Carbonic Anhydrase Sensitivity to Pesticides: Perspectives for Biomarker Development



Abstract
:1. Introduction
2. In Vitro Sensitivity of CA Activity to Pesticides in Animals
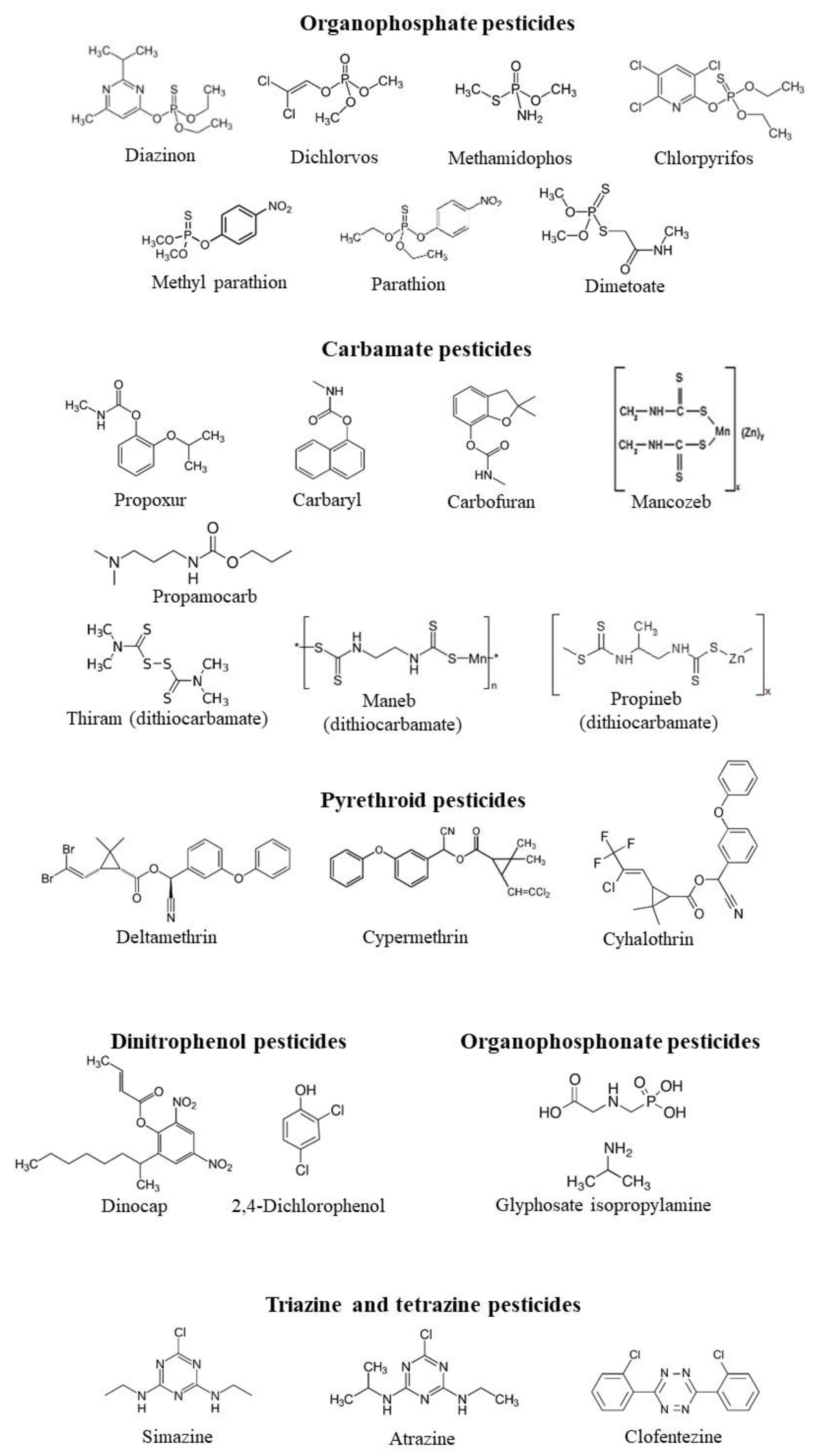
2.1. In Vitro Effect of Different Classes of Pesticides on CA Activity
2.2. Comparison among Species
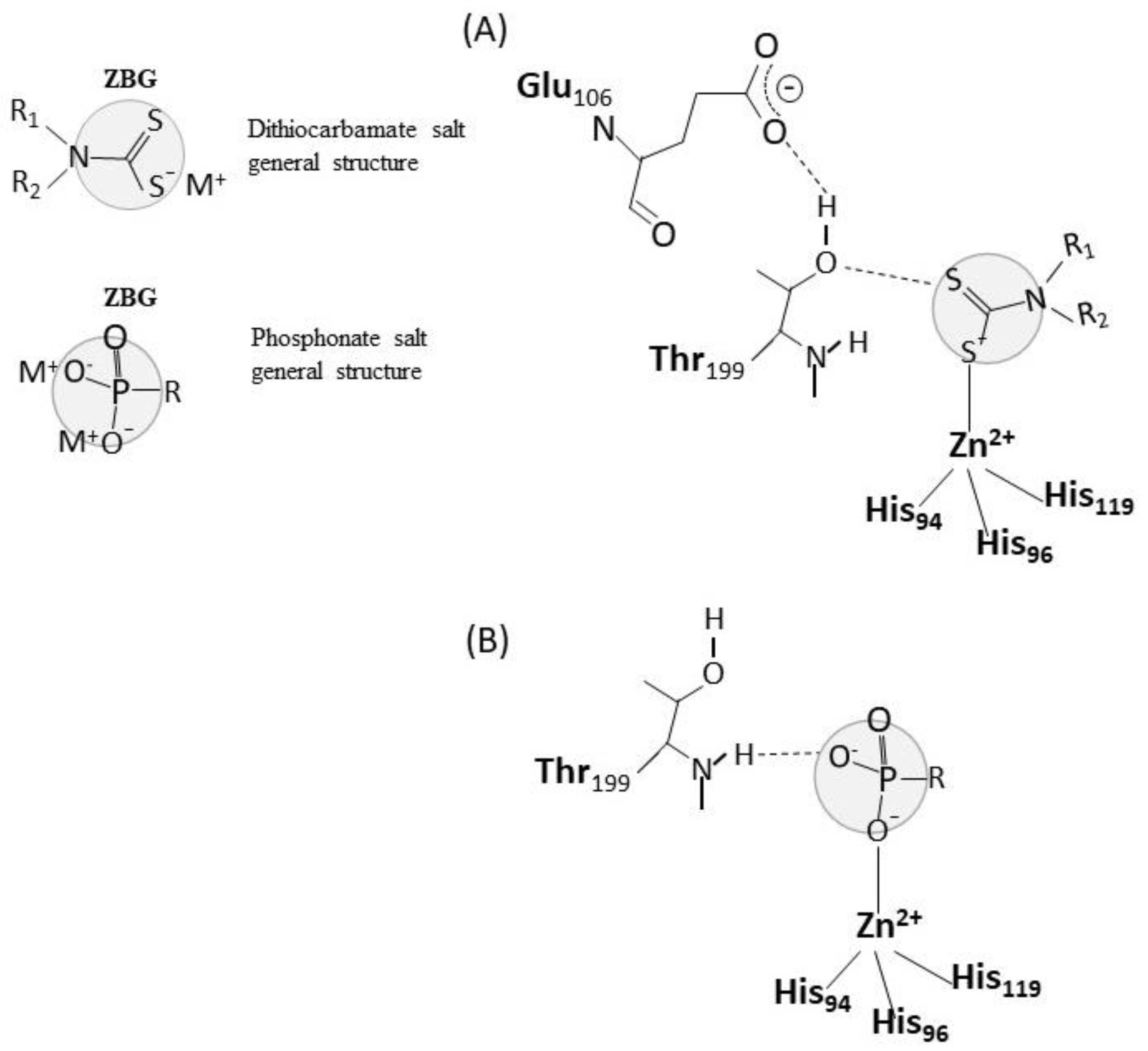
2.3. Mechanisms of Action
3. In Vivo Sensitivity of CA Activity to Pesticides in Animals
4. Perspectives for Pesticide Biomarker Development
4.1. Potential Biomarker of Effect
4.2. Potential Biomarker of Exposure
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- DiMario, R.J.; Machingura, M.C.; Waldrop, G.L.; Moroney, J.V. The many types of carbonic anhydrases in photosynthetic organisms. Plant Sci. 2018, 268, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Nocentini, A.; Supuran, C.T.; Capasso, C. Bacterial ι-carbonic anhydrase: A new active class of carbonic anhydrase identified in the genome of the Gram-negative bacterium Burkholderia territorii. J. Enzyme Inhib. Med. Chem. 2020, 35, 1060–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lionetto, M.G.; Caricato, R.; Giordano, M.E.; Schettino, T. The complex relationship between metals and carbonic anhydrase: New insights and perspectives. Int. J. Mol. Sci. 2016, 17, 127. [Google Scholar] [CrossRef] [Green Version]
- Di Fiore, A.; Monti, M.; Scaloni, A.; De Simone, G.; Monti, S.M. Protective role of carbonic anhydrases III and VII in cellular defense mechanisms upon redox unbalance. Oxid. Med. Cell. Longev. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Supuran, C.T. How many carbonic anhydrase inhibition mechanisms exist? J. Enzyme Inhib. Med. Chem. 2016, 31, 345–360. [Google Scholar] [CrossRef]
- Supuran, C.T.; Scozzafava, A. Carbonic anhydrases as targets for medicinal chemistry. Bioorg. Med. Chem. Lett. 2007, 15, 4336–4350. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrase inhibitors. Bioorg. Med. Chem. Lett. 2010, 15, 3467–3474. [Google Scholar] [CrossRef]
- Ye, M.; Beach, J.; Martin, J.W.; Senthilselvan, A. Pesticide exposures and respiratory health in general populations. J. Environ. Sci. 2017, 51, 361–370. [Google Scholar] [CrossRef]
- Lamberth, C.; Jeanmart, S.; Luksch, T.; Plant, A. Current Challenges and Trends in the Discovery of Agrochemicals. Science 2013, 341, 742–746. [Google Scholar] [CrossRef]
- Ntzani, E.E.; Chondrogiorgi, M.; Ntritsos, G.; Evangelou, E.; Tzoulaki, I. Literature review on epidemiological studies linking exposure to pesticides and health effects. EFSA 2013, EN-497, 159. [Google Scholar] [CrossRef]
- Stanley, J.; Preetha, G. Pesticide Toxicity to Non-Target Organisms: Exposure, Toxicity and Risk Assessment Methodologies; Springer: Basel, Switzerland, 2016. [Google Scholar]
- Rojas-García, A.E.; Medina-Díaz, I.M.; Robledo-Marenco, M.L.; Barrón-Vivanco, B.S.; Pérez-Herrera, N. Pesticide biomarkers. In Pesticides in the Modern World—Pests Control and Pesticides Exposure and Toxicity Assessment; Stoytcheva, M., Ed.; InTech: London, UK, 2011; pp. 161–190. [Google Scholar]
- Araud, M. Biological markers of human exposure to pesticides. In Pesticides in the Modern World—Pests Control and Pesticides Exposure and Toxicity Assessment; Stoytcheva, M., Ed.; InTech: London, UK, 2011; pp. 154–196. [Google Scholar]
- Lionetto, M.G.; Caricato, R.; Calisi, A.; Giordano, M.E.; Schettino, T. Acetylcholinesterase as a biomarker in Environmental and occupational medicine: New insights and future perspectives. BioMed Res. Intern. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lionetto, M.G.; Caricato, R.; Giordano, M.E. Pollution Biomarkers in Environmental and Human Biomonitoring. Open Biomark. J. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lionetto, M.G.; Caricato, R.; Giordano, M.E.; Erroi, E.; Schettino, T. Carbonic anhydrase as pollution biomarker: An ancient enzyme with a new use. Int. J. Environ. Res. Public Health 2012, 9, 3965–3977. [Google Scholar] [CrossRef] [Green Version]
- Gangemi, S.; Miozzi, E.; Teodoro, M.; Briguglio, G.; De Luca, A.; Alibrando, C.; Polito, I.; Libra, M. Occupational exposure to pesticides as a possible risk factor for the development of chronic diseases in humans. Mol. Med. Rep. 2016, 14, 4475–4488. [Google Scholar] [CrossRef] [Green Version]
- Dodgson, S.J. Why are there carbonic anhydrases in the liver? Biochem. Cell. Biol. 1991, 69, 761–763. [Google Scholar] [CrossRef]
- Wistrand, P.J. Carbonic Anhydrase III in Liver and Muscle of Male Rats Purification and Properties. Upsala J. Med Sci. 2009, 107, 77–88. [Google Scholar] [CrossRef]
- Gilmour, K.M. New insights into the many functions of carbonic anhydrase in fish gills. Respir. Physiol. Neurobiol. 2012, 184, 223–230. [Google Scholar] [CrossRef]
- Ceyhun, S.B.; Sentürk, M.; Erdogăn, O.; Küfrevioğlu, Ö.I. In vitro and in vivo effects of some pesticides on carbonic anhydrase enzyme from rainbow trout (Oncorhynchus mykiss) gills. Pestic. Biochem. Phys. 2010, 97, 177–181. [Google Scholar] [CrossRef]
- Dogăn, S. The in vitro effects of some pesticides on carbonic anhydrase activity of Oncorhynchus mykiss and Cyprinus carpio carpio fish. J. Hazard. Mater. 2006, A132, 171–176. [Google Scholar] [CrossRef]
- Demirdağ, R.; Yerlikaya, E.; Aksakal, E.; Küfrevioğlu, Ö.I.; Ekinci, D. Influence of pesticides on the pH regulatory enzyme, carbonic anhydrase, from European Seabass liver and bovine erythrocytes. Environ. Toxicol. Pharmacol. 2012, 34, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Kılınç, N.; Isgor, M.; Şengül, B.; Beydemir, Ş. Influence of pesticide exposure on carbonic anhydrase II from sheep stomach. Toxicol. Ind. Health 2015, 31, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Işik, S.; Köçkar, F.; Özensoy, Ö.; Guler, O.O.; Arslan, O. Differential in vitro effects of some pesticides on carbonic anhydrase activities from some freshwater and seawater fish erythrocytes. Fresen. Environ. Bull. 2004, 13, 25–29. [Google Scholar]
- Sinan, S.; Gencer, N.; Turana, Y.; Arslanb, O. In vitro inhibition of the carbonic anhydrase from saanen goat (Capra hircus) with pesticides. Pestic. Biochem. Phys. 2007, 88, 307–311. [Google Scholar] [CrossRef]
- Kolayli, S.; Karahalil, F.; Sahin, H.; Dincer, B.; Supuran, C.T. Characterization and inhibition studies of an α-carbonic anhydrase from the endangered sturgeon species Acipenser gueldenstaedti. J. Enzyme Inhib. Med. Chem. 2011, 26, 895–900. [Google Scholar] [CrossRef] [Green Version]
- Yerlinkaya, E. Inhibitory effects of some pesticides and metals on carbonic anhydrase purified from shabut fish (Barbus grypus) gill tissue. Appl. Ecol. Env. Res. 2019, 17, 13473–13483. [Google Scholar]
- Gençer, N.; Ergün, A.; Demir, D. In vitro effects of some herbicides and fungicides on human erythrocyte carbonic anhydrase activity. Fresen. Environ. Bull. 2012, 21, 549–552. [Google Scholar]
- Soydan, E.; Güler, A.; Bıyık, S.; Şentürk, M.; Supuran, C.T.; Ekinci, D. Carbonic anhydrase from Apis mellifera: Purification and inhibition by pesticides. J. Enzyme Inhib. Med. Chem. 2017, 32, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Lionetto, M.G.; Caricato, R.; Erroi, E.; Giordano, M.E.; Schettino, T. Carbonic anhydrase-based environmental bioassay. Int. J. Envir. Anal. Chem. 2005, 85, 895–903. [Google Scholar] [CrossRef]
- Ekinci, E.; Beydemir, S. Risk assessment of pesticides and fungicides for acid–base regulation and salt transport in rainbow trout tissues. Pest. Biochem. Physiol. 2010, 97, 66–70. [Google Scholar] [CrossRef]
- Caglayan, C.; Taslimi, P.; Türk, C.; Kandemir, F.M.; Demir, Y.; Gulcin, İ. Purification and characterization of the carbonic anhydrase enzyme from horse mackerel (Trachurus trachurus) muscle and the impact of some metal ions and pesticides on enzyme activity. Comp. Biochem. Physiol. C. 2019, 226, 108605. [Google Scholar] [CrossRef] [PubMed]
- Caglayan, C.; Taslimi, P.; Türk, C.; Gulcin, İ.; Kandemir, F.M.; Demir, Y.; Beydemir, Ş. Inhibition effects of some pesticides and heavy metals on carbonic anhydrase enzyme activity purified from horse mackerel (Trachurus trachurus) gill tissues. Environ. Sci. Pollut. R. 2020, 27, 10607–10616. [Google Scholar] [CrossRef] [PubMed]
- Kose, L.P.; Gülçin, I.; Özdemir, H.; Atasever, A.; Alwasel, S.H.; Supuran, C.T. The effects of some avermectins on bovine carbonic anhydrase enzyme. J. Enzyme Inhib. Med. Chem. 2016, 31, 773–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranauskiene, L.; Matulis, D. Herbicide oryzalin inhibits human carbonic anhydrases in vitro. J. Biochem. Mol. Toxicol. 2017, 31, e21894. [Google Scholar] [CrossRef] [PubMed]
- Bloomquist, J.R. Chloride channels as tools for developing selective insecticides. Arch Insect Biochem Physiol. 2003, 54, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Sant’anna, V.; de Souza, W.; Vommaro, R.C. Anthelmintic effect of herbicidal dinitroanilines on the nematode model Caenorhabditis elegans. Exp Parasitol. 2016, 167, 43–49. [Google Scholar] [CrossRef]
- Carta, F.; Aggarwal, M.; Maresca, A.; Scozzafava, A.; McKenna, R.; Masini, E.; Supuran, C.T. Dithiocarbamates Strongly Inhibit Carbonic Anhydrases and Show Antiglaucoma Action in Vivo. J. Med. Chem. 2012, 55, 1721–1730. [Google Scholar] [CrossRef] [Green Version]
- Bozdag, M.; Carta, F.; Vullo, D.; Akdemir, A.; Isik, S.; Lanzi, C.; Scozzafava, A.; Masini, E.; Supuran, C.T. Synthesis of a new series of dithiocarbamates with effective human carbonic anhydrase inhibitory activity and antiglaucoma action. Bioorg. Med. Chem. 2015, 23, 2368–2376. [Google Scholar] [CrossRef]
- Temperini, C.; Innocenti, A.; Guerri, A.; Scozzafava, A.; Rusconi, S.; Supuran, C.T. Phosph(on)ate as a zinc-binding group in metalloenzyme inhibitors: X-ray crystal structure of the antiviral drug foscarnet complexed to human carbonic anhydrase I. Bioorganic Med. Chem. Lett. 2007, 17, 2210–2215. [Google Scholar] [CrossRef]
- Ilies, M.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase activators. In Carbonic Anhydrase—Its Inhibitors and Activators, Supuran, C.T.; Scozzafava, A., Conway, J., Eds.; CRC Press: Boca Raton, FL, USA, 2004; p. 317. [Google Scholar]
- Martínez-Tabche, L.; Beatriz, E.M.; Galar, I. Parathion and salinity effects on gills and mesonephros carbonic anhydrase activity of the fish Oreochromis hornorum. B Environ. Contam. Toxicol. 1992, 49, 929–934. [Google Scholar] [CrossRef]
- Topal, A.; Atamanalp, M.; Oruç, E.; Demir, Y.; Beydemir, Ş.; Işık, A. In vivo changes in carbonic anhydrase activity and histopathology of gill and liver tissues after acute exposure to chlorpyrifos in rainbow trout. Arh. Hig. Rada Toksikol. 2014, 65, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulukan, E.; Köktürk, M.; Ceylan, H.; Beydemir, Ş.; Işik, M.; Atamanalp, M.; Ceyhun, S.B. An approach to clarify the effect mechanism of glyphosate on body malformations during embryonic development of zebrafish (Daino rerio). Chemosphere 2017, 180, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Paulino, M.G.; Sakuragui, M.M.; Fernandes, M.N. Effects of atrazine on the gill cells and ionic balance in a neotropical fish, Prochilodus lineatus. Chemosphere 2012, 86, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esbaugh, A.J.; Tufts, B.L. The structure and function of carbonic anhydrase isozymes in the respiratory system of vertebrates. Respir. Physiol. Neurobiol. 2006, 154, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Mboge, M.Y.; Mahon, B.P.; McKenna, R.; Frost, S.C. Carbonic anhydrases: Role in pH control and cancer. Metabolites 2018, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.H.; Muhammad, E.; Zheng, C.; Hershkovitz, E.; Alkrinawi, S.; Loewenthal, N.; Parvari, R.; Muallem, S. Essential role of carbonic anhydrase XII in secretory gland fluid and HCO3− secretion revealed by diseasecausing human mutation. J. Physiol. 2015, 593, 5299–5312. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T. Carbonic anhydrases and metabolism. Metabolites 2018, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Dai, H.Y.; Hong, C.C.; Liang, S.C.; Yan, M.D.; Lai, G.M.; Cheng, A.L.; Chuang, S.E. Carbonic anhydrase III promotes transformation and invasion capability in hepatoma cells through FAK signaling pathway. Mol. Carcinog. 2008, 47, 956–963. [Google Scholar] [CrossRef]
- Monti, D.M.; De Simone, G.; Langella, E.; Supuran, C.T.; Di Fiore, A.; Monti, S.M. Insights into the role of reactive sulfhydryl groups of Carbonic Anhydrase III and VII during oxidative damage. J. Enzyme Inhib. Med. Chem. 2017, 32, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Caricato, R.; Giordano, M.E.; Schettino, T.; Lionetto, M.G. Functional involvement of carbonic anhydrase in the lysosomal response to cadmium exposure in Mytilus galloprovincialis digestive gland. Front. Physiol. 2018, 9, 319. [Google Scholar] [CrossRef]
- Caricato, R.; Giordano, M.E.; Schettino, T.; Maisano, M.; Mauceri, A.; Giannetto, A.; Cappello, T.; Parrino, V.; Ancora, S.; Caliani, I.; et al. Carbonic anhydrase integrated into a multimarker approach for the detection of the stress status induced by pollution exposure in Mytilus galloprovincialis: A field case study. Sci. Total Environ. 2019, 690, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Harju, A.K.; Bootorabi, F.; Kuuslahti, M.; Supuran, C.T.; Parkkila, S. Carbonic anhydrase III: A neglected isozyme is stepping into the limelight. J. Enzyme Inhib. Med. Chem. 2013, 28, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, W.L.; Chu, S.C.; Lai, J.C.; Yang, S.F.; Chiou, H.L.; Hsieh, Y.S. Alternations in quantities and activities of erythrocyte cytosolic carbonic anhydrase isoenzymes in glucose-6-phosphate dehydrogenase-deficient individuals. Clin. Chim. Acta 2001, 314, 195–201. [Google Scholar] [CrossRef]
- Kuo, W.H.; Yang, S.F.; Hsieh, Y.S.; Tsai, C.-S.; Hwang, W.-L.; Chu, S.-C. Differential expression of carbonic anhydrase isoenzymes invarious types of anemia. Clin. Chim. Acta 2005, 351, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Cabiscol, E.; Levine, R.L. The phosphatase activity of carbonic anhydrase III is reversibly regulated by glutathiolation. PNAS 1996, 93, 4170–4174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallis, R.J.; Poland, B.W.; Chatterjee, T.K.; Fisher, R.A.; Richard, S.D.; Honzatko, B.; Thomas, J.A. Crystal structure of S-glutathiolated carbonic anhydrase III. FEBS Lett. 2011, 482, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Ceyhun, S.B.; Şentürk, M.; Ekinci, D.; Erdoğan, O.; Çiltaş, A.; Kocaman, E.M. Deltamethrin attenuates antioxidant defense system and induces the expression of heat shock protein 70 in rainbow trout. Comp. Biochem. Physiol. C 2010, 152, 215–223. [Google Scholar] [CrossRef]
- Rossi, A.S.; Fantón, N.; Michlig, M.P.; Repettic, M.R.; Cazenave, J. Fish inhabiting rice fields: Bioaccumulation, oxidative stress and neurotoxic effects after pesticides application. Ecol. Indic. 2020, 113, 106186. [Google Scholar] [CrossRef]
- Banerjee, B.D.; Seth, V.; Bhattacharya, A.; Pasha, S.T.; Chakraborty, A.K. Biochemical effects of some pesticides on lipid peroxidation and free-radical scavengers. Toxicol. Lett. 1999, 107, 33–47. [Google Scholar] [CrossRef]
- Ranjbar, A.; Pasalar, P.; Abdollahi, M. Induction of oxidative stress and acetylcholinesterase inhibition in organophosphorus pesticide manufacturing workers. Hum. Exp. Toxicol. 2002, 21, 179–182. [Google Scholar] [CrossRef]
- Olakkaran, S.; Purayl, A.K.; Antony, A.; Mallikarjunaiah, S.; Hunasanahally, G.; Puttaswamygowda, G.H. Oxidative stress-mediated genotoxicity of malathion in human lymphocytes. Mutat Res-Gen. Tox. En. 2020, 849, 503138. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, H.; Rappaport, S.M.; Törnqvist, M. Protein adductomics: Methodologies for untargeted screening of adducts to serum albumin and hemoglobin in human blood samples. High Throughput. 2019, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsillach, J.; Costa, L.G.; Furlong, C.E. Protein adducts as biomarkers of exposure to organophosphorus compounds. Toxicology 2013, 307, 46–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Pesticides | IC50 (µM) | Ki (µM) | Inhibition Type | Species | Tissue | Ref. |
|---|---|---|---|---|---|---|
| Organophosphate | ||||||
| Diazinon | 0.267 | n.d. | n.d. | Oncorynchus mykiss | gills | [22] |
| 6840 | n.d. | n.d. | Oncorynchus mykiss | erythroytes | [23] | |
| 3920 | n.d. | n.d. | Cyprinus carpio | erytrocytes | [23] | |
| Dichlorvos | 23.19 | 14.17 | n.d. | Dicentrarchus labrax | liver | [24] |
| 10.178 | 5.872 | n.d. | Bos taurus | erytrocytes | [24] | |
| 2.690 | 3.700 ± 1.670 | non competitive | Ovis aries | stomach | [25] | |
| Methamidophos | 38.73 | 26.16 | n.d. | Dicentrarchus labrax | liver | [24] |
| 2.129 | 1.636 | n.d. | Bos taurus | erytrocytes | [24] | |
| Chlorpyrifos | 2.640 | 2.175 | n.d. | Dicentrarchus labrax | liver | [24] |
| 84.12 | 53.28 | n.d. | Bos taurus | erytrocytes | [24] | |
| Methyl parathion | 0.850 | 0.376 | n.d. | Dicentrarchus labrax | liver | [24] |
| 2.150 | 1.174 | n.d. | Bos taurus | erytrocytes | [24] | |
| 620.0 | n.d. | n.d. | Scorpaena porcus | erytrocyte | [26] | |
| 2900 | n.d. | n.d. | Diplodus vulgaris | erytrocyte | [26] | |
| 1770 | n.d. | n.d. | Salmo gairdnerii | erytrocyte | [26] | |
| 2450 | n.d. | n.d. | Barbus barbus | erytrocyte | [26] | |
| 3190 | n.d. | n.d. | Diplodus vulgaris | erytrocyte | [26] | |
| stimul. | n.d. | n.d. | Capra hircus | erytrocytes | [27] | |
| Parathion | 102 | n.d. | n.d. | A. gueldenstaedti | erythrocytes | [28] |
| Dimetoate | 520 | n.d. | n.d. | Barbus grypus | gills | [29] |
| Organophosphonate | ||||||
| Glyphosate isopropylamine | 0.150 | 0.319 ± 0.067 | non competitive | Ovis aries | stomach | [25] |
| 15.2 | n.d. | n.d. | Homo sapiens (CA I) | erytrocyte | [30] | |
| 62.8 | n.d. | n.d. | Homo sapiens (CA II) | erytrocyte | [30] | |
| Carbamate pesticides | ||||||
| Propoxur | 0.420 | n.d. | Oncorynchus mykiss | gills | [22] | |
| 0.032 | n.d. | n.d. | Apis mellifera | whole animal | [31] | |
| Carbaryl | 0.003 | n.d. | n.d. | Apis mellifera | whole animal | [31] |
| 0.100 | n.d. | n.d. | Bos taurus | erytrocytes | [32] | |
| 398 | n.d. | n.d. | A. gueldenstaedti | erythrocytes | [28] | |
| Carbofuran | 0.009 | n.d. | n.d. | Apis mellifera | whole animal | [31] |
| Mancozeb | 0.368 | n.d. | n.d. | Oncorhynchus mykiss | muscle | [33] |
| 0.505 | n.d. | n.d. | Oncorhynchus mykiss | brain | [33] | |
| 0.151 | n.d. | n.d. | Oncorhynchus mykiss | liver | [33] | |
| 0.306 | n.d. | n.d. | Oncorhynchus mykiss | kidney | [33] | |
| Propamoarb | 90.4 | n.d. | n.d. | Homo sapiens (CA I) | erytrocyte | [30] |
| 62.0 | n.d. | n.d. | Homo sapiens (CA II) | erytrocyte | [30] | |
| Maneb (dithiocarbam.) | 18.0 | n.d. | n.d. | A. gueldenstaedti | erythrocytes | [28] |
| Propineb (dithiocarbam.) | 16.0 | n.d. | n.d. | A. gueldenstaedti | erythrocytes | [28] |
| 0.0094 | 0.0098 ± 0.0048 | uncompetitive | Trachurus trachrus | muscle | [34] | |
| 0.0084 | 0.0111 ± 0.0050 | uncompetitive | Trachurus trachrus | gills | [35] | |
| Thiram (dithiocarbam.) | 0.0058 | 0.0057 ± 0.0023 | uncompetitive | Trachurus trachrus | muscle | [34] |
| 0.0032 | 0.0043 ± 0.0020 | uncompetitive | Trachurus trachrus | gills | [35] | |
| Pyrethroid pesticides | ||||||
| Deltamethrin | 0.137 | n.d. | n.d. | Oncorynchus mykiss | gills | [22] |
| 0.097 | n.d. | n.d. | Oncorynchus mykiss | liver | [33] | |
| 0.237 | n.d. | n.d. | Oncorynchus mykiss | muscle | [33] | |
| 0.161 | n.d. | n.d. | Oncorynchus mykiss | kidney | [33] | |
| 0.160 | n.d. | n.d. | Oncorynchus mykiss | brain | [33] | |
| 14.8 | n.d. | n.d. | Oncorynchus mykiss | erytrocytes | [23] | |
| 470 | n.d. | n.d. | Cyprinus carpio | erytrocytes | [23] | |
| 0.0085 | 0.0076 ± 0.0011 | uncompetitive | Trachurus trachrus | muscle | [34] | |
| 0.012 | 0.0011 ± 0.0031 | uncompetitive | Trachurus trachrus | gills | [35] | |
| Cypermethrin | 0.460 | n.d. | n.d. | Oncorynchus mykiss | gills | [22] |
| 1.248 | 0.832 | n.d. | Dicentrarchus labrax | liver | [24] | |
| 28.440 | 16.17 | n.d. | Bos taurus | erythrocytes | [24] | |
| 0.256 | n.d. | n.d. | Oncorynchus mykiss | liver | [33] | |
| 0.700 | n.d. | n.d. | Oncorynchus mykiss | muscle | [33] | |
| 0.220 | n.d. | n.d. | Oncorynchus mykiss | kidney | [33] | |
| 0.491 | n.d. | n.d. | Oncorynchus mykiss | brain | [33] | |
| Cyhalothrin | 1.895 | 1.074 | n.d. | Dicentrarchus labrax | liver | [24] |
| 2.336 | 1.863 | n.d. | Bos taurus | erythrocytes | [24] | |
| 605 | n.d. | n.d. | Oncorynchus mykiss | erythrocytes | [23] | |
| 686 | n.d. | n.d. | Cyprinus carpio | erythrocytes | [23] | |
| Dinitrophenol pesticides | ||||||
| Dinocap | 0.102 | n.d. | n.d. | Oncorhynchus mykiss | liver | [33] |
| 0.263 | n.d. | n.d. | Oncorhynchus mykiss | muscle | [33] | |
| 0.199 | n.d. | n.d. | Oncorhynchus mykiss | kidney | [33] | |
| 0.190 | n.d. | n.d. | Oncorhynchus mykiss | brain | [33] | |
| 2,4-Dichlorophenol | 240 | n.d. | n.d. | A. gueldenstaedti | erythrocytes | [28] |
| Triazine and tetrazine pesticides | ||||||
| Simazine | 0.0273 | n.d. | n.d. | Apis mellifera | whole animal | [31] |
| Atrazine | 0.0165 | n.d. | n.d. | Apis mellifera | whole animal | [31] |
| Clofentezine | 0.0038 | 0.0023 ± 0.0002 | competitive | Trachurus trachrus | muscle | [34] |
| 0.0035 | 0.0053 ± 0.0022 * | competitive | Trachurus trachrus | gills | [35] | |
| Pyrimidine pesticides | ||||||
| NuarimolTM | 352 | n.d. | n.d. | Capra hircus | erytrocyte | [27] |
| 380 | n.d. | n.d. | Cyprinus carpio | erytrocyte | [26] | |
| 200 | n.d. | n.d. | Scorpaena porcus | erytrocyte | [26] | |
| 280 | n.d. | n.d. | Barbus barbus | erytrocyte | [26] | |
| 230 | n.d. | n.d. | Salmo gairdnerii | erytrocyte | [26] | |
| 380 | n.d. | n.d. | Diplodus vulgaris | erytrocyte | [26] | |
| FenarimolTM | 924 | n.d. | Capra hircus | erytrocyte | [27] | |
| 550 | n.d. | n.d. | Cyprinus carpio | erytrocyte | [26] | |
| 180 | n.d. | n.d. | Scorpaena porcus | erytrocyte | [26] | |
| 590 | n.d. | n.d. | Barbus barbus | erytrocyte | [26] | |
| 510 | n.d. | n.d. | Salmo gairdnerii | erytrocyte | [26] | |
| 370 | n.d. | n.d. | Diplodus vulgaris | erytrocyte | [26] | |
| Triazole pesticides | ||||||
| Tebuconazole | 0.003 | n.d. | Apis mellifera | wholeanimal | [31] | |
| Phenoxy carboxylic acid pesticides | ||||||
| 2,4-D | 2040 | n.d. | n.d. | Capra hircus | erytrocyte | [27] |
| 61.7 | n.d. | n.d. | Homo sapiens (CA I) | erytrocyte | [30] | |
| 66.0 | n.d. | n.d. | Homo sapiens (CA II) | erytrocyte | [30] | |
| 650 | n.d. | n.d. | Scorpaena porcus | erytrocyte | [26] | |
| 2720 | n.d. | n.d. | Cyprinus carpio | erytrocyte | [26] | |
| 1730 | n.d. | n.d. | Barbus barbus | erytrocyte | [26] | |
| 1260 | n.d. | n.d. | Salmo gairdnerii | erytrocyte | [26] | |
| 2670 | n.d. | n.d. | Diplodus vulgaris | erytrocyte | [26] | |
| Spinosoid pesticides | ||||||
| Spinosad | 410 | n.d. | n.d. | Barbus grypus | gills | [29] |
| Imidazolinone herbicides | ||||||
| Imazethapyr | 93.0 | n.d. | n.d. | Homo sapiens (CA I) | erytrocyte | [30] |
| 46.3 | n.d. | n.d. | Homo sapiens (CA II) | erytrocyte | [30] | |
| Strobilurin fungicides | ||||||
| Azoxystrobin | 0.0301 | 0.0307 ± 0.0100 * | competitive | Trachurus trachrus | muscle | [34] |
| 0.0309 | 0.0139 ± 0.0032 | competitive | Trachurus trachrus | gills | [35] | |
| Benzimidazole fungicides | ||||||
| Thiophanate | 0.0705 | 0.0898 ± 0.0322 | uncompetitive | Trachurus trachrus | muscle | [34] |
| 0.0367 | 0.0484 ± 0.0140 | uncompetitive | Trachurus trachrus | gills | [35] | |
| Avermectin pesticides | ||||||
| Abamectin | 0.0144 | 0.0097 ± 0.0019 ** | n.d. | Bos taurus | erytrocytes | [36] |
| Doramectin | 0.0146 | 0.0174 ± 0.0048 ** | n.d. | Bos taurus | erytrocytes | [36] |
| Emamectin | 0.0187 | 0.0020 ± 0.0095 ** | n.d. | Bos taurus | erytrocytes | [36] |
| Eprinomectin | 0.0146 | 0.0134 ± 0.0025 ** | n.d. | Bos taurus | erytrocytes | [36] |
| Ivermectin | 0.0145 | 0.0164 ± 0.0053 ** | n.d. | Bos taurus | erytrocytes | [36] |
| Moxidectin | 0.0208 | 0.0177 ± 0.0037 ** | n.d. | Bos taurus | erytrocytes | [36] |
| Dinitroaniline herbicides | ||||||
| Oryzalin | n.d. | 0.029 | n.d. | Homo sapiens (CA I) | reconbinant | [37] |
| Oryzalin | n.d. | 0.008 | n.d. | Homo sapiens (CA II) | reconbinant | [37] |
| Oryzalin | n.d. | 0.195 | n.d. | Homo sapiens (CAIV) | reconbinant | [37] |
| Oryzalin | n.d. | 0.002 | n.d. | Homo sapiens (CA XIV) | reconbinant | [37] |
| Pesticides | Species | Tissue | Concentration Tested | Duration of Exposure | Effect Observed on CA | Ref |
|---|---|---|---|---|---|---|
| Organophosphate pesticides | ||||||
| Parathion | Oreochromis hornorum | Gills, mesonephron | 8.3, 10.4, 13.9, 20.8 µg/L | 72 h | Dose- and salinity-dependent induction/inhibition | [44] |
| Chlorpyrifos | Oncorhynchus mykiss | Gills, liver | 2.25, 4.5, 6.75 μg/L | 24, 48, 72, 96 h | Dose- and time-dependent inhibition | [45] |
| Organophosphonate pesticides | ||||||
| Glyphosate | Daino rerio | Whole embrios | 1, 5, 10 and 100 mg/L | 96h | Dose- and time-dependent inhibition | [46] |
| Pyrethroid pesticides | ||||||
| Deltamethrin | Onchorynchus mykiss | Gills | 0.25 μg/L, 1 μg/L, and 2.5 μg/L | 6, 12, 24, 48,72 h | Dose- and time-dependent inhibition | [22] |
| Onchorynchus mykiss | Muscle, liver, kidney | 0.25 μg/L, 1 μg/L, and 1 μg/L | 6, 12, 24, 48 h | Dose- and time-dependent inhibition | [33] | |
| Triazine pesticides | ||||||
| Atrazine | Prochilodus lineatus | Gills | 2, 10, 25 μg/l | 48 h and 14 days | Inhibition after 14 days | [47] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lionetto, M.G.; Caricato, R.; Giordano, M.E. Carbonic Anhydrase Sensitivity to Pesticides: Perspectives for Biomarker Development. Int. J. Mol. Sci. 2020, 21, 3562. https://doi.org/10.3390/ijms21103562
Lionetto MG, Caricato R, Giordano ME. Carbonic Anhydrase Sensitivity to Pesticides: Perspectives for Biomarker Development. International Journal of Molecular Sciences. 2020; 21(10):3562. https://doi.org/10.3390/ijms21103562
Chicago/Turabian StyleLionetto, Maria Giulia, Roberto Caricato, and Maria Elena Giordano. 2020. "Carbonic Anhydrase Sensitivity to Pesticides: Perspectives for Biomarker Development" International Journal of Molecular Sciences 21, no. 10: 3562. https://doi.org/10.3390/ijms21103562
APA StyleLionetto, M. G., Caricato, R., & Giordano, M. E. (2020). Carbonic Anhydrase Sensitivity to Pesticides: Perspectives for Biomarker Development. International Journal of Molecular Sciences, 21(10), 3562. https://doi.org/10.3390/ijms21103562