B.; Yan, Z.; Xie, Y.H.; Gilles, J.; Yang, Y.; Weiss, P.S. Copper Ion Binding Site in beta-Amyloid Peptide. Nano Lett. 2016, 16, 6282–6289. [Google Scholar] [CrossRef] [PubMed]
- Mira, L.; Fernandez, M.T.; Santos, M.; Rocha, R.; Florencio, M.H.; Jennings, K.R. Interactions of flavonoids with iron and copper ions: A mechanism for their antioxidant activity. Free Radic. Res. 2002, 36, 1199–1208. [Google Scholar] [CrossRef]
- Vestergaard, M.; Kerman, K.; Tamiya, E. An electrochemical approach for detecting copper-chelating properties of flavonoids using disposable pencil graphite electrodes: Possible implications in copper-mediated illnesses. Anal. Chim. Acta 2005, 538, 273–281. [Google Scholar] [CrossRef]
- Teng, Y.; Zhao, J.; Ding, L.; Ding, Y.; Zhou, P. Complex of EGCG with Cu(II) Suppresses Amyloid Aggregation and Cu(II)-Induced Cytotoxicity of alpha-Synuclein. Molecules (Basel, Switz.) 2019, 24. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anti-Aβ Aggregation Effect | Polyphenol | Toxic Effects Associated with Pro-Oxidant Properties | Concentration or Dose Showing Toxic Effects of Polyphenols |
|---|---|---|---|
| Inhibiting nucleation | Myricetin | Cytotoxicity | |
| Cytotoxicity linked with ROS generation | Cell: 20 μM [25], 50 μM [26,27] | ||
| Genotoxicity | |||
| Oxidative DNA damage | Cell: 20 μM [28], 50 μM [29] DNA: 5 μM [30], 200 μM [31] | ||
| Mutagenic activity | Bacteria: 0.628 μmol/plate [32] Cell: 42 μM [33] | ||
| Inhibiting nucleation and elongation | Morin | Genotoxicity | |
| Oxidative DNA damage | Cell: 100 μM [34] DNA: 5 μM [30],10 μM [35], 20 μM [36], 100 μM [37] | ||
| Mutagenic activity | Bacteria: 0.149 μmol/plate [38] | ||
| Datiscetin | No report | ||
| Inhibiting elongation | Curcumin | Cytotoxicity | |
| Cytotoxicity linked with ROS generation | Cell: 5 μM [39], 50 μM [40] | ||
| Genotoxicity | |||
| DNA damage in cultured cell | Cell: 50 μM [41] | ||
| Curcumin metabolite-mediated oxidative damage in isolated DNA | DNA: 2 μM [42] | ||
| Tumorigenicity | |||
| Colon mucosal hyperplasia and hepatocellular adenoma in rats and mice treated with turmeric oleoresin containing curcumin (79%-85%), respectively | Colon hyperplasia: 2000 mg/kg/day (male rats) [43] Hepatocellular adenoma: 520 mg/kg/day (male mice) [43], 1620 mg/kg/day (female mice) [43] | ||
| Inhibiting elongation (continued) | Quercetin | Cytotoxicity | |
| Cytotoxicity linked with ROS generation | Cell: 50 μM [44] | ||
| Genotoxicity | |||
| Oxidative DNA damage | Cell: 30 μM [45], 50 μM [29], 100 μM [34] DNA: 10 μM [46] | ||
| Mutagenic activity | Bacteria: 0.121 μmol/plate [47] Cell: 2.2 μM [48], 32.5 μM [49] | ||
| Carcinogenesis | |||
| Renal tubule adenocarcinomas and intestinal and bladder cancer in rats | Renal tubule adenocarcinomas: 1900 mg/kg/day (male rats) [48] Intestinal and bladder cancer: 27.8 mM/rat (male, cumulative dose) [50], 25.3 mM/rat (female, cumulative dose) [50] | ||
| Kaempferol | Genotoxicity | ||
| Oxidative DNA damage | Cell: 50 μM [29] | ||
| Mutagenic activity | Bacteria: 0.143 μmol/plate [47] | ||
| Inhibiting elongation and redirecting to “off-pathway” aggregation | EGCG | Cytotoxicity | |
| Cytotoxicity linked with ROS generation | Cell: 2 μM [51], 12.5 μM [52] | ||
| Genotoxicity | |||
| Oxidative DNA damage | Cell: 100 μM [53], 200 μM [54] DNA: 5 μM [54] | ||
| Hepatotoxicity and gastrointestinal toxicity | |||
| Gastrointestinal tract and liver lesion in rats and mice treated with green tea extract containing EGCG (48.4%) | Gastrointestinal tract lesion: 1000 mg/kg/day (male and female rats) [55] Liver lesion: 1000 mg/kg/day (male and female rats) [55], 300 mg/kg/day (male mice) [55] | ||
| High dose intake-associated liver damage in humans | Human: 704 mg/day [56] | ||
| Gallic acid | Cytotoxicity | ||
| Cytotoxicity linked with ROS generation | Cell: 74 μM [57], 294 μM [58,59] | ||
| Genotoxicity | |||
| Oxidative DNA damage | DNA: 5 μM [60], 200 μM [61] | ||
| Hepatotoxicity and nephrotoxicity | |||
| Liver damage in mice and rats, and renal injury in rats | Liver damage: 200 mg/kg/day (male mice) [62], 100 mg/kg/day (male rats) [63] Renal injury: 100 mg/kg/day (male rats) [63] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, H.; Murata, M.; Kawanishi, S.; Oikawa, S. Polyphenols with Anti-Amyloid β Aggregation Show Potential Risk of Toxicity Via Pro-Oxidant Properties. Int. J. Mol. Sci. 2020, 21, 3561. https://doi.org/10.3390/ijms21103561
Kobayashi H, Murata M, Kawanishi S, Oikawa S. Polyphenols with Anti-Amyloid β Aggregation Show Potential Risk of Toxicity Via Pro-Oxidant Properties. International Journal of Molecular Sciences. 2020; 21(10):3561. https://doi.org/10.3390/ijms21103561
Chicago/Turabian StyleKobayashi, Hatasu, Mariko Murata, Shosuke Kawanishi, and Shinji Oikawa. 2020. "Polyphenols with Anti-Amyloid β Aggregation Show Potential Risk of Toxicity Via Pro-Oxidant Properties" International Journal of Molecular Sciences 21, no. 10: 3561. https://doi.org/10.3390/ijms21103561
APA StyleKobayashi, H., Murata, M., Kawanishi, S., & Oikawa, S. (2020). Polyphenols with Anti-Amyloid β Aggregation Show Potential Risk of Toxicity Via Pro-Oxidant Properties. International Journal of Molecular Sciences, 21(10), 3561. https://doi.org/10.3390/ijms21103561