Role of microRNAs in Hemophilia and Thrombosis in Humans

Abstract
:1. Introduction
2. Bleeding Disorders and miRNAs
2.1. Hemophilia
2.1.1. Hemophilia A
2.1.2. Hemophilia B
2.1.3. Hemophilia C
2.2. Von Willebrand Disease
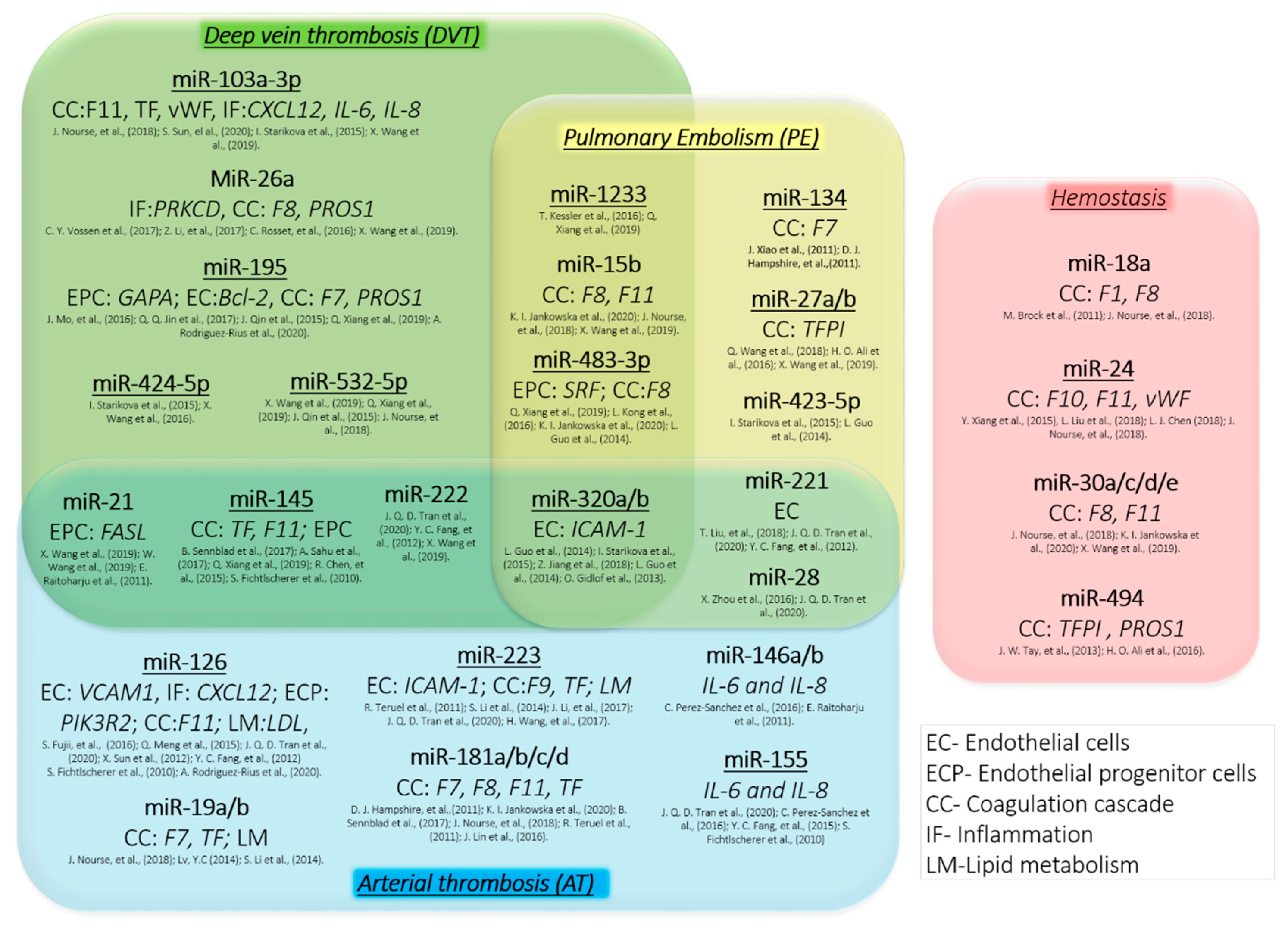
3. Thrombosis and miRNA
3.1. Venous Thromboembolism
3.1.1. Deep Vein Thrombosis
3.1.2. Pulmonary Embolism (PE)
3.2. Arterial Thrombosis
3.3. Coagulation Cascade and Thrombosis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Akt | Protein kinase B |
| APC | Activated protein C |
| APC-PCI | Plasma concentrations of activated protein C-inhibitor of protein C |
| APE | Acute pulmonary embolism |
| ATP | Adenosine triphosphate |
| Bcl-2 | B-cell lymphoma 2 |
| CAD | Coronary artery disease |
| CCL | Chemokine C-C motif ligand |
| CTEPH | Chronic thromboembolic pulmonary hypertension |
| CXCL12 | Chemokine C-X-C motif ligand 12 |
| Del/Ins | Deletion-insertion |
| DVT | Venous thrombosis that includes both deep vein thrombosis |
| EC | Endothelial cells |
| EPC | Endothelial progenitor cells |
| ET-1 | Endothelin-1 |
| F11 | Coagulation factor FXI |
| F2 | Coagulation factor II gene |
| F7 | Coagulation factor VII gene |
| F8 | Coagulation factor VIII gene |
| F9 | Coagulation factor IX gene |
| FasL | Fas ligant (FasL or CD95L or CD178) is a type-II transmembrane protein that belongs to the tumor necrosis factor (TNF) family |
| FASL | Gene encoded Fas ligand |
| FGA | Fibrinogen alpha gene |
| FGB | Fibrinogen beta gene |
| FGG | Fibrinogen gamma gene |
| FI | Coagulation factor I, fibrinogen |
| FII | Prothrombin, coagulation factor II |
| FIX | Coagulation factor IX |
| FVII | Coagulation factor VII |
| FVIII | Coagulation factor VIII |
| FX | Coagulation factor X |
| FXI | Coagulation factor XI |
| GABA | Gamma-aminobutyric acid receptors |
| GABARAPL1 | GABA Type A receptor associated protein like 1 |
| HA | Hemophilia A, coagulation FVIII deficiency |
| HB | Hemophilia B, coagulation FIX deficiency |
| HC | Hemophilia C, coagulation FXI deficiency |
| HepG2 | Liver hepatocellular cells |
| Huh-7 | Human liver cell line |
| HUVEC | Human umbilical vein endothelial cells |
| ICAM-1 | Intercellular adhesion molecule 1 |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| KLKB1 | gene encoded plasma kallikrein |
| LDL | Low-density lipoprotein |
| MAPK | Mitogen-activated protein kinase |
| MCF7 | Breast cancer cell line acronym of Michigan Cancer Foundation-7 |
| miRNA | MicroRNAs |
| MMP2 | Proteins of the matrix metalloproteinase |
| mRNA | Messenger RNA |
| NF-κB | Nuclear factor-κB |
| NMD | Nonsense-mediated mRNA decay |
| OS | Osteosarcoma |
| PBMC | Peripheral blood mononuclear cells |
| PE | Pulmonary embolism |
| PI3K | Phosphoinositide 3-kinases |
| PIK3R2 | Phosphoinositide-3-kinase regulatory subunit 2 which affected PI3K/Akt |
| PLG | Plasminogen, fibrinolytic protease |
| PROS | Protein S gene |
| PROZ | protein Z, vitamin K dependent plasma glycoprotein gene |
| PS | Protein S |
| PTC | Premature termination codon |
| PTEN | Phosphatase and tensin homolog |
| PTS | Post-thrombotic syndrome |
| SERPINA | Serpin family A member 1 gene |
| SERPINC1 | Serpin family C member 1 gene |
| SERPIND1 | SERPIND1 gene encoded heparin cofactor II (HCII) |
| SRC | Proto-oncogene tyrosine-protein kinase Src |
| SRCIN1 | SRC kinase signaling inhibitor 1 |
| SRF | Serum response factor |
| TGFBR1 | Transforming growth factor beta receptor 1 |
| TF | Tissue factor, coagulation factor III |
| TFPI | Tissue factor pathway inhibitor |
| TIC | Trauma-induced coagulopathy patients |
| TNF-α | Tumor necrosis factor alpha |
| UTR | Untranslated regions |
| VCAM-1 | Vascular cell adhesion molecule-1 |
| VWF | Von Willebrand factor |
| VWD | Von Willebrand disease |
References
- Peter, M.E. Targeting of mRNAs by multiple miRNAs: The next step. Oncogene 2010, 29, 2161–2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Michlewski, G.; Caceres, J.F. Post-transcriptional control of miRNA biogenesis. RNA 2019, 25, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, G.M.; Haas, M.A.; Pocock, R. MicroRNAs: Not “fine-tuners” but key regulators of neuronal development and function. Front. Neurol. 2015, 6, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Qin, Y.W.; Brewer, G.; Jing, Q. MicroRNA degradation and turnover: Regulating the regulators. Wiley Interdiscip. Rev. RNA 2012, 3, 593–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schratt, G. Fine-tuning neural gene expression with microRNAs. Curr. Opin. Neurobiol. 2009, 19, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Sevignani, C.; Calin, G.A.; Siracusa, L.D.; Croce, C.M. Mammalian microRNAs: A small world for fine-tuning gene expression. Mamm. Genome 2006, 17, 189–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowska, K.I.; McGill, J.; Pezeshkpoor, B.; Oldenburg, J.; Atreya, C.D.; Sauna, Z.E. Clinical manifestation of hemophilia A in the absence of mutations in the F8 gene that encodes FVIII: Role of microRNAs. Transfusion 2020, 60, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Sarachana, T.; Dahiya, N.; Simhadri, V.L.; Pandey, G.S.; Saini, S.; Guelcher, C.; Guerrera, M.F.; Kimchi-Sarfaty, C.; Sauna, Z.E.; Atreya, C.D. Small ncRNA expression-profiling of blood from hemophilia A patients identifies miR-1246 as a potential regulator of Factor 8 gene. PLoS ONE 2015, 10, e0132433. [Google Scholar] [CrossRef] [Green Version]
- Jerjes-Sanchez, C. Venous and arterial thrombosis: A continuous spectrum of the same disease? Eur. Heart J. 2005, 26, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Kamphuisen, P.W.; Eikenboom, J.C.; Bertina, R.M. Elevated factor VIII levels and the risk of thrombosis. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 731–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, M.; Trenkmann, M.; Gay, R.E.; Gay, S.; Speich, R.; Huber, L.C. MicroRNA-18a enhances the interleukin-6-mediated production of the acute-phase proteins fibrinogen and haptoglobin in human hepatocytes. J. Biol. Chem. 2011, 286, 40142–40150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.J.; Deng, J.; Wang, J.Y.; Zhang, P.J.; Xin, Z.; Xiao, K.; Feng, D.; Jia, Y.H.; Liu, Y.N.; Xie, L.X. Serum miR-122 levels are related to coagulation disorders in sepsis patients. Clin. Chem. Lab. Med. 2014, 52, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Fort, A.; Borel, C.; Migliavacca, E.; Antonarakis, S.E.; Fish, R.J.; Neerman-Arbez, M. Regulation of fibrinogen production by microRNAs. Blood 2010, 116, 2608–2615. [Google Scholar] [CrossRef] [Green Version]
- Nourse, J.; Braun, J.; Lackner, K.; Huttelmaier, S.; Danckwardt, S. Large-scale identification of functional microRNA targeting reveals cooperative regulation of the hemostatic system. J. Thromb. Haemost. 2018, 16, 2233–2245. [Google Scholar] [CrossRef] [Green Version]
- Vossen, C.Y.; Van Hylckama Vlieg, A.; Teruel-Montoya, R.; Salloum-Asfar, S.; De Haan, H.; Corral, J.; Reitsma, P.; Koeleman, B.P.C.; Martinez, C. Identification of coagulation gene 3′UTR variants that are potentially regulated by microRNAs. Br. J. Haematol. 2017, 177, 782–790. [Google Scholar] [CrossRef] [Green Version]
- Carter, A.M.; Catto, A.J.; Kohler, H.P.; Ariens, R.A.; Stickland, M.H.; Grant, P.J. alpha-fibrinogen Thr312Ala polymorphism and venous thromboembolism. Blood 2000, 96, 1177–1179. [Google Scholar] [CrossRef]
- Remijn, J.A.; Van Wijk, R.; De Groot, P.G.; Van Solinge, W.W. Nature of the fibrinogen Aalpha gene TaqI polymorphism. Thromb. Haemost. 2001, 86, 935–936. [Google Scholar]
- Hampshire, D.J.; Goodeve, A.C. The international society on thrombosis and haematosis von Willebrand disease database: An update. Semin. Thromb. Hemost. 2011, 37, 470–479. [Google Scholar] [CrossRef]
- Rodriguez-Rius, A.; Lopez, S.; Martinez-Perez, A.; Souto, J.C.; Soria, J.M. Identification of a plasma MicroRNA profile associated with venous thrombosis. Arterioscler. Thromb. Vasc. Biol. 2020, 40. [Google Scholar] [CrossRef] [Green Version]
- Rosset, C.; Vieira, I.A.; Salzano, F.M.; Bandinelli, E. A germline variant affects putative miRNA-binding sites at the F8 3′UTR and acts as a potential haemophilia A phenotype modifier in Southern Brazilian patients. Haemophilia 2016, 22, e327–e329. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chai, B.; Yang, L. MiR-128 and miR-125 regulate expression of coagulation Factor IX gene with nonsense mutation by repressing nonsense-mediated mRNA decay. Biomed. Pharmacother. 2016, 80, 331–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.J.; Yang, L.; Cheng, X.; Xue, Y.K.; Chen, L.B. Overexpression of miR-24 Is Involved in the Formation of Hypocoagulation State after Severe Trauma by Inhibiting the Synthesis of Coagulation Factor X. Dis Markers 2017, 2017, 3649693. [Google Scholar] [CrossRef] [PubMed]
- Sennblad, B.; Basu, S.; Mazur, J.; Suchon, P.; Martinez-Perez, A.; Van Hylckama Vlieg, A.; Truong, V.; Li, Y.; Gadin, J.R.; Tang, W.; et al. Genome-wide association study with additional genetic and post-transcriptional analyses reveals novel regulators of plasma factor XI levels. Hum. Mol. Genet. 2017, 26, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Tay, J.W.; Romeo, G.; Hughes, Q.W.; Baker, R.I. Micro-ribonucleic Acid 494 regulation of protein S expression. J. Thromb. Haemost. 2013, 11, 1547–1555. [Google Scholar] [CrossRef]
- Sun, S.; Chai, S.; Zhang, F.; Lu, L. Overexpressed microRNA-103a-3p inhibits acute lower-extremity deep venous thrombosis via inhibition of CXCL12. IUBMB Life 2020, 72, 492–504. [Google Scholar] [CrossRef]
- Sahu, A.; Jha, P.K.; Prabhakar, A.; Singh, H.D.; Gupta, N.; Chatterjee, T.; Tyagi, T.; Sharma, S.; Kumari, B.; Singh, S.; et al. MicroRNA-145 impedes thrombus formation via targeting tissue factor in venous thrombosis. EBioMedicine 2017, 26, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; He, S.; Sun, X.; Franck, G.; Deng, Y.; Yang, D.; Haemmig, S.; Wara, A.K.; Icli, B.; Li, D.; et al. MicroRNA-181b inhibits thrombin-mediated endothelial activation and arterial thrombosis by targeting caspase recruitment domain family member 10. FASEB J. 2016, 30, 3216–3226. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Ren, J.; Xu, N.; Zhang, J.; Geng, Q.; Cao, C.; Lee, C.; Song, J.; Li, J.; Chen, H. MicroRNA-19b functions as potential anti-thrombotic protector in patients with unstable angina by targeting tissue factor. J. Mol. Cell Cardiol. 2014, 75, 49–57. [Google Scholar] [CrossRef]
- Li, S.; Chen, H.; Ren, J.; Geng, Q.; Song, J.; Lee, C.; Cao, C.; Zhang, J.; Xu, N. MicroRNA-223 inhibits tissue factor expression in vascular endothelial cells. Atherosclerosis 2014, 237, 514–520. [Google Scholar] [CrossRef]
- Ali, H.O.; Arroyo, A.B.; Gonzalez-Conejero, R.; Stavik, B.; Iversen, N.; Sandset, P.M.; Martinez, C.; Skretting, G. The role of microRNA-27a/b and microRNA-494 in estrogen-mediated downregulation of tissue factor pathway inhibitor alpha. J. Thromb. Haemost. 2016, 14, 1226–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Y.; Cheng, J.; Wang, D.; Hu, X.; Xie, Y.; Stitham, J.; Atteya, G.; Du, J.; Tang, W.H.; Lee, S.H.; et al. Hyperglycemia repression of miR-24 coordinately upregulates endothelial cell expression and secretion of von Willebrand factor. Blood 2015, 125, 3377–3387. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Pan, J.; Wang, H.; Ma, Z.; Yin, J.; Yuan, F.; Yuan, Q.; Zhou, L.; Liu, X.; Zhang, Y.; et al. von Willebrand factor rescued by miR-24 inhibition facilitates the proliferation and migration of osteosarcoma cells in vitro. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsen, J.M.; Fletcher, S.N.; Huston, H.; Roberge, S.; Martin, B.K.; Kircher, M.; Josephson, N.C.; Shendure, J.; Ruuska, S.; Koerper, M.A.; et al. Novel approach to genetic analysis and results in 3000 hemophilia patients enrolled in the My Life, Our Future initiative. Blood Adv. 2017, 1, 824–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaraj, S.R.; Pipe, S.W. Not in the genotype: Can unexplained hemophilia A result from “micro(RNA) management”? Transfusion. 2020, 60, 227–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezeshkpoor, B.; Berkemeier, A.C.; Czogalla, K.J.; Oldenburg, J.; El-Maarri, O. Evidence of pathogenicity of a mutation in 3′ untranslated region causing mild haemophilia A. Haemophilia 2016, 22, 598–603. [Google Scholar] [CrossRef]
- Duga, S.; Salomon, O. Congenital factor XI deficiency: An update. Semin. Thromb. Hemost. 2013, 39, 621–631. [Google Scholar] [CrossRef]
- Wheeler, A.P.; Gailani, D. Why factor XI deficiency is a clinical concern. Expert Rev. Hematol. 2016, 9, 629–637. [Google Scholar] [CrossRef]
- Salloum-Asfar, S.; Teruel-Montoya, R.; Arroyo, A.B.; Garcia-Barbera, N.; Chaudhry, A.; Schuetz, E.; Luengo-Gil, G.; Vicente, V.; Gonzalez-Conejero, R.; Martinez, C. Regulation of coagulation factor XI expression by microRNAs in the human liver. PLoS ONE 2014, 9, e111713. [Google Scholar] [CrossRef]
- Mackman, N. Triggers, targets and treatments for thrombosis. Nature 2008, 451, 914–918. [Google Scholar] [CrossRef]
- Meng, Q.; Wang, W.; Yu, X.; Li, W.; Kong, L.; Qian, A.; Li, C.; Li, X. Upregulation of MicroRNA-126 contributes to endothelial progenitor cell function in deep vein thrombosis via its target PIK3R2. J. Cell. Biochem. 2015, 116, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sundquist, K.; Elf, J.L.; Strandberg, K.; Svensson, P.J.; Hedelius, A.; Palmer, K.; Memon, A.A.; Sundquist, J.; Zoller, B. Diagnostic potential of plasma microRNA signatures in patients with deep-vein thrombosis. Thromb. Haemost. 2016, 116, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhu, X.; Du, X.; Xu, A.; Yuan, X.; Zhan, Y.; Liu, M.; Wang, S. MiR-150 promotes angiogensis and proliferation of endothelial progenitor cells in deep venous thrombosis by targeting SRCIN1. Microvasc. Res. 2019, 123, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.Q.; Sun, J.H.; Du, Q.X.; Lu, X.J.; Zhu, X.Y.; Fan, H.L.; Holscher, C.; Wang, Y.Y. Integrating microRNA and messenger RNA expression profiles in a rat model of deep vein thrombosis. Int. J. Mol. Med. 2017, 40, 1019–1028. [Google Scholar] [CrossRef]
- Mo, J.; Zhang, D.; Yang, R. MicroRNA-195 regulates proliferation, migration, angiogenesis and autophagy of endothelial progenitor cells by targeting GABARAPL1. Biosci. Rep. 2016, 36. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.; Liang, H.; Shi, D.; Dai, J.; Xu, Z.; Chen, D.; Chen, X.; Jiang, Q. A panel of microRNAs as a new biomarkers for the detection of deep vein thrombosis. J. Thromb. Thrombolysis 2015, 39, 215–221. [Google Scholar] [CrossRef]
- Sun, L.L.; Xiao, L.; Du, X.L.; Hong, L.; Li, C.L.; Jiao, J.; Li, W.D.; Li, X.Q. MiR-205 promotes endothelial progenitor cell angiogenesis and deep vein thrombosis recanalization and resolution by targeting PTEN to regulate Akt/autophagy pathway and MMP2 expression. J. Cell. Mol. Med. 2019, 23, 8493–8504. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Hong, L.; Sun, L.; Sang, H.; Qian, A.; Li, W.; Zhuang, H.; Liang, H.; Song, D.; Li, C.; et al. miR-21 induces endothelial progenitor cells proliferation and angiogenesis via targeting FASLG and is a potential prognostic marker in deep venous thrombosis. J. Transl. Med. 2019, 17, 270. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Ni, J. Role of microRNA-26a in the diagnosis of lower extremity deep vein thrombosis in patients with bone trauma. Exp. Ther. Med. 2017, 14, 5069–5074. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Ma, J.; Wang, Q.; Wu, F.; Ping, J.; Ming, L. Combination of circulating miRNA-320a/b and D-dimer improves diagnostic accuracy in deep vein thrombosis patients. Med. Sci. Monit. 2018, 24, 2031–2037. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Z.; Wei, R.; Miao, X.; Sun, S.; Liang, G.; Chu, C.; Zhao, L.; Zhu, X.; Guo, Q.; et al. IL (Interleukin)-6 contributes to deep vein thrombosis and is negatively regulated by miR-338-5p. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Hu, N.; Du, X.; Wang, W.; Chen, H.; Li, W.; Wei, S.; Zhuang, H.; Li, X.; Li, C. Upregulation of miR-483-3p contributes to endothelial progenitor cells dysfunction in deep vein thrombosis patients via SRF. J. Transl. Med. 2016, 14, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Yang, Y.; Liu, J.; Wang, L.; Li, J.; Wang, Y.; Liu, Y.; Gu, S.; Gan, H.; Cai, J.; et al. Differentially expressed plasma microRNAs and the potential regulatory function of Let-7b in chronic thromboembolic pulmonary hypertension. PLoS ONE 2014, 9, e101055. [Google Scholar] [CrossRef] [PubMed]
- Kessler, T.; Erdmann, J.; Vilne, B.; Bruse, P.; Kurowski, V.; Diemert, P.; Schunkert, H.; Sager, H.B. Serum microRNA-1233 is a specific biomarker for diagnosing acute pulmonary embolism. J. Transl. Med. 2016, 14, 120. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Jing, Z.C.; Ellinor, P.T.; Liang, D.; Zhang, H.; Liu, Y.; Chen, X.; Pan, L.; Lyon, R.; Liu, Y.; et al. MicroRNA-134 as a potential plasma biomarker for the diagnosis of acute pulmonary embolism. J. Transl. Med. 2011, 9, 159. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Kang, J.; Liu, F. Plasma levels of microRNA-221 (miR-221) are increased in patients with acute pulmonary embolism. Med. Sci. Monit. 2018, 24, 8621–8626. [Google Scholar] [CrossRef]
- Wang, Q.; Ma, J.; Jiang, Z.; Wu, F.; Ping, J.; Ming, L. Diagnostic value of circulating microRNA-27a/b in patients with acute pulmonary embolism. Int. Angiol. 2018, 37, 19–25. [Google Scholar] [CrossRef]
- Zhou, X.; Wen, W.; Shan, X.; Qian, J.; Li, H.; Jiang, T.; Wang, W.; Cheng, W.; Wang, F.; Qi, L.; et al. MiR-28-3p as a potential plasma marker in diagnosis of pulmonary embolism. Thromb. Res. 2016, 138, 91–95. [Google Scholar] [CrossRef]
- Starikova, I.; Jamaly, S.; Sorrentino, A.; Blondal, T.; Latysheva, N.; Sovershaev, M.; Hansen, J.B. Differential expression of plasma miRNAs in patients with unprovoked venous thromboembolism and healthy control individuals. Thromb. Res. 2015, 136, 566–572. [Google Scholar] [CrossRef]
- Wang, X.; Sundquist, K.; Svensson, P.J.; Rastkhani, H.; Palmer, K.; Memon, A.A.; Sundquist, J.; Zoller, B. Association of recurrent venous thromboembolism and circulating microRNAs. Clin. Epigenet. 2019, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Q.; Zhang, H.X.; Wang, Z.; Liu, Z.Y.; Xie, Q.F.; Hu, K.; Zhang, Z.; Mu, G.Y.; Ma, L.Y.; Jiang, J.; et al. The predictive value of circulating microRNAs for venous thromboembolism diagnosis: A systematic review and diagnostic meta-analysis. Thromb. Res. 2019, 181, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Tran, J.Q.D.; Pedersen, O.H.; Larsen, M.L.; Grove, E.L.; Kristensen, S.D.; Hvas, A.M.; Nissen, P.H. Platelet microRNA expression and association with platelet maturity and function in patients with essential thrombocythemia. Platelets 2020, 31, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Perez-Sanchez, C.; Aguirre, M.A.; Ruiz-Limon, P.; Barbarroja, N.; Jimenez-Gomez, Y.; De la Rosa, I.A.; Rodriguez-Ariza, A.; Collantes-Estevez, E.; Segui, P.; Velasco, F.; et al. Atherothrombosis-associated microRNAs in Antiphospholipid syndrome and Systemic Lupus Erythematosus patients. Sci. Rep. 2016, 6, 31375. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Sugiura, T.; Dohi, Y.; Ohte, N. MicroRNA in atherothromobosis: Is it useful as a disease marker? Thromb. J. 2016, 14, 21. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.C.; Yeh, C.H. Role of microRNAs in vascular remodeling. Curr. Mol. Med. 2015, 15, 684–696. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zhang, M.; Sanagawa, A.; Mori, C.; Ito, S.; Iwaki, S.; Satoh, H.; Fujii, S. Circulating microRNA-126 in patients with coronary artery disease: Correlation with LDL cholesterol. Thromb. J. 2012, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Fichtlscherer, S.; De Rosa, S.; Fox, H.; Schwietz, T.; Fischer, A.; Liebetrau, C.; Weber, M.; Hamm, C.W.; Roxe, T.; Muller-Ardogan, M.; et al. Circulating microRNAs in patients with coronary artery disease. Circ. Res. 2010, 107, 677–684. [Google Scholar] [CrossRef] [Green Version]
- Raitoharju, E.; Lyytikainen, L.P.; Levula, M.; Oksala, N.; Mennander, A.; Tarkka, M.; Klopp, N.; Illig, T.; Kahonen, M.; Karhunen, P.J.; et al. miR-21, miR-210, miR-34a, and miR-146a/b are up-regulated in human atherosclerotic plaques in the Tampere Vascular Study. Atherosclerosis 2011, 219, 211–217. [Google Scholar] [CrossRef]
- Li, J.; Tan, M.; Xiang, Q.; Zhou, Z.; Yan, H. Thrombin-activated platelet-derived exosomes regulate endothelial cell expression of ICAM-1 via microRNA-223 during the thrombosis-inflammation response. Thromb. Res. 2017, 154, 96–105. [Google Scholar] [CrossRef]
- Franco, R.F.; Reitsma, P.H. Genetic risk factors of venous thrombosis. Hum. Genet. 2001, 109, 369–384. [Google Scholar] [CrossRef]
- Hembrom, A.A.; Srivastava, S.; Garg, I.; Kumar, B. MicroRNAs in venous thrombo-embolism. Clin. Chim. Acta 2020, 504, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Ma, J.; Wang, Q.; Wu, F.; Ping, J.; Ming, L. Circulating microRNA expression and their target genes in deep vein thrombosis: A systematic review and bioinformatics analysis. Medicine (Baltimore) 2017, 96, e9330. [Google Scholar] [CrossRef] [PubMed]
- Li, W.D.; Li, X.Q. Endothelial progenitor cells accelerate the resolution of deep vein thrombosis. Vascul. Pharmacol. 2016, 83, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Tahamtan, A.; Teymoori-Rad, M.; Nakstad, B.; Salimi, V. Anti-inflammatory MicroRNAs and their potential for inflammatory diseases treatment. Front. Immunol. 2018, 9, 1377. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, A.B.; Salloum-Asfar, S.; Perez-Sanchez, C.; Teruel-Montoya, R.; Navarro, S.; Garcia-Barbera, N.; Luengo-Gil, G.; Roldan, V.; Hansen, J.B.; Lopez-Pedrera, C.; et al. Regulation of TFPIalpha expression by miR-27a/b-3p in human endothelial cells under normal conditions and in response to androgens. Sci. Rep. 2017, 7, 43500. [Google Scholar] [CrossRef] [Green Version]
- Voetsch, B.; Loscalzo, J. Genetic determinants of arterial thrombosis. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 216–229. [Google Scholar] [CrossRef] [Green Version]
- Menghini, R.; Stohr, R.; Federici, M. MicroRNAs in vascular aging and atherosclerosis. Ageing Res. Rev. 2014, 17, 68–78. [Google Scholar] [CrossRef]
- Hosin, A.A.; Prasad, A.; Viiri, L.E.; Davies, A.H.; Shalhoub, J. MicroRNAs in atherosclerosis. J. Vasc. Res. 2014, 51, 338–349. [Google Scholar] [CrossRef]
- Feinberg, M.W.; Moore, K.J. MicroRNA regulation of atherosclerosis. Circ. Res. 2016, 118, 703–720. [Google Scholar] [CrossRef] [Green Version]
- Tomaiuolo, M.; Brass, L.F.; Stalker, T.J. Regulation of platelet activation and coagulation and its role in vascular injury and arterial thrombosis. Interv. Cardiol. Clin. 2017, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pafili, K.; Penlioglou, T.; Mikhailidis, D.P.; Papanas, N. Mean platelet volume and coronary artery disease. Curr. Opin. Cardiol. 2019, 34, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Icli, A.; Aksoy, F.; Turker, Y.; Uysal, B.A.; Alpay, M.F.; Dogan, A.; Nar, G.; Varol, E. Relationship between mean platelet volume and pulmonary embolism in patients with deep vein thrombosis. Heart Lung Circ. 2015, 24, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Warny, M.; Helby, J.; Birgens, H.S.; Bojesen, S.E.; Nordestgaard, B.G. Arterial and venous thrombosis by high platelet count and high hematocrit: 108 521 individuals from the Copenhagen General Population Study. J. Thromb. Haemost. 2019, 17, 1898–1911. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.; Olejnickova, V.; Tkacova, N.; Santulli, G. Mechanistic role of MicroRNAs in coupling lipid metabolism and atherosclerosis. Adv. Exp. Med. Biol. 2015, 887, 79–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazari-Jahantigh, M.; Wei, Y.; Schober, A. The role of microRNAs in arterial remodelling. Thromb. Haemost. 2012, 107, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.C.; Tang, Y.Y.; Peng, J.; Zhao, G.J.; Yang, J.; Yao, F.; Ouyang, X.P.; He, P.P.; Xie, W.; Tan, Y.L.; et al. MicroRNA-19b promotes macrophage cholesterol accumulation and aortic atherosclerosis by targeting ATP-binding cassette transporter A1. Atherosclerosis 2014, 236, 215–226. [Google Scholar] [CrossRef]
- Jansen, F.; Schafer, L.; Wang, H.; Schmitz, T.; Flender, A.; Schueler, R.; Hammerstingl, C.; Nickenig, G.; Sinning, J.M.; Werner, N. Kinetics of circulating MicroRNAs in response to cardiac stress in patients with coronary artery disease. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef]
- Gidlof, O.; Van der Brug, M.; Ohman, J.; Gilje, P.; Olde, B.; Wahlestedt, C.; Erlinge, D. Platelets activated during myocardial infarction release functional miRNA, which can be taken up by endothelial cells and regulate ICAM1 expression. Blood 2013, 121, 3908–3917. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Nakajima, T.; Tanabe, N.; Hinohara, K.; Sakao, S.; Kasahara, Y.; Tatsumi, K.; Inoue, Y.; Kimura, A. Susceptibility to chronic thromboembolic pulmonary hypertension may be conferred by miR-759 via its targeted interaction with polymorphic fibrinogen alpha gene. Hum. Genet. 2010, 128, 443–452. [Google Scholar] [CrossRef]
- Nesbitt, J.E.; Fuller, G.M. Transcription and translation are required for fibrinogen mRNA degradation in hepatocytes. Biochim. Biophys. Acta 1991, 1089, 88–94. [Google Scholar] [CrossRef]
- Teruel, R.; Corral, J.; Perez-Andreu, V.; Martinez-Martinez, I.; Vicente, V.; Martinez, C. Potential role of miRNAs in developmental haemostasis. PLoS ONE 2011, 6, e17648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campello, E.; Spiezia, L.; Radu, C.M.; Bulato, C.; Gavasso, S.; Tormene, D.; Woodhams, B.; Dalla Valle, F.; Simioni, P. Circulating microparticles and the risk of thrombosis in inherited deficiencies of antithrombin, protein C and protein S. Thromb. Haemost. 2016, 115, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, R.D.; Bauer, K.A. Thrombosis in inherited deficiencies of antithrombin, protein C, and protein S. Hum. Pathol. 1987, 18, 253–262. [Google Scholar] [CrossRef]
- Siddiqi, F.A.; Tepler, J.; Fantini, G.A. Acquired protein S and antithrombin III deficiency caused by nephrotic syndrome: An unusual cause of graft thrombosis. J. Vasc. Surg. 1997, 25, 576–580. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Target Gene | miRNA | References |
|---|---|---|
| F1 | miR-18a | [12] |
| miR-211 | [13] | |
| miR-218 | [14] | |
| miR-29a/b/c | [14] | |
| miR-365 | [14] | |
| F1 (FGA) | miR-193b-3p | [15] |
| miR-194-5p | [15] | |
| F1(FGA) * | miR-186 | [16] |
| miR-3133 | [16] | |
| miR-3173 | [16] | |
| miR-434 | [16] | |
| miR-759 | [17,18] | |
| miR-4476 | [16] | |
| F1 (FGB) | miR-409-3p | [14] |
| F1(FGB) * | miR-29b-1-5p | [16] |
| miR-4294 | [16] | |
| miR-627 | [16] | |
| miR-759 | [16] | |
| miR-924 | [16] | |
| F1(FGG_A) | miR-151a-5p | [15] |
| miR-193-5p | [15] | |
| miR-452-5p | [15] | |
| miR-99b-3p | [15] | |
| F7 | miR-134 and | [19] |
| miR-181a | [19] | |
| miR-195-5p | [20] | |
| miR-19a/b-3p | [15] | |
| MiR-885-5p | [20] | |
| F8 | let-7i-5p | [8] |
| miR-1246, | [9] | |
| miR-128-3p | [8] | |
| miR-144-5p | [8] | |
| miR-15b-3p | [8] | |
| miR-181d | [9] | |
| miR-18a-5p | [15] | |
| miR-30c | [8] | |
| miR-30e-3p | [15] | |
| miR-34-5p | [15] | |
| miR-374b | [8] | |
| miR-4521 | [9] | |
| miR-454-3p | [15] | |
| miR-483-3p | [8] | |
| miR-532-5p | [15] | |
| miR-6803-3p, | [8] | |
| miR-7-5p | [15] | |
| miR-874-3p | [15] | |
| miR-1297 | [21] | |
| F8 * | miR-26a-5p | [21] |
| miR-26b-5p | [21] | |
| miR-34a/c | [16] | |
| miR-449a/b | [16] | |
| F9 | miR-128 | [22] |
| miR-125 | [22] | |
| F10 | miR-24 | [23] |
| F11 | miR-103a-3p | [15] |
| miR-1255a | [15] | |
| miR-145 | [24] | |
| miR-148b-3p | [15] | |
| miR-151a-3p | [15] | |
| miR-15b-5p | [15] | |
| miR-181a | [24] | |
| miR-181b-5p | [15] | |
| miR-24-3p | [15] | |
| miR-30a-3p | [15] | |
| miR-30d-3p | [15] | |
| miR-96-5p | [15] | |
| miR-126-3p | [20] | |
| F11 * | miR-137 | [16] |
| miR-1975 | [16] | |
| miR2355 | [16] | |
| miR-4286 | [16] | |
| miR-513a-3p | [16] | |
| miR-544 | [16] | |
| miR-622 | [16] | |
| miR-889 | [16] | |
| miR-93-5p | [16] | |
| PROS1 | miR-195-5p | [20] |
| MiR-494 | [25] | |
| MiR-885-5p | [20] | |
| PROS1 * | miR-26-5p | [16] |
| miR-375 | [16] | |
| TF | miR-103-3p | [26] |
| miR-145 | [27] | |
| miR-181b | [28] | |
| miR-19b | [29] | |
| miR-223 | [30] | |
| TFPI | miR-27a/b | [31] |
| miR-494 | [31] | |
| TFPI * | miR-200a/b | [16] |
| miR-2355 | [16] | |
| miR-429 | [16] | |
| miR-4302 | [16] | |
| miR-605 | [16] | |
| VWF | miR-24 | [32,33] |
| miR-103-3p | [26] | |
| ADAMS13 | MiR-192-5p | [20] |
| miRNA | Target Gene | References | |
|---|---|---|---|
| DVT | miR-103a-3p | CXCL12 | [26] |
| miR-126 | PIK3R2 | [41] | |
| miR-136-5p | [42] | ||
| miR-150 | SRCIN1 | [43] | |
| miR-195 | Bcl-2 | [44] | |
| GAPA | [45] | ||
| [46] | |||
| miR-205 | PTEN | [47] | |
| miR-21 | FASL | [48] | |
| MiR-26a | PRKCD | [49] | |
| miR-320a/b | [50] | ||
| miR-338-5 | [51] | ||
| miR-424-5p | [42] | ||
| miR-483-3p | SRF | [52] | |
| miR-532 | [46] | ||
| miR-582 | [46] | ||
| PE | let-17b | ET-1, TGFBR1 | [53] |
| miR-106b | [53] | ||
| miR-1233 | [54] | ||
| miR-1260 | [53] | ||
| miR129-5p | [53] | ||
| miR-134 | [55] | ||
| miR-140-3p | [53] | ||
| miR-185 | [53] | ||
| miR-1908 | [53] | ||
| miR-22 | [53] | ||
| miR-221 | [56] | ||
| miR-27a/b | [57] | ||
| miR-28 | [58] | ||
| miR-320a/b/c | [53] | ||
| miR-423-5p | [53] | ||
| miR-483-5p | [53] | ||
| miR-486 | [53] | ||
| miR-602 | [53] | ||
| miR-93 | [53] | ||
| miR-933 | [53] | ||
| VTE | miR-103a-3p | [59] | |
| [60] | |||
| miR-106a-5p | [60] | ||
| miR-10b-5p | [59] | ||
| miR-145 | [61] | ||
| miR-15b-5p | [60] | ||
| miR-191-5p | [59] | ||
| miR-195 | [61] | ||
| miR-197-3p | [60] | ||
| miR-199b-3p | [59] | ||
| miR-21-5p | [60] | ||
| miR-222-3p | [60] | ||
| miR-26b-5p | [60] | ||
| miR-27b-3p | [60] | ||
| miR-301a-3p | [59] | ||
| miR-30c-5p | [60] | ||
| miR-320a/b | [59] | ||
| miR-361-5p | [60] | ||
| miR-423-5p | [59] | ||
| miR-424-5p | [59] | ||
| miR-483-3p | [61] | ||
| miR-532 | [61] | ||
| miR-532-5p | [60] | ||
| miR-652-3p | [60] | ||
| AT | miR-10a | [62] | |
| miR-124a | [63] | ||
| miR-125a | [63] | ||
| miR-126 | VCAM-1, CXCL12 | [64] | |
| [65] | |||
| [66] | |||
| [62] | |||
| [67] | |||
| miR-146a | [63] | ||
| [68] | |||
| miR-155 | [65] | ||
| [63] | |||
| [62] | |||
| [67] | |||
| miR-17 | [67] | ||
| miR-21 | [68] | ||
| miR-210 | [68] | ||
| miR-221 | [65] | ||
| [62] | |||
| miR-222 | [65] | ||
| [62] | |||
| miR-223 | ICAM-1 | [69] | |
| [62] | |||
| [13] | |||
| miR-28 | [62] | ||
| miR-320b | ICAM-1 | [27] | |
| miR-34a | [68] | ||
| miR-431 | [62] | ||
| miR-490 | [62] | ||
| miR-9 | [62] | ||
| miR-92a | [68] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankowska, K.I.; Sauna, Z.E.; Atreya, C.D. Role of microRNAs in Hemophilia and Thrombosis in Humans. Int. J. Mol. Sci. 2020, 21, 3598. https://doi.org/10.3390/ijms21103598
Jankowska KI, Sauna ZE, Atreya CD. Role of microRNAs in Hemophilia and Thrombosis in Humans. International Journal of Molecular Sciences. 2020; 21(10):3598. https://doi.org/10.3390/ijms21103598
Chicago/Turabian StyleJankowska, Katarzyna I., Zuben E. Sauna, and Chintamani D. Atreya. 2020. "Role of microRNAs in Hemophilia and Thrombosis in Humans" International Journal of Molecular Sciences 21, no. 10: 3598. https://doi.org/10.3390/ijms21103598
APA StyleJankowska, K. I., Sauna, Z. E., & Atreya, C. D. (2020). Role of microRNAs in Hemophilia and Thrombosis in Humans. International Journal of Molecular Sciences, 21(10), 3598. https://doi.org/10.3390/ijms21103598