Protein Glycation in Plants—An Under-Researched Field with Much Still to Discover



Abstract
:1. Protein Glycation in Plants: Three Papers Setting the Scene
2. Glycation: The Maillard Reaction
3. Glycation in Arabidopsis thaliana
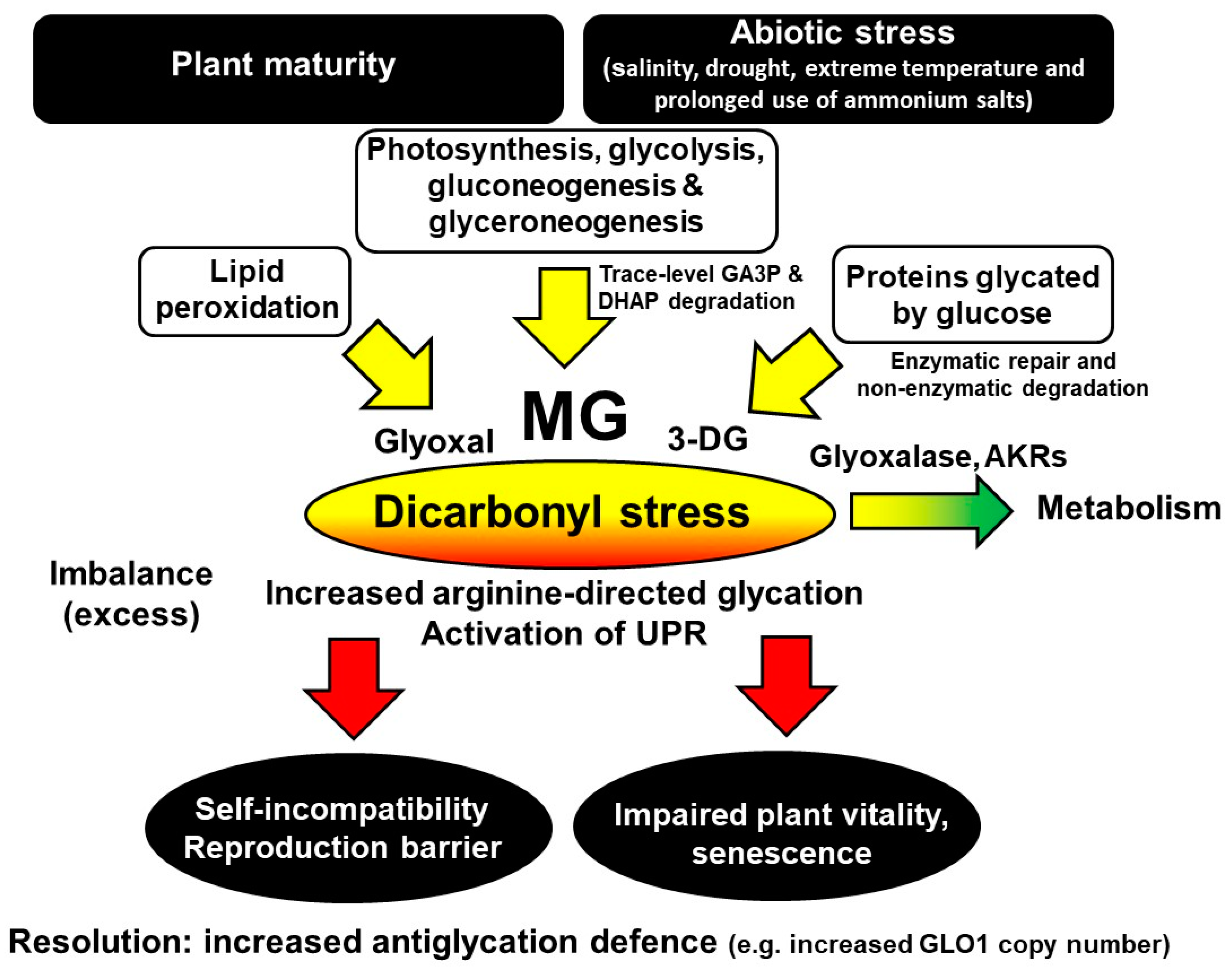
4. Dicarbonyl Stress in Plants
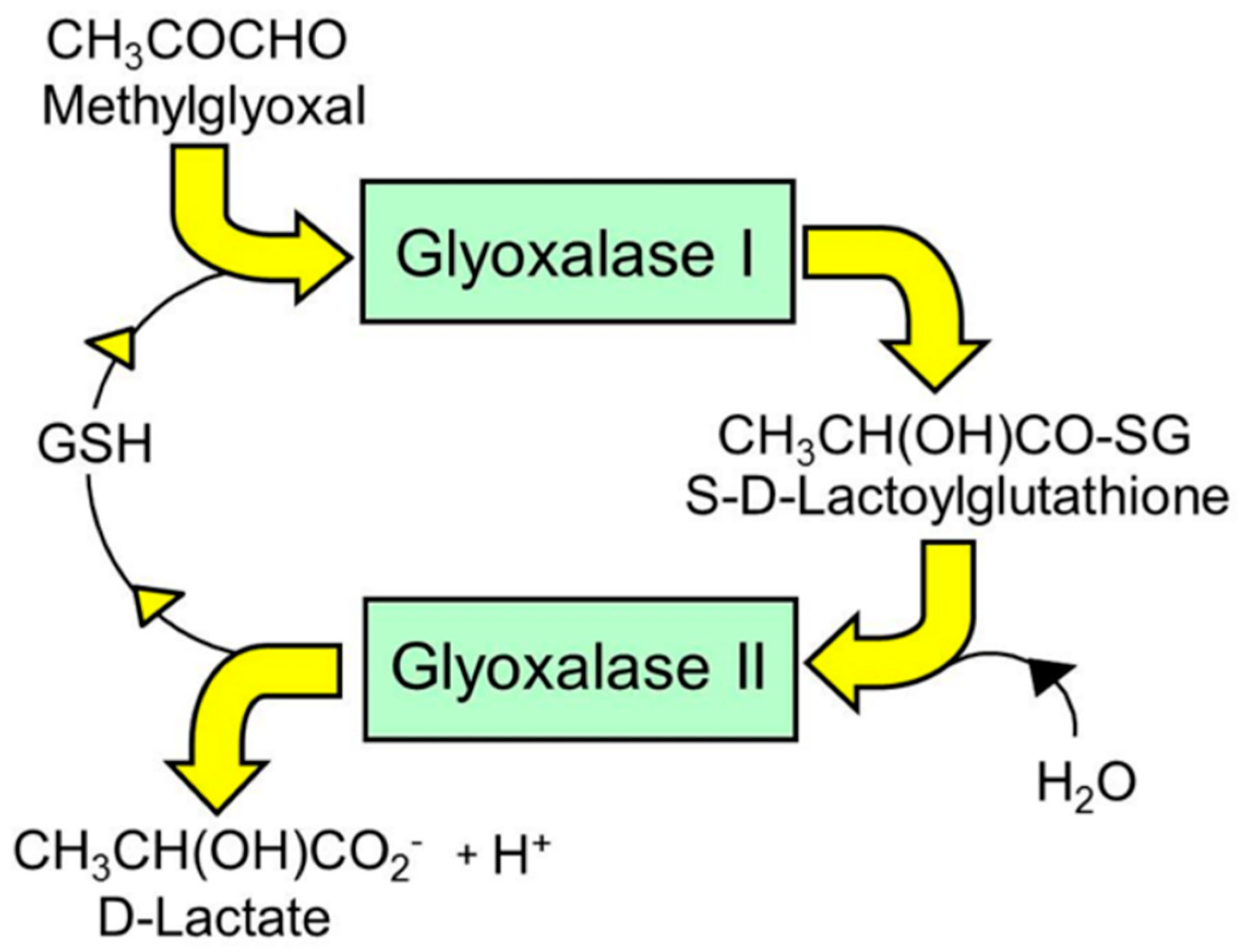
5. Enzymatic Defense Against Glycation—The Glyoxalase System and Aldoketo Reductases
6. Glycation in Plants—Considerations for Crops and Other Commercial Aspects
7. Why Is Glycation Potentially Damaging to Plants?
8. Role of Dicarbonyl Stress in the Unfolded Protein Response in Plants
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AGEs | advanced glycation endproducts; |
| AKR | aldoketoreductase; |
| CEL | Nε-(1-carboxyethyl)lysine; |
| CMA | Nω-carboxymethylarginine; |
| CML | Nε-carboxymethyl-lysine; |
| 3-DG | 3-deoxyglucosone; |
| DHAP | dihydroxyacetonephosphate; |
| ER | endoplasmic reticulum; |
| FL | Nε-fructosyl-lysine; |
| GA3P | glyceraldehyde-3-phosphate; |
| G-H1 | glyoxal-derived hydroimidazolone, Nδ-(5-hydro-4-imidazolon-2-yl)ornithine; |
| Glo1 | glyoxalase 1; |
| Glo2 | glyoxalase 2; |
| G6P | glucose-6-phosphate; |
| GSH | reduced glutathione; |
| HA | hemithioacetal; |
| HAS | human serum albumin; |
| LC-MS/MS | liquid chromatography-tandem mass spectrometry; |
| MG | methylglyoxal; |
| MG-H1 | methylglyoxal-derived hydroimidazolone, |
| Nδ-(5-hydro-5-methyl-4-imidazolon-2-yl)-ornithine; | |
| MOLD | methylglyoxal-derived lysine dimer, |
| 1,3-di(Nε-lysino)-4-methyl-imidazolium; | |
| OsAKR1 | aldoketo reductase isoform-1 of Oryza sativa; |
| RBD | receptor binding domain; |
| R5P | ribose-5-phosphate; |
| SLG | S-D-lactoylglutathione; |
| UPR | unfolded protein response. |
References
- Bechtold, U.; Rabbani, N.; Mullineaux, P.M.; Thornalley, P.J. Quantitative measurement of specific biomarkers for protein oxidation, nitration and glycation in Arabidopsis leaves. Plant J. 2009, 59, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Takagi, D.; Inoue, H.; Odawara, M.; Shimakawa, G.; Miyake, C. The Calvin Cycle Inevitably Produces Sugar-Derived Reactive Carbonyl Methylglyoxal During Photosynthesis: A Potential Cause of Plant Diabetes. Plant Cell Physiol. 2014, 55, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilova, T.; Paudel, G.; Shilyaev, N.; Schmidt, R.; Brauch, D.; Tarakhovskaya, E.; Milrud, S.; Smolikova, G.; Tissier, A.; Vogt, T.; et al. Global proteomic analysis of advanced glycation end products in the Arabidopsis proteome provides evidence for age-related glycation hot spots. J. Biol. Chem. 2017, 292, 15758–15776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soboleva, A.; Vikhnina, M.; Grishina, T.; Frolov, A. Probing Protein Glycation by Chromatography and Mass Spectrometry: Analysis of Glycation Adducts. Int. J. Mol. Sci. 2017, 18, 2557. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, N.; Thornalley, P.J. Dicarbonyl stress in cell and tissue dysfunction contributing to ageing and disease. Biochem. Biophys. Res. Commun. 2015, 458, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Irshad, Z.; Xue, M.; Ashour, A.; Larkin, J.R.; Thornalley, P.J.; Rabbani, N. Activation of the unfolded protein response in high glucose treated endothelial cells is mediated by methylglyoxal. Sci. Rep. 2019, 9, 7889. [Google Scholar] [CrossRef] [Green Version]
- Shumilina, J.; Kusnetsova, A.; Tsarev, A.; Janse van Rensburg, H.C.; Medvedev, S.; Demidchik, V.; Van den Ende, W.; Frolov, A. Glycation of Plant Proteins: Regulatory Roles and Interplay with Sugar Signalling? Int. J. Mol. Sci. 2019, 20, 2366. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, N.; Thornalley, P.J. Glycation research in Amino Acids: A place to call home. Amino Acids 2012, 42, 1087–1096. [Google Scholar] [CrossRef] [Green Version]
- Venkatraman, J.; Aggarwal, K.; Balaram, P. Helical peptide models for protein glycation: Proximity effects in catalysis of the Amadori rearrangement. Chem. Biol. 2001, 8, 611–625. [Google Scholar] [CrossRef] [Green Version]
- Fortpied, J.; Gemayel, R.; Stroobant, V.; Van Schaftingen, E. Plant ribulosamine/erythrulosamine 3-kinase, a putative protein-repair enzyme. Biochem. J. 2005, 388, 795–802. [Google Scholar] [CrossRef]
- Thornalley, P.J.; Langborg, A.; Minhas, H.S. Formation of glyoxal, methylglyoxal and 3-deoxyglucosone in the glycation of proteins by glucose. Biochem.J. 1999, 344, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Argirov, O.K.; Minhas, H.S.; Cordeiro, C.A.; Thornalley, P.J. Assay of advanced glycation endproducts (AGEs): Surveying AGEs by chromatographic assay with derivatisation by aminoquinolyl-N-hydroxysuccimidyl-carbamate and application to Nε-carboxymethyl-lysine- and Nε-(1-carboxyethyl)lysine-modified albumin. Biochem. J. 2002, 364, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Thornalley, P.J.; Battah, S.; Ahmed, N.; Karachalias, N.; Agalou, S.; Babaei-Jadidi, R.; Dawnay, A. Quantitative screening of advanced glycation endproducts in cellular and extracellular proteins by tandem mass spectrometry. Biochem. J. 2003, 375, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Glomb, M.A.; Lang, G. Isolation and characterization of glyoxal-arginine modifications. J. Agric. Food Chem. 2001, 49, 1493–1501. [Google Scholar] [CrossRef]
- Ahmed, M.U.; Thorpe, S.R.; Baynes, J.W. Identification of Nε-carboxymethyl-lysine as a degradation product of fructoselysine in glycated protein. J. Biol. Chem. 1986, 261, 4889–4894. [Google Scholar]
- Dunn, J.A.; Ahmed, M.U.; Murtiashaw, M.H.; Richardson, J.M.; Walla, M.D.; Thorpe, S.R.; Baynes, J.W. Reaction of ascorbate with lysine and protein under autoxidising conditions: Formation of Nε(carboxymethyl)lysine by reaction between lysine and products of autoxidation of ascorbate. Biochemistry 1990, 29, 10964–10970. [Google Scholar] [CrossRef]
- Phillips, S.A.; Thornalley, P.J. The formation of methylglyoxal from triose phosphates. Investigation using a specific assay for methylglyoxal. Eur. J. Biochem. 1993, 212, 101–105. [Google Scholar] [CrossRef]
- Szwergold, B.S.; Kappler, F.; Brown, T.R. Identification of fructose-3-phosphate in the lens of diabetic rats. Science 1990, 247, 451–454. [Google Scholar] [CrossRef]
- Paudel, G.; Bilova, T.; Schmidt, R.; Greifenhagen, U.; Berger, R.; Tarakhovskaya, E.; Stockhardt, S.; Balcke, G.U.; Humbeck, K.; Brandt, W.; et al. Osmotic stress is accompanied by protein glycation in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 6283–6295. [Google Scholar] [CrossRef] [Green Version]
- Matamoros, M.A.; Kim, A.; Penuelas, M.; Ihling, C.; Griesser, E.; Hoffmann, R.; Fedorova, M.; Frolov, A.; Becana, M. Protein Carbonylation and Glycation in Legume Nodules. Plant Physiol. 2018, 177, 1510–1528. [Google Scholar] [CrossRef] [Green Version]
- Leonova, T.; Popova, V.; Tsarev, A.; Henning, C.; Antonova, K.; Rogovskaya, N.; Vikhnina, M.; Baldensperger, T.; Soboleva, A.; Dinastia, E.; et al. Does Protein Glycation Impact on the Drought-Related Changes in Metabolism and Nutritional Properties of Mature Pea (Pisum sativum L.) Seeds? Int. J. Mol. Sci. 2020, 21, 567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaplin, A.K.; Chernukhin, I.; Bechtold, U. Profiling of advanced glycation end products uncovers abiotic stress-specific target proteins in Arabidopsis. J. Exp. Bot. 2019, 70, 653–670. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Ejiri, Y.; Tanaka, K. Glycation by ascorbic acid causes loss of activity of ribulose-1,5-bisphosphate carboxylase/oxygenase and its increased susceptibility to proteases. Plant Cell Physiol. 2002, 43, 1334–1341. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, N.; Thornalley, P.J. Measurement of methylglyoxal by stable isotopic dilution analysis LC-MS/MS with corroborative prediction in physiological samples. Nat. Protoc. 2014, 9, 1969–1979. [Google Scholar] [CrossRef]
- Ramaswamy, O.; Pal, S.; Guha-Mukerjee, S.; Sopory, S.K. Presence of glyoxalase I in pea. Biochem. Int. 1983, 3, 307–318. [Google Scholar]
- Turoczy, Z.; Kis, P.; Török, K.; Cserháti, M.; Lendvai, A.; Dudits, D.; Horváth, G. Overproduction of a rice aldo–keto reductase increases oxidative and heat stress tolerance by malondialdehyde and methylglyoxal detoxification. Plant Mol. Biol. 2011, 75, 399–412. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Ejiri, Y.; Toyoda, Y.; Tanaka, K. Identification and Biochemical Characterization of Plant Acylamino Acid-Releasing Enzyme. J. Biochem. 2003, 134, 251–257. [Google Scholar] [CrossRef]
- Lo, T.W.C.; Westwood, M.E.; McLellan, A.C.; Selwood, T.; Thornalley, P.J. Binding and modification of proteins by methylglyoxal under physiological conditions. A kinetic and mechanistic study with Na-acetylarginine, Na-acetylcysteine, Na-acetyl-lysine, and bovine serum albumin. J. Biol. Chem. 1994, 269, 32299–32305. [Google Scholar]
- Murthy, U.M.N.; Sun, W.Q. Protein modification by Amadori and Maillard reactions during seed storage: Roles of sugar hydrolysis and lipid peroxidation. J. Exp. Bot. 2000, 51, 1221–1228. [Google Scholar] [CrossRef]
- Singla-Pareek, S.L.; Reddy, M.K.; Sopory, S.K. Genetic engineering of the glyoxalase pathway in tobacco leads to enhanced salinity tolerance. Proc. Natl. Acad. Sci. USA 2003, 100, 14672–14677. [Google Scholar] [CrossRef] [Green Version]
- Thornalley, P.J. The enzymatic defence against glycation in health, disease and therapeutics: A symposium to examine the concept. Biochem. Soc. Trans. 2003, 31, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Loidl-Stahlhofen, A.; Spiteller, G. alpha-Hydroxyaldehydes, products of lipid peroxidation. Biochim. Biophys. Acta 1994, 1211, 156–160. [Google Scholar] [CrossRef]
- Rabbani, N.; Thornalley, P.J. Dicarbonyls (Glyoxal, Methylglyoxal, and 3-Deoxyglucosone). In Uremic Toxins; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 177–192. [Google Scholar]
- Delpierre, G.; Rider, M.H.; Collard, F.; Stroobant, V.; Vanstapel, F.; Santos, H.; Van Schaftingen, E. Identification, cloning, and heterologous expression of a mammalian fructosamine-3-kinase. Diabetes 2000, 49, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- Delpierre, G.; Vertommen, D.; Communi, D.; Rider, M.H.; Van Schaftingen, E. Identification of fructosamine residues deglycated by fructosamine-3-kinase in human hemoglobin. J. Biol. Chem. 2004, 279, 27613–27620. [Google Scholar] [CrossRef] [Green Version]
- Biemel, K.M.; Friedl, D.A.; Lederer, M.O. Identification and quantification of major Maillard cross-links in human serum albumin and lens protein—Evidence for glucosepane as the dominant compound. J. Biol. Chem. 2002, 277, 24907–24915. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zhao, Y.; Niu, Y.; Wang, C.; Wang, M.; Li, Y.; Sun, C. Activated glucose-6-phosphate dehydrogenase is associated with insulin resistance by upregulating pentose and pentosidine in diet-induced obesity of rats. Horm. Metab. Res. 2012, 44, 938–942. [Google Scholar] [CrossRef]
- Borysiuk, K.; Ostaszewska-Bugajska, M.; Vaultier, M.N.; Hasenfratz-Sauder, M.P.; Szal, B. Enhanced Formation of Methylglyoxal-Derived Advanced Glycation End Products in Arabidopsis Under Ammonium Nutrition. Front. Plant Sci. 2018, 9, 14. [Google Scholar] [CrossRef]
- Rabbani, N.; Shaheen, F.; Anwar, A.; Masania, J.; Thornalley, P.J. Assay of methylglyoxal-derived protein and nucleotide AGEs. Biochem. Soc. Trans. 2014, 42, 511–517. [Google Scholar] [CrossRef]
- Rabbani, N.; Thornalley, P.J. Reading patterns of proteome damage by glycation, oxidation and nitration: Quantitation by stable isotopic dilution analysis LC-MS/MS. Essays Biochem. 2020, 64, 169–183. [Google Scholar] [CrossRef]
- Bechtold, U.; Murphy, D.J.; Mullineaux, P.M. Arabidopsis peptide methionine sulfoxide reductase2 prevents cellular oxidative damage in long nights. Plant Cell 2004, 16, 908–919. [Google Scholar] [CrossRef] [Green Version]
- Conklin, P.L.; Pallanca, J.E.; Last, R.L.; Smirnoff, N. L-ascorbic acid metabolism in the ascorbate-deficient Arabidopsis mutant vtc1. Plant Physiol. 1997, 115, 1277–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbani, N.; Ashour, A.; Thornalley, P.J. Mass spectrometric determination of early and advanced glycation in biology. Glycoconj. J. 2016, 33, 553–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, N.; Thornalley, P.J. Chromatographic assay of glycation adducts in human serum albumin glycated in vitro by derivatisation with aminoquinolyl-N-hydroxysuccimidyl-carbamate and intrinsic fluorescence. Biochem.J. 2002, 364, 15–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornalley, P.J. Monosaccharide autoxidation in health and disease. Environ. Health Prospect. 1985, 64, 297–307. [Google Scholar] [CrossRef]
- Bechtold, U. Evidence of advanced glycation endproducts, oxidation and nitrosation damage in Arabidopsis leaves. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, S56. [Google Scholar] [CrossRef]
- Yadav, S.K.; Singla-Pareek, S.L.; Ray, M.; Reddy, M.K.; Sopory, S.K. Methylglyoxal levels in plants under salinity stress are dependent on glyoxalase I and glutathione. Biochem. Biophys. Res. Commun. 2005, 337, 61–67. [Google Scholar] [CrossRef]
- McLellan, A.C.; Phillips, S.A.; Thornalley, P.J. The assay of methylglyoxal in biological systems by derivatization with 1,2-diamino-4,5-dimethoxybenzene. Anal. Biochem. 1992, 206, 17–23. [Google Scholar] [CrossRef]
- Viškelienė, A.; Samuoliene, G.; Karklelienė, R.; Viskelis, P.; Sasnauskas, A.; Duchovskis, P. Quality and developmental changes in white head cabbage (Brassica oleracea L.) and radish (Raphanus sativus L.) during winter storage. Zemdirb. Agric. 2017, 104, 229–234. [Google Scholar] [CrossRef]
- Morcos, M.; Du, X.; Pfisterer, F.; Hutter, H.; Sayed, A.A.R.; Thornalley, P.; Ahmed, N.; Baynes, J.; Thorpe, S.; Kukudov, G.; et al. Glyoxalase-1 prevents mitochondrial protein modification and enhances lifespan in Caenorhabditis elegans. Aging Cell 2008, 7, 260–269. [Google Scholar] [CrossRef]
- Schlotterer, A.; Kukudov, G.; Bozorgmehr, F.; Hutter, H.; Du, X.L.; Oikonomou, D.; Ibrahim, Y.; Pfisterer, F.; Rabbani, N.; Thornalley, P.; et al. C-elegans as Model for the Study of High Glucose-Mediated Life Span Reduction. Diabetes 2009, 58, 2450–2456. [Google Scholar] [CrossRef] [Green Version]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Review: Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. 2016, 248, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbani, N.; Xue, M.; Thornalley, P.J. Methylglyoxal-induced dicarbonyl stress in aging and disease: First steps towards glyoxalase 1-based treatments. Clin. Sci. 2016, 130, 1677–1696. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Singla-Pareek, S.L.; Sopory, S.K. Glyoxalase and Methylglyoxal as Biomarkers for Plant Stress Tolerance. Crit. Rev. Plant Sci. 2014, 33, 429–456. [Google Scholar] [CrossRef]
- Sankaranarayanan, S.; Jamshed, M.; Kumar, A.; Skori, L.; Scandola, S.; Wang, T.; Spiegel, D.; Samuel, M.A. Glyoxalase Goes Green: The Expanding Roles of Glyoxalase in Plants. Int. J. Mol. Sci. 2017, 18, 898. [Google Scholar] [CrossRef]
- Ghosh, A.; Kushwaha, H.R.; Hasan, M.R.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Presence of unique glyoxalase III proteins in plants indicates the existence of shorter route for methylglyoxal detoxification. Sci. Rep. 2016, 6, 18358. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, N.; Xue, M.; Thornalley, P.J. Activity, regulation, copy number and function in the glyoxalase system. Biochem. Soc. Trans. 2014, 42, 419–424. [Google Scholar] [CrossRef]
- Jain, M.; Batth, R.; Kumari, S.; Mustafiz, A. Arabidopsis thaliana Contains Both Ni2+ and Zn2+ Dependent Glyoxalase I Enzymes and Ectopic Expression of the Latter Contributes More towards Abiotic Stress Tolerance in E. coli. PLoS ONE 2016, 11, e0159348. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Herrmann, C.J.; Simonovic, M.; Szklarczyk, D.; von Mering, C. Version 4.0 of PaxDb: Protein abundance data, integrated across model organisms, tissues, and cell-lines. Proteomics 2015, 15, 3163–3168. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, A.; Iwasa, M.; Nakabayashi, R.; Kobayashi, M.; Nishizawa, T.; Okazaki, Y.; Saito, K.; Kusano, M. Effects of Combined Low Glutathione with Mild Oxidative and Low Phosphorus Stress on the Metabolism of Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1464. [Google Scholar] [CrossRef] [Green Version]
- Vander Jagt, D.L.; Daub, F.; Krohn, J.A.; Han, L.P.B. Effects of pH and thiols on the kinetics of yeast glyoxalase I. An evaluation of the random pathway mechanism. Biochemistry 1975, 14, 3669–3675. [Google Scholar] [CrossRef]
- Proietti, S.; Falconieri, G.S.; Bertini, L.; Baccelli, I.; Paccosi, E.; Belardo, A.; Timperio, A.M.; Caruso, C. GLYI4 Plays A Role in Methylglyoxal Detoxification and Jasmonate-Mediated Stress Responses in Arabidopsis thaliana. Biomolecules 2019, 9, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramaswamy, O.; Pal, S.; Guha-Mukerjee, S.; Sopory, S.K. Correlation of glyoxalase I activity with cell proliferation in Datura callus culture. Plant Cell Rep. 1984, 3, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Mustafiz, A.; Singh, A.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Genome-wide analysis of rice and Arabidopsis identifies two glyoxalase genes that are highly expressed in abiotic stresses. Funct. Integr. Genom. 2011, 11, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Shimakawa, G.; Ifuku, K.; Suzuki, Y.; Makino, A.; Ishizaki, K.; Fukayama, H.; Morita, R.; Sakamoto, K.; Nishi, A.; Miyake, C. Responses of the chloroplast glyoxalase system to high CO2 concentrations. Biosci. Biotechnol. Biochem. 2018, 82, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Giacomelli, L.; Rudella, A.; van Wijk, K.J. High Light Response of the Thylakoid Proteome in Arabidopsis Wild Type and the Ascorbate-Deficient Mutant vtc2-2. A Comparative Proteomics Study. Plant Physiol. 2006, 141, 685–701. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, D.; Naik, D.; Reddy, A.R. Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plant metabolic processes and stress defense: A structure-function update. J. Plant Physiol. 2015, 179, 40–55. [Google Scholar] [CrossRef]
- Wani, S.H.; Thakur, A.K.; Jeshima, Y. Brassica Improvement: Molecular, Genetics and Genomic Perspectives; Springer: Heidelberg, Germany, 2020; p. 224. [Google Scholar]
- Muñoz-Sanz, J.V.; Zuriaga, E.; Cruz-García, F.; McClure, B.; Romero, C. Self-(In)compatibility Systems: Target Traits for Crop-Production, Plant Breeding, and Biotechnology. Front. Plant Sci. 2020, 11, 195. [Google Scholar] [CrossRef] [Green Version]
- Sankaranarayanan, S.; Jamshed, M.; Samuel, M. Degradation of glyoxalase I in Brassica napus stigma leads to self-incompatibility response. Nat. Plants 2015, 1, 1–7. [Google Scholar] [CrossRef]
- Azibi, T.; Hadj-Arab, H.; Lodé, M.; Ferreira de Carvalho, J.; Trotoux, G.; Nègre, S.; Gilet, M.-M.; Boutte, J.; Lucas, J.; Vekemans, X.; et al. Impact of whole genome triplication on the evolutionary history and the functional dynamics of regulatory genes involved in Brassica self-incompatibility signalling pathway. Plant Reprod. 2020, 33, 43–58. [Google Scholar] [CrossRef]
- Frahm, G.E.; Smith, D.G.S.; Kane, A.; Lorbetskie, B.; Cyr, T.D.; Girard, M.; Johnston, M.J.W. Determination of Supplier-to-Supplier and Lot-to-Lot Variability in Glycation of Recombinant Human Serum Albumin Expressed in Oryza sativa. PLoS ONE 2014, 9, e109893. [Google Scholar] [CrossRef]
- Chan, H.-T.; Daniell, H. Plant-made oral vaccines against human infectious diseases—Are we there yet? Plant Biotechnol. J. 2015, 13, 1056–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, N.; Dobler, D.; Dean, M.; Thornalley, P.J. Peptide mapping identifies hotspot site of modification in human serum albumin by methylglyoxal involved in ligand binding and esterase activity. J. Biol. Chem. 2005, 280, 5724–5732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbani, N.; Thornalley, P.J. Methylglyoxal, glyoxalase 1 and the dicarbonyl proteome. Amino Acids 2012, 42, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Gallet, X.; Charloteaux, B.; Thomas, A.; Braseur, R. A fast method to predict protein interaction sites from sequences. J. Mol. Biol. 2000, 302, 917–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.P.; Holmgren, A.; Larsson, N.G.; Halliwell, B.; Chang, C.J.; Kalyanaraman, B.; Rhee, S.G.; Thornalley, P.J.; Partridge, L.; Gems, D.; et al. Unraveling the Biological Roles of Reactive Oxygen Species. Cell Metab. 2011, 13, 361–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winterbourn, C.C. Reconciling the chemistry and biology of reactive oxygen species. Nat. Chem. Biol. 2008, 4, 278–286. [Google Scholar] [CrossRef]
- Doubnerova, V.; Ryslava, H. Roles of HSP70 in Plant Abiotic Stress; CRC Press: Boca Raton, FL, USA, 2014; pp. 44–66. [Google Scholar]
- Afrin, T.; Diwan, D.; Sahawneh, K.; Pajerowska-Mukhtar, K. Multilevel regulation of endoplasmic reticulum stress responses in plants: Where old roads and new paths meet. J. Exp. Bot. 2019, 71, 1659–1667. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Glycating Agent | Comment |
 Glyoxal | Formed by the degradation of reducing sugars, glycated proteins, nucleotides and lipid peroxidation [11,32]. Metabolized by the glyoxalase system [5]. Glyoxal is present in solution mainly as mono- and di-hydrates [33]. |
 Methylglyoxal (MG) | Formed mainly by the trace-level degradation of GA3P and DHAP [17]. Relatively high flux reactive dicarbonyl. Metabolized by the glyoxalase system [5]. MG is present in solution mainly as mono- and di-hydrates [33]. Precursor of the major AGE, MG-H1. |
 3-Deoxyglucosone (3-DG) | Formed by the degradation of reducing sugars and glycated proteins. Additionally formed by the enzymatic repair of FL [34]. Metabolized by aldoketo reductases [5]. 3-DG is present in solution as a complex mixture of cyclic hemiacetals and hemiketals [33]. |
| Glycation Adduct | Comment |
 | Early-stage glycation adduct [1]. Formed from glucose non-enzymatically and exposure to increased glucose concentration. Repaired intracellularly by fructosamine 3-phosphokinase [35]. |
 | A major quantitative arginine-derived AGE formed from MG. Influenced by the rate of the formation of MG, rate of metabolism of MG by Glo1 of the glyoxalase system and cellular proteolysis. Major AGE in Arabidopsis thaliana. Implicated in protein misfolding and, in excess, activation of the UPR. |
 | A major quantitative lysine-derived AGE. Formed by the oxidative degradation of FL (major), glycation by glyoxal and by ascorbic acid (usually minor). Increased by light stress in Arabidopsis thaliana. The CML/FL ratio is a marker of oxidative stress. |
 | Major quantitative crosslink formed in protein glycation [36]. Produced from the degradation of FL residues with a proximate arginine residue. Content in plant proteins is unknown. |
 | Low-level pentose sugar-derived glycation crosslink and intense fluorophore. Considered to reflect pentosephosphate pathway activity [37]. |
| Growth Condition | Effect on Protein Glycation of Glycating | Reference |
|---|---|---|
| Daylight to dark growth cycle | Early glycation adduct, FL: 3 mmol/mol lys (daylight entry), increasing to 10 mmol/mol lys (dark entry). | [1] |
| Diurnal period, heat, light and drought | Glycation adducts detected: CML, CMA, FL, G-H1 and MG-H1. Protein targets: a core group of 112 proteins, including chloroplast ATP synthase (β-subunit) and phosphoglycerate kinase. Glycated protein abundances were similar in heat, light and drought stresses. Glycated proteins with altered abundance were: light stress—2 (RPI3 and TPI, decreased); heat stress—1 (TPI, decreased); diurnal variation—8 (ASP5, FTSH2 and RAN3, increased; AOC2, BAS1, CORI3, OASB, PRK, PRXQ and PURA, decreased); and drought stress 17 (A2, GSA2 and P83484, increased; CAT2, CICDH, CTIMC, CYP18-4, FBP, GGAT1, GLU1, LOX2, P25697, PER34, RBCS-1A, RBCS-3B, TGG2 and TL29, decreased). | [22] |
| Excess light stress | AGEs increased: G-H1 (0.14 to 0.35 mmol/mol arg) and CML (0.77 to 1.65 mmol/mol lys). | [1] |
| Osmotic stress | Major glycation adducts detected: CML, CMA and G-H1; 785 glycation sites detected on 724 proteins—33 and 62 glycation sites were unique for control and osmotically stressed plants, respectively. Abundance changes of AGE-modified proteins under osmotic stress (range—2-fold decrease to 27-fold increase): 12 proteins involved in lipid metabolism, DNA supercoils and methylation; protein ubiquitination and degradation; energy metabolism; cell organization and development; cell wall formation; and the regulation of transcription and stress. | [19] |
| Ammonium NH4+ salts | MG-H1 and CEL-modified proteins detected by immunoblotting and immunoassays; 15% increase in CEL in ammonium NH4+-grown plants compared with those in nitrate NO3−-grown control plants. | [38] |
| Days Post-Sowing | Plant Appearance | Dicarbonyl Metabolite (nmol/g Fresh Weight; Mean ± SD, n = 6) | ||
|---|---|---|---|---|
| Glyoxal | MG | 3-DG | ||
| 6 |  | 0.38 ± 0.04 | 2.90 ± 0.81 | 0.76 ± 0.29 |
| 30 |  | 0.46 ± 0.12 | 3.47 ± 1.21 | 1.80 ± 1.05 * |
| 65 |  | 0.81 ± 0.32 **,OO | 4.08 ± 0.27 * | 0.49 ± 0.23 O |
| Amino Acid | Count | ||||
|---|---|---|---|---|---|
| Proteome | RBD | % AA in Proteome | % AA in RBD | Fold Enrichment | |
| Ala | 463,770 | 25,941 | 6.5 | 3.3 | 0.5 |
| Arg | 380,640 | 150,922 | 5.3 | 19.5 | 3.7 |
| Asn | 317,995 | 44,745 | 4.4 | 5.8 | 1.3 |
| Asp | 384,200 | 52,528 | 5.3 | 6.8 | 1.3 |
| Cys | 130,271 | 10,915 | 1.8 | 1.4 | 0.8 |
| Gln | 250,179 | 38,180 | 3.5 | 4.9 | 1.4 |
| Glu | 474,124 | 70,661 | 6.6 | 9.1 | 1.4 |
| Gly | 473,373 | 30,225 | 6.6 | 3.9 | 0.6 |
| His | 160,243 | 20,712 | 2.2 | 2.7 | 1.2 |
| Ile | 392,264 | 8682 | 5.5 | 1.1 | 0.2 |
| Leu | 697,276 | 28,075 | 9.7 | 3.6 | 0.4 |
| Lys | 449,328 | 101,031 | 6.3 | 13.0 | 2.1 |
| Met | 164,360 | 11,802 | 2.3 | 1.5 | 0.7 |
| Phe | 314,311 | 8387 | 4.4 | 1.1 | 0.2 |
| Pro | 341,009 | 29,637 | 4.7 | 3.8 | 0.8 |
| Ser | 636,209 | 67,405 | 8.9 | 8.7 | 1.0 |
| Thr | 369,142 | 36,395 | 5.1 | 4.7 | 0.9 |
| Trp | 90,588 | 5539 | 1.3 | 0.7 | 0.6 |
| Tyr | 209,664 | 17,971 | 2.9 | 2.3 | 0.8 |
| Val | 487,953 | 15,139 | 6.8 | 2.0 | 0.3 |
| Total: | 7,186,899 | 774,892 | 100 | 100 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabbani, N.; Al-Motawa, M.; Thornalley, P.J. Protein Glycation in Plants—An Under-Researched Field with Much Still to Discover. Int. J. Mol. Sci. 2020, 21, 3942. https://doi.org/10.3390/ijms21113942
Rabbani N, Al-Motawa M, Thornalley PJ. Protein Glycation in Plants—An Under-Researched Field with Much Still to Discover. International Journal of Molecular Sciences. 2020; 21(11):3942. https://doi.org/10.3390/ijms21113942
Chicago/Turabian StyleRabbani, Naila, Maryam Al-Motawa, and Paul J. Thornalley. 2020. "Protein Glycation in Plants—An Under-Researched Field with Much Still to Discover" International Journal of Molecular Sciences 21, no. 11: 3942. https://doi.org/10.3390/ijms21113942
APA StyleRabbani, N., Al-Motawa, M., & Thornalley, P. J. (2020). Protein Glycation in Plants—An Under-Researched Field with Much Still to Discover. International Journal of Molecular Sciences, 21(11), 3942. https://doi.org/10.3390/ijms21113942