Norepinephrine Inhibits the Proliferation of Human Bone Marrow-Derived Mesenchymal Stem Cells via β2-Adrenoceptor-Mediated ERK1/2 and PKA Phosphorylation

 , , and
, , and
Abstract
:1. Introduction
2. Results
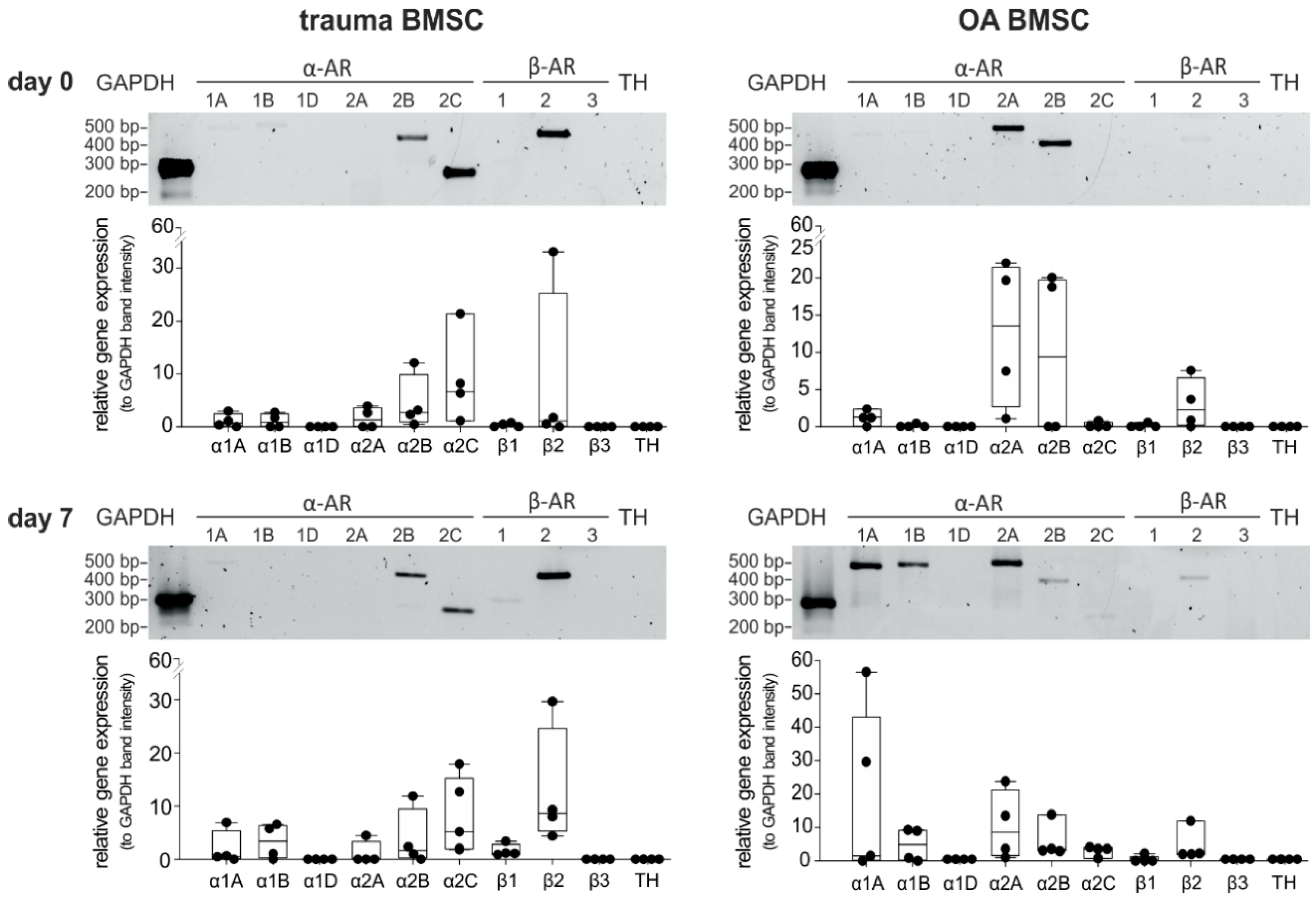
2.1. AR and TH Gene Expression Profile of BMSCs
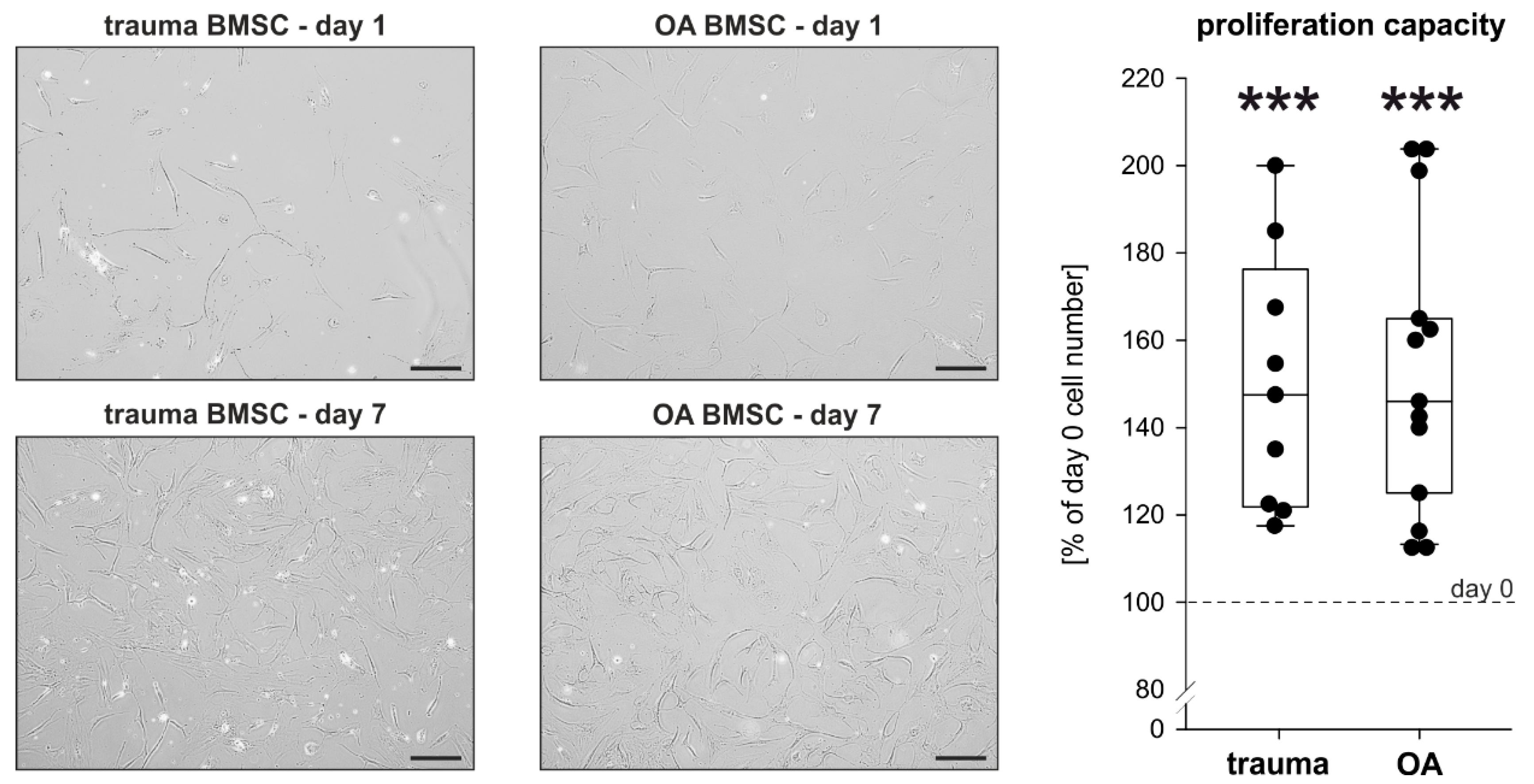
2.2. Proliferation Capacity of Trauma and OA BMSCs
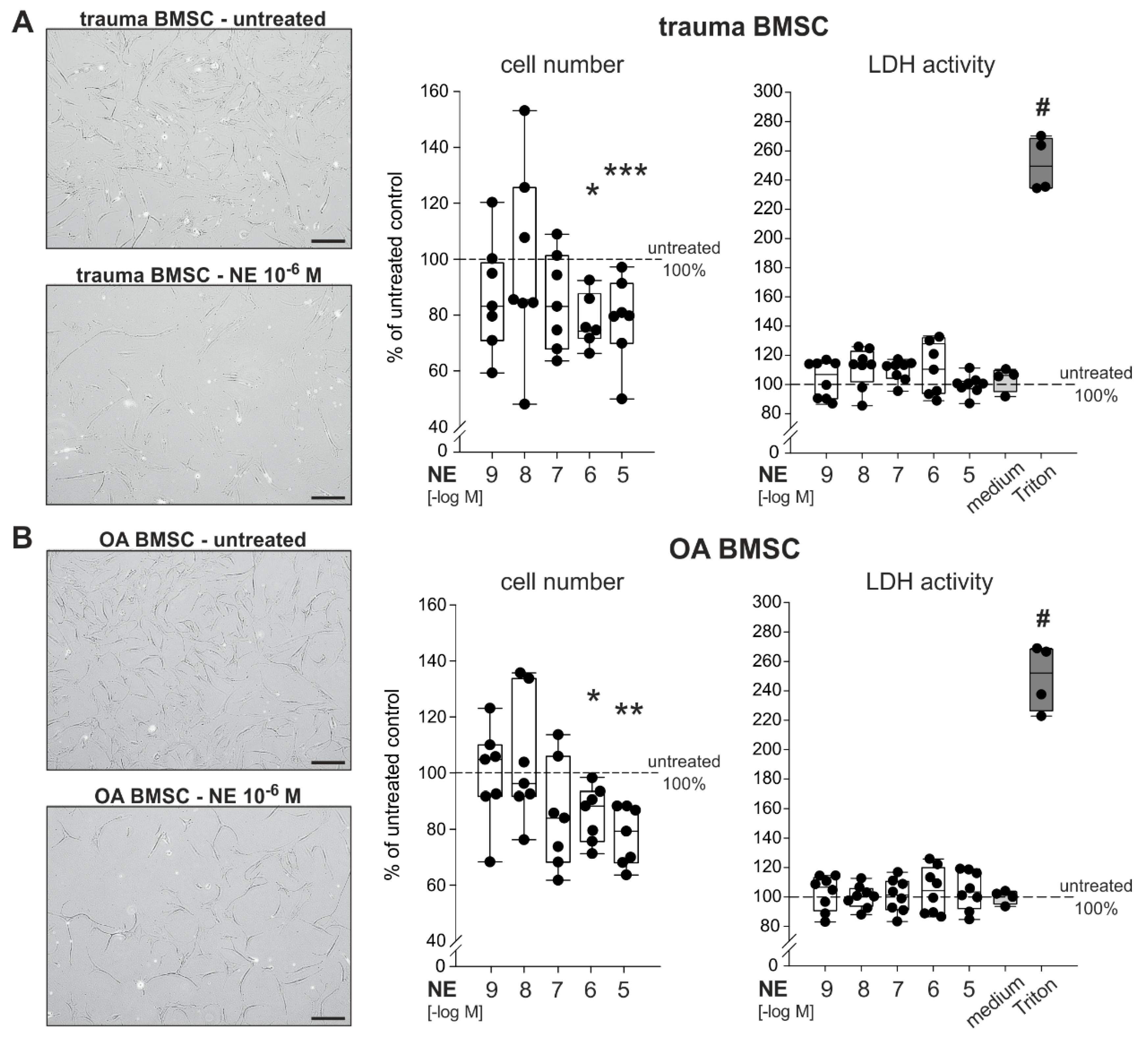
2.3. Effect of NE on Proliferation of Trauma and OA BMSCs
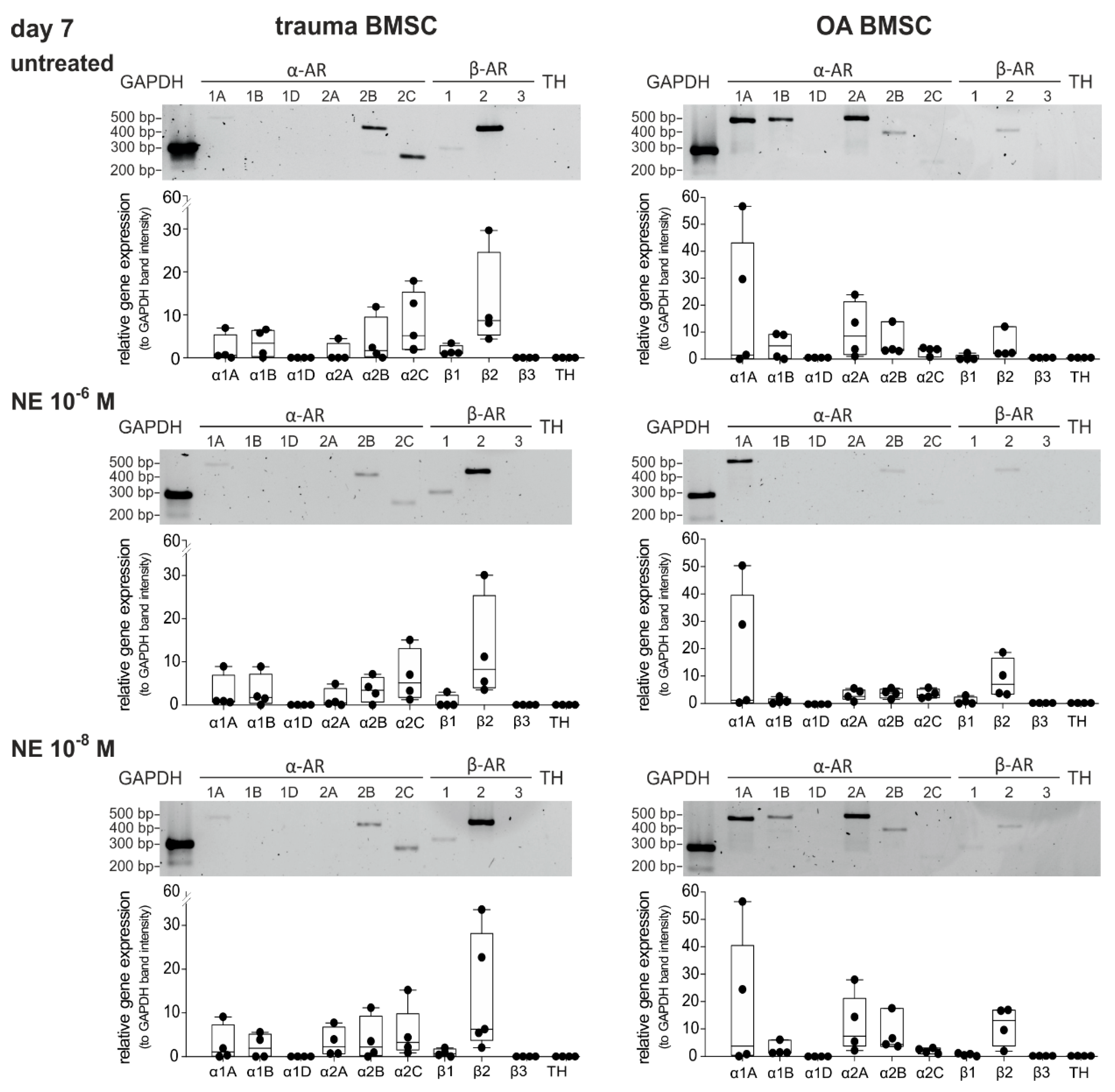
2.4. Changes in AR Gene Expression after Treatment with NE
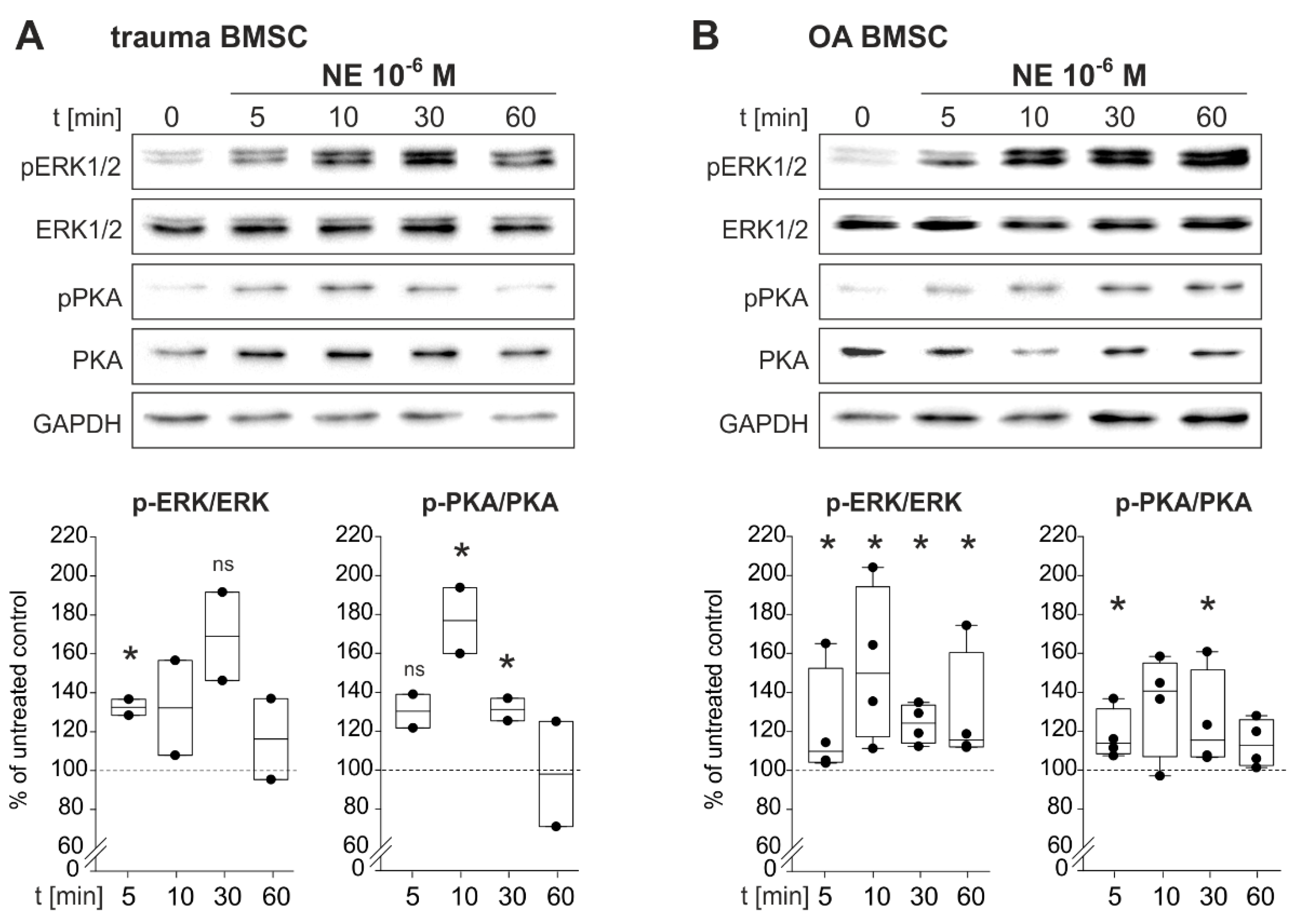
2.5. NE-Induced Intracellular Signaling
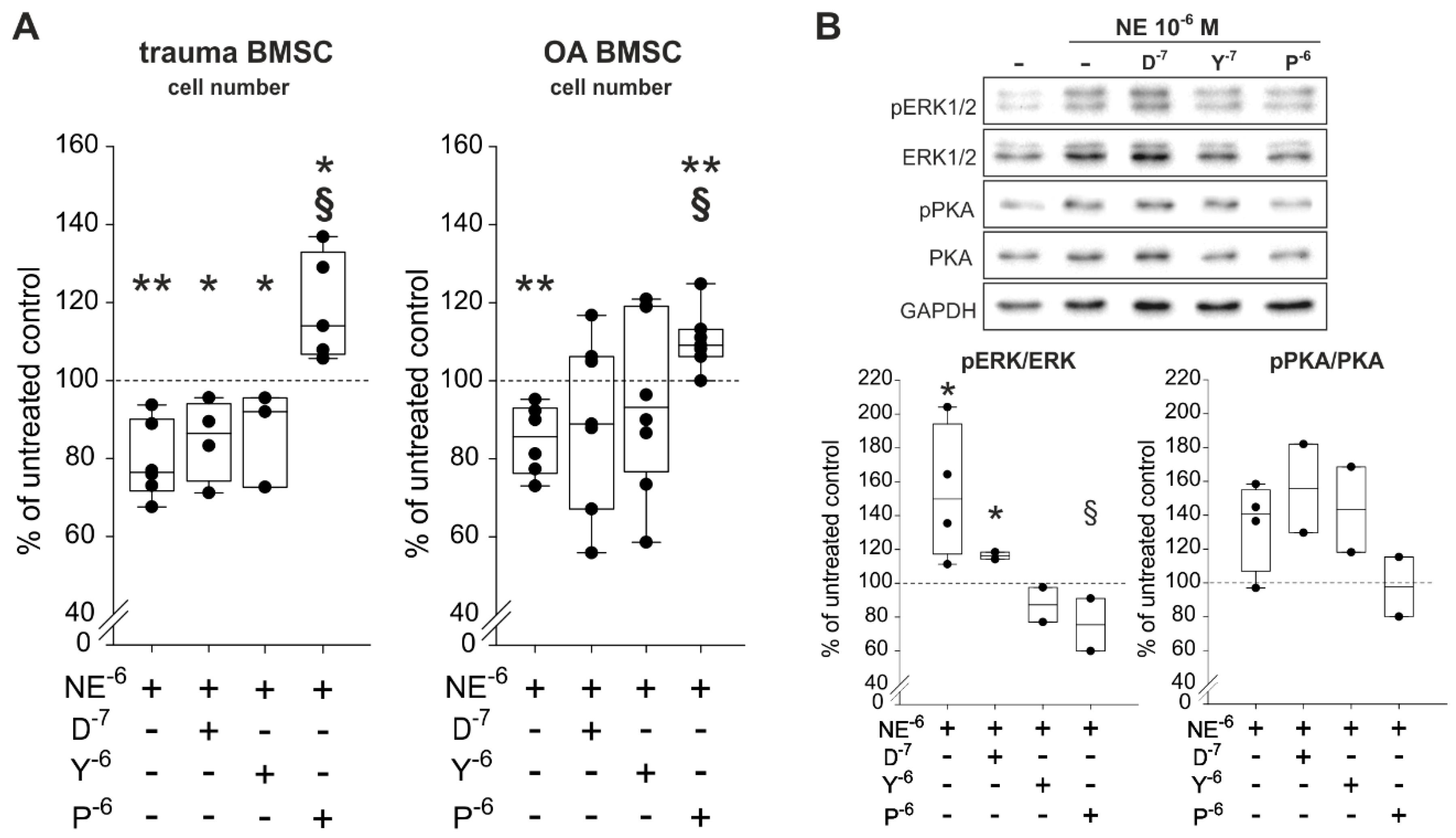
2.6. Reversal of NE-Mediated Effects by β2-AR-Antagonist
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Cell Culture
4.3. Proliferation Experiments
4.4. Determination of Cell Viability
4.5. Western Blot Analysis
4.6. RNA Isolation and cDNA Synthesis
4.7. Real-Time RT-PCR and Gel Electrophoresis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AR | Adrenergic receptor |
| BMSC cAMP | Bone marrow-derived stem cells Cyclic adenosine monophosphate |
| cDNA | Complementary desoxy ribonucleic acid |
| DMEM/F12 | Dulbecco’s modified eagle’s medium and Ham’s F-12 Medium |
| DPBS | Dulbecco’s phosphate buffered saline |
| ECL | Enhanced chemiluminescence |
| ERK | Extracellular signal-regulated kinases |
| FBS | Fetal bovine serum |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| HRP IL-1β LDH | Horse radish peroxidase Interleukin-1β Lactate dehydrogenase |
| mRNA | Messenger ribonucleic acid |
| MSC | Mesenchymal stem cells |
| NE | Norepinephrine |
| OA | Osteoarthritis |
| P/S | Penicillin streptomycin |
| PCR | Polymerase chain reaction |
| pERK | Phosphorylated extracellular signal-regulated kinases |
| PKA | Protein kinase A |
| pPKA | Phosphorylated protein kinase A |
| PVDF | Polyvinylidenfluorid |
| RT-PCR | Reverse-transcriptase PCR |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| TBST | Tris-buffered saline with Tween20 |
| TH | Tyrosine hydroxylase |
References
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudics, V.; Kunstár, A.; Kovács, J.; Lakatos, T.; Géher, P.; Gömör, B.; Monostori, E.; Uher, F. Chondrogenic potential of mesenchymal stem cells from patients with rheumatoid arthritis and osteoarthritis: Measurements in a microculture system. Cells Tissues Organs 2009, 189, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Koelling, S.; Kruegel, J.; Irmer, M.; Path, J.R.; Sadowski, B.; Miro, X.; Miosge, N. Migratory chondrogenic progenitor cells from repair tissue during the later stages of human osteoarthritis. Cell Stem Cell 2009, 4, 324–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, S.M.; Kalamegam, G.; Pushparaj, P.N.; Matta, C.; Memic, A.; Khademhosseini, A.; Mobasheri, R.; Poletti, F.L.; Hoyland, J.A.; Mobasheri, A. Mesenchymal stem cells in regenerative medicine: Focus on articular cartilage and intervertebral disc regeneration. Methods 2016, 99, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Granero-Molto, F.; Weis, J.A.; Longobardi, L.; Spagnoli, A. Role of mesenchymal stem cells in regenerative medicine: Application to bone and cartilage repair. Expert Opin. Biol. Ther. 2008, 8, 255–268. [Google Scholar] [CrossRef]
- Caplan, A.I.; Dennis, J.E. Mesenchymal stem cells as trophic mediators. J. Cell. Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef]
- Ha, C.-W.; Park, Y.-B.; Kim, S.H.; Lee, H.-J. Intra-articular Mesenchymal Stem Cells in Osteoarthritis of the Knee: A Systematic Review of Clinical Outcomes and Evidence of Cartilage Repair. Arthroscopy 2019, 35, 277–288.e2. [Google Scholar] [CrossRef]
- Liu, S.; Zhou, J.; Zhang, X.; Liu, Y.; Chen, J.; Hu, B.; Song, J.; Zhang, Y. Strategies to Optimize Adult Stem Cell Therapy for Tissue Regeneration. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.M.; Cho, H.H.; Lee, S.Y.; Hong, C.P.; Yang, J.W.; Kim, Y.S.; Suh, K.T.; Jung, J.S. Role of IRAK1 on TNF-induced proliferation and NF-ĸB activation in human bone marrow mesenchymal stem cells. Cell. Physiol. Biochem. 2012, 30, 49–60. [Google Scholar] [CrossRef]
- Gopi, I.K.; Rattan, S.I.S. Biphasic Dose-Response and Hormetic Effects of Stress Hormone Hydrocortisone on Telomerase-Immortalized Human Bone Marrow Stem Cells In Vitro. Dose Response 2019, 17, 1559325819889819. [Google Scholar] [CrossRef] [PubMed]
- Golan, K.; Kollet, O.; Markus, R.P.; Lapidot, T. Daily light and darkness onset and circadian rhythms metabolically synchronize hematopoietic stem cell differentiation and maintenance: The role of bone marrow norepinephrine, tumor necrosis factor, and melatonin cycles. Exp. Hematol. 2019, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenei-Lanzl, Z.; Grässel, S.; Pongratz, G.; Kees, F.; Miosge, N.; Angele, P.; Straub, R.H. Norepinephrine inhibition of mesenchymal stem cell and chondrogenic progenitor cell chondrogenesis and acceleration of chondrogenic hypertrophy. Arthritis Rheumatol. 2014, 66, 2472–2481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grässel, S.G. The role of peripheral nerve fibers and their neurotransmitters in cartilage and bone physiology and pathophysiology. Arthritis Res. Ther. 2014, 16, 485. [Google Scholar] [CrossRef] [Green Version]
- Kotova, P.D.; Sysoeva, V.Y.; Rogachevskaja, O.A.; Bystrova, M.F.; Kolesnikova, A.S.; Tyurin-Kuzmin, P.A.; Fadeeva, J.I.; Tkachuk, V.A.; Kolesnikov, S.S. Functional expression of adrenoreceptors in mesenchymal stromal cells derived from the human adipose tissue. Biochim. Biophys. Acta 2014, 1843, 1899–1908. [Google Scholar] [CrossRef] [Green Version]
- Hein, L. Adrenoceptors and signal transduction in neurons. Cell Tissue Res. 2006, 326, 541–551. [Google Scholar] [CrossRef]
- Cotecchia, S.; Kobilka, B.K.; Daniel, K.W.; Nolan, R.D.; Lapetina, E.Y.; Caron, M.G.; Lefkowitz, R.J.; Regan, J.W. Multiple second messenger pathways of alpha-adrenergic receptor subtypes expressed in eukaryotic cells. J. Biol. Chem. 1990, 265, 63–69. [Google Scholar]
- Lai, L.P.; Mitchell, J. Beta2-adrenergic receptors expressed on murine chondrocytes stimulate cellular growth and inhibit the expression of Indian hedgehog and collagen type X. J. Cell. Biochem. 2008, 104, 545–553. [Google Scholar] [CrossRef]
- Alblas, J.; van Corven, E.J.; Hordijk, P.L.; Milligan, G.; Moolenaar, W.H. Gi-mediated activation of the p21ras-mitogen-activated protein kinase pathway by alpha 2-adrenergic receptors expressed in fibroblasts. J. Biol. Chem. 1993, 268, 22235–22238. [Google Scholar]
- Lorenz, J.; Schäfer, N.; Bauer, R.; Jenei-Lanzl, Z.; Springorum, R.H.; Grässel, S. Norepinephrine modulates osteoarthritic chondrocyte metabolism and inflammatory responses. Osteoarthr. Cartil. 2016, 24, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Kodama, D.; Togari, A. Noradrenaline stimulates cell proliferation by suppressing potassium channels via G(i/o) -protein-coupled α(1B) -adrenoceptors in human osteoblasts. Br. J. Pharmacol. 2013, 168, 1230–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, J.; Lai, L.P.; Peralta, F.; Xu, Y.; Sugamori, K. β2-adrenergic receptors inhibit the expression of collagen type II in growth plate chondrocytes by stimulating the AP-1 factor Jun-B. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E633–E639. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.M.; Dixon, K.; Beck, S.; Fabian, D.; Feldman, A.; Barry, F. Reduced chondrogenic and adipogenic activity of mesenchymal stem cells from patients with advanced osteoarthritis. Arthritis Rheum. 2002, 46, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Suri, S.; Gill, S.E.; Massena de Camin, S.; Wilson, D.; McWilliams, D.F.; Walsh, D.A. Neurovascular invasion at the osteochondral junction and in osteophytes in osteoarthritis. Ann. Rheum. Dis. 2007, 66, 1423–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Zou, Z.; Zhu, C.; Deng, J.; Wang, J.; Ran, X.; Shi, C.; Ai, G.; Li, R.; Cheng, T.; et al. DNA synthesis of rat bone marrow mesenchymal stem cells through alpha1-adrenergic receptors. Arch. Biochem. Biophys. 2009, 490, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Blake, L.E.; Roux, J.; Hernando-Herraez, I.; Banovich, N.E.; Perez, R.G.; Hsiao, C.J.; Eres, I.; Cuevas, C.; Marques-Bonet, T.; Gilad, Y. A comparison of gene expression and DNA methylation patterns across tissues and species. Genome Res. 2020, 30, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Speichert, S.; Molotkov, N.; El Bagdadi, K.; Meurer, A.; Zaucke, F.; Jenei-Lanzl, Z. Role of Norepinephrine in IL-1β-Induced Chondrocyte Dedifferentiation under Physioxia. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, T.; Cannon, B.; Nedergaard, J. Differential adrenergic regulation of the gene expression of the beta-adrenoceptor subtypes beta1, beta2 and beta3 in brown adipocytes. Biochem. J. 2000, 347 Pt 3, 643–651. [Google Scholar] [CrossRef]
- Lei, B.; Zhang, Y.; Han, C. Sustained norepinephrine stimulation induces different regulation of expression in three alpha1-adrenoceptor subtypes. Life Sci. 2001, 69, 301–308. [Google Scholar] [CrossRef]
- Spector, S.; Gordon, R.; Sjoerdsma, A.; Udenfriend, S. End-product inhibition of tyrosine hydroxylase as a possible mechanism for regulation of norepinephrine synthesis. Mol. Pharmacol. 1967, 3, 549–555. [Google Scholar]
- Sokolove, J.; Lepus, C.M. Role of inflammation in the pathogenesis of osteoarthritis: Latest findings and interpretations. Ther. Adv. Musculoskelet. Dis. 2013, 5, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Dexheimer, V.; Mueller, S.; Braatz, F.; Richter, W. Reduced Reactivation from Dormancy but Maintained Lineage Choice of Human Mesenchymal Stem Cells with Donor Age. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Kido, A.; Yoshitani, K.; Shimizu, T.; Akahane, M.; Fujii, H.; Tsukamoto, S.; Kondo, Y.; Honoki, K.; Imano, M.; Tanaka, Y. Effect of mesenchymal stem cells on hypoxia-induced desensitization of β2-adrenergic receptors in rat osteosarcoma cells. Oncol. Lett. 2012, 4, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Haller, J.M.; McFadden, M.; Kubiak, E.N.; Higgins, T.F. Inflammatory cytokine response following acute tibial plateau fracture. J. Bone Joint Surg. Am. 2015, 97, 478–483. [Google Scholar] [CrossRef]
- Liu-Bryan, R.; Terkeltaub, R. Emerging regulators of the inflammatory process in osteoarthritis. Nat. Rev. Rheumatol. 2015, 11, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Jenei-Lanzl, Z.; Zwingenberg, J.; Lowin, T.; Anders, S.; Straub, R.H. Proinflammatory receptor switch from Gαs to Gαi signaling by β-arrestin-mediated PDE4 recruitment in mixed RA synovial cells. Brain Behav. Immun. 2015, 50, 266–274. [Google Scholar] [CrossRef]
- Wang, D.W.; Fermor, B.; Gimble, J.M.; Awad, H.A.; Guilak, F. Influence of oxygen on the proliferation and metabolism of adipose derived adult stem cells. J. Cell. Physiol. 2005, 204, 184–191. [Google Scholar] [CrossRef]
- Refsnes, M.; Thoresen, G.H.; Sandnes, D.; Dajani, O.F.; Dajani, L.; Christoffersen, T. Stimulatory and inhibitory effects of catecholamines on DNA synthesis in primary rat hepatocyte cultures: Role of alpha 1- and beta-adrenergic mechanisms. J. Cell. Physiol. 1992, 151, 164–171. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, C.; Chen, W.; Qiu, L.; Li, S.; Wang, T.; Xie, H.; Li, Y.; Li, C.; Li, L. The stress hormone norepinephrine promotes tumor progression through β2-adrenoreceptors in oral cancer. Arch. Oral Biol. 2020, 113, 104712. [Google Scholar] [CrossRef]
- Nilsson, M.B.; Le, X.; Heymach, J.V. β-Adrenergic Signaling in Lung Cancer: A Potential Role for Beta-Blockers. J. Neuroimmune Pharmacol. 2020, 15, 27–36. [Google Scholar] [CrossRef]
- Coelho, M.; Soares-Silva, C.; Brandão, D.; Marino, F.; Cosentino, M.; Ribeiro, L. β-Adrenergic modulation of cancer cell proliferation: Available evidence and clinical perspectives. J. Cancer Res. Clin. Oncol. 2017, 143, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Neaves, W.B. Normal stem cells and cancer stem cells: The niche matters. Cancer Res. 2006, 66, 4553–4557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronnikov, G.; Houstĕk, J.; Nedergaard, J. Beta-adrenergic, cAMP-mediated stimulation of proliferation of brown fat cells in primary culture. Mediation via beta 1 but not via beta 3 adrenoceptors. J. Biol. Chem. 1992, 267, 2006–2013. [Google Scholar] [PubMed]
- Dai, Z.; Li, Y.; Quarles, L.D.; Song, T.; Pan, W.; Zhou, H.; Xiao, Z. Resveratrol enhances proliferation and osteoblastic differentiation in human mesenchymal stem cells via ER-dependent ERK1/2 activation. Phytomedicine 2007, 14, 806–814. [Google Scholar] [CrossRef]
- Pullar, C.E.; Isseroff, R.R. The beta 2-adrenergic receptor activates pro-migratory and pro-proliferative pathways in dermal fibroblasts via divergent mechanisms. J. Cell Sci. 2006, 119, 592–602. [Google Scholar] [CrossRef] [Green Version]
- Tombes, R.M.; Auer, K.L.; Mikkelsen, R.; Valerie, K.; Wymann, M.P.; Marshall, C.J.; McMahon, M.; Dent, P. The mitogen-activated protein (MAP) kinase cascade can either stimulate or inhibit DNA synthesis in primary cultures of rat hepatocytes depending upon whether its activation is acute/phasic or chronic. Biochem. J. 1998, 330, 1451–1460. [Google Scholar] [CrossRef] [Green Version]
- Meloche, S.; Pouysségur, J. The ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1- to S-phase transition. Oncogene 2007, 26, 3227–3239. [Google Scholar] [CrossRef] [Green Version]
- Dumaz, N.; Marais, R. Integrating signals between cAMP and the RAS/RAF/MEK/ERK signalling pathways. Based on the anniversary prize of the Gesellschaft für Biochemie und Molekularbiologie Lecture delivered on 5 July 2003 at the Special FEBS Meeting in Brussels. FEBS J. 2005, 272, 3491–3504. [Google Scholar] [CrossRef]
- Stork, P.J.S.; Schmitt, J.M. Crosstalk between cAMP and MAP kinase signaling in the regulation of cell proliferation. Trends Cell Biol. 2002, 12, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Huang, Y.; Bu, D.; Liu, A.D.; Holmberg, L.; Jia, Y.; Tang, C.; Du, J.; Jin, H. Sulfur dioxide inhibits vascular smooth muscle cell proliferation via suppressing the Erk/MAP kinase pathway mediated by cAMP/PKA signaling. Cell Death Dis. 2014, 5, e1251. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- Jenei-Lanzl, Z.; Straub, R.H.; Dienstknecht, T.; Huber, M.; Hager, M.; Grässel, S.; Kujat, R.; Angele, M.K.; Nerlich, M.; Angele, P. Estradiol inhibits chondrogenic differentiation of mesenchymal stem cells via nonclassic signaling. Arthritis Rheum. 2010, 62, 1088–1096. [Google Scholar] [CrossRef]
- Anderson, D.E.; Markway, B.D.; Weekes, K.J.; McCarthy, H.E.; Johnstone, B. Physioxia Promotes the Articular Chondrocyte-Like Phenotype in Human Chondroprogenitor-Derived Self-Organized Tissue. Tissue Eng. Part A 2018, 24, 264–274. [Google Scholar] [CrossRef]
- Herrmann, M.; Anders, S.; Straub, R.H.; Jenei-Lanzl, Z. TNF inhibits catecholamine production from induced sympathetic neuron-like cells in rheumatoid arthritis and osteoarthritis in vitro. Sci. Rep. 2018, 8, 9645. [Google Scholar] [CrossRef]
- El Bagdadi, K.; Zaucke, F.; Meurer, A.; Straub, R.H.; Jenei-Lanzl, Z. Norepinephrine Inhibits Synovial Adipose Stem Cell Chondrogenesis via α2a-Adrenoceptor-Mediated ERK1/2 Activation. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Characteristics | Trauma BMSC Number/Mean/(%)/(Range) | OA BMSC Number/Mean/(%)/(Range) |
|---|---|---|
| total (number, %) | 14 (100%) | 17 (100%) |
| female/male (number, %) | 2/9 * (18.18%/81.82%) | 1 / 16 (5.88% / 94.12%) |
| age (years (mean ± stdd., (range))) | 34.08 ± 10.79 (23–51) | 66.44 ± 6.72 (52–76) |
| Antibody | Target Protein/Species/Antibody/Manufacture/Dilution |
|---|---|
| ERK | p44/42 MAPK (ERK1/2) (3A7)/mouse/mAb/Cell Signaling Technology®®/1:2500 |
| p-ERK | p-p44/42 MAPK (T202/ Y204)/rabbit/mAb/Cell Signaling Technology®®/1:2500 |
| PKA | ab32514, PKA R2, (Y116), 0,15 mg/mL/rabbit/mAb/Abcam/ 1:5000 |
| p-PKA | ab32390, PKA R2 (E151) (phospho S99), 0,351 mg/mL/rabbit/ mAb/Abcam/1:2500 |
| GAPDH | GAPDH Loading control antibody (GA1R)/mouse/mAb/ invitrogen by Thermo Fisher Scientific/1:2500 |
| anti-mouse | Polyclonal Rabbit Anti-Mouse Immunglobulins/HRP/Dako, Glostrup, Denmark/1:1000 |
| anti-rabbit | Polyclonal Swine Anti-Rabbit Immunglobulins/HRP/Dako, Glostrup, Denmark/1:1000 |
| Gene | NCBI Reference | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|---|
| GAPDH | NM_001289745.2 | CTC CTG TTC GAC AGT CAG CC | TTC CCG TTC TCA GCC TTG AC |
| ADRA1A | NM_000680.3 | CCA TGC TCC AGC CAA GAG TT | TCC TGT CCT AGA CTT CCT CCC |
| ADRA1B | NM_000679.3 | GTC CAC CGT CAT CTC CAT CG | GAA CAA GGA GCC AAG CGG TAG |
| ADRA1D | NM_000678.3 | TGA CTT TCC GCG ATC TCC TG | TTA CCT GCC ACG GCC ATA AG |
| ADRA2A | NM_000681.3 | TGG TCA TCG GAG TGT TCG TG | GCC CAC TAG GAA GAT GGC TC |
| ADRA2B | NM_000682.6 | GAC ATT TCA CCG GCA ACA CC | GGG ACT GAG AAC CAG GAA GC |
| ADRA2C | NM_000683.3 | CGA TGT GCT GTT TTG CAC CT | GGA TGT ACC AGG TCT CGT CG |
| ADRB1 | NM_000684.2 | TAG CAG GTG AAC TCG AAG CC | ATC TTC CAC TCC GGT CCT CT |
| ADRB2 | NM_000024.5 | CAG AGC CTG CTG ACC AAG AA | GCC TAA CGT CTT GAG GGC TT |
| ADRB3 | NM_000025.2 | GCC AAT TCT GCC TTCAAC CC | GCC AGA GGT TTT CCA CAG GT |
| TH | NM_000360.3 | CAG GCA GAG GCC ATC ATG T | GTG GTC CAA GTC CAG GTC AG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hedderich, J.; El Bagdadi, K.; Angele, P.; Grässel, S.; Meurer, A.; Straub, R.H.; Zaucke, F.; Jenei-Lanzl, Z. Norepinephrine Inhibits the Proliferation of Human Bone Marrow-Derived Mesenchymal Stem Cells via β2-Adrenoceptor-Mediated ERK1/2 and PKA Phosphorylation. Int. J. Mol. Sci. 2020, 21, 3924. https://doi.org/10.3390/ijms21113924
Hedderich J, El Bagdadi K, Angele P, Grässel S, Meurer A, Straub RH, Zaucke F, Jenei-Lanzl Z. Norepinephrine Inhibits the Proliferation of Human Bone Marrow-Derived Mesenchymal Stem Cells via β2-Adrenoceptor-Mediated ERK1/2 and PKA Phosphorylation. International Journal of Molecular Sciences. 2020; 21(11):3924. https://doi.org/10.3390/ijms21113924
Chicago/Turabian StyleHedderich, Jessica, Karima El Bagdadi, Peter Angele, Susanne Grässel, Andrea Meurer, Rainer H. Straub, Frank Zaucke, and Zsuzsa Jenei-Lanzl. 2020. "Norepinephrine Inhibits the Proliferation of Human Bone Marrow-Derived Mesenchymal Stem Cells via β2-Adrenoceptor-Mediated ERK1/2 and PKA Phosphorylation" International Journal of Molecular Sciences 21, no. 11: 3924. https://doi.org/10.3390/ijms21113924
APA StyleHedderich, J., El Bagdadi, K., Angele, P., Grässel, S., Meurer, A., Straub, R. H., Zaucke, F., & Jenei-Lanzl, Z. (2020). Norepinephrine Inhibits the Proliferation of Human Bone Marrow-Derived Mesenchymal Stem Cells via β2-Adrenoceptor-Mediated ERK1/2 and PKA Phosphorylation. International Journal of Molecular Sciences, 21(11), 3924. https://doi.org/10.3390/ijms21113924