Production and Sensing of Butyrate in a Probiotic E. coli Strain


Abstract
:1. Introduction
2. Results
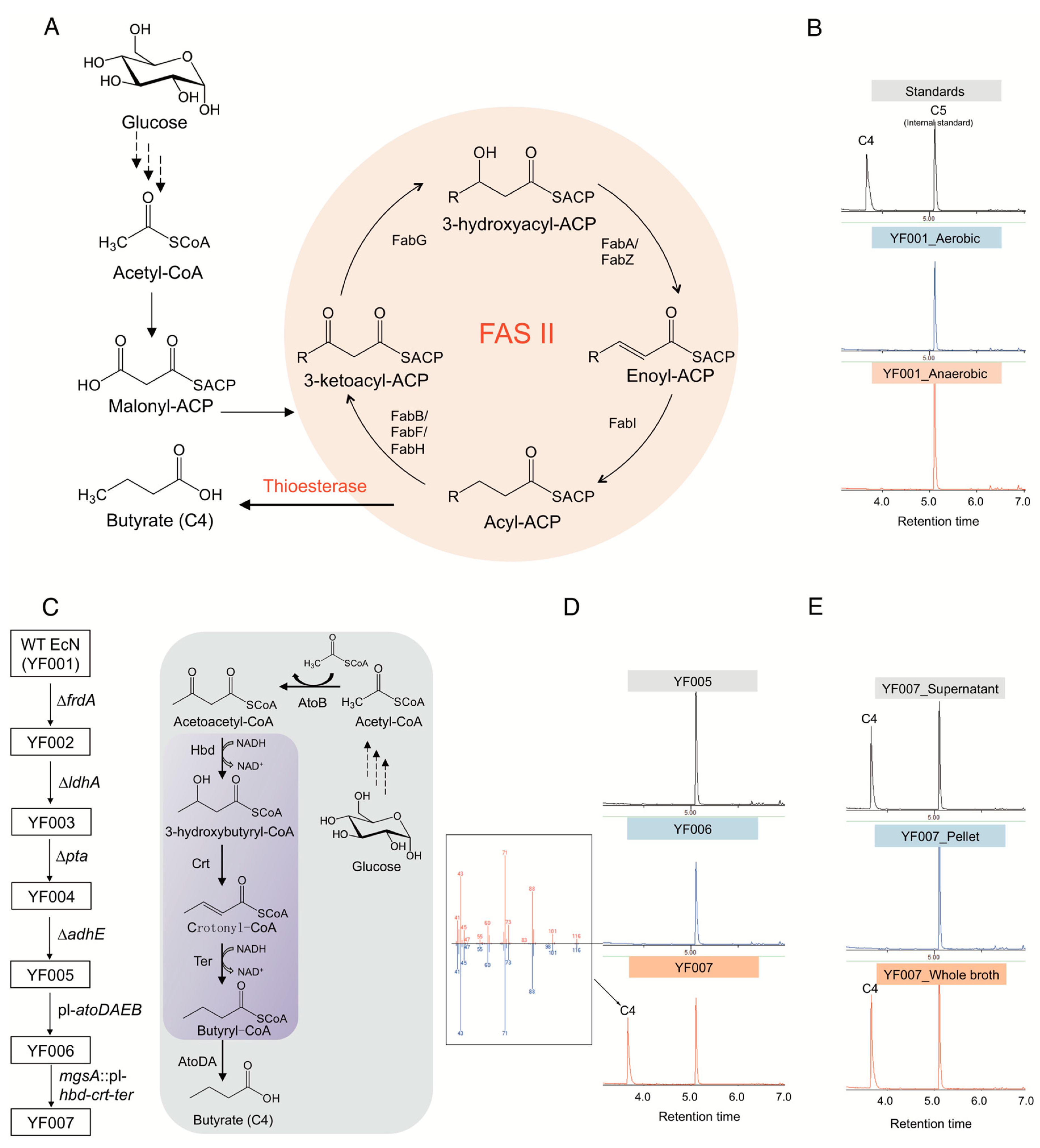
2.1. Construction of a Butyrate Biosynthetic Pathway in E. coli Nissle 1917
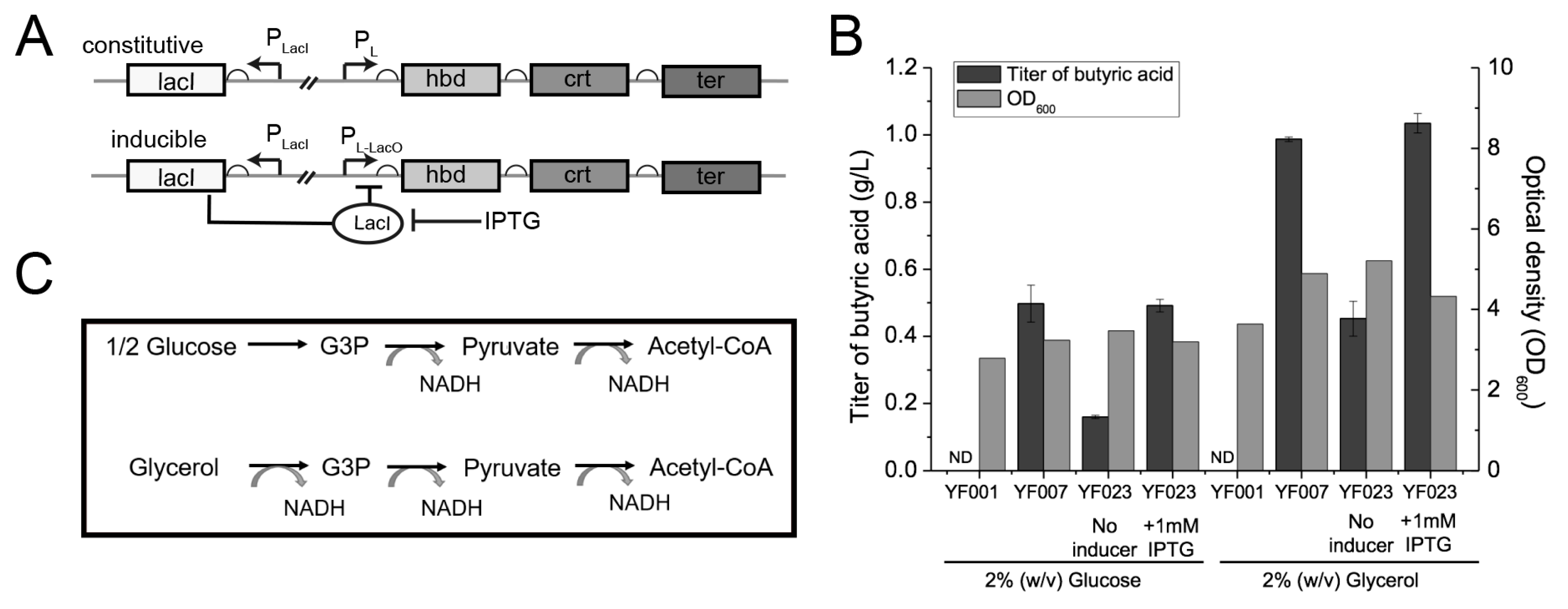
2.2. Effects of Inducible Expression System and Different Carbon Sources on C4 Production
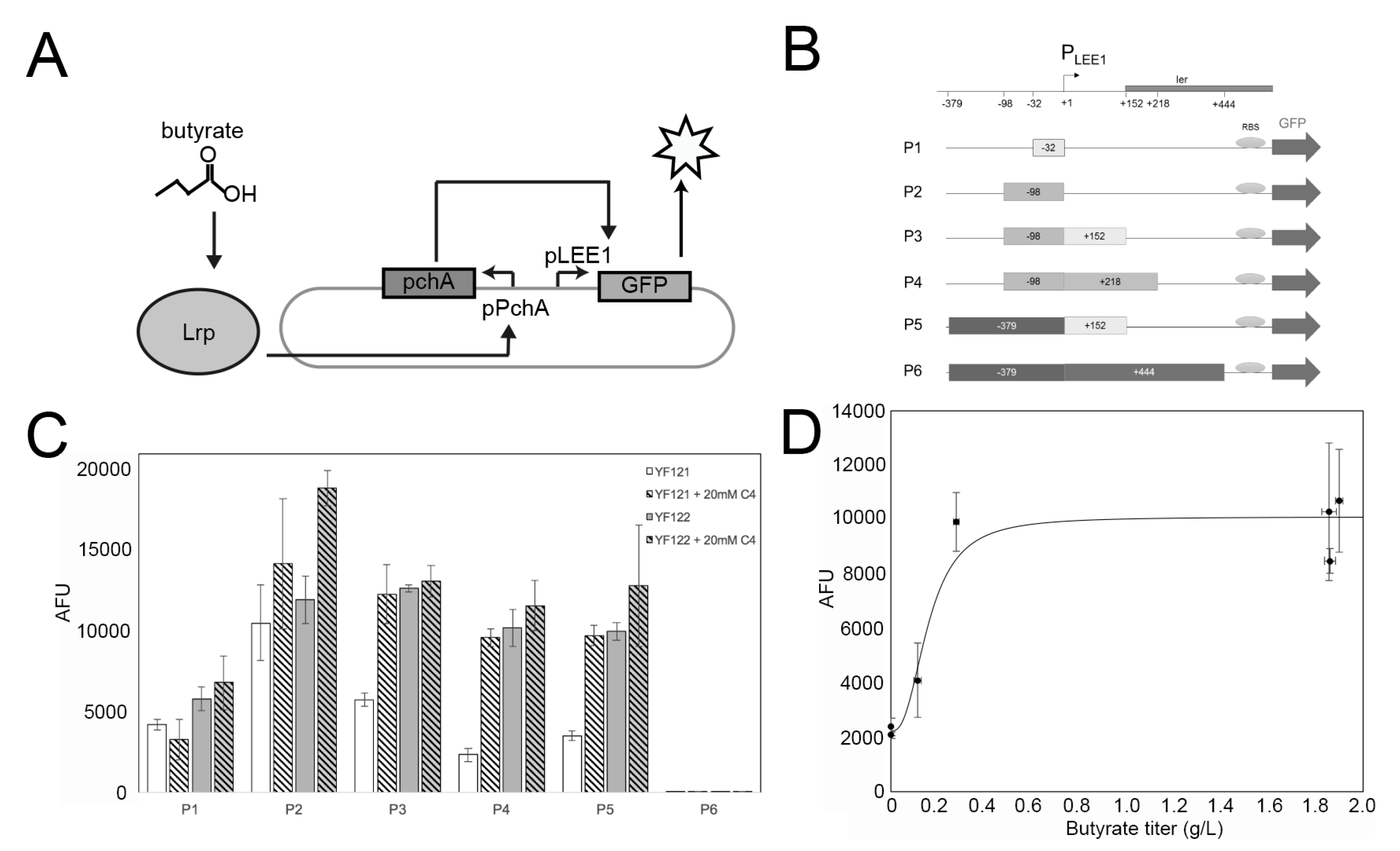
2.3. Construction of High-Throughput Butyrate Sensor System in EcN
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Culture Conditions
4.2. Strain Construction
4.3. Biosensor Plasmid Construction
4.4. Butyrate Production by E. coli Nissle 1917
4.5. Butyrate Extraction and Detection by GC-MS
4.6. Detection of Sensor System of Butyrate
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, M.Y.; Turner, N.D.; Murphy, M.E.; Carroll, R.J.; Chapkin, R.S.; Lupton, J.R. In vivo regulation of colonic cell proliferation, differentiation, apoptosis, and P27Kip1 by dietary fish oil and butyrate in rats. Cancer Prev. Res. 2015, 8, 1076–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O’Connell, T.M.; Bunger, M.K.; Bultman, S.J. The Microbiome and Butyrate Regulate Energy Metabolism and Autophagy in the Mammalian Colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, M.; Hong Chen, M.; Chiang, C.J.; Chao, Y.P. Potential production platform of n-butanol in E. coli. Metab. Eng. 2015, 27, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Bultman, S.J. Butyrate consumption of differentiated colonocytes in the upper crypt promotes homeostatic proliferation of stem and progenitor cells near the crypt base. Transl. Cancer Res. 2016, 5, S526–S528. [Google Scholar] [CrossRef]
- Rivera-Chavez, F.; Zhang, L.F.; Faber, F.; Lopez, C.A.; Byndloss, M.X.; Olsan, E.E.; Xu, G.; Velazquez, E.M.; Lebrilla, C.B.; Winter, S.E.; et al. Depletion of Butyrate-Producing Clostridia from the Gut Microbiota Drives an Aerobic Luminal Expansion of Salmonella. Cell Host Microbe. 2016, 19, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geirnaert, A.; Calatayud, M.; Grootaert, C.; Laukens, D.; Devriese, S.; Smagghe, G.; De Vos, M.; Boon, N.; Van de Wiele, T. Butyrate-producing bacteria supplemented in vitro to Crohn’s disease patient microbiota increased butyrate production and enhanced intestinal epithelial barrier integrity. Sci. Rep. 2017, 7, 11450. [Google Scholar] [CrossRef] [Green Version]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal Clostridia: Leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5. [Google Scholar] [CrossRef] [Green Version]
- Rios-Covian, D.; Gueimonde, M.; Duncan, S.H.; Flint, H.J.; Reyes-Gavilan, C.G.D.L. Enhanced butyrate formation by cross-feeding between Faecalibacterium prausnitzii and Bifidobacterium adolescentis. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Nishida, A.; Fujimoto, T.; Fujii, M.; Shioya, M.; Imaeda, H.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Andoh, A. Reduced Abundance of Butyrate-Producing Bacteria Species in the Fecal Microbial Community in Crohn’s Disease. Digestion 2016, 93, 59–65. [Google Scholar] [CrossRef]
- Breuer, R.I.; Soergel, K.H.; Lashner, B.A.; Christ, M.L.; Hanauer, S.B.; Vanagunas, A.; Harig, J.M.; Keshavarzian, A.; Robinson, M.; Sellin, J.H.; et al. Short chain fatty acid rectal irrigation for left-sided ulcerative colitis: A randomised, placebo controlled trial. Gut 1997, 40, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Sabatino, A.; Morera, R.; Ciccocioppo, R.; Cazzola, P.; Gotti, S.; Tinozzi, F.P.; Tinozzi, S.; Corazza, G.R. Oral butyrate for mildly to moderately active Crohn’s disease. Aliment. Pharm. Ther. 2005, 22, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Vieira, E.L.; Leonel, A.J.; Sad, A.P.; Beltrao, N.R.; Costa, T.F.; Ferreira, T.M.; Gomes-Santos, A.C.; Faria, A.M.; Peluzio, M.C.; Cara, D.C.; et al. Oral administration of sodium butyrate attenuates inflammation and mucosal lesion in experimental acute ulcerative colitis. J. Nutr. Biochem. 2012, 23, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Gaudier, E.; Rival, M.; Buisine, M.-P.; Robineau, I.; Hoebler, C. Butyrate Enemas Upregulate Muc Genes Expression but Decrease Adherent Mucus Thickness in Mice Colon. Physiol. Res. 2009, 58, 111–119. [Google Scholar]
- Van Immerseel, F.; Ducatelle, R.; De Vos, M.; Boon, N.; Van De Wiele, T.; Verbeke, K.; Rutgeerts, P.; Sas, B.; Louis, P.; Flint, H.J. Butyric acid-producing anaerobic bacteria as a novel probiotic treatment approach for inflammatory bowel disease. J. Med. Microbiol. 2010, 59, 141–143. [Google Scholar] [CrossRef] [Green Version]
- Trachsel, J.; Bayles, D.O.; Looft, T.; Levine, U.Y.; Allen, H.K. Function and Phylogeny of Bacterial Butyryl Coenzyme A:Acetate Transferases and Their Diversity in the Proximal Colon of Swine. Appl. Environ. Microbiol. 2016, 82, 6788–6798. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol Hepatol 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Henker, J.; Laass, M.W.; Blokhin, B.M.; Maydannik, V.G.; Bolbot, Y.K.; Elze, M.; Wolff, C.; Schreiner, A.; Schulze, J. Probiotic E. coli Nissle 1917 versus placebo for treating diarrhea of greater than 4 days duration in infants and toddlers. Pediatr. Infect. Dis. J. 2008, 27, 494–499. [Google Scholar] [CrossRef]
- Ukena, S.N.; Singh, A.; Dringenberg, U.; Engelhardt, R.; Seidler, U.; Hansen, W.; Bleich, A.; Bruder, D.; Franzke, A.; Rogler, G.; et al. Probiotic E. coli Nissle 1917 inhibits leaky gut by enhancing mucosal integrity. PLoS ONE 2007, 2, e1308. [Google Scholar] [CrossRef]
- Duan, F.F.; Liu, J.H.; March, J.C. Engineered Commensal Bacteria Reprogram Intestinal Cells Into Glucose-Responsive Insulin-Secreting Cells for the Treatment of Diabetes. Diabetes 2015, 64, 1794–1803. [Google Scholar] [CrossRef] [Green Version]
- Stritzker, J.; Weibel, S.; Hill, P.J.; Oelschlaeger, T.A.; Goebel, W.; Szalay, A.A. Tumor-specific colonization, tissue distribution, and gene induction by probiotic E. coli Nissle 1917 in live mice. Int. J. Med. Microbiol. 2007, 297, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Xia, L.; Zhang, X.; Ding, X.; Yan, F.; Wu, F. E. coli Nissle 1917 targets and restrains mouse B16 melanoma and 4T1 breast tumors through expression of azurin protein. Appl. Environ. Microbiol. 2012, 78, 7603–7610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeidi, N.; Wong, C.K.; Lo, T.M.; Nguyen, H.X.; Ling, H.; Leong, S.S.; Poh, C.L.; Chang, M.W. Engineering microbes to sense and eradicate Pseudomonas aeruginosa, a human pathogen. Mol. Syst. Biol. 2011, 7, 521. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Guo, L.; Zhang, Y.; Walzem, R.L.; Pendergast, J.S.; Printz, R.L.; Morris, L.C.; Matafonova, E.; Stien, X.; Kang, L.; et al. Incorporation of therapeutically modified bacteria into gut microbiota inhibits obesity. J. Clin. Invest. 2014, 124, 3391–3406. [Google Scholar] [CrossRef] [PubMed]
- Somabhai, C.A.; Raghuvanshi, R.; Nareshkumar, G. Genetically Engineered E. coli Nissle 1917 Synbiotics Reduce Metabolic Effects Induced by Chronic Consumption of Dietary Fructose. PLoS ONE 2016, 11, e0164860. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Ahn, J.H.; Park, S.H.; Do, K.H.; Kim, J.; Moon, Y. Enhanced wound healing by recombinant E. coli Nissle 1917 via human epidermal growth factor receptor in human intestinal epithelial cells: Therapeutic implication using recombinant probiotics. Infect Immun. 2012, 80, 1079–1087. [Google Scholar] [CrossRef] [Green Version]
- Isabella, V.M.; Ha, B.N.; Castillo, M.J.; Lubkowicz, D.J.; Rowe, S.E.; Millet, Y.A.; Anderson, C.L.; Li, N.; Fisher, A.B.; West, K.A.; et al. Development of a synthetic live bacterial therapeutic for the human metabolic disease phenylketonuria. Nat. Biotechnol. 2018, 36, 857–864. [Google Scholar] [CrossRef]
- Bermudez-Humaran, L.G.; Langella, P. Live bacterial biotherapeutics in the clinic. Nat. Biotechnol. 2018, 36, 816–818. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, H.; Yang, F.; Ma, Y. Current progress on butyric acid production by fermentation. Curr. Microbiol. 2009, 59, 656–663. [Google Scholar] [CrossRef]
- Joung, S.-M.; Kurumbang, N.P.; Sang, B.-I.; Oh, M.-K. Effects of carbon source and metabolic engineering on butyrate production in E. coli. Korean J. Chem. Eng. 2011, 28, 1587–1592. [Google Scholar] [CrossRef]
- Baek, J.M.; Mazumdar, S.; Lee, S.W.; Jung, M.Y.; Lim, J.H.; Seo, S.W.; Jung, G.Y.; Oh, M.K. Butyrate production in engineered E. coli with synthetic scaffolds. Biotechnol. Bioeng. 2013, 110, 2790–2794. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Marraffini, L.A. CRISPR-Cas: New Tools for Genetic Manipulations from Bacterial Immunity Systems. Annu. Rev. Microbiol. 2015, 69, 209–228. [Google Scholar] [CrossRef] [PubMed]
- Jawed, K.; Mattam, A.J.; Fatma, Z.; Wajid, S.; Abdin, M.Z.; Yazdani, S.S. Engineered Production of Short Chain Fatty Acid in E. coli Using Fatty Acid Synthesis Pathway. PLoS ONE 2016, 11, e0160035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vital, M.; Howe, A.C.; Tiedje, J.M. Revealing the bacterial butyrate synthesis pathways by analyzing (meta)genomic data. mBio 2014, 5, e00889. [Google Scholar] [CrossRef] [Green Version]
- Boynton, Z.L.; Bennett, G.N.; Rudolph, F.B. Cloning, Sequencing, and Expression of Clustered Genes Encoding -Hydroxybutyryl-Coenzyme A (CoA) Dehydrogenase, Crotonase, and Butyryl-CoA Dehydrogenase from Clostridium acetobutylicum ATCC 824. J. Bacteriol. 1996, 178, 3015–3024. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; A. Patterson, T. Construction and use of X PL promoter vectors for direct cloning and high level expression of PCR amplified DNA coding sequences. Nucleic Acid Res. 1992, 20, 4591–4598. [Google Scholar] [CrossRef]
- Inui, M.; Suda, M.; Kimura, S.; Yasuda, K.; Suzuki, H.; Toda, H.; Yamamoto, S.; Okino, S.; Suzuki, N.; Yukawa, H. Expression of Clostridium acetobutylicum butanol synthetic genes in E. coli. Appl. Microbiol. Biotechnol. 2008, 77, 1305–1316. [Google Scholar] [CrossRef]
- Tucci, S.; Martin, W. A novel prokaryotic trans-2-enoyl-CoA reductase from the spirochete Treponema denticola. FEBS Lett. 2007, 581, 1561–1566. [Google Scholar] [CrossRef] [Green Version]
- Totemeyer, S.; Booth, N.A.; Nichols, W.W.; Dunbar, B.; Booth, I.R. From famine to feast: The role of methylglyoxal production in E. coli. Mol. Microbiol. 1998, 27, 553–562. [Google Scholar] [CrossRef]
- Royce, L.A.; Liu, P.; Stebbins, M.J.; Hanson, B.C.; Jarboe, L.R. The damaging effects of short chain fatty acids on E. coli membranes. Appl. Microbiol. Biotechnol. 2013, 97, 8317–8327. [Google Scholar] [CrossRef] [Green Version]
- Tan, Z.; Yoon, J.M.; Nielsen, D.R.; Shanks, J.V.; Jarboe, L.R. Membrane engineering via trans unsaturated fatty acids production improves E. coli robustness and production of biorenewables. Metab. Eng. 2016, 35, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontrelli, S.; Chiu, T.Y.; Lan, E.I.; Chen, F.Y.; Chang, P.; Liao, J.C. E. coli as a host for metabolic engineering. Metab. Eng. 2018, 50, 16–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, N.; Tashiro, K.; Kuhara, S.; Hayashi, T.; Sugimoto, N.; Tobe, T. Regulation of virulence by butyrate sensing in enterohaemorrhagic E. coli. Microbiology 2009, 155, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takao, M.; Yen, H.; Tobe, T. LeuO enhances butyrate-induced virulence expression through a positive regulatory loop in enterohaemorrhagic E. coli. Mol. Microbiol. 2014, 93, 1302–1313. [Google Scholar] [CrossRef]
- Torres, A.G.; Lopez-Sanchez, G.N.; Milflores-Flores, L.; Patel, S.D.; Rojas-Lopez, M.; Martinez de la Pena, C.F.; Arenas-Hernandez, M.M.; Martinez-Laguna, Y. Ler and H-NS, regulators controlling expression of the long polar fimbriae of E. coli O157:H7. J. Bacteriol. 2007, 189, 5916–5928. [Google Scholar] [CrossRef] [Green Version]
- Umanski, T.; Rosenshine, I.; Friedberg, D. Thermoregulated expression of virulence genes in enteropathogenic E. coli. Microbiology 2002, 148, 2735–2744. [Google Scholar] [CrossRef] [Green Version]
- Tan, Z.; Khakbaz, P.; Chen, Y.; Lombardo, J.; Yoon, J.M.; Shanks, J.V.; Klauda, J.B.; Jarboe, L.R. Engineering E. coli membrane phospholipid head distribution improves tolerance and production of biorenewables. Metab. Eng. 2017, 44, 1–12. [Google Scholar] [CrossRef]
- Tan, Z.; Zhu, X.; Chen, J.; Li, Q.; Zhang, X. Activating phosphoenolpyruvate carboxylase and phosphoenolpyruvate carboxykinase in combination for improvement of succinate production. Appl. Environ. Microbiol. 2013, 79, 4838–4844. [Google Scholar] [CrossRef] [Green Version]
- Lennen, R.M.; Politz, M.G.; Kruziki, M.A.; Pfleger, B.F. Identification of transport proteins involved in free fatty acid efflux in E. coli. J. Bacteriol. 2013, 195, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Deutscher, J. The mechanisms of carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 2008, 11, 87–93. [Google Scholar] [CrossRef]
- Mattson, M.P. Hormesis Defined. Ageing Res. Rev. 2008, 7, 1–7. [Google Scholar] [CrossRef]
- Mannan, A.A.; Liu, D.; Zhang, F.; Oyarzun, D.A. Fundamental Design Principles for Transcription-Factor-Based Metabolite Biosensors. ACS Synth. Biol. 2017, 6, 1851–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Chen, B.; Duan, C.; Sun, B.; Yang, J.; Yang, S. Multigene editing in the E. coli genome via the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2015, 81, 2506–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shis, D.L.; Bennett, M.R. Library of synthetic transcriptional AND gates built with split T7 RNA polymerase mutants. Proc. Natl. Acad. Sci. USA 2013, 110, 5028–5033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansell, T.J.; Linderman, S.W.; Fisher, A.C.; DeLisa, M.P. A rapid protein folding assay for the bacterial periplasm. Protein Sci. 2010, 19, 1079–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A., 3rd; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Kalejta, R.F.; Brideau, A.D.; Banfield, B.W.; Beavis, A.J. An Integral Membrane Green Fluorescent Protein Marker, Us9-GFP, Is Quantitatively Retained in Cells during Propidium Iodide-Based Cell Cycle Analysis by Flow Cytometry. Exp. Cell Res. 1999, 248, 322–328. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Plasmids/Strains | Genetic Characteristics | Source |
|---|---|---|
| Plasmids | ||
| pCas | repA101(Ts), Pcas-cas9, ParaR-Red lacIq, Ptrc-sgRNA-pMB1; KanR | [53] |
| pTargetF | pMB1, aadA; SpeR | [53] |
| pTargetF-mgsA | pMB, sgRNA-mgsA; SpeR | This study |
| pTargetF-frdA | pMB1, sgRNA-frdA; SpeR | This study |
| pTargetF-ldhA | pMB1, sgRNA-ldhA; SpeR | This study |
| pTargetF-pta | pMB1, sgRNA-pta; SpeR | This study |
| pTargetF-adhE | pMB1, sgRNA-adhE; SpeR | This study |
| pTargetF-atoDA | pMB1, sgRNA-atoDA; SpeR | This study |
| pTargetF-hns | pMB1, sgRNA-hns; SpeR | This study |
| pET28a-GFP | pBR322 ori with PT7; GFP; KanR | [54] |
| pDMB | Modified pTrc99A containing ssDsbA and bla; CmR | [55] |
| pC4S-P1 | pDMB,PpchA-pchA-PLEE1(−32 bp)-gfp; CmR | This study |
| pC4S-P2 | pDMB,PpchA-pchA-PLEE1(−98 bp)-gfp; CmR | This study |
| pC4S-P3 | pDMB,PpchA-pchA-PLEE1(−98 + 152 bp)-gfp; CmR | This study |
| pC4S-P4 | pDMB,PpchA-pchA-PLEE1(−98 + 218 bp)-gfp; CmR | This study |
| pC4S-P5 | pDMB,PpchA-pchA-PLEE1(−379 + 152 bp)-gfp; CmR | This study |
| pC4S-P6 | pDMB,PpchA-pchA-PLEE1(−379 + 444 bp)-gfp; CmR | This study |
| E.coli Strains | ||
| YF001 | E.coli Nissle 1917, wild-type | Gift of Milton Allison, Iowa State University |
| YF005 | E.coli Nissle 1917, ∆frdA, ∆ldhA, ∆adhE, ∆pta | This study |
| YF006 | YF005, PL-atoDABE | This study |
| YF007 | YF006, mgsA::PL-hbd-crt-ter | This study |
| YF023 | YF006, mgsA::PL-LacO-hbd-crt-ter | This study |
| YF021 | YF001, ∆pyrF | This study |
| YF022 | YF007, ∆pyrF | This study |
| YF121 | YF006, ∆hns | This study |
| YF122 | YF007, ∆hns | This study |
| YF126 | YF023, ∆hns | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, Y.; Mansell, T.J. Production and Sensing of Butyrate in a Probiotic E. coli Strain. Int. J. Mol. Sci. 2020, 21, 3615. https://doi.org/10.3390/ijms21103615
Bai Y, Mansell TJ. Production and Sensing of Butyrate in a Probiotic E. coli Strain. International Journal of Molecular Sciences. 2020; 21(10):3615. https://doi.org/10.3390/ijms21103615
Chicago/Turabian StyleBai, Yanfen, and Thomas J. Mansell. 2020. "Production and Sensing of Butyrate in a Probiotic E. coli Strain" International Journal of Molecular Sciences 21, no. 10: 3615. https://doi.org/10.3390/ijms21103615
APA StyleBai, Y., & Mansell, T. J. (2020). Production and Sensing of Butyrate in a Probiotic E. coli Strain. International Journal of Molecular Sciences, 21(10), 3615. https://doi.org/10.3390/ijms21103615