Interactive Effects of Perinatal BPA or DES and Adult Testosterone and Estradiol Exposure on Adult Urethral Obstruction and Bladder, Kidney, and Prostate Pathology in Male Mice

 and
and 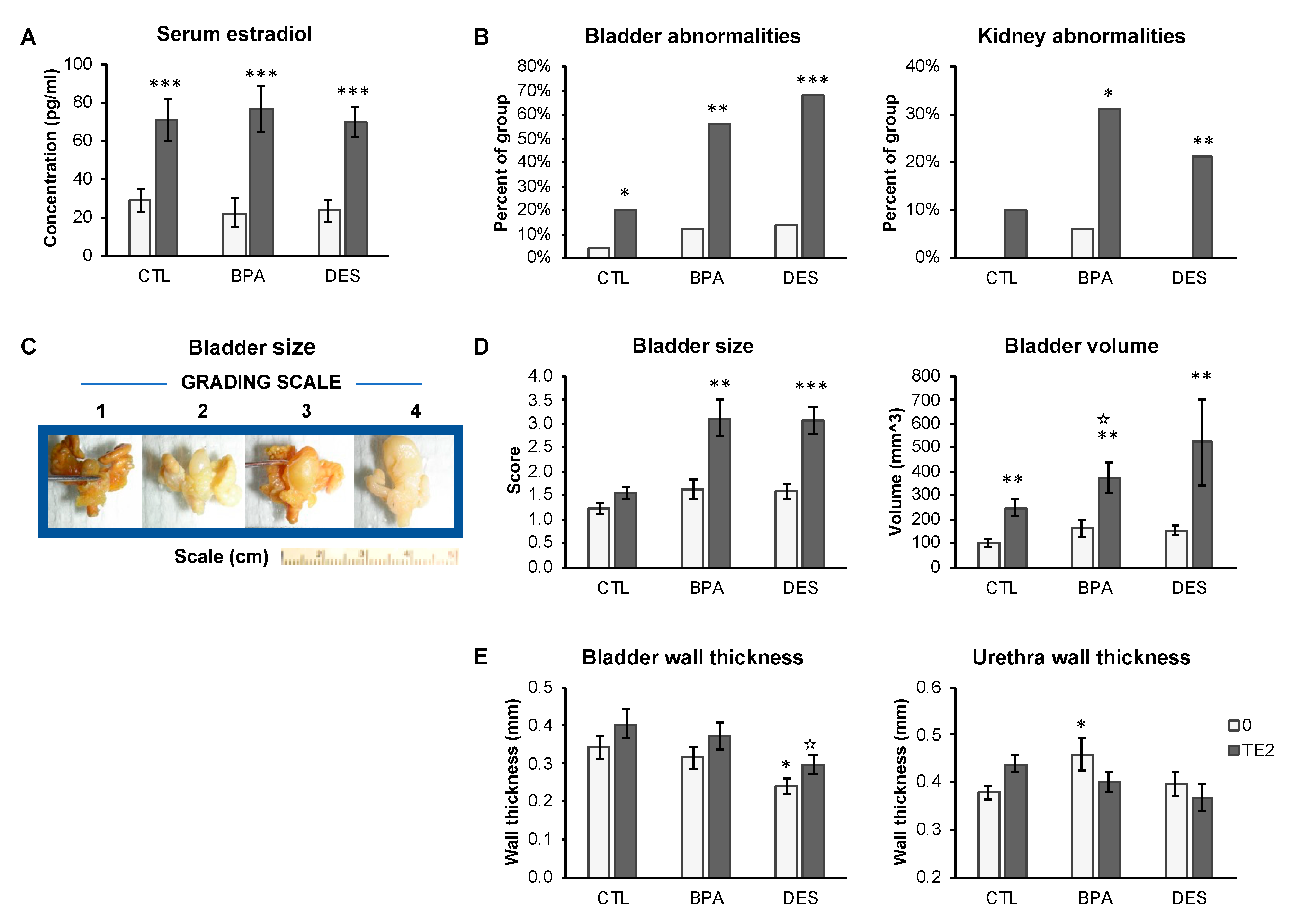{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Animal Group Designation, Sample Size, and Survival
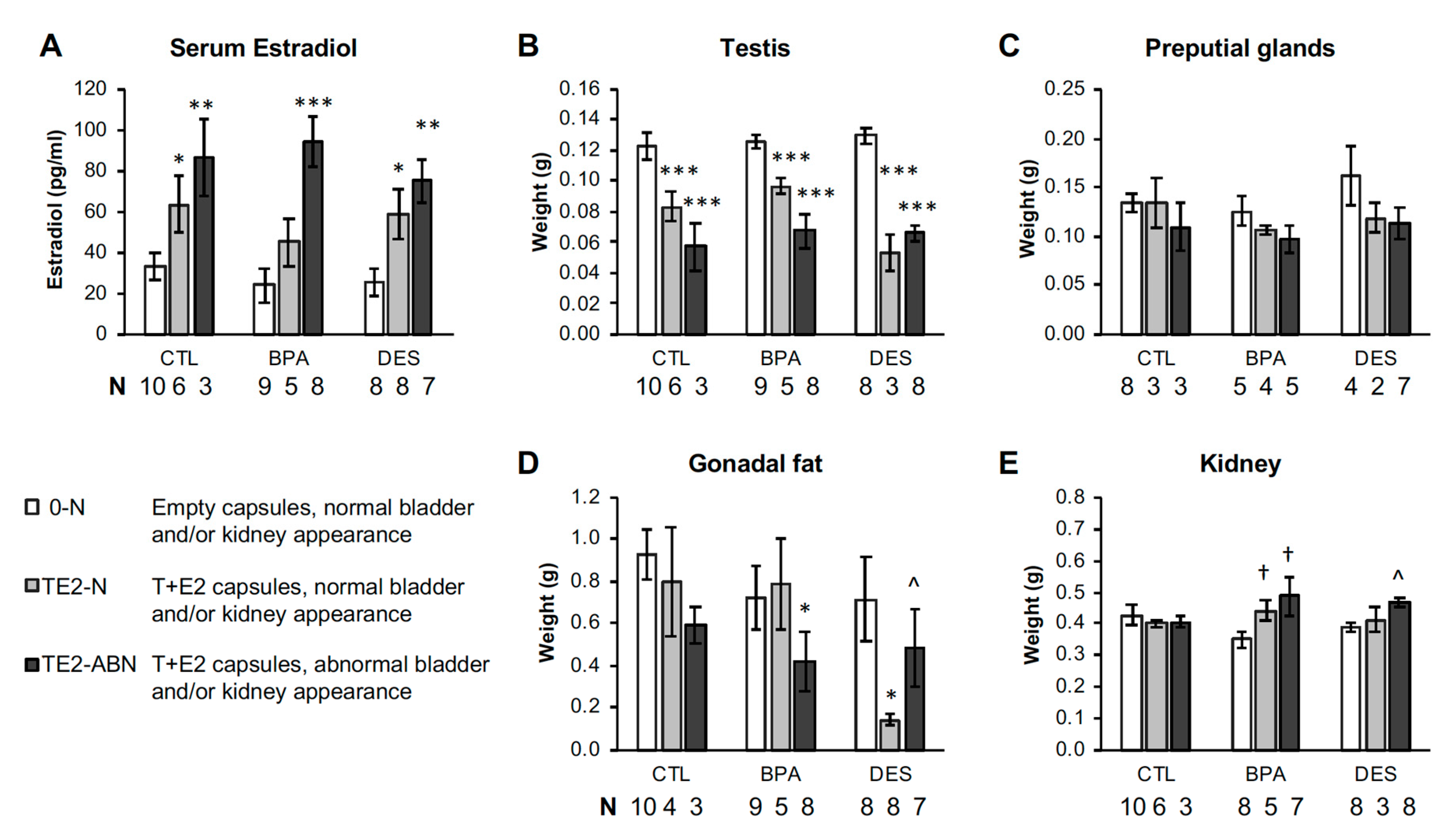
2.2. Serum Estradiol Concentrations
2.3. Bioassays for Release from Silastic Capsules of Testosterone and Estradiol
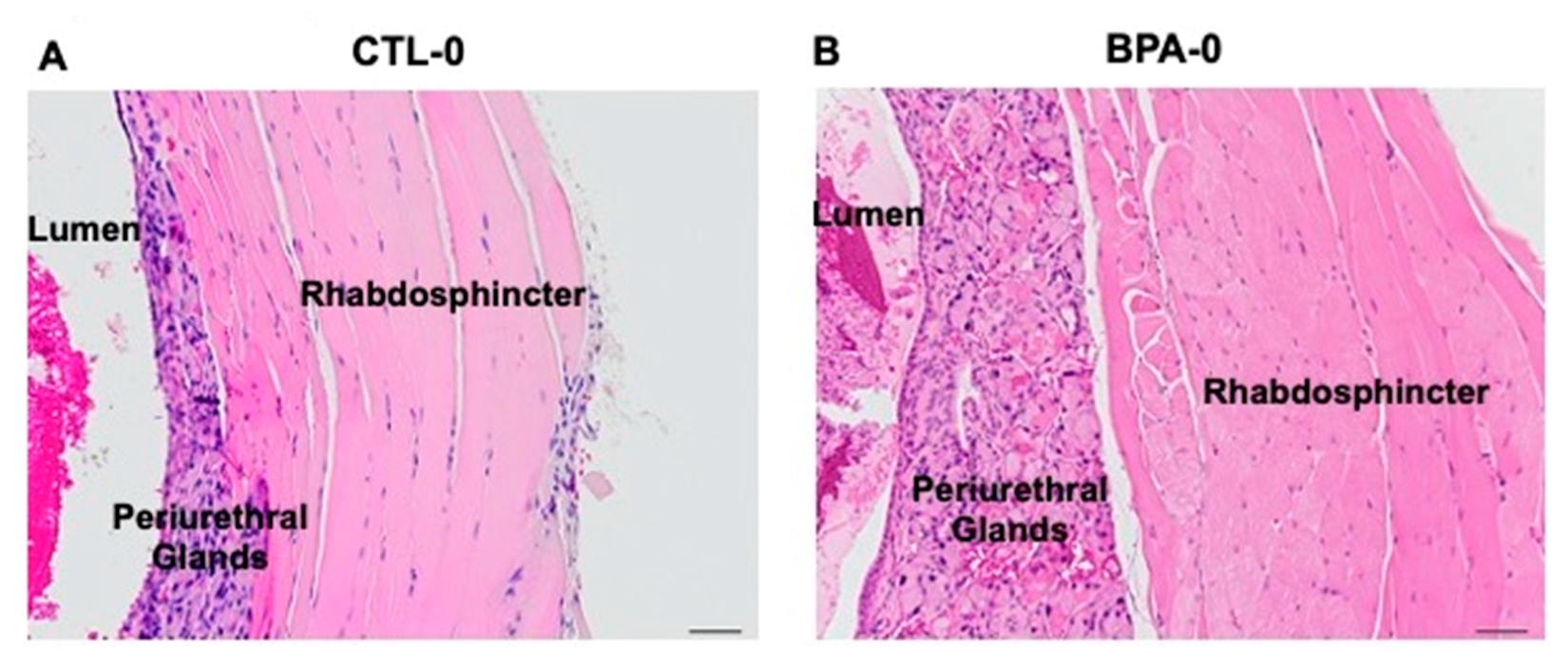
2.4. Bladder and Urethra Necropsy Findings
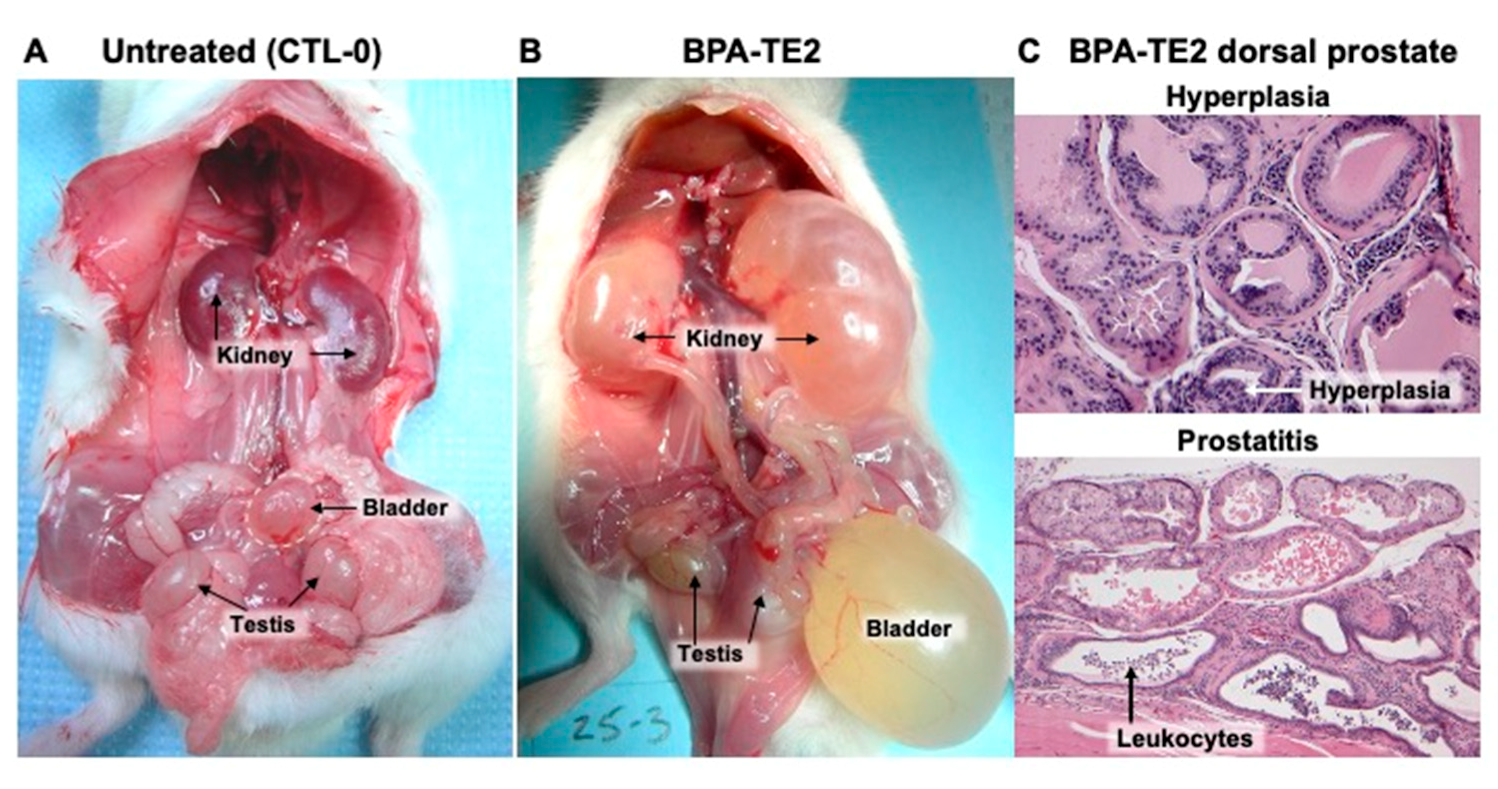
2.5. Perinatal BPA Plus Adult T+E2 Impacts the Dorsal Prostate as Well as Urine Retention in the Bladder
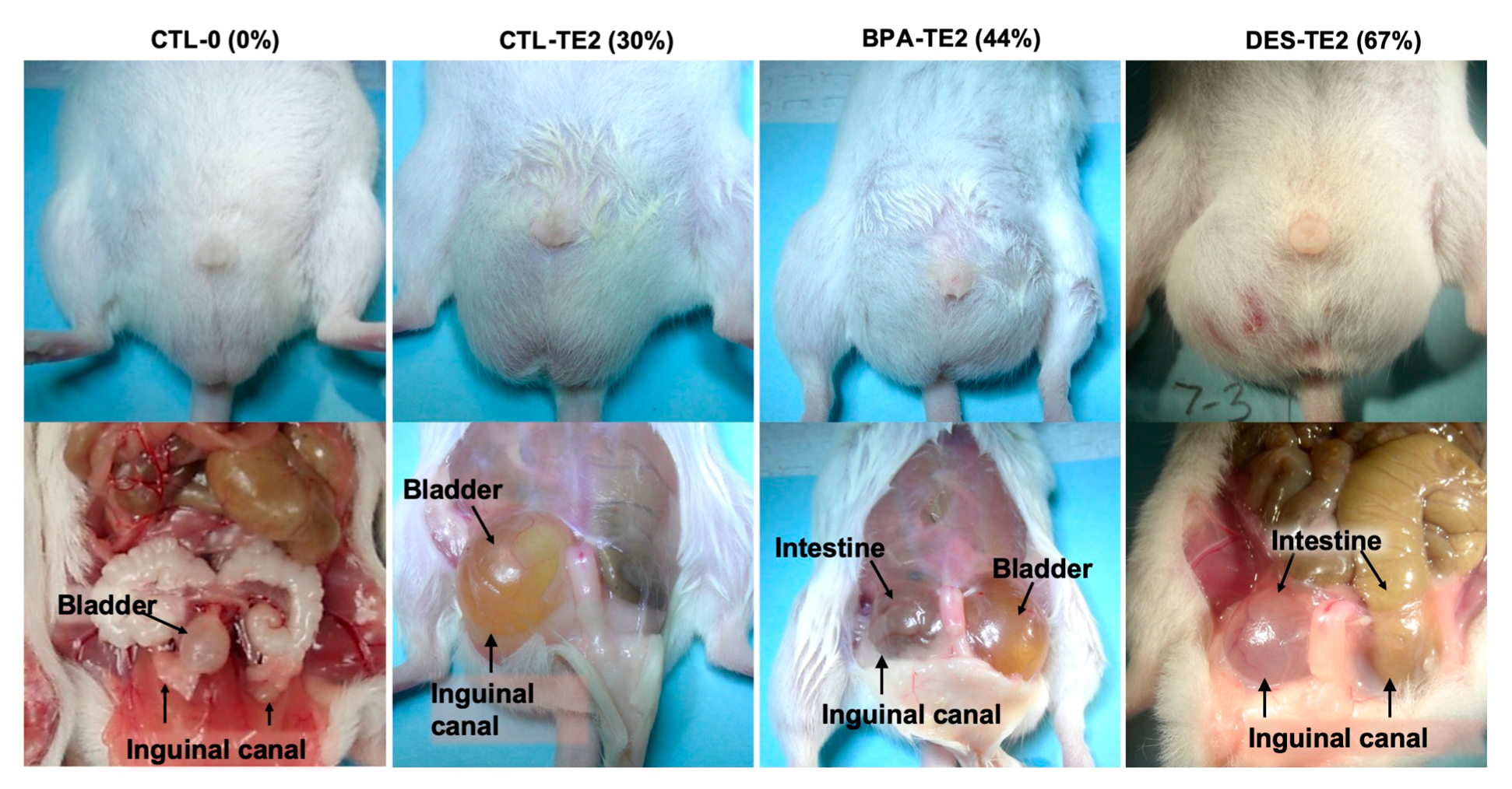
2.6. Inguinal Hernias
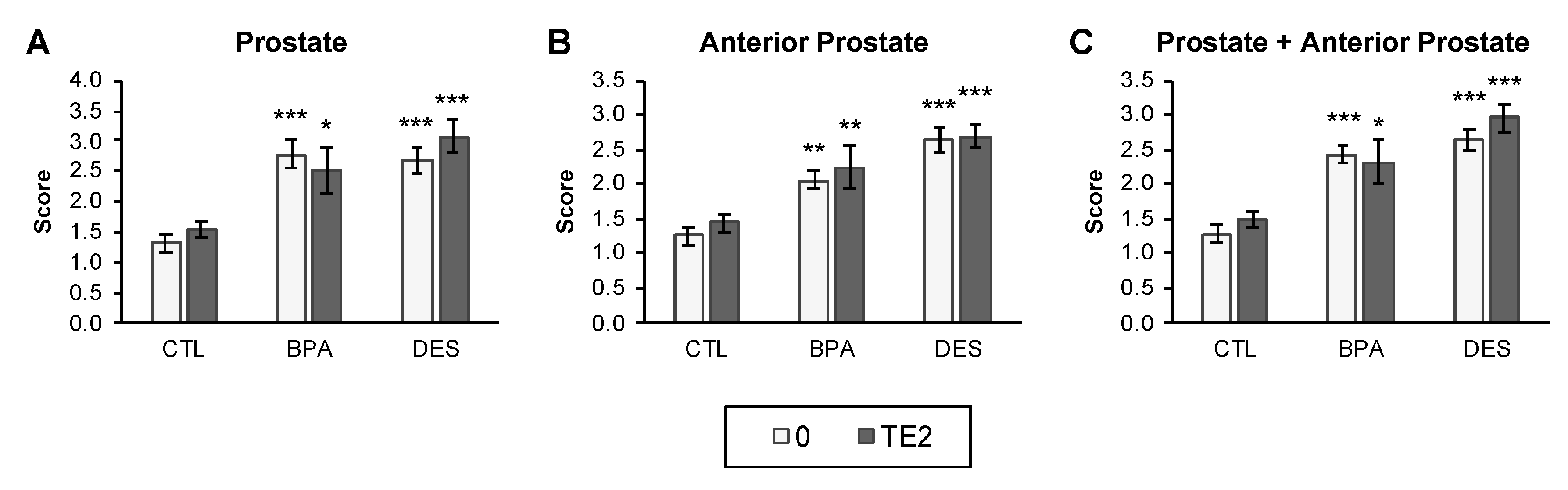
2.7. Prostate Size
2.8. Other Organ Weights at Necropsy
2.9. Daily Sperm Production and Testis Weight
2.10. Differences in Serum Estradiol in T+E2-Treated Males Impacted Phenotype
3. Discussion
3.1. Obstructive Voiding Disorder
3.2. Modulation of Androgen Receptors (AR) and Estrogen Receptors (ER)
3.3. Inguinal Hernias
3.4. Prostate Pathology
3.5. Estrogen, Obstructive Voiding Disorder, and Prostate Pathology
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Treatment of Pregnant Females and Neonates
4.4. Treatment of Neonatal Males with DES or BPA
4.5. Adult Hormone Treatment
4.6. Silastic Capsule Bioassay
4.7. Necropsy
4.8. External Examination of Fixed Urogenital Tract Specimens
4.9. Micro-CT of Urogenital Tract
4.10. Serum Estradiol Assay
4.11. Histology
4.12. Daily Sperm Production
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AR | Androgen receptor |
| BPA | Bisphenol A |
| BPH | Benign prostatic hyperplasia |
| CDC | Centers for Disease Control and Prevention |
| CT | Computed tomography |
| DES | Diethylstilbestrol |
| DOAJ | Directory of open access journals |
| DSP | Daily sperm production |
| E2 | Estradiol |
| EE2 | Ethinylestradiol |
| ER | Estrogen receptor |
| FDA | Food and Drug Administration |
| GD | Gestation day |
| MDPI | Multidisciplinary Digital Publishing Institute |
| NBF | Neutral buffered formalin |
| OVD | Obstructive voiding disorder |
| PND | Postnatal day |
| SHBG | Sex hormone binding globulin |
| T | Testosterone |
| UGS | Urogenital sinus |
References
- Kok, E.T.; Schouten, B.W.; Bohnen, A.M.; Groeneveld, F.P.; Thomas, S.; Bosch, J.L. Risk factors for lower urinary tract symptoms suggestive of benign prostatic hyperplasia in a community based population of healthy aging men: The Krimpen Study. J. Urol. 2009, 181, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Lekskulchai, O.; Dietz, H.P. Is detrusor hypertrophy in women associated with voiding dysfunction? Aust. N. Z. J. Obstet. Gynaecol. 2009, 49, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, T.M.; Ricke, E.A.; Marker, P.C.; Miano, J.M.; Mayer, R.D.; Timms, B.G.; vom Saal, F.S.; Wood, R.W.; Ricke, W.A. Testosterone and 17beta-estradiol induce glandular prostatic growth, bladder outlet obstruction, and voiding dysfunction in male mice. Endocrinology 2012, 153, 5556–5565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- vom Saal, F.S.; Timms, B.G.; Montano, M.M.; Palanza, P.; Thayer, K.A.; Nagel, S.C.; Dhar, M.D.; Ganjam, V.K.; Parmigiani, S.; Welshons, W.V. Prostate enlargement in mice due to fetal exposure to low doses of estradiol or diethylstilbestrol and opposite effects at high doses. Proc. Natl. Acad. Sci. USA 1997, 94, 2056–2061. [Google Scholar] [CrossRef] [Green Version]
- Timms, B.G.; Howdeshell, K.L.; Barton, L.; Bradley, S.; Richter, C.A.; vom Saal, F.S. Estrogenic chemicals in plastic and oral contraceptives disrupt development of the mouse prostate and urethra. Proc. Natl. Acad. Sci. USA 2005, 102, 7014–7019. [Google Scholar] [CrossRef] [Green Version]
- Uchtmann, K.S.; Taylor, J.A.; Timms, B.G.; Stahlhut, R.W.; Ricke, E.A.; Ellersieck, M.R.; vom Saal, F.S.; Ricke, W.A. Fetal bisphenol A and ethinylestradiol exposure alters male rat urogenital tract morphology at birth: Confirmation of prior low-dose findings in CLARITY-BPA. Reprod. Toxicol. 2020, 91, 131–141. [Google Scholar] [CrossRef]
- Putz, O.; Schwartz, C.B.; Kim, S.; LeBlanc, G.A.; Cooper, R.L.; Prins, G.S. Neonatal low- and high-dose exposure to estradiol benzoate in the male rat: I. Effects on the prostate gland. Biol. Reprod. 2001, 65, 1496–1505. [Google Scholar] [CrossRef] [Green Version]
- Ogura, Y.; Ishii, K.; Kanda, H.; Kanai, M.; Arima, K.; Wang, Y.; Sugimura, Y. Bisphenol A induces permanent squamous change in mouse prostatic epithelium. Differ. Res. Biol. Divers. 2007, 75, 745–756. [Google Scholar] [CrossRef]
- Taylor, J.A.; Richter, C.A.; Ruhlen, R.L.; vom Saal, F.S. Estrogenic environmental chemicals and drugs: Mechanisms for effects on the developing male urogenital system. J. Steroid. Biochem. Mol. Biol. 2011, 127, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Bjornerem, A.; Straume, B.; Midtby, M.; Fonnebo, V.; Sundsfjord, J.; Svartberg, J.; Acharya, G.; Oian, P.; Berntsen, G.K. Endogenous sex hormones in relation to age, sex, lifestyle factors, and chronic diseases in a general population: The Tromso Study. J. Clin. Endocrinol. Metab. 2004, 89, 6039–6047. [Google Scholar] [CrossRef] [Green Version]
- Coffey, D.S.; Walsh, P.C. Clinical and experimental studies of benign prostatic hyperplasia. Urol. Clin. N. Am. 1990, 17, 461–475. [Google Scholar]
- Richter, C.A.; Timms, B.G.; vom Saal, F.S. Prostate Development: Mechanisms for opposite effects of low and high doses of estrogenic chemicals. In Endocrine Disruptors: Effects on Male and Female Reproductive Systems, 2nd ed.; Naz, R.K., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 379–410. [Google Scholar]
- Glenister, T.W. The development of the utricle and of the so-called “middle” or “median” lobe of the human prostate. J. Anat. (Lond.) 1962, 96, 443–455. [Google Scholar]
- Nicholson, T.M.; Nguyen, J.L.; Leverson, G.E.; Taylor, J.A.; vom Saal, F.S.; Wood, R.W.; Ricke, W.A. Endocrine disruptor bisphenol A is implicated in urinary voiding dysfunction in male mice. Am. J. Physiol. Renal. Physiol. 2018, 315, F1208–F1216. [Google Scholar] [CrossRef] [PubMed]
- Prins, G.S.; Hu, W.Y.; Xie, L.; Shi, G.B.; Hu, D.P.; Birch, L.; Bosland, M.C. Evaluation of Bisphenol A (BPA) Exposures on Prostate Stem Cell Homeostasis and Prostate Cancer Risk in the NCTR-Sprague-Dawley Rat: An NIEHS/FDA CLARITY-BPA Consortium Study. Environ. Health Perspect. 2018, 126, 117001. [Google Scholar] [CrossRef] [PubMed]
- Cunha, G.R.; Vezina, C.M.; Isaacson, D.; Ricke, W.A.; Timms, B.G.; Cao, M.; Franco, O.; Baskin, L.S. Development of the human prostate. Differentiation 2018, 103, 24–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonneman, D.J.; Ganjam, V.K.; Welshons, W.V.; vom Saal, F.S. Intrauterine position effects on steroid metabolism and steroid receptors of reproductive organs in male mice. Biol. Reprod. 1992, 47, 723–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timms, B.G.; Petersen, S.L.; vom Saal, F.S. Prostate gland growth during development is stimulated in both male and female rat fetuses by intrauterine proximity to female fetuses. J. Urol. 1999, 161, 1694–1701. [Google Scholar] [CrossRef]
- Risbridger, G.P.; Wang, H.; Frydenberg, M.; Cunha, G.R. The metaplastic effects of estrogen on mouse prostate epithelium: Proliferation of cells with basal cell phenotype. Endocrinology 2001, 142, 2443–2450. [Google Scholar] [CrossRef]
- Gupta, C. Reproductive malformation of the male offspring following maternal exposure to estrogenic chemicals. Proc. Soc. Exp. Biol. Med. 2000, 224, 61–68. [Google Scholar] [CrossRef]
- Richter, C.A.; Taylor, J.A.; Ruhlen, R.R.; Welshons, W.V.; vom Saal, F.S. Estradiol and bisphenol A stimulate androgen receptor and estrogen receptor gene expression in fetal mouse prostate cells. Environ. Health Perspect. 2007, 115, 902–908. [Google Scholar] [CrossRef]
- Taylor, J.A.; Richter, C.A.; Suzuki, A.; Watanabe, H.; Iguchi, T.; Coser, K.R.; Shioda, T.; vom Saal, F.S. Dose-Related Estrogen Effects on Gene Expression in Fetal Mouse Prostate Mesenchymal Cells. PLoS ONE 2012, 7, e48311. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, R.K.; Taylor, J.A.; Sommerfeld-Sager, J.; Tillitt, D.E.; Ricke, W.A.; vom Saal, F.S. Estrogen receptor 1 expression and methylation of Esr1 promoter in mouse fetal prostate mesenchymal cells induced by gestational exposure to bisphenol A or ethinylestradiol. Environ. Epigenet. 2019, 5, dvz012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thayer, K.A.; Ruhlen, R.L.; Howdeshell, K.L.; Buchanan, D.L.; Cooke, P.S.; Preziosi, D.; Welshons, W.V.; Haseman, J.; vom Saal, F.S. Altered prostate growth and daily sperm production in male mice exposed prenatally to subclinical doses of 17alpha-ethinyl oestradiol. Hum. Reprod. 2001, 16, 988–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calafat, A.M.; Ye, X.; Wong, L.Y.; Reidy, J.A.; Needham, L.L. Exposure of the U.S. population to bisphenol A and 4-tertiary-octylphenol: 2003–2004. Environ. Health Perspect. 2008, 116, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howdeshell, K.L.; Hotchkiss, A.K.; Thayer, K.A.; Vandenbergh, J.G.; vom Saal, F.S. Exposure to bisphenol A advances puberty. Nature 1999, 401, 763–764. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, N.; Silva, E.; Kortenkamp, A. Combining xenoestrogens at levels below individual no-observed-effect-concentrations dramatically enhances steroid hormone action. Environ. Health Perspect. 2002, 110, 917–921. [Google Scholar] [CrossRef] [Green Version]
- Gerona, R.; vom Saal, F.S.; Hunt, P.A. BPA: Have flawed analytical techniques compromised risk assessments? Lancet Diabetes Endocrinol. 2020, 8, 11–13. [Google Scholar] [CrossRef] [Green Version]
- OpenPR. Bisphenol A Market is Accounted for $17.69 Billion in 2017 and is Expected to Reach $31.19 Billion by 2026 Growing at a CAGR of 6.5%. 2018. Available online: https://www.openpr.com/news/1247056/Bisphenol-A-market-is-accounted-for-17-69-billion-in-2017-and-is-expected-to-reach-31-19-billion-by-2026-growing-at-a-CAGR-of-6-5.html (accessed on 23 February 2020).
- Alworth, L.C.; Howdeshell, K.L.; Ruhlen, R.L.; Day, J.K.; Lubahn, D.B.; Huang, T.H.; Besch-Williford, C.L.; vom Saal, F.S. Uterine responsiveness to estradiol and DNA methylation are altered by fetal exposure to diethylstilbestrol and methoxychlor in CD-1 mice: Effects of low versus high doses. Toxicol. Appl. Pharmacol. 2002, 183, 10–22. [Google Scholar] [CrossRef]
- Vom Saal, F.S. TRIENNIAL REPRODUCTION SYMPOSIUM: Environmental programming of reproduction during fetal life: Effects of intrauterine position and the endocrine disrupting chemical bisphenol A. J. Anim. Sci. 2016, 94, 2722–2736. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Arnold, A.P. Reframing sexual differentiation of the brain. Nat. Neurosci. 2011, 14, 677–683. [Google Scholar] [CrossRef] [Green Version]
- Newbold, R.R.; Jefferson, W.N.; Padilla-Banks, E.; Haseman, J. Developmental exposure to diethylstilbestrol (DES) alters uterine response to estrogens in prepubescent mice: Low versus high dose effects. Reprod. Toxicol. 2004, 18, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R., Jr.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhou, L.; Li, L.; Coon, V.J.; Chatterton, R.T.; Brooks, D.C.; Jiang, E.; Liu, L.; Xu, X.; Dong, Z.; et al. Shift from androgen to estrogen action causes abdominal muscle fibrosis, atrophy, and inguinal hernia in a transgenic male mouse model. Proc. Natl. Acad. Sci. USA 2018, 115, E10427–E10436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, R.D.; Neumayer, L. Inguinal hernia in the 21st century: An evidence-based review. Curr. Probl. Surg. 2008, 45, 261–312. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.C.; vom Saal, F.S.; Thayer, K.A.; Dhar, M.G.; Boechler, M.; Welshons, W.V. Relative binding affinity-serum modified access (RBA-SMA) assay predicts the relative in vivo bioactivity of the xenoestrogens bisphenol A and octylphenol. Environ. Health Perspect. 1997, 105, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Z.; Wong, Y.C. Sex hormone-induced prostatic carcinogenesis in the noble rat: The role of insulin-like growth factor-I (IGF-I) and vascular endothelial growth factor (VEGF) in the development of prostate cancer. Prostate 1998, 35, 165–177. [Google Scholar] [CrossRef]
- Parsons, J.K. Lifestyle factors, benign prostatic hyperplasia, and lower urinary tract symptoms. Curr. Opin. Urol. 2011, 21, 1–4. [Google Scholar] [CrossRef]
- Ricke, W.A.; Lee, C.W.; Clapper, T.R.; Schneider, A.J.; Moore, R.W.; Keil, K.P.; Abler, L.L.; Wynder, J.L.; Lopez Alvarado, A.; Beaubrun, I.; et al. In Utero and Lactational TCDD Exposure Increases Susceptibility to Lower Urinary Tract Dysfunction in Adulthood. Toxicol. Sci. 2016, 150, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Angle, B.M.; Do, R.P.; Ponzi, D.; Stahlhut, R.W.; Drury, B.E.; Nagel, S.C.; Welshons, W.V.; Besch-Williford, C.L.; Palanza, P.; Parmigiani, S.; et al. Metabolic disruption in male mice due to fetal exposure to low but not high doses of bisphenol A (BPA): Evidence for effects on body weight, food intake, adipocytes, leptin, adiponectin, insulin and glucose regulation. Reprod. Toxicol. 2013, 42, 256–268. [Google Scholar] [CrossRef] [Green Version]
- Sugimura, Y.; Cunha, G.R.; Donjacour, A.A. Morphogenesis of ductal networks in the mouse prostate. Biol. Reprod. 1986, 34, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Heindel, J.J.; Blumberg, B.; Cave, M.; Machtinger, R.; Mantovani, A.; Mendez, M.A.; Nadal, A.; Palanza, P.; Panzica, G.; Sargis, R.; et al. Metabolism disrupting chemicals and metabolic disorders. Reprod. Toxicol. 2017, 68, 3–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howdeshell, K.L.; Peterman, P.H.; Judy, B.M.; Taylor, J.A.; Orazio, C.E.; Ruhlen, R.L.; vom Saal, F.S.; Welshons, W.V. Bisphenol A is released from used polycarbonate animal cages into water at room temperature. Environ. Health Perspect. 2003, 111, 1180–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.A.; vom Saal, F.S.; Welshons, W.V.; Drury, B.; Rottinghaus, G.; Hunt, P.A.; Toutain, P.L.; Laffont, C.M.; Vandevoort, C.A. Similarity of bisphenol a pharmacokinetics in rhesus monkeys and mice: Relevance for human exposure. Environ. Health Perspect. 2011, 119, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Welshons, W.V.; vom Saal, F.S.; Toutain, P.L.; Myers, J.P. Should oral gavage be abandoned in toxicity testing of endocrine disruptors? Environ. Health 2014, 13, 46. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Rebuli, M.E.; Rogers, J.; Todd, K.L.; Leyrer, S.M.; Ferguson, S.A.; Patisaul, H.B. Prenatal bisphenol A exposure alters sex-specific estrogen receptor expression in the neonatal rat hypothalamus and amygdala. Toxicol. Sci. 2013, 133, 157–173. [Google Scholar] [CrossRef] [Green Version]
- vom Saal, F.S.; Cooke, P.S.; Buchanan, D.L.; Palanza, P.; Thayer, K.A.; Nagel, S.C.; Parmigiani, S.; Welshons, W.V. A physiologically based approach to the study of bisphenol A and other estrogenic chemicals on the size of reproductive organs, daily sperm production, and behavior. Toxicol. Ind. Health 1998, 14, 239–260. [Google Scholar] [CrossRef]
- Vom Saal, F.S. Flaws in design, execution and interpretation limit CLARITY-BPA’s value for risk assessments of bisphenol A. Basic Clin. Pharmacol. Toxicol. 2019, 125 (Suppl. 3), 32–43. [Google Scholar] [CrossRef]
- Narushin, V.G. Egg geometry calculation using the measurements of length and breadth. Poultry Sci. 2005, 84, 482–484. [Google Scholar] [CrossRef]
- Pauwels, E.; Van Loo, D.; Cornillie, P.; Brabant, L.; Van Hoorebeke, L. An exploratory study of contrast agents for soft tissue visualization by means of high resolution X-ray computed tomography imaging. J. Microsc. 2013, 250, 21–31. [Google Scholar] [CrossRef]
- Loening, A.M.; Gambhir, S.S. AMIDE: A free software tool for multimodality medical image analysis. Mol. Imaging 2003, 2, 131–137. [Google Scholar] [CrossRef]
- vom Saal, F.S.; Quadagno, D.M.; Even, M.D.; Keisler, L.W.; Keisler, D.H.; Khan, S. Paradoxical effects of maternal stress on fetal steroids and postnatal reproductive traits in female mice from different intrauterine positions. Biol. Reprod. 1990, 43, 751–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UCLA (University of California, Los Angeles Statistics Department). Statistics 10, Chapter 8: Hypothesis Testing for Population Proportions. Available online: https://www.coursehero.com/file/12555039/Stat-Chapter-8/ (accessed on 5 April 2020).
- Ricke, W.A.; McPherson, S.J.; Bianco, J.J.; Cunha, G.R.; Wang, Y.; Risbridger, G.P. Prostatic hormonal carcinogenesis is mediated by in situ estrogen production and estrogen receptor alpha signaling. FASEB J. 2008, 22, 1512–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellem, S.J.; Wang, H.; Poutanen, M.; Risbridger, G.P. Increased endogenous estrogen synthesis leads to the sequential induction of prostatic inflammation (prostatitis) and prostatic pre-malignancy. Am. J. Pathol. 2009, 175, 1187–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prins, G.S.; Ye, S.H.; Birch, L.; Ho, S.M.; Kannan, K. Serum bisphenol A pharmacokinetics and prostate neoplastic responses following oral and subcutaneous exposures in neonatal Sprague-Dawley rats. Reprod. Toxicol. 2011, 31, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taylor, J.A.; Jones, M.B.; Besch-Williford, C.L.; Berendzen, A.F.; Ricke, W.A.; vom Saal, F.S. Interactive Effects of Perinatal BPA or DES and Adult Testosterone and Estradiol Exposure on Adult Urethral Obstruction and Bladder, Kidney, and Prostate Pathology in Male Mice. Int. J. Mol. Sci. 2020, 21, 3902. https://doi.org/10.3390/ijms21113902
Taylor JA, Jones MB, Besch-Williford CL, Berendzen AF, Ricke WA, vom Saal FS. Interactive Effects of Perinatal BPA or DES and Adult Testosterone and Estradiol Exposure on Adult Urethral Obstruction and Bladder, Kidney, and Prostate Pathology in Male Mice. International Journal of Molecular Sciences. 2020; 21(11):3902. https://doi.org/10.3390/ijms21113902
Chicago/Turabian StyleTaylor, Julia A., Maren Bell Jones, Cynthia L. Besch-Williford, Ashley F. Berendzen, William A. Ricke, and Frederick S. vom Saal. 2020. "Interactive Effects of Perinatal BPA or DES and Adult Testosterone and Estradiol Exposure on Adult Urethral Obstruction and Bladder, Kidney, and Prostate Pathology in Male Mice" International Journal of Molecular Sciences 21, no. 11: 3902. https://doi.org/10.3390/ijms21113902
APA StyleTaylor, J. A., Jones, M. B., Besch-Williford, C. L., Berendzen, A. F., Ricke, W. A., & vom Saal, F. S. (2020). Interactive Effects of Perinatal BPA or DES and Adult Testosterone and Estradiol Exposure on Adult Urethral Obstruction and Bladder, Kidney, and Prostate Pathology in Male Mice. International Journal of Molecular Sciences, 21(11), 3902. https://doi.org/10.3390/ijms21113902