Broad-Spectrum Coronavirus Fusion Inhibitors to Combat COVID-19 and Other Emerging Coronavirus Diseases




Abstract
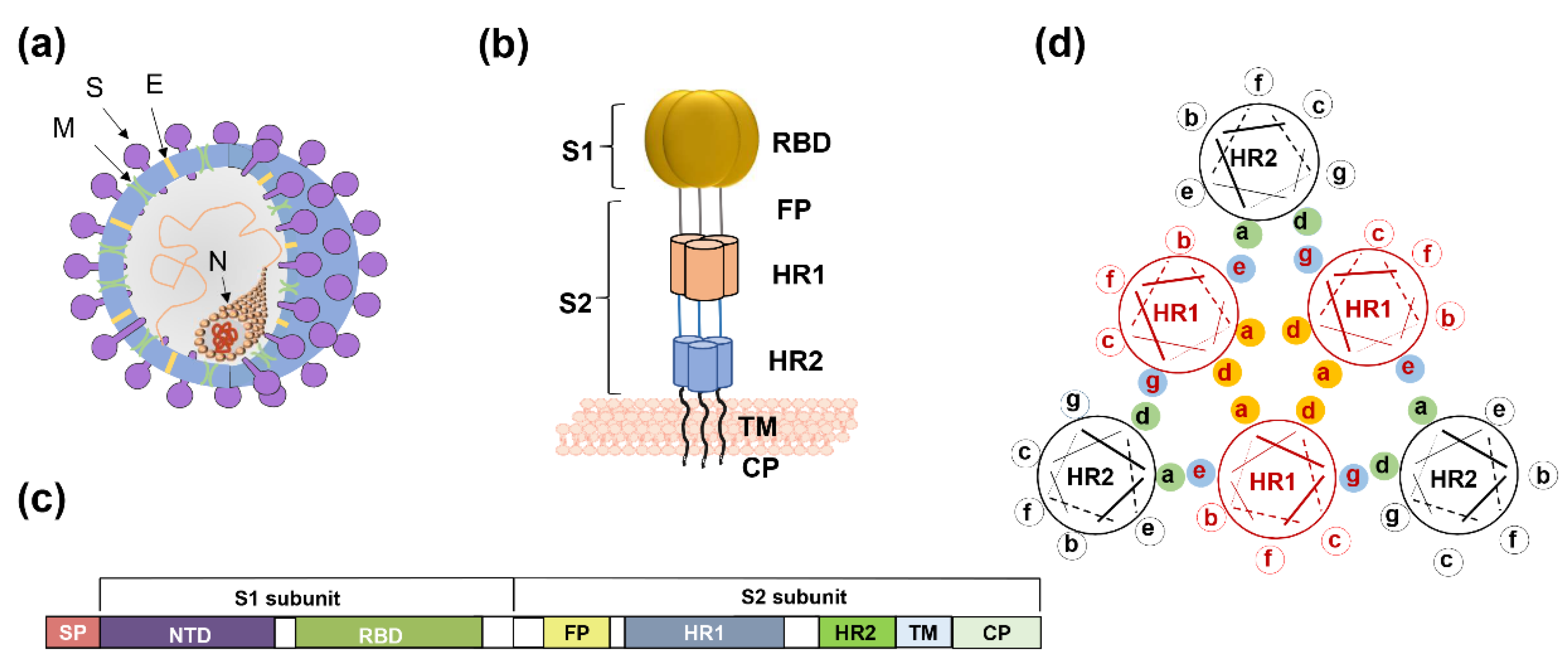
:1. Introduction
2. The Mechanism of Membrane Fusion
2.1. Receptor Recognition and Binding
2.2. Proteolytic Function in Membrane Fusion
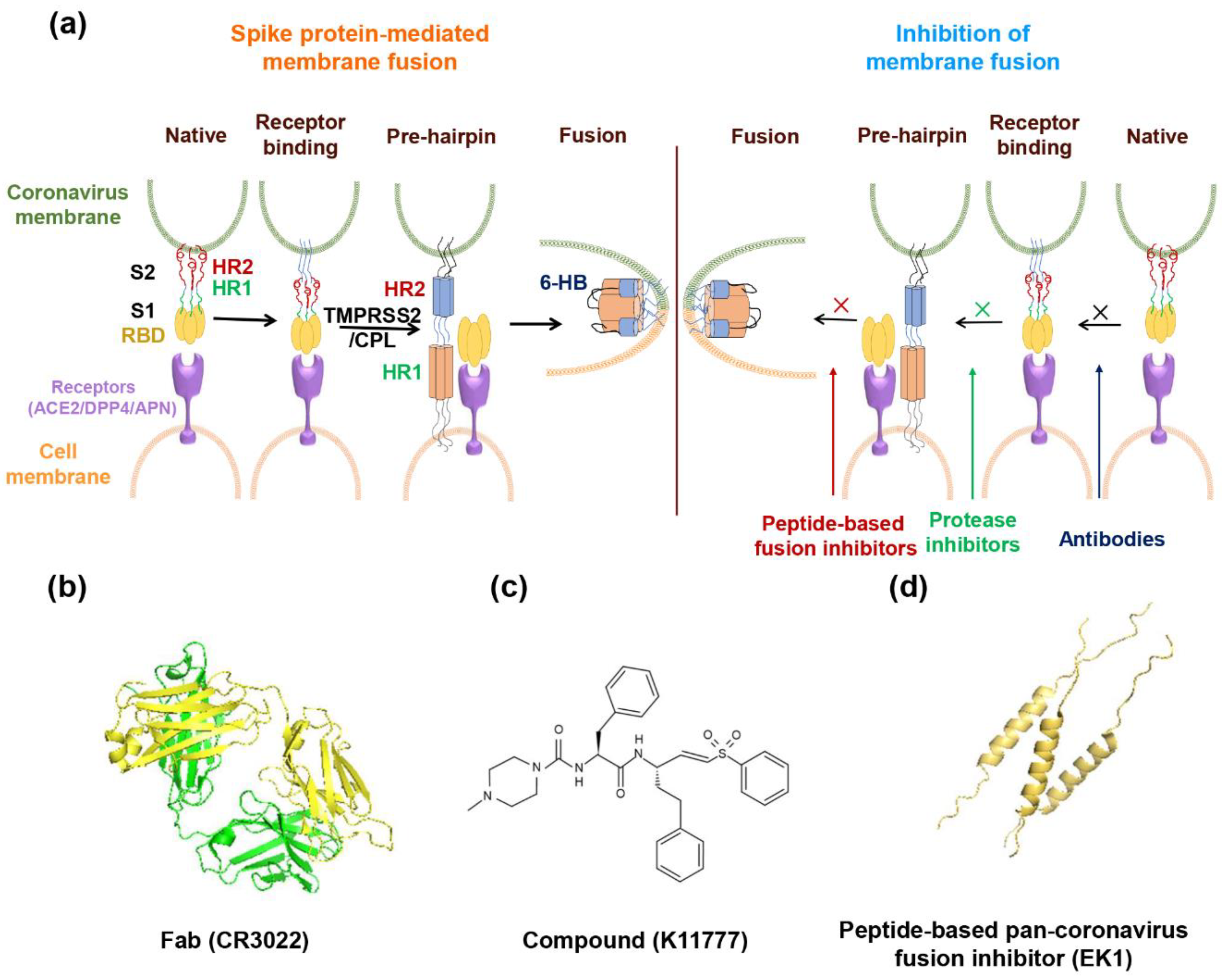
2.3. Mechanism of S2 Subunit-Mediated Membrane Fusion
3. Broad-Spectrum Coronavirus Fusion Inhibitors
3.1. Broad-Spectrum Coronavirus Fusion Inhibitors Targeting RBD
3.2. Fusion Inhibitors Targeting Fusion-Related Proteases
3.3. Broad-Spectrum Coronavirus Fusion Inhibitors Targeting the HR1 Domain
4. Advantages and Disadvantages of the Broad-Spectrum Coronavirus Fusion Inhibitors
5. Summary
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SARS | Severe acute respiratory syndrome |
| MERS | Middle East respiratory syndrome |
| COVID-19 | Coronavirus Disease 2019 |
| SARS-CoV-2 | SARS coronavirus 2 |
| 2019-nCoV | Novel coronavirus 2019 |
| HCoV-19 | Human coronavirus 2019 |
| SARSr-CoV | SARS-related coronavirus |
| NTD | N-terminal domain |
| CTD | C-terminal domain |
| RBD | Receptor-binding domain |
| ACE2 | Angiotensin-converting enzyme 2 |
| APN | Aminopeptidase N |
| DPP4 | Dipeptidyl peptidase 4 |
| CPL | Cysteine protease cathepsin L |
| TMPRSS2 | Transmembrane protease serine 2 |
| FP | Fusion peptide |
| 6-HB | Six-helix bundle |
| PRNT | Plaque reduction neutralization test |
| IFITM | Interferon-inducible transmembrane |
| NM | Nafamostat mesylate |
| FOY | Ggabexate mesylate |
| IC50 | 50% inhibitory concentration |
| CC50 | 50% cell cytotoxicity concentration |
| EC50 | Concentration for 50% of maximal effect |
| EC90 | Concentration for 90% of maximal effect |
| RAS | Renin-angiotensin system |
| AT2 | Angiotensin II receptor |
| pan-CoV | Pan-coronavirus |
References
- Coronavirus Disease (COVID-19) Pandemic. Available online: https://www.who.int/ (accessed on 26 April 2020).
- Jiang, S.; Shi, Z.; Shu, Y.; Song, J.; Gao, G.F.; Tan, W.; Guo, D. A distinct name is needed for the new coronavirus. Lancet 2020, 395, 949. [Google Scholar] [CrossRef]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Du, L.; Shi, Z. An emerging coronavirus causing pneumonia outbreak in Wuhan, China: Calling for developing therapeutic and prophylactic strategies. Emerg. Microbes Infect. 2020, 9, 275–277. [Google Scholar] [CrossRef] [PubMed]
- Prioritizing Diseases for Research and Development in Emergency Contexts. Available online: https://www.who.int/activities/prioritizing-diseases-for-research-and-development-in-emergency-contexts (accessed on 26 April 2020).
- Jiang, S.; Shi, Z. The First Disease X Is Caused by a Highly Transmissible Acute Respiratory Syndrome Coronavirus. Virol. Sin. 2020. [Google Scholar] [CrossRef] [Green Version]
- De Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Middle East Respiratory Syndrome Coronavirus (MERS-CoV). MERS Monthly Summary, November 2019. Available online: https://www.who.int/emergencies/mers-cov/en/ (accessed on 26 April 2020).
- Coronavirus Disease (COVID-19) Pandemic. Coronavirus Disease (COVID-19) Outbreak Situation. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 26 April 2020).
- Cheng, V.C.; Lau, S.K.; Woo, P.C.; Yuen, K.Y. Severe acute respiratory syndrome coronavirus as an agent of emerging and reemerging infection. Clin. Microbiol. Rev. 2007, 20, 660–694. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.F.; Lau, S.K.; To, K.K.; Cheng, V.C.; Woo, P.C.; Yuen, K.Y. Middle East Respiratory Syndrome Coronavirus: Another Zoonotic Betacoronavirus Causing SARS-Like Disease. Clin. Microbiol. Rev. 2015, 28, 465–522. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Yang, X.; Wang, X.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.; Zhu, Y.; Li, B.; Huang, C.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Bidon, M.; Jaimes, J.A.; Whittaker, G.R.; Daniel, S. Coronavirus membrane fusion mechanism offers a potential target for antiviral development. Antivir. Res. 2020, 178, 104792. [Google Scholar] [CrossRef]
- Baranowski, E.; Ruiz-Jarabo, C.M.; Domingo, E. Evolution of cell recognition by viruses. Science 2001, 292, 1102–1105. [Google Scholar] [CrossRef] [PubMed]
- Wrapp, D.; Wang, N.S.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292 e286. [Google Scholar] [CrossRef] [PubMed]
- Kuba, K.; Imai, Y.; Rao, S.A.; Gao, H.; Guo, F.; Guan, B.; Huan, Y.; Yang, P.; Zhang, Y.L.; Deng, W.; et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Bourgonje, A.R.; Abdulle, A.E.; Timens, W.; Hillebrands, J.L.; Navis, G.J.; Gordijn, S.J.; Bolling, M.C.; Dijkstra, G.; Voors, A.A.; Osterhaus, A.; et al. Angiotensin-converting enzyme-2 (ACE2), SARS-CoV-2 and pathophysiology of coronavirus disease 2019 (COVID-19). J. Pathol. 2020. [Google Scholar] [CrossRef]
- Wu, K.; Li, W.; Peng, G.; Li, F. Crystal structure of NL63 respiratory coronavirus receptor-binding domain complexed with its human receptor. Proc. Natl. Acad. Sci. USA 2009, 106, 19970–19974. [Google Scholar] [CrossRef] [Green Version]
- Yeager, C.L.; Ashmun, R.A.; Williams, R.K.; Cardellichio, C.B.; Shapiro, L.H.; Look, A.T.; Holmes, K.V. Human Aminopeptidase-N Is a Receptor for Human Coronavirus-229E. Nature 1992, 357, 420–422. [Google Scholar] [CrossRef] [Green Version]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.; Muller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.; Zaki, A.; Fouchier, R.A.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef] [Green Version]
- Li, F. Receptor recognition and cross-species infections of SARS coronavirus. Antivir. Res. 2013, 100, 246–254. [Google Scholar] [CrossRef]
- Li, F. Evidence for a Common Evolutionary Origin of Coronavirus Spike Protein Receptor-Binding Subunits. J. Virol. 2012, 86, 2856–2858. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Xia, S.; Wang, X.; Lan, Q.; Xu, W.; Wang, Q.; Jiang, S.; Lu, L. Inefficiency of Sera from Mice Treated with Pseudotyped SARS-CoV to Neutralize 2019-nCoV Infection. Virol. Sin. 2020. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Zhao, G.; Yang, Y.; Qiu, H.; Wang, L.; Kou, Z.; Tao, X.; Yu, H.; Sun, S.; Tseng, C.T.; et al. A Conformation-Dependent Neutralizing Monoclonal Antibody Specifically Targeting Receptor-Binding Domain in Middle East Respiratory Syndrome Coronavirus Spike Protein. J. Virol. 2014, 88, 7045–7053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Lu, H.; Siddiqui, P.; Zhou, Y.; Jiang, S. Receptor-binding domain of severe acute respiratory syndrome coronavirus spike protein contains multiple conformation-dependent epitopes that induce highly potent neutralizing antibodies. J. Immunol. 2005, 174, 4908–4915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, J.; Li, W.; Murakami, A.; Tamin, A.; Matthews, L.J.; Wong, S.K.; Moore, M.J.; Tallarico, A.S.; Olurinde, M.; Choe, H.; et al. Potent neutralization of severe acute respiratory syndrome (SARS) coronavirus by a human mAb to S1 protein that blocks receptor association. Proc. Natl. Acad. Sci. USA 2004, 101, 2536–2541. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; He, Y.; Zhou, Y.; Liu, S.; Zheng, B.; Jiang, S. The spike protein of SARS-CoV—A target for vaccine and therapeutic development. Nat. Rev. Microbiol. 2009, 7, 226–236. [Google Scholar] [CrossRef]
- White, J.M.; Whittaker, G.R. Fusion of Enveloped Viruses in Endosomes. Traffic 2016, 17, 593–614. [Google Scholar] [CrossRef] [Green Version]
- Madu, I.G.; Roth, S.L.; Belouzard, S.; Whittaker, G.R. Characterization of a Highly Conserved Domain within the Severe Acute Respiratory Syndrome Coronavirus Spike Protein S2 Domain with Characteristics of a Viral Fusion Peptide. J. Virol. 2009, 83, 7411–7421. [Google Scholar] [CrossRef] [Green Version]
- Millet, J.K.; Whittaker, G.R. Host cell proteases: Critical determinants of coronavirus tropism and pathogenesis. Virus Res. 2015, 202, 120–134. [Google Scholar] [CrossRef]
- Millet, J.K.; Whittaker, G.R. Host cell entry of Middle East respiratory syndrome coronavirus after two-step, furin-mediated activation of the spike protein. Proc. Natl. Acad. Sci. USA 2014, 111, 15214–15219. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Liu, M.; Wang, C.; Xu, W.; Lan, Q.; Feng, S.; Qi, F.F.; Bao, L.; Du, L.; Liu, S.; et al. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res. 2020, 30, 343–355. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.Y.; Li, J.L.; Yang, X.L.; Chmura, A.A.; Zhu, G.; Epstein, J.H.; Mazet, J.K.; Hu, B.; Zhang, W.; Peng, C.; et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 2013, 503, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Menachery, V.D.; Yount, B.L.; Sims, A.C.; Debbink, K.; Agnihothram, S.S.; Gralinski, L.E.; Graham, R.L.; Scobey, T.; Plante, J.A.; Royal, S.R.; et al. SARS-like WIV1-CoV poised for human emergence. Proc. Natl. Acad. Sci. USA 2016, 113, 3048–3053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earnest, J.T.; Hantak, M.P.; Li, K.; McCray, P.B., Jr.; Perlman, S.; Gallagher, T. The tetraspanin CD9 facilitates MERS-coronavirus entry by scaffolding host cell receptors and proteases. PLoS Pathog. 2017, 13, e1006546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e278. [Google Scholar] [CrossRef]
- Liu, S.; Xiao, G.; Chen, Y.; He, Y.; Niu, J.; Escalante, C.R.; Xiong, H.; Farmar, J.; Debnath, A.K.; Tien, P.; et al. Interaction between heptad repeat 1 and 2 regions in spike protein of SARS-associated coronavirus: Implications for virus fusogenic mechanism and identification of fusion inhibitors. Lancet 2004, 363, 938–947. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Liu, Q.; Zhu, Y.; Chan, K.H.; Qin, L.; Li, Y.; Wang, Q.; Chan, J.F.; Du, L.; Yu, F.; et al. Structure-based discovery of Middle East respiratory syndrome coronavirus fusion inhibitor. Nat. Commun. 2014, 5, 3067. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Zhu, Y.; Liu, M.; Lan, Q.; Xu, W.; Wu, Y.; Ying, T.; Liu, S.; Shi, Z.; Jiang, S.; et al. Fusion mechanism of 2019-nCoV and fusion inhibitors targeting HR1 domain in spike protein. Cell Mol. Immunol. 2020. [Google Scholar] [CrossRef]
- Du, L.; Yang, Y.; Zhou, Y.; Lu, L.; Li, F.; Jiang, S. MERS-CoV spike protein: A key target for antivirals. Expert Opin. Ther. Targets 2017, 21, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Huentelman, M.J.; Zubcevic, J.; Prada, J.A.H.; Xiao, X.D.; Dimitrov, D.S.; Raizada, M.K.; Ostrov, D.A. Structure-based discovery of a novel angiotensin converting enzyme 2 inhibitor. Hypertension 2004, 44, 903–906. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Yan, L.; Xu, W.; Agrawal, A.S.; Algaissi, A.; Tseng, C.K.; Wang, Q.; Du, L.; Tan, W.; Wilson, I.A.; et al. A pan-coronavirus fusion inhibitor targeting the HR1 domain of human coronavirus spike. Sci. Adv. 2019, 5, eaav4580. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Xu, W.; Wang, Q.; Wang, C.; Hua, C.; Li, W.; Lu, L.; Jiang, S. Peptide-Based Membrane Fusion Inhibitors Targeting HCoV-229E Spike Protein HR1 and HR2 Domains. Int. J. Mol. Sci 2018, 19, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Chakraborti, S.; He, Y.; Roberts, A.; Sheahan, T.; Xiao, X.; Hensley, L.E.; Prabakaran, P.; Rockx, B.; Sidorov, I.A.; et al. Potent cross-reactive neutralization of SARS coronavirus isolates by human monoclonal antibodies. Proc. Natl. Acad. Sci. USA 2007, 104, 12123–12128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockx, B.; Corti, D.; Donaldson, E.; Sheahan, T.; Stadler, K.; Lanzavecchia, A.; Baric, R. Structural basis for potent cross-neutralizing human monoclonal antibody protection against lethal human and zoonotic severe acute respiratory syndrome coronavirus challenge. J. Virol. 2008, 82, 3220–3235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, T.; Du, L.; Ju, T.W.; Prabakaran, P.; Lau, C.C.; Lu, L.; Liu, Q.; Wang, L.; Feng, Y.; Wang, Y.; et al. Exceptionally potent neutralization of Middle East respiratory syndrome coronavirus by human monoclonal antibodies. J. Virol. 2014, 88, 7796–7805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Yang, Y.; Huang, J.; Jiang, S.; Du, L. Advances in MERS-CoV Vaccines and Therapeutics Based on the Receptor-Binding Domain. Viruses 2019, 11, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Li, R.; Pan, Z.; Qian, C.; Yang, Y.; You, R.; Zhao, J.; Liu, P.; Gao, L.; Li, Z.; et al. Human monoclonal antibodies block the binding of SARS-CoV-2 spike protein to angiotensin converting enzyme 2 receptor. Cell Mol. Immunol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, W.; Drabek, D.; Okba, N.M.; van Haperen, R.; Osterhaus, A.D.; van Kuppeveld, F.J.; Haagmans, B.L.; Grosveld, F.; Bosch, B.J. A human monoclonal antibody blocking SARS-CoV-2 infection. Nat. Commun. 2020. [Google Scholar] [CrossRef]
- Tai, W.; He, L.; Zhang, X.; Pu, J.; Voronin, D.; Jiang, S.; Zhou, Y.; Du, L. Characterization of the receptor-binding domain (RBD) of 2019 novel coronavirus: Implication for development of RBD protein as a viral attachment inhibitor and vaccine. Cell Mol. Immunol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Li, C.; Huang, A.; Xia, S.; Lu, S.; Shi, Z.; Lu, L.; Jiang, S.; Yang, Z.; Wu, Y.; et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg. Microbes Infect. 2020, 9, 382–385. [Google Scholar] [CrossRef] [Green Version]
- Lei, C.; Qian, K.; Li, T.; Zhang, S.; Fu, W.; Ding, M.; Hu, S. Neutralization of SARS-CoV-2 spike pseudotyped virus by recombinant ACE2-Ig. Nat. Commun 2020, 11, 2070. [Google Scholar] [CrossRef] [Green Version]
- Imai, Y.; Kuba, K.; Rao, S.; Huan, Y.; Guo, F.; Guan, B.; Yang, P.; Sarao, R.; Wada, T.; Leong-Poi, H.; et al. Angiotensin-converting enzyme 2 protects from severe acute lung failure. Nature 2005, 436, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Magrone, T.; Magrone, M.; Jirillo, E. Focus on Receptors for Coronaviruses with Special Reference to Angiotensin-converting Enzyme 2 as a Potential Drug Target—A Perspective. Endocr. Metab. Immune Disord. Drug Targets 2020. [Google Scholar] [CrossRef] [PubMed]
- Wrensch, F.; Winkler, M.; Pohlmann, S. IFITM Proteins Inhibit Entry Driven by the MERS-Coronavirus Spike Protein: Evidence for Cholesterol-Independent Mechanisms. Viruses-Basel 2014, 6, 3683–3698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Zhou, J.; Zhang, K.; Chu, H.; Liu, D.; Poon, V.K.; Chan, C.C.; Leung, H.C.; Fai, N.; Lin, Y.; et al. A novel peptide with potent and broad-spectrum antiviral activities against multiple respiratory viruses. Sci. Rep. 2016, 6, 22008. [Google Scholar] [CrossRef] [PubMed]
- Shirato, K.; Kawase, M.; Matsuyama, S. Middle East respiratory syndrome coronavirus infection mediated by the transmembrane serine protease TMPRSS2. J. Virol. 2013, 87, 12552–12561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Schroeder, S.; Kleine-Weber, H.; Muller, M.A.; Drosten, C.; Pohlmann, S. Nafamostat mesylate blocks activation of SARS-CoV-2: New treatment option for COVID-19. Antimicrob. Agents Chemother. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, M.; Matsuyama, S.; Li, X.; Takeda, M.; Kawaguchi, Y.; Inoue, J.; Matsuda, Z. Identification of Nafamostat as a Potent Inhibitor of Middle East Respiratory Syndrome Coronavirus S Protein-Mediated Membrane Fusion Using the Split-Protein-Based Cell-Cell Fusion Assay. Antimicrob. Agents Chemother. 2016, 60, 6532–6539. [Google Scholar] [CrossRef] [Green Version]
- Engel, J.C.; Doyle, P.S.; Hsieh, I.; McKerrow, J.H. Cysteine protease inhibitors cure an experimental Trypanosoma cruzi infection. J. Exp. Med. 1998, 188, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Vedantham, P.; Lu, K.; Agudelo, J.; Carrion, R.; Nunneley, J.W.; Barnard, D.; Pohlmann, S.; McKerrow, J.H.; Renslo, A.R.; et al. Protease inhibitors targeting coronavirus and filovirus entry. Antivir. Res. 2015, 116, 76–84. [Google Scholar] [CrossRef]
- Ou, X.Y.; Liu, Y.; Lei, X.B.; Li, P.; Mi, D.; Ren, L.L.; Guo, L.; Guo, R.X.; Chen, T.; Hu, J.X.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [Green Version]
- Zhou, N.; Pan, T.; Zhang, J.S.; Li, Q.W.; Zhang, X.; Bai, C.; Huang, F.; Peng, T.; Zhang, J.H.; Liu, C.; et al. Glycopeptide Antibiotics Potently Inhibit Cathepsin L in the Late Endosome/Lysosome and Block the Entry of Ebola Virus, Middle East Respiratory Syndrome Coronavirus (MERS-CoV), and Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV). J. Biol. Chem. 2016, 291, 9218–9232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Ma, X.; Yu, F.; Liu, J.; Zou, F.; Pan, T.; Zhang, H. Teicoplanin potently blocks the cell entry of 2019-nCoV. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Lin, K.; Strick, N.; Neurath, A.R. Hiv-1 Inhibition by a Peptide. Nature 1993, 365, 113. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, C.A.; Nault, G.; Galkina, S.A.; Thibaudeau, K.; Bakis, P.; Bousquet-Gagnon, N.; Robitaille, M.; Bellomo, M.; Paradis, V.; Liscourt, P.; et al. Albumin-conjugated C34 Peptide HIV-1 Fusion Inhibitor: Equipotent to C34 and T-20 in Vitro With Sustained Activity in SCID-hu Thy/Liv Mice. J. Biol. Chem. 2008, 283, 34045–34052. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Lu, L.; Qi, Z.; Lu, H.; Wang, J.; Yu, X.; Chen, Y.; Jiang, S. Novel Recombinant Engineered gp41 N-terminal Heptad Repeat Trimers and Their Potential as Anti-HIV-1 Therapeutics or Microbicides. J. Biol. Chem. 2010, 285, 25506–25515. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Shi, W.; Chappell, J.D.; Joyce, M.G.; Zhang, Y.; Kanekiyo, M.; Becker, M.M.; van Doremalen, N.; Fischer, R.; Wang, N.; et al. Importance of Neutralizing Monoclonal Antibodies Targeting Multiple Antigenic Sites on the Middle East Respiratory Syndrome Coronavirus Spike Glycoprotein To Avoid Neutralization Escape. J. Virol. 2018, 92, e02002–e02017. [Google Scholar] [CrossRef] [Green Version]
- Ota, S.; Hara, Y.; Kanoh, S.; Shinoda, M.; Kawano, S.; Fujikura, Y.; Kawana, A.; Shinkai, M. Acute eosinophilic pneumonia caused by camostat mesilate: The first case report. Respir. Med. Case Rep. 2016, 19, 21–23. [Google Scholar] [CrossRef] [Green Version]
- Keck, T.; Balcom, J.H.; Antoniu, B.A.; Lewandrowski, K.; Warshaw, A.L.; Fernandez-del Castillo, C.F. Regional effects of nafamostat, a novel potent protease and complement inhibitor, on severe necrotizing pancreatitis. Surgery 2001, 130, 175–181. [Google Scholar] [CrossRef]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.Y.; Kilby, J.M.; Saag, M.S. Enfuvirtide. Expert Opin. Investig. Drugs 2002, 11, 1837–1843. [Google Scholar] [CrossRef]
- Su, S.; Rasquinha, G.; Du, L.; Wang, Q.; Xu, W.; Li, W.; Lu, L.; Jiang, S. A Peptide-Based HIV-1 Fusion Inhibitor with Two Tail-Anchors and Palmitic Acid Exhibits Substantially Improved In Vitro and Ex Vivo Anti-HIV-1 Activity and Prolonged In Vivo Half-Life. Molecules 2019, 24, 1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Inhibitor Name | Molecular Weight | Target Site | Testing Model | Activity In Vitro (IC50) | Activity In Vivo (Protect Rate) | Ref. | ||
|---|---|---|---|---|---|---|---|---|
| Cell-Cell Fusion | Pseudovirus Infection | Live Virus Infection | ||||||
| ACE2-Ig | 111.38 KD (protein) | ACE2 | In vitro 293T/ACE2 cell | 0.8 μg/mL (SARS-CoV) 0.1 μg/mL (SARS-CoV-2) | 0.85 μg/mL (SARS-CoV) 0.65 μg/mL (SARS-CoV-2) | No reported | No reported | [54] |
| 47D11 | 150 KD (antibody) | RBD | In vitro Vero E6 cell | 20 μg/mL (significantly inhibit, SARS-CoV) 20 μg/mL (significantly inhibit, SARS-CoV-2) | 0.06 μg/mL (SARS-CoV) 0.08 μg/mL (SARS-CoV-2) | 0.19 μg/mL (SARS-CoV) 0.57 μg/mL (SARS-CoV-2) | No reported | [51] |
| Inhibitor Name | Molecular Weight | Target Site | Testing Model | Activity In Vitro (IC50) | Activity In Vivo (Protect Rate) | Ref. | ||
|---|---|---|---|---|---|---|---|---|
| Cell-Cell Fusion | Pseudovirus Infection | Live Virus Infection | ||||||
| Camostat mesylate | 494.52 (compound) | TMPRSS2 | In vitro: Calu-3; 293FT; Vero/TMPRSS2 cell In vivo: BALB/c mice | About 100 nM (MERS-CoV, 293FT cell) | 444 nM (MERS-CoV) 198 nM (SARS-CoV) 87 nM (SARS-CoV-2) | About 100 nM (significantly inhibit, MERS-CoV) | ~60% (SARS-CoV lethal model) | [60,61,63] |
| Nafamostat mesylate | 539.58 (compound) | TMPRSS2 | In vitro: Caclu-3 293FT; Vero/TMPRSS2 cell | 100 nM (MERS-CoV, 293FT cell) 1 nM (MERS-CoV, calu-3 cell) | 5.9 nM (MERS-CoV) 1.4 nM (SARS-CoV) 5 nM (SARS-CoV-2) | 1 nM (significantly inhibit, MERS-CoV) | No reported | [60,61] |
| Gabexate mesylate | 417.48 (compound) | TMPRSS2 | In vitro: Calu-3 cell 293FT cell | >1 × 105 nM (MERS-CoV, 293FT cell) | 1.15 × 105 nM (SARS-CoV) 1.2 × 109 nM (SARS-CoV-2) | No reported | No reported | [60,61] |
| K11777 | No reported (compound) | CPL | In vitro: 293T/ACE2 293T/CD13 Vero cells Vero 76 cells | No reported | 0.68 nM (SARS-CoV) 1.48 nM (HCoV-229E) 6.78 nM (HCoV-NL63) 46.12 nM (MERS-CoV) | <0.05 nM (SARS-CoV) | No reported | [63] |
| Teicoplanin | 1709.39 (compound) | CPL | In vitro: A549 cells HEK293T | No reported | 630 nM (MERS-CoV) 3760 nM (SARS-CoV) 1660 nM (SARS-CoV-2) | No reported | No reported | [65,66] |
| Inhibitor Name | Molecular Weight | Target Site | Testing Model | Activity In Vitro (IC50) | Activity In Vivo (Protect Rate) | Ref. | ||
|---|---|---|---|---|---|---|---|---|
| Cell-Cell Fusion | Pseudovirus Infection | Live Virus Infection | ||||||
| OC43-HR2P | 4270.86 (peptide) | HR1 | In vitro Huh-7 cells | 390 nM (MERS-CoV) 540 nM (SARS-CoV) 660 nM (HCoV-OC43) 840 nM (HCoV-229E) 940 nM (HCoV-NL63) | Similar to EK1 (1810 nM, HCoV-OC43)) | 930 nM (HCoV-OC43) | No reported | [44] |
| EK1 | 4331.98 (peptide) | HR1 | In vitro Huh-7 cells 293T/ACE2 cells In vivo Balb/c mice | 315 nM (SARS-CoV-2) 180 nM (MERS-CoV) 270 nM (SARS-CoV) 330 nM (HCoV-OC43) 150 nM (HCoV-229E) 630 nM (HCoV-NL63) | 2375 nM (SARS-CoV-2) 260 nM (MERS-CoV) 2230 0M (SARS-CoV) 3350 nM (HCoV-229E) 6020 nM (HCoV-NL63) 1810 nM (HCoV-OC43) 2250 nM (Rs 3367) 2010 nM (WIVI) | 2468 nM (SARS-CoV-2) 110 nM (MERS-CoV) 620 nM (HCoV-OC43) 690 nM (HCoV-229E) 480 nM (HCoV-NL63) | 100% (prophylactic; 0.5 h pre-infection) 66.7% (therapeutic; 0.5 h post-infection) | [34,44] |
| EK1C4 | 5436.08 (lipopeptide) | HR1 | In vitro Huh-7 cells 293T/ACE2 cells In vivo Balb/c mice | 1.3 nM (SARS-CoV-2) 4.3 nM (SARS-CoV) 2.5 nM (MERS-CoV) 7.7 nM (HCoV-OC43) 5.2 nM (HCoV-229E) 21.4 nM (HCoV-NL63) 4.5 nM (WIVI) 8.1 nM (Rs 3367) 4.3 nM (SHC014) | 15.8 nM (SARS-CoV-2) 11.7 nM (SARS-CoV) 11.1 nM (MERS-CoV) 37.7 nM (HCoV-OC43) 12.4 nM (HCoV-229E) 76.6 nM (HCoV-NL63) 30.8 nM (WIVI) 66.9 nM (Rs 3367) | 36.5 nM (SARS-CoV-2) 4.2 nM (MERS-CoV) 24.8 nM (HCoV-OC43) 101.5 nM (HCoV-229E) 187.6 nM (HCoV-NL63) | 100% (prophylactic; 4 h pre-infection) 100% (therapeutic; 0.5 h post- infection) | [34,44] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Xia, S.; Wang, Q.; Xu, W.; Li, W.; Lu, L.; Jiang, S. Broad-Spectrum Coronavirus Fusion Inhibitors to Combat COVID-19 and Other Emerging Coronavirus Diseases. Int. J. Mol. Sci. 2020, 21, 3843. https://doi.org/10.3390/ijms21113843
Wang X, Xia S, Wang Q, Xu W, Li W, Lu L, Jiang S. Broad-Spectrum Coronavirus Fusion Inhibitors to Combat COVID-19 and Other Emerging Coronavirus Diseases. International Journal of Molecular Sciences. 2020; 21(11):3843. https://doi.org/10.3390/ijms21113843
Chicago/Turabian StyleWang, Xinling, Shuai Xia, Qian Wang, Wei Xu, Weihua Li, Lu Lu, and Shibo Jiang. 2020. "Broad-Spectrum Coronavirus Fusion Inhibitors to Combat COVID-19 and Other Emerging Coronavirus Diseases" International Journal of Molecular Sciences 21, no. 11: 3843. https://doi.org/10.3390/ijms21113843
APA StyleWang, X., Xia, S., Wang, Q., Xu, W., Li, W., Lu, L., & Jiang, S. (2020). Broad-Spectrum Coronavirus Fusion Inhibitors to Combat COVID-19 and Other Emerging Coronavirus Diseases. International Journal of Molecular Sciences, 21(11), 3843. https://doi.org/10.3390/ijms21113843