Association of the Epithelial–Mesenchymal Transition (EMT) with Cisplatin Resistance

 ,
,  ,
, 
Abstract
:1. Introduction
2. Cisplatin in Cancer Therapy
3. Cisplatin Resistance
4. EMT Process in Healthy and Cancerous Tissues
5. Cisplatin Induces EMT-Mediated Cancer Chemoresistance
6. Strategies to Attenuate EMT-Related Cisplatin Resistance
6.1. Cluster of Differentiation Role
6.2. The Contribution of Exosomes
6.3. Forkhead Box Protein O1 (FOXO1) Signaling Pathway
6.4. MicroRNAs
6.5. PI3K/Akt Signaling Pathway
6.6. Wnt Signaling Pathway
6.7. Long Non-Coding RNAs
6.8. Nuclear Factor-κB (NF-ĸB) Signaling Pathway
6.9. Other Molecular Signaling Pathways
7. Possible Pathways for Further Targeting
7.1. PI3k/Akt Signaling Pathway
7.2. Laminin Subunit Beta-3
7.3. ZEB Proteins
7.4. Long Non-Coding RNAs
7.5. Targeting Drug Transporters
7.6. Other Molecular Signaling Pathways
8. Conclusions and Remarks
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| EMT | Epithelial–mesenchymal transition |
| CP | Cisplatin |
| ROS | Reactive oxygen species |
| DOX | Docetaxel |
| NPs | Nanoparticles |
| AMPK | AMP-activated protein kinase |
| lncRNA | Long non-coding RNAs |
| TAMs | Tumor associated macrophages |
| GC | Gastric cancer |
| OSCC | Oral squamous cell carcinoma |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| SIRT1 | Sirtuin 1 |
| MSI2 | Musashi RNA-binding protein 2 |
| FHL | Four-and-half LIM domain |
| TGF | Tumor growth factor |
| YAP1 | Yes-associated protein 1 |
| TAZ1 | Tazaffin |
| CRC | Colorectal cancer |
| UBE2O | Ubiquitin-conjugating enzyme E2O |
| mTOR | Mammalian target of rapamycin |
| TKI | Tyrosine kinase inhibitor |
| PTX | Paclitaxel |
| Cams | Classically activated macrophages |
| HCC | Hepatocellular carcinoma |
| CCL20 | Chemokine ligand 20 |
| CCR6 | Chemokine receptor 6 |
| EMP3 | Epithelial membrane protein 3 |
| NPC | Nasopharyngeal carcinoma |
| EBV | Epstein-Barr virus |
| PTP1B | Protein tyrosine phosphatase 1B |
| CSCs | Cancer stem cells |
| NSCLC | Non-small cell lung cancer |
| CIP2A | Cancerous inhibitor of protein phosphatase 2A |
| PP2A | Protein phosphatase 2A |
| PPI | Polyphyllin I |
| PPVII | Polyphyllin VII |
| PRS | Paris polyphylla |
| MM | Malignant mesothelioma |
| HOXA3 | Homeobox A3 |
| FOXC2 | Forkhead box protein C2 |
| FOX | Forkhead box |
| PKCα | Protein kinase Cα |
| UCA1 | Urogenital carcinoma antigen 1 |
| SIK2 | Salt inducible kinase 2 |
| VEGF | Vascular endothelial growth factor |
| PD-L1 | Programmed death-ligand 1 |
| MDM2 | Mouse double minute 2 homolog |
| ZCCHC10 | Zinc finger CCHC-type containing 10 |
| NF-κB | Nuclear factor-κB |
| IKKβ | Inhibitor of NF-κB kinase subunit β |
| Mcl-1 | Myeloid cell leukemia 1 |
| NCTD | Norcantharidin |
| DDR | DNA damage response |
| CTGF | Connective tissue growth factor |
| CYR61 | Cysteine rich angiogenic factor 61 |
| PDCD4 | Programmed cell death 4 |
| elF4E | Eukaryotic translation initiation factor 4E |
| MDR1 | Multidrug resistance protein 1 |
| LAMB3 | Laminin subunit β-3; |
| ZEB | Zinc-finger E-box binding homeobox |
| ERCC1 | Excision repair cross-complementary gene 1 |
| ABCG2 | ATP-binding cassette subfamily G member 2 |
| elF5A2 | Eukaryotic initiation factor 5A2 |
| DUBs | Deubiquitinating enzymes |
| USP37 | Ubiquitin specific peptidase 37 |
| Hh | Hedgehog |
| ATM | Ataxia telangiectasia mutated |
| GDF15 | Growth differentiation factor 15 |
| HIF-α, | Hypoxia inducible factor-α |
| COX-2 | Cyclooxygenase-2 |
| PDC | Pyruvate dehydrogenase complex |
| PDK1 | Pyruvate dehydrogenase kinase 1 |
| EGFR | Epidermal growth factor receptor |
| IGF1R | Insulin-like growth factor 1 |
| HOXA11-AS | Homeobox A11 antisense RNA |
| ACTN4 | Actinin alpha 4 |
| LUAD | Lung adenocarcinoma |
| BCL9 | B-cell lymphoma 9 |
| RKIP | Raf kinase inhibitor protein |
| FAT10 | HLA-F adjacent transcript 10 |
| H2A Z | H2A histone family member Z |
| ABC | ATP-binding cassette |
| P-gp | P-glycoprotein |
| MSDs | Membrane spinning domains |
| NBDs | Nucleotide binding domains |
| BBB | Blood–brain barrier |
| BTB | Blood–testis barrier |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA A Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, C.E.; Miller, K.D.; Dale, W.; Mohile, S.G.; Cohen, H.J.; Leach, C.R.; Goding Sauer, A.; Jemal, A.; Siegel, R.L. Cancer statistics for adults aged 85 years and older, 2019. CA A Cancer J. Clin. 2019, 69, 452–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA A Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Zhang, J.; Arfuso, F.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Kumar, A.P.; Ahn, K.S.; Sethi, G. Targeting TNF-related apoptosis-inducing ligand (TRAIL) receptor by natural products as a potential therapeutic approach for cancer therapy. Exp. Biol. Med. (Maywood) 2015, 240, 760–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugam, M.K.; Warrier, S.; Kumar, A.P.; Sethi, G.; Arfuso, F. Potential Role of Natural Compounds as Anti-Angiogenic Agents in Cancer. Curr. Vasc. Pharmacol. 2017, 15, 503–519. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.; Mortezaee, K.; Majidpoor, J. Stromal reprogramming: A target for tumor therapy. Life Sci. 2019, 239, 117049. [Google Scholar] [CrossRef] [PubMed]
- Prasannan, R.; Kalesh, K.A.; Shanmugam, M.K.; Nachiyappan, A.; Ramachandran, L.; Nguyen, A.H.; Kumar, A.P.; Lakshmanan, M.; Ahn, K.S.; Sethi, G. Key cell signaling pathways modulated by zerumbone: role in the prevention and treatment of cancer. Biochem. Pharmacol. 2012, 84, 1268–1276. [Google Scholar] [CrossRef] [Green Version]
- Mortezaee, K.; Potes, Y.; Mirtavoos-Mahyari, H.; Motevaseli, E.; Shabeeb, D.; Musa, A.E.; Najafi, M.; Farhood, B. Boosting immune system against cancer by melatonin: A mechanistic viewpoint. Life Sci. 2019, 238, 116960. [Google Scholar] [CrossRef]
- Mortezaee, K.; Najafi, M.; Farhood, B.; Ahmadi, A.; Shabeeb, D.; Musa, A.E. NF-κB targeting for overcoming tumor resistance and normal tissues toxicity. J. Cell. Physiol. 2019, 234, 17187–17204. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Manu, K.A.; Ong, T.H.; Ramachandran, L.; Surana, R.; Bist, P.; Lim, L.H.; Kumar, A.P.; Hui, K.M.; Sethi, G. Inhibition of CXCR4/CXCL12 signaling axis by ursolic acid leads to suppression of metastasis in transgenic adenocarcinoma of mouse prostate model. Int. J. Cancer 2011, 129, 1552–1563. [Google Scholar] [CrossRef]
- Ramachandran, L.; Manu, K.A.; Shanmugam, M.K.; Li, F.; Siveen, K.S.; Vali, S.; Kapoor, S.; Abbasi, T.; Surana, R.; Smoot, D.T.; et al. Isorhamnetin inhibits proliferation and invasion and induces apoptosis through the modulation of peroxisome proliferator-activated receptor γ activation pathway in gastric cancer. J. Biol. Chem. 2012, 287, 38028–38040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Xie, W.; Gao, L.; Huang, G.; Zhou, J.; Mei, B.; Chen, J. Impact of neoadjuvant chemotherapy on survival prognosis and pathological downstaging in patients presenting with high-risk upper tract urothelial carcinoma: A protocol for systematic review and meta analysis. Medicine 2020, 99, e20184. [Google Scholar] [CrossRef] [PubMed]
- Minami, K.; Ueda, N.; Ishimoto, K.; Tsujiuchi, T. Lysophosphatidic acid receptor-2 (LPA2)-mediated signaling enhances chemoresistance in melanoma cells treated with anticancer drugs. Mol. Cell. Biochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Qiu, N.; Yin, J.; Zhang, J.; Liu, H.; Guo, W.; Liu, M.; Liu, T.; Chen, D.; Luo, K.; et al. SRGN crosstalks with YAP to maintain chemoresistance and stemness in breast cancer cells by modulating HDAC2 expression. Theranostics 2020, 10, 4290–4307. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, G.; Gao, Y.; Liang, H. HOTAIR/miR-125 axis-mediated Hexokinase 2 expression promotes chemoresistance in human glioblastoma. J. Cell. Mol. Med. 2020. [Google Scholar] [CrossRef] [Green Version]
- Paciello, F.; Rita Fetoni, A.; Mezzogori, D.; Rolesi, R.; Di Pino, A.; Paludetti, G.; Grassi, C.; Troiani, D. The dual role of curcumin and ferulic acid in counteracting chemoresistance and cisplatin-induced ototoxicity. Sci. Rep. 2020, 10, 1063. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z.; Li, X.; Sun, Y.; Li, L.; Zhang, Q.; Zhu, L.; Zhong, Z.; Wang, M.; Wang, Q.; Liu, Z.; et al. NOS1 expression promotes proliferation and invasion and enhances chemoresistance in ovarian cancer. Oncol. Lett. 2020, 19, 2989–2995. [Google Scholar] [CrossRef] [Green Version]
- Dou, N.; Hu, Q.; Li, L.; Wu, Q.; Li, Y.; Gao, Y. USP32 promotes tumorigenesis and chemoresistance in gastric carcinoma via upregulation of SMAD2. Int. J. Biol. Sci. 2020, 16, 1648–1657. [Google Scholar] [CrossRef] [Green Version]
- Xi, Y.; Yuan, P.; Li, T.; Zhang, M.; Liu, M.F.; Li, B. hENT1 reverses chemoresistance by regulating glycolysis in pancreatic cancer. Cancer Lett. 2020, 479, 112–122. [Google Scholar] [CrossRef]
- Tang, L.; Chen, Y.; Chen, H.; Jiang, P.; Yan, L.; Mo, D.; Tang, X.; Yan, F. DCST1-AS1 Promotes TGF-beta-Induced Epithelial-Mesenchymal Transition and Enhances Chemoresistance in Triple-Negative Breast Cancer Cells via ANXA1. Front. Oncol. 2020, 10, 280. [Google Scholar] [CrossRef]
- Lin, G.; Zhao, R.; Wang, Y.; Han, J.; Gu, Y.; Pan, Y.; Ren, C.; Ren, S.; Xu, C. Dynamic analysis of N-glycomic and transcriptomic changes in the development of ovarian cancer cell line A2780 to its three cisplatin-resistant variants. Ann. Transl. Med. 2020, 8, 289. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Quan, L.N.; Meng, Q.; Wang, H.Y.; Wang, J.; Yu, P.; Fu, J.T.; Li, Y.J.; Chen, J.; Cheng, H.; et al. miR-548e Sponged by ZFAS1 Regulates Metastasis and Cisplatin Resistance of OC by Targeting CXCR4 and let-7a/BCL-XL/S Signaling Axis. Mol. Ther. Nucleic Acids 2020, 20, 621–638. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.C.; Teo, W.H.; Huang, T.F.; Lee, T.C.; Lo, J.F. Combinatorial Low Dose Arsenic Trioxide and Cisplatin Exacerbates Autophagy via AMPK/STAT3 Signaling on Targeting Head and Neck Cancer Initiating Cells. Front. Oncol. 2020, 10, 463. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, D.P.; Heyza, J.; Zimmermann, C.M.; Patrick, S.M.; Merkel, O.M. Nanoparticle-Mediated Gene Silencing for Sensitization of Lung Cancer to Cisplatin Therapy. Molecules 2020, 25, 1994. [Google Scholar] [CrossRef]
- Chen, T.W.; Jan, I.S.; Chang, D.Y.; Lin, C.H.; Chen, I.C.; Chen, H.M.; Cheng, A.L.; Lu, Y.S. Systemic treatment of breast cancer with leptomeningeal metastases using bevacizumab, etoposide and cisplatin (BEEP regimen) significantly improves overall survival. J. Neuro-Oncol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.W.; Ryu, H.; Song, I.C.; Yun, H.J.; Jo, D.Y.; Ko, Y.B.; Lee, H.J. Efficacy of cisplatin combined with topotecan in patients with advanced or recurrent ovarian cancer as second- or higher-line palliative chemotherapy. Medicine 2020, 99, e19931. [Google Scholar] [CrossRef]
- Li, W.H.; Zhang, L.; Wu, Y.H. CDKN3 regulates cisplatin resistance to colorectal cancer through TIPE1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3614–3623. [Google Scholar] [CrossRef]
- Murakami, M.; Izumi, H.; Kurita, T.; Koi, C.; Morimoto, Y.; Yoshino, K. UBE2L6 is Involved in Cisplatin Resistance by Regulating the Transcription of ABCB6. Anti-Cancer Agents Med. Chem. 2020. [Google Scholar] [CrossRef]
- Jin, L.; Zhang, N.; Zhang, Q.; Ding, G.; Yang, Z.; Zhang, Z. Serum microRNAs as potential new biomarkers for cisplatin resistance in gastric cancer patients. PeerJ 2020, 8, e8943. [Google Scholar] [CrossRef] [Green Version]
- Kasikova, L.; Hensler, M.; Truxova, I.; Skapa, P.; Laco, J.; Belicova, L.; Praznovec, I.; Vosahlikova, S.; Halaska, M.J.; Brtnicky, T.; et al. Calreticulin exposure correlates with robust adaptive antitumor immunity and favorable prognosis in ovarian carcinoma patients. J. Immunother. Cancer 2019, 7, 312. [Google Scholar] [CrossRef]
- Jain, A.; Jahagirdar, D.; Nilendu, P.; Sharma, N.K. Molecular approaches to potentiate cisplatin responsiveness in carcinoma therapeutics. Expert Rev. Anticancer Ther. 2017, 17, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Rezaee, R.; Momtazi, A.A.; Monemi, A.; Sahebkar, A. Curcumin: A potentially powerful tool to reverse cisplatin-induced toxicity. Pharmacol. Res. 2017, 117, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Nance, E.A.; Mastorakos, P.; Chisholm, J.; Berry, S.; Eberhart, C.; Tyler, B.; Brem, H.; Suk, J.S.; Hanes, J. Convection enhanced delivery of cisplatin-loaded brain penetrating nanoparticles cures malignant glioma in rats. J. Controll. Release 2017, 263, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Wu, Q.; Fang, M.; Huang, W.; Zhu, H. miR-152-3p Sensitizes Glioblastoma Cells Towards Cisplatin Via Regulation Of SOS1. OncoTargets Ther. 2019, 12, 9513–9525. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Gong, Q.; Yu, Y.; Zhu, J.; Li, W. Knockdown of circ-ABCB10 promotes sensitivity of lung cancer cells to cisplatin via miR-556-3p/AK4 axis. BMC Pulm. Med. 2020, 20, 10. [Google Scholar] [CrossRef] [Green Version]
- Balog, J.A.; Hackler, L., Jr.; Kovacs, A.K.; Neuperger, P.; Alfoldi, R.; Nagy, L.I.; Puskas, L.G.; Szebeni, G.J. Single Cell Mass Cytometry Revealed the Immunomodulatory Effect of Cisplatin Via Downregulation of Splenic CD44+, IL-17A+ MDSCs and Promotion of Circulating IFN-gamma+ Myeloid Cells in the 4T1 Metastatic Breast Cancer Model. Int. J. Mol. Sci. 2019, 21, 170. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Li, W.; Jin, X.; Niu, T.; Cao, Y.; Zhou, P.; Zheng, M. Silencing long non-coding RNA NEAT1 enhances the suppression of cell growth, invasion, and apoptosis of bladder cancer cells under cisplatin chemotherapy. Int. J. Clin. Exp. Pathol. 2019, 12, 549–558. [Google Scholar]
- Jiang, H.; Xiong, W.; Chen, L.; Lv, Z.; Yang, C.; Li, Y. Knockdown of the long noncoding RNA HOTTIP inhibits cell proliferation and enhances cell sensitivity to cisplatin by suppressing the Wnt/beta-catenin pathway in prostate cancer. J. Cell. Biochem. 2019, 120, 8965–8974. [Google Scholar] [CrossRef]
- Zang, H.; Qian, G.; Arbiser, J.; Owonikoko, T.K.; Ramalingam, S.S.; Fan, S.; Sun, S.Y. Overcoming acquired resistance of EGFR-mutant NSCLC cells to the third generation EGFR inhibitor, osimertinib, with the natural product honokiol. Mol. Oncol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Bostan, M.; Petrica-Matei, G.G.; Ion, G.; Radu, N.; Mihaila, M.; Hainarosie, R.; Brasoveanu, L.I.; Roman, V.; Constantin, C.; Neagu, M.T. Cisplatin effect on head and neck squamous cell carcinoma cells is modulated by ERK1/2 protein kinases. Exp. Ther. Med. 2019, 18, 5041–5051. [Google Scholar] [CrossRef] [Green Version]
- Achkar, I.W.; Abdulrahman, N.; Al-Sulaiti, H.; Joseph, J.M.; Uddin, S.; Mraiche, F. Cisplatin based therapy: The role of the mitogen activated protein kinase signaling pathway. J. Transl. Med. 2018, 16, 96. [Google Scholar] [CrossRef] [PubMed]
- Fadejeva, I.; Olschewski, H.; Hrzenjak, A. MicroRNAs as regulators of cisplatin-resistance in non-small cell lung carcinomas. Oncotarget 2017, 8, 115754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Xia, M.; Wang, B.; Thapa, N.; Gan, L.; Sun, C.; Guo, E.; Huang, J.; Lu, Y.; Cai, H. Clarithromycin synergizes with cisplatin to inhibit ovarian cancer growth in vitro and in vivo. J. Ovarian Res. 2019, 12, 107. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, B.; Nitulescu, G.M.; Olaru, O.T.; Venables, L.; van de Venter, M. Anti-Cancer Activity of a 5-Aminopyrazole Derivative Lead Compound (BC-7) and Potential Synergistic Cytotoxicity with Cisplatin against Human Cervical Cancer Cells. Int. J. Mol. Sci. 2019, 20, 5559. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, M.; Shirakawa, Y.; Maeda, N.; Tanabe, S.; Noma, K.; Sakurama, K.; Katsui, K.; Nishizaki, M.; Fujiwara, T. Induction chemoradiotherapy including docetaxel, cisplatin, and 5-fluorouracil for locally advanced esophageal cancer. Esophagus Off. J. Jpn. Esophageal Soc. 2020. [Google Scholar] [CrossRef]
- Khafaji, M.; Zamani, M.; Vossoughi, M.; Iraji Zad, A. Doxorubicin/Cisplatin-Loaded Superparamagnetic Nanoparticles As A Stimuli-Responsive Co-Delivery System For Chemo-Photothermal Therapy. Int. J. Nanomed. 2019, 14, 8769–8786. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Kang, Y.; Sewastianik, T.; Wang, J.; Tanton, H.; Alder, K.; Dennis, P.; Xin, Y.; Wang, Z.; Liu, R.; et al. BCL9 provides multi-cellular communication properties in colorectal cancer by interacting with paraspeckle proteins. Nat. Commun. 2020, 11, 19. [Google Scholar] [CrossRef] [Green Version]
- Gan, D.; He, W.; Yin, H.; Gou, X. beta-elemene enhances cisplatin-induced apoptosis in bladder cancer cells through the ROS-AMPK signaling pathway. Oncol. Lett. 2020, 19, 291–300. [Google Scholar] [CrossRef]
- Heeren, A.M.; van Luijk, I.F.; Lakeman, J.; Pocorni, N.; Kole, J.; de Menezes, R.X.; Kenter, G.G.; Bosse, T.; de Kroon, C.D.; Jordanova, E.S. Neoadjuvant cisplatin and paclitaxel modulate tumor-infiltrating T cells in patients with cervical cancer. Cancer Immunol. Immunother. CII 2019, 68, 1759–1767. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Zhang, R.; An, X.; Li, Z.; Fang, C.; Pan, B.; Chen, W.; Xu, G.; Han, W. LncRNA HOXA-AS3 confers cisplatin resistance by interacting with HOXA3 in non-small-cell lung carcinoma cells. Oncogenesis 2019, 8, 60. [Google Scholar] [CrossRef]
- Nan, Y. Lung carcinoma therapy using epidermal growth factor receptortargeted lipid polymeric nanoparticles coloaded with cisplatin and doxorubicin. Oncol. Rep. 2019, 42, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Roy, S.; Kar, M.; Chakraborty, A.; Kumar, A.; Delogu, F.; Asthana, S.; Hande, M.P.; Banerjee, B. Combined treatment with cisplatin and the tankyrase inhibitor XAV-939 increases cytotoxicity, abrogates cancer-stem-like cell phenotype and increases chemosensitivity of head-and-neck squamous-cell carcinoma cells. Mutat. Res. 2019, 846, 503084. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Hayashi, Y.; Watanabe, K.; Yoshimi, R.; Hibi, H. Concurrent chemoradiotherapy with intravenous cisplatin and docetaxel for advanced oral cancer. Nagoya J. Med Sci. 2019, 81, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Wang, L.; Li, Y.; Song, H.; Chu, H.; Yang, H.; Guo, A.; Jiao, J. SiRNA-Mediated RRM2 Gene Silencing Combined with Cisplatin in the Treatment of Epithelial Ovarian Cancer In Vivo: An Experimental Study of Nude Mice. Int. J. Med. Sci. 2019, 16, 1510–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Shanmugam, M.K.; Xiang, P.; Yam, T.Y.A.; Kumar, V.; Chew, W.S.; Chang, J.K.; Ali, M.Z.B.; Reolo, M.J.Y.; Peh, Y.X.; et al. Sphingosine 1-Phosphate Receptor 2 Induces Otoprotective Responses to Cisplatin Treatment. Cancers 2020, 12, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayurova, E.; Jansons, J.; Skrastina, D.; Smirnova, O.; Mezale, D.; Kostyusheva, A.; Kostyushev, D.; Petkov, S.; Podschwadt, P.; Valuev-Elliston, V.; et al. HIV-1 Reverse Transcriptase Promotes Tumor Growth and Metastasis Formation via ROS-Dependent Upregulation of Twist. Oxidative Med. Cell. Longev. 2019, 2019, 6016278. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.S.; Noh, M.R.; Jung, E.M.; Kim, W.Y.; Southekal, S.; Guda, C.; Foster, K.W.; Oupicky, D.; Ferrer, F.A.; Padanilam, B.J. Proximal tubule cyclophilin D regulates fatty acid oxidation in cisplatin-induced acute kidney injury. Kidney Int. 2019. [Google Scholar] [CrossRef]
- Li, Q.; Liang, X.; Yang, Y.; Zeng, X.; Zhong, X.; Huang, C. Panax notoginseng saponins ameliorate cisplatin-induced mitochondrial injury via the HIF-1alpha/mitochondria/ROS pathway. FEBS Open Bio 2020, 10, 118–126. [Google Scholar] [CrossRef]
- Dai, X.; Wang, L.; Deivasigamni, A.; Looi, C.Y.; Karthikeyan, C.; Trivedi, P.; Chinnathambi, A.; Alharbi, S.A.; Arfuso, F.; Dharmarajan, A.; et al. A novel benzimidazole derivative, MBIC inhibits tumor growth and promotes apoptosis via activation of ROS-dependent JNK signaling pathway in hepatocellular carcinoma. Oncotarget 2017, 8, 12831–12842. [Google Scholar] [CrossRef] [Green Version]
- Hanigan, M.H.; Devarajan, P. Cisplatin nephrotoxicity: Molecular mechanisms. Cancer Ther. 2003, 1, 47. [Google Scholar]
- Tan, R.Z.; Liu, J.; Zhang, Y.Y.; Wang, H.L.; Li, J.C.; Liu, Y.H.; Zhong, X.; Zhang, Y.W.; Yan, Y.; Lan, H.Y.; et al. Curcumin relieved cisplatin-induced kidney inflammation through inhibiting Mincle-maintained M1 macrophage phenotype. Phytomedicine 2019, 52, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Said, R.S.; Mantawy, E.M.; El-Demerdash, E. Mechanistic perspective of protective effects of resveratrol against cisplatin-induced ovarian injury in rats: Emphasis on anti-inflammatory and anti-apoptotic effects. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 1225–1238. [Google Scholar] [CrossRef] [PubMed]
- Domitrovic, R.; Cvijanovic, O.; Pernjak-Pugel, E.; Skoda, M.; Mikelic, L.; Crncevic-Orlic, Z. Berberine exerts nephroprotective effect against cisplatin-induced kidney damage through inhibition of oxidative/nitrosative stress, inflammation, autophagy and apoptosis. Food Chem. Toxicol. 2013, 62, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Mortezaee, K. CXCL12/CXCR4 axis in the microenvironment of solid tumors: A critical mediator of metastasis. Life Sci. 2020, 249, 117534. [Google Scholar] [CrossRef] [PubMed]
- Mortezaee, K. Hypoxia induces core-to-edge transition of progressive tumoral cells: A critical review on differential yet corroborative roles for HIF-1α and HIF-2α. Life Sci. 2020, 242, 117145. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, A.; Bianconi, V.; Pirro, M.; Orafai, H.M.; Sahebkar, A. The role of TFEB in tumor cell autophagy: Diagnostic and therapeutic opportunities. Life Sci. 2020, 244, 117341. [Google Scholar] [CrossRef]
- Meng, D.; Li, Z.; Ma, X.; Wu, L.; Fu, L.; Qin, G. ETV5 overexpression contributes to tumor growth and progression of thyroid cancer through PIK3CA. Life Sci. 2020, 253, 117693. [Google Scholar] [CrossRef]
- Gu, Y.; Fei, Z.; Zhu, R. miR-21 modulates cisplatin resistance of gastric cancer cells by inhibiting autophagy via the PI3K/Akt/mTOR pathway. Anti-cancer Drugs 2020. [Google Scholar] [CrossRef]
- Yin, J.; Chen, D.; Luo, K.; Lu, M.; Gu, Y.; Zeng, S.; Chen, X.; Song, Y.; Zhang, Z.; Zheng, G.; et al. Cip2a/miR-301a feedback loop promotes cell proliferation and invasion of triple-negative breast cancer. J. Cancer 2019, 10, 5964–5974. [Google Scholar] [CrossRef]
- Hazari, Y.; Bravo-San Pedro, J.M.; Hetz, C.; Galluzzi, L.; Kroemer, G. Autophagy in hepatic adaptation to stress. J. Hepatol. 2020, 72, 183–196. [Google Scholar] [CrossRef]
- Galluzzi, L.; Green, D.R. Autophagy-Independent Functions of the Autophagy Machinery. Cell 2019, 177, 1682–1699. [Google Scholar] [CrossRef] [PubMed]
- Jamali, Z.; Taheri-Anganeh, M.; Shabaninejad, Z.; Keshavarzi, A.; Taghizadeh, H.; Razavi, Z.S.; Mottaghi, R.; Abolhassan, M.; Movahedpour, A.; Mirzaei, H. Autophagy regulation by microRNAs: Novel insights into osteosarcoma therapy. IUBMB Life 2020, n/a. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Lee, J.Y.; Ermolenko, L.; Mazumder, A.; Ji, S.; Ryu, H.; Kim, H.; Kim, D.W.; Lee, J.W.; Dicato, M.; et al. Tetrahydrobenzimidazole TMQ0153 triggers apoptosis, autophagy and necroptosis crosstalk in chronic myeloid leukemia. Cell Death Dis. 2020, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Sala de Oyanguren, F.J.; Rainey, N.E.; Moustapha, A.; Saric, A.; Sureau, F.; O’Connor, J.E.; Petit, P.X. Highlighting Curcumin-Induced Crosstalk between Autophagy and Apoptosis as Supported by Its Specific Subcellular Localization. Cells 2020, 9, 361. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N. Fusobacterium nucleatum promotes chemoresistance to colorectal cancer by modulating autophagy. Cell 2017, 170, 548–563. e516. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhou, Y.; He, J.; Sun, H.; Jin, Z. Long non-coding RNA H19 mediates ovarian cancer cell cisplatin-resistance and migration during EMT. Int. J. Clin. Exp. Pathol. 2019, 12, 2506–2515. [Google Scholar]
- Liu, Y.; Hu, Q.; Wang, X. AFAP1-AS1 induces cisplatin resistance in non-small cell lung cancer through PI3K/AKT pathway. Oncol. Lett. 2020, 19, 1024–1030. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, I.; Kasamatsu, A.; Yamatoji, M.; Nakashima, D.; Saito, K.; Higo, M.; Endo-Sakamoto, Y.; Shiiba, M.; Tanzawa, H.; Uzawa, K. Kinesin family member 14 in human oral cancer: A potential biomarker for tumoral growth. Biochem. Biophys. Rep. 2015, 3, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Zhou, S.; Wang, X.; Mao, E.; Huang, L. SNHG14 stimulates cell autophagy to facilitate cisplatin resistance of colorectal cancer by regulating miR-186/ATG14 axis. Biomed. Pharmacother. Biomed. Pharmacother. 2020, 121, 109580. [Google Scholar] [CrossRef]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-associated macrophages as treatment targets in oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef]
- Liu, F.; Qiu, F.; Fu, M.; Chen, H.; Wang, H. Propofol Reduces Epithelial to Mesenchymal Transition, Invasion and Migration of Gastric Cancer Cells through the MicroRNA-195-5p/Snail Axis. Med. Sci. Monit. 2020, 26, e920981. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Li, C.S.; Zhou, Y.; Lu, X.H. Propofol facilitates cisplatin sensitivity via lncRNA MALAT1/miR-30e/ATG5 axis through suppressing autophagy in gastric cancer. Life Sci. 2020, 244, 117280. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xia, H. Oridonin elevates sensitivity of ovarian carcinoma cells to cisplatin via suppressing cisplatin-mediated autophagy. Life Sci. 2019, 233, 116709. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Cai, M.; Zhang, Y.; Tao, L.; Guo, R. miR-29c-3p inhibits autophagy and cisplatin resistance in ovarian cancer by regulating FOXP1/ATG14 pathway. Cell Cycle 2020, 19, 193–206. [Google Scholar] [CrossRef]
- Pan, X.; Chen, Y.; Shen, Y.; Tantai, J. Knockdown of TRIM65 inhibits autophagy and cisplatin resistance in A549/DDP cells by regulating miR-138-5p/ATG7. Cell Death Dis. 2019, 10, 429. [Google Scholar] [CrossRef]
- Wang, Y.; Lina, L.; Xu, L.; Yang, Z.; Qian, Z.; Zhou, J.; Suoni, L. Arctigenin enhances the sensitivity of cisplatin resistant colorectal cancer cell by activating autophagy. Biochem. Biophys. Res. Commun. 2019, 520, 20–26. [Google Scholar] [CrossRef]
- Liu, J.; Huang, Y.; Wang, H.; Wu, D. MiR-106a-5p promotes 5-FU resistance and the metastasis of colorectal cancer by targeting TGFbetaR2. Int. J. Clin. Exp. Pathol. 2018, 11, 5622–5634. [Google Scholar]
- Pourhanifeh, M.H.; Mahjoubin-Tehran, M.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S.; Mirzaei, H.; Asemi, Z. MicroRNAs and exosomes: Small molecules with big actions in multiple myeloma pathogenesis. IUBMB Life 2020, 72, 314–333. [Google Scholar] [CrossRef]
- Naeli, P.; Yousefi, F.; Ghasemi, Y.; Savardashtaki, A.; Mirzaei, H. The Role of MicroRNAs in Lung Cancer: Implications for Diagnosis and Therapy. Curr. Mol. Med. 2020, 20, 90–101. [Google Scholar] [CrossRef]
- Feng, W.T.; Yao, R.; Xu, L.J.; Zhong, X.M.; Liu, H.; Sun, Y.; Zhou, L.L. Effect of miR-363 on the proliferation, invasion and apoptosis of laryngeal cancer by targeting Mcl-1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4564–4572. [Google Scholar] [CrossRef]
- Xu, C.; Du, Z.; Ren, S.; Liang, X.; Li, H. MiR-129-5p sensitization of lung cancer cells to etoposide-induced apoptosis by reducing YWHAB. J. Cancer 2020, 11, 858–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.-T.; Wang, L.; Wang, H.; Tang, F.-R.; Cai, W.-Q.; Sethi, G.; Xin, H.-W.; Ma, Z. Insights into Biological Role of LncRNAs in Epithelial-Mesenchymal Transition. Cells 2019, 8, 1178. [Google Scholar] [CrossRef] [Green Version]
- Vafadar, A.; Shabaninejad, Z.; Movahedpour, A.; Mohammadi, S.; Fathullahzadeh, S.; Mirzaei, H.R.; Namdar, A.; Savardashtaki, A.; Mirzaei, H. Long Non-Coding RNAs As Epigenetic Regulators in Cancer. Curr. Pharm. Design 2019, 25, 3563–3577. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhu, Q.; Lu, L.; Liu, Y. MiR-132 inhibits migration and invasion and increases chemosensitivity of cisplatin-resistant oral squamous cell carcinoma cells via targeting TGF-beta1. Bioengineered 2020, 11, 91–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.X.; Zhang, Q.; Hu, H.; Jin, Y.; Zeng, A.L.; Xia, Y.B.; Xu, L. A novel circular RNA circFN1 enhances cisplatin resistance in gastric cancer via sponging miR-182-5p. J. Cell. Biochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Li, H.; Bi, Y.; Zhang, Z.; Wang, S.; Li, J.; Yang, Z.; Lv, X.; Zhou, B.; Yin, Z. The Polymorphisms of lncRNA HOXA11-AS and the risk of Lung Cancer in Northeastern Chinese population. J. Cancer 2020, 11, 592–598. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zheng, M.; Zhu, L.; Deng, L.; Li, X.; Gao, L.; Wang, C.; Wang, H.; Liu, J.; Lin, B. Low BCL9 expression inhibited ovarian epithelial malignant tumor progression by decreasing proliferation, migration, and increasing apoptosis to cancer cells. Cancer Cell Int. 2019, 19, 330. [Google Scholar] [CrossRef]
- Cheng, Y.; Shen, X.; Zheng, M.; Zou, G.; Shen, Y. Knockdown Of lncRNA NCK-AS1 Regulates Cisplatin Resistance Through Modulating miR-137 In Osteosarcoma Cells. OncoTargets Ther. 2019, 12, 11057–11068. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Yang, F.; Wu, K.; Kai, Z.; Xinxing, X.; An, Y.; Xu, F.; Xun, J.; Lv, X.; Zhang, X.; et al. Lost miR-141 and upregulated TM4SF1 expressions associate with poor prognosis of pancreatic cancer: Regulation of EMT and angiogenesis by miR-141 and TM4SF1 via AKT. Cancer Biol. Ther. 2020, 1–10. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, C.; Tan, T.; Li, S.; Tang, S.; Chen, X. Sinomenine sensitizes human gastric cancer cells to cisplatin through negative regulation of PI3K/AKT/Wnt signaling pathway. Anti-cancer Drugs 2019, 30, 983–990. [Google Scholar] [CrossRef]
- Silva, M.M.; Rocha, C.R.R.; Kinker, G.S.; Pelegrini, A.L.; Menck, C.F.M. The balance between NRF2/GSH antioxidant mediated pathway and DNA repair modulates cisplatin resistance in lung cancer cells. Sci. Rep. 2019, 9, 17639. [Google Scholar] [CrossRef] [PubMed]
- Asaka, R.; Miyamoto, T.; Yamada, Y.; Ando, H.; Mvunta, D.H.; Kobara, H.; Shiozawa, T. Sirtuin 1 promotes the growth and cisplatin resistance of endometrial carcinoma cells: A novel therapeutic target. Lab. Investig. J. Tech. Methods Pathol. 2015, 95, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Loh, C.-Y.; Chai, J.Y.; Tang, T.F.; Wong, W.F.; Sethi, G.; Shanmugam, M.K.; Chong, P.P.; Looi, C.Y. The E-Cadherin and N-Cadherin Switch in Epithelial-to-Mesenchymal Transition: Signaling, Therapeutic Implications, and Challenges. Cells 2019, 8, 1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roshan, M.K.; Soltani, A.; Soleimani, A.; Kahkhaie, K.R.; Afshari, A.R.; Soukhtanloo, M. Role of AKT and mTOR signaling pathways in the induction of epithelial-mesenchymal transition (EMT) process. Biochimie 2019, 165, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.J.; Son, D.; Ko, T.H.; Hong, W.; Yun, W.; Jang, J.; Choi, J.I.; Song, G.; Lee, J.; Kim, I.Y.; et al. mRNA-Driven Generation of Transgene-Free Neural Stem Cells from Human Urine-Derived Cells. Cells 2019, 8, 1043. [Google Scholar] [CrossRef] [Green Version]
- Tentler, D.; Lomert, E.; Novitskaya, K.; Barlev, N.A. Role of ACTN4 in Tumorigenesis, Metastasis, and EMT. Cells 2019, 8, 1427. [Google Scholar] [CrossRef] [Green Version]
- Teeuwssen, M.; Fodde, R. Wnt Signaling in Ovarian Cancer Stemness, EMT, and Therapy Resistance. J. Clin. Med. 2019, 8, 1658. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Chen, L.; Liu, L.; Niu, X. EMT-Mediated Acquired EGFR-TKI Resistance in NSCLC: Mechanisms and Strategies. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Ayyar, B.V.; Arora, S.; O’Kennedy, R. Coming-of-Age of Antibodies in Cancer Therapeutics. Trends Pharmacol. Sci. 2016, 37, 1009–1028. [Google Scholar] [CrossRef]
- Yan, W.; Wu, Q.; Yao, W.; Li, Y.; Liu, Y.; Yuan, J.; Han, R.; Yang, J.; Ji, X.; Ni, C. MiR-503 modulates epithelial-mesenchymal transition in silica-induced pulmonary fibrosis by targeting PI3K p85 and is sponged by lncRNA MALAT1. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Zhu, Q.; Zhou, J. Long non-coding RNA MALAT1 interaction with miR-429 regulates the proliferation and EMT of lung adenocarcinoma cells through RhoA. Int. J. Clin. Exp. Pathol. 2019, 12, 419–430. [Google Scholar] [PubMed]
- Cai, H.; Yan, L.; Liu, N.; Xu, M.; Cai, H. IFI16 promotes cervical cancer progression by upregulating PD-L1 in immunomicroenvironment through STING-TBK1-NF-kB pathway. Biomed. Pharmacother. Biomed. Pharmacother. 2020, 123, 109790. [Google Scholar] [CrossRef] [PubMed]
- Qu, T.; Zhao, Y.; Chen, Y.; Jin, S.; Fang, Y.; Jin, X.; Sun, L.; Ma, Y. Down-regulated MAC30 expression inhibits breast cancer cell invasion and EMT by suppressing Wnt/beta-catenin and PI3K/Akt signaling pathways. Int. J. Clin. Exp. Pathol. 2019, 12, 1888–1896. [Google Scholar] [PubMed]
- Baek, S.H.; Ko, J.H.; Lee, J.H.; Kim, C.; Lee, H.; Nam, D.; Lee, J.; Lee, S.G.; Yang, W.M.; Um, J.Y.; et al. Ginkgolic Acid Inhibits Invasion and Migration and TGF-beta-Induced EMT of Lung Cancer Cells Through PI3K/Akt/mTOR Inactivation. J. Cell. Physiol. 2017, 232, 346–354. [Google Scholar] [CrossRef]
- Zhang, H.; Li, M.; Xu, X. MicroRNA-204 attenuates the migration and invasion of pancreatic cancer cells by targeting ZEB1/EMT axis. Int. J. Clin. Exp. Pathol. 2018, 11, 3802–3811. [Google Scholar]
- Yang, L.; Yu, Y.; Xiong, Z.; Chen, H.; Tan, B.; Hu, H. Downregulation of SEMA4C Inhibit Epithelial-Mesenchymal Transition (EMT) and the Invasion and Metastasis of Cervical Cancer Cells via Inhibiting Transforming Growth Factor-beta 1 (TGF-beta1)-Induced Hela cells p38 Mitogen-Activated Protein Kinase (MAPK) Activation. Med. Sci. Monit. 2020, 26, e918123. [Google Scholar] [CrossRef]
- Yang, M.H.; Lee, J.H.; Ko, J.H.; Jung, S.H.; Sethi, G.; Ahn, K.S. Brassinin Represses Invasive Potential of Lung Carcinoma Cells through Deactivation of PI3K/Akt/mTOR Signaling Cascade. Molecules 2019, 24, 1584. [Google Scholar] [CrossRef] [Green Version]
- Bunz, F. EMT and Back Again: Visualizing the Dynamic Phenotypes of Metastasis. Cancer Res. 2020, 80, 153–155. [Google Scholar] [CrossRef]
- Ko, J.H.; Nam, D.; Um, J.Y.; Jung, S.H.; Sethi, G.; Ahn, K.S. Bergamottin Suppresses Metastasis of Lung Cancer Cells through Abrogation of Diverse Oncogenic Signaling Cascades and Epithelial-to-Mesenchymal Transition. Molecules 2018, 23, 1601. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Zhang, Y.; Geng, Y.; Lu, W.; Yin, J.; Li, Z.; Huang, L.; Liu, H.; Xu, N. IGFBP2 promotes the EMT of colorectal cancer cells by regulating E-cadherin expression. Int. J. Clin. Exp. Pathol. 2019, 12, 2559–2565. [Google Scholar]
- Wu, Z.; Xue, S.; Zheng, B.; Ye, R.; Xu, G.; Zhang, S.; Zeng, T.; Zheng, W.; Chen, C. Expression and significance of c-kit and epithelial-mesenchymal transition (EMT) molecules in thymic epithelial tumors (TETs). J. Thorac. Dis. 2019, 11, 4602–4612. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Wang, K.; Feng, Z.; Zhang, M.Y.; Wu, J.; Geng, J.J.; Chen, Z.N. CD147 deficiency in T cells prevents thymic involution by inhibiting the EMT process in TECs in the presence of TGFbeta. Cell. Mol. Immunol. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seddiki, R.; Narayana, G.H.N.S.; Strale, P.-O.; Balcioglu, H.E.; Peyret, G.; Yao, M.; Le, A.P.; Teck Lim, C.; Yan, J.; Ladoux, B. Force-dependent binding of vinculin to α-catenin regulates cell–cell contact stability and collective cell behavior. Mol. Biol. Cell 2018, 29, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.T.; Ma, Y.C.; Wang, C.H.; Xu, Y.; Gu, G.J. Prognostic and clinicopathologic significance of AEG-1/MTDH and E-cadherin expression in human gallbladder carcinoma. Int. J. Clin. Exp. Pathol. 2018, 11, 6025–6031. [Google Scholar]
- Haensel, D.; Dai, X. Epithelial-to-mesenchymal transition in cutaneous wound healing: Where we are and where we are heading. Dev. Dyn. 2018, 247, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Xing, T.; Yang, Z.; Dudek, R.; Lu, Q.; Chen, Y.-H. Epithelial mesenchymal transition in embryonic development, tissue repair and cancer: A comprehensive overview. J. Clin. Med. 2018, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Rout-Pitt, N.; Farrow, N.; Parsons, D.; Donnelley, M. Epithelial mesenchymal transition (EMT): A universal process in lung diseases with implications for cystic fibrosis pathophysiology. Respir. Res. 2018, 19, 136. [Google Scholar] [CrossRef]
- Kassouf, T.; Larive, R.M.; Morel, A.; Urbach, S.; Bettache, N.; Marcial Medina, M.C.; Merezegue, F.; Freiss, G.; Peter, M.; Boissiere-Michot, F.; et al. The Syk Kinase Promotes Mammary Epithelial Integrity and Inhibits Breast Cancer Invasion by Stabilizing the E-Cadherin/Catenin Complex. Cancers 2019, 11, 1974. [Google Scholar] [CrossRef] [Green Version]
- Shu, J.; Wang, L.; Han, F.; Chen, Y.; Wang, S.; Luo, F. BTBD7 Downregulates E-Cadherin and Promotes Epithelial-Mesenchymal Transition in Lung Cancer. BioMed Res. Int. 2019, 2019, 5937635. [Google Scholar] [CrossRef] [Green Version]
- Dehli, J.; Karlsson, C.; Bizelli-Silveira, C.; Jiang, X.; Kraft, D.; Foss, M. E-cadherin mediated cell-biomaterial interaction reduces migration of keratinocytes in-vitro. Colloids Surf. B Biointerfaces 2019, 180, 326–333. [Google Scholar] [CrossRef]
- Sun, S.; Gong, Q. The expressions and prognostic implications of Twist and E-cadherin in adenocarcinomas of the gastroesophageal junction and proximal gastric carcinoma. Medicine 2019, 98, e18449. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; He, H.; Lin, D.; Wang, C.; Zhu, Y.; Xu, D. Up-regulation of E-cadherin by saRNA inhibits the migration and invasion of renal carcinoma cells. Int. J. Clin. Exp. Pathol. 2018, 11, 5792–5800. [Google Scholar] [PubMed]
- Fang, D.D.; Tang, Q.; Kong, Y.; Wang, Q.; Gu, J.; Fang, X.; Zou, P.; Rong, T.; Wang, J.; Yang, D.; et al. MDM2 inhibitor APG-115 synergizes with PD-1 blockade through enhancing antitumor immunity in the tumor microenvironment. J. Immunother. Cancer 2019, 7, 327. [Google Scholar] [CrossRef]
- Miro, C.; Di Cicco, E.; Ambrosio, R.; Mancino, G.; Di Girolamo, D.; Cicatiello, A.G.; Sagliocchi, S.; Nappi, A.; De Stefano, M.A.; Luongo, C. Thyroid hormone induces progression and invasiveness of squamous cell carcinomas by promoting a ZEB-1/E-cadherin switch. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Manshouri, R.; Coyaud, E.; Kundu, S.T.; Peng, D.H.; Stratton, S.A.; Alton, K.; Bajaj, R.; Fradette, J.J.; Minelli, R.; Peoples, M.D.; et al. ZEB1/NuRD complex suppresses TBC1D2b to stimulate E-cadherin internalization and promote metastasis in lung cancer. Nat. Commun. 2019, 10, 5125. [Google Scholar] [CrossRef]
- Liu, B.; Li, X.; Li, C.; Xu, R.; Sun, X. miR-25 mediates metastasis and epithelial-mesenchymal-transition in human esophageal squamous cell carcinoma via regulation of E-cadherin signaling. Bioengineered 2019, 10, 679–688. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Ueo, H.; Kai, Y.; Noda, M.; Hu, Q.; Sato, K.; Fujii, A.; Hayashi, N.; Tsuruda, Y.; Otsu, H.; et al. N-Cadherin mRNA Levels in Peripheral Blood Could Be a Potential Indicator of New Metastases in Breast Cancer: A Pilot Study. Int. J. Mol. Sci. 2020, 21, 511. [Google Scholar] [CrossRef] [Green Version]
- Po, J.W.; Roohullah, A.; Lynch, D.; DeFazio, A.; Harrison, M.; Harnett, P.R.; Kennedy, C.; de Souza, P.; Becker, T.M. Improved ovarian cancer EMT-CTC isolation by immunomagnetic targeting of epithelial EpCAM and mesenchymal N-cadherin. J. Circ. Biomark. 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef]
- Zhu, G.J.; Song, P.P.; Zhou, H.; Shen, X.H.; Wang, J.G.; Ma, X.F.; Gu, Y.J.; Liu, D.D.; Feng, A.N.; Qian, X.Y.; et al. Role of epithelial-mesenchymal transition markers E-cadherin, N-cadherin, beta-catenin and ZEB2 in laryngeal squamous cell carcinoma. Oncol. Lett. 2018, 15, 3472–3481. [Google Scholar] [CrossRef] [Green Version]
- Azimi, I.; Petersen, R.M.; Thompson, E.W.; Roberts-Thomson, S.J.; Monteith, G.R. Hypoxia-induced reactive oxygen species mediate N-cadherin and SERPINE1 expression, EGFR signalling and motility in MDA-MB-468 breast cancer cells. Sci. Rep. 2017, 7, 15140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobritoiu, M.; Stepan, A.E.; Margaritescu, C.; Simionescu, C.E.; Vere, C.C.; Schenker, M.; Crisan, A.E. Immunoexpression of E-cadherin, P-cadherin and fibronectin in gastric carcinomas. Rom. J. Morphol. Embryol. Rev. Roum. Morphol. Embryol. 2019, 60, 573–579. [Google Scholar]
- Zhan, Y.; Chen, Z.; Li, Y.; He, A.; He, S.; Gong, Y.; Li, X.; Zhou, L. Long non-coding RNA DANCR promotes malignant phenotypes of bladder cancer cells by modulating the miR-149/MSI2 axis as a ceRNA. J. Exp. Clin. Cancer Res. CR 2018, 37, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.L.; Wang, C.; Liu, W.; Ai, Z.L. Emerging roles of the long non-coding RNA 01296/microRNA-143-3p/MSI2 axis in development of thyroid cancer. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Yeh, Y.; Guo, Q.; Connelly, Z.; Cheng, S.; Yang, S.; Prieto-Dominguez, N.; Yu, X. Wnt/Beta-Catenin Signaling and Prostate Cancer Therapy Resistance. Adv. Exp. Med. Biol. 2019, 1210, 351–378. [Google Scholar] [CrossRef]
- Duggimpudi, S.; Kloetgen, A.; Maney, S.K.; Munch, P.C.; Hezaveh, K.; Shaykhalishahi, H.; Hoyer, W.; McHardy, A.C.; Lang, P.A.; Borkhardt, A.; et al. Transcriptome-wide analysis uncovers the targets of the RNA-binding protein MSI2 and effects of MSI2’s RNA-binding activity on IL-6 signaling. J. Biol. Chem. 2018, 293, 15359–15369. [Google Scholar] [CrossRef] [Green Version]
- Sheng, W.; Shi, X.; Lin, Y.; Tang, J.; Jia, C.; Cao, R.; Sun, J.; Wang, G.; Zhou, L.; Dong, M. Musashi2 promotes EGF-induced EMT in pancreatic cancer via ZEB1-ERK/MAPK signaling. J. Exp. Clin. Cancer Res. CR 2020, 39, 16. [Google Scholar] [CrossRef] [Green Version]
- Brun, J.; Dieudonne, F.X.; Marty, C.; Muller, J.; Schule, R.; Patino-Garcia, A.; Lecanda, F.; Fromigue, O.; Marie, P.J. FHL2 silencing reduces Wnt signaling and osteosarcoma tumorigenesis in vitro and in vivo. PLoS ONE 2013, 8, e55034. [Google Scholar] [CrossRef] [Green Version]
- Zienert, E.; Eke, I.; Aust, D.; Cordes, N. LIM-only protein FHL2 critically determines survival and radioresistance of pancreatic cancer cells. Cancer Lett. 2015, 364, 17–24. [Google Scholar] [CrossRef]
- Ding, L.; Wang, Z.; Yan, J.; Yang, X.; Liu, A.; Qiu, W.; Zhu, J.; Han, J.; Zhang, H.; Lin, J.; et al. Human four-and-a-half LIM family members suppress tumor cell growth through a TGF-beta-like signaling pathway. J. Clin. Investig. 2009, 119, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.L.; Liang, S.Q.; Fu, W.; Zhu, B.K.; Li, S.Z.; Han, H.; Qin, H.Y. The LIM domain protein FHL1C interacts with tight junction protein ZO-1 contributing to the epithelial-mesenchymal transition (EMT) of a breast adenocarcinoma cell line. Gene 2014, 542, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Deng, X.; Zhao, Y.; Deng, B.; Deng, J.; Fan, H.; Du, Y.; Hao, L. Ubiquitin-like protein FAT10 promotes osteosarcoma growth by modifying the ubiquitination and degradation of YAP1. Exp. Cell Res. 2020, 387, 111804. [Google Scholar] [CrossRef] [PubMed]
- Zanconato, F.; Cordenonsi, M.; Piccolo, S. YAP/TAZ at the Roots of Cancer. Cancer Cell 2016, 29, 783–803. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-H.; Wu, H.-Y.; Gao, J.; Wang, X.-H.; Gao, T.-H.; Zhang, S.-F. IGF1R facilitates epithelial-mesenchymal transition and cancer stem cell properties in neuroblastoma via the STAT3/AKT axis. Cancer Manag. Res. 2019, 11, 5459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokota, T.; Nagai, H.; Harada, H.; Mine, N.; Terada, Y.; Fujiwara, H.; Yabe, A.; Miyazaki, K.; Emi, M. Identification, tissue expression, and chromosomal position of a novel gene encoding human ubiquitin-conjugating enzyme E2-230k. Gene 2001, 267, 95–100. [Google Scholar] [CrossRef]
- Xing, Z.; Li, S.; Liu, Z.; Zhang, C.; Bai, Z. CTCF-induced upregulation of HOXA11-AS facilitates cell proliferation and migration by targeting miR-518b/ACTN4 axis in prostate cancer. Prostate 2020. [Google Scholar] [CrossRef]
- Georgakilas, A.G.; Tsantoulis, P.; Kotsinas, A.; Michalopoulos, I.; Townsend, P.; Gorgoulis, V.G. Are common fragile sites merely structural domains or highly organized “functional” units susceptible to oncogenic stress? Cell. Mol. Life Sci. 2014, 71, 4519–4544. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, B.; Zhang, P.; Lian, L.; Li, L.; Qiu, Z.; Qian, K.; Chen, A.; Liu, Q.; Jiang, Y.; et al. Norcantharidin regulates ERalpha signaling and tamoxifen resistance via targeting miR-873/CDK3 in breast cancer cells. PLoS ONE 2019, 14, e0217181. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Chen, Y.; Li, F.; Bao, L.; Liu, W. Atezolizumab and Bevacizumab attenuate cisplatin resistant ovarian cancer cells progression synergistically via suppressing epithelial-mesenchymal transition. Front. Immunol. 2019, 10, 867. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Yu, M.; Yang, R.; Zhang, L.; Zhang, L.; Zhu, D.; Luo, H.; Hong, Y.; Yu, T.; Sun, J. A PTAL-miR-101-FN1 axis promotes EMT and invasion-metastasis in serous ovarian cancer. Mol. Ther. Oncolytics 2020, 16, 53–62. [Google Scholar] [CrossRef]
- Montanuy, H.; Martinez-Barriocanal, A.; Casado, J.A.; Rovirosa, L.; Ramirez, M.J.; Nieto, R.; Carrascoso-Rubio, C.; Riera, P.; Gonzalez, A.; Lerma, E.; et al. Gefitinib and afatinib show potential efficacy for Fanconi anemia-related head and neck cancer. Clin. Cancer Res. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chih-Hsin Yang, J.; Schuler, M.; Popat, S.; Miura, S.; Heeke, S.; Park, K.; Marten, A.; Kim, E.S. Afatinib for the Treatment of Non-Small Cell Lung Cancer Harboring Uncommon EGFR Mutations: A Database of 693 Cases. J. Thorac. Oncol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Poh, M.E.; Liam, C.K.; Rajadurai, P.; Chai, C.S. Epithelial-to-mesenchymal transition (EMT) causing acquired resistance to afatinib in a patient with epidermal growth factor receptor (EGFR)-mutant lung adenocarcinoma. J. Thorac. Dis. 2018, 10, E560–E563. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.F.; Wang, A.Y.; Liu, J. HIFs-MiR-33a-Twsit1 axis can regulate invasiveness of hepatocellular cancer cells. Eur. Rev. Med Pharmacol. Sci. 2016, 20, 3011–3016. [Google Scholar] [PubMed]
- Ryan, M.R.; Sohl, C.D.; Luo, B.; Anderson, K.S. The FGFR1 V561M Gatekeeper Mutation Drives AZD4547 Resistance through STAT3 Activation and EMT. Mol. Cancer Res. MCR 2019, 17, 532–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowinsky, E.K.; Donehower, R.C. Paclitaxel (taxol). N. Engl. J. Med. 1995, 332, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Chen, L. Progress in research on paclitaxel and tumor immunotherapy. Cell. Mol. Biol. Lett. 2019, 24, 40. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Xu, Q.; Dong, Y.; Li, G.; Yang, L.; Wang, L.; Li, H. MiR-181a upregulation is associated with epithelial-to-mesenchymal transition (EMT) and multidrug resistance (MDR) of ovarian cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2004–2010. [Google Scholar]
- Jing, L.; Bo, W.; Yourong, F.; Tian, W.; Shixuan, W.; Mingfu, W. Sema4C mediates EMT inducing chemotherapeutic resistance of miR-31-3p in cervical cancer cells. Sci. Rep. 2019, 9, 17727. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, F.; Cui, J.Y.; Chen, L.; Chen, Y.T.; Liu, B.W. CAFs enhance paclitaxel resistance by inducing EMT through the IL6/JAK2/STAT3 pathway. Oncol. Rep. 2018, 39, 2081–2090. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Zhang, J.; Liu, M.; Huang, Y.; Yin, L. SMAD3 inducing the transcription of STYK1 to promote the EMT process and improve the tolerance of ovarian carcinoma cells to paclitaxel. J. Cell. Biochem. 2019, 120, 10796–10811. [Google Scholar] [CrossRef]
- Sale, M.J.; Balmanno, K.; Saxena, J.; Ozono, E.; Wojdyla, K.; McIntyre, R.E.; Gilley, R.; Woroniuk, A.; Howarth, K.D.; Hughes, G.; et al. MEK1/2 inhibitor withdrawal reverses acquired resistance driven by BRAF(V600E) amplification whereas KRAS(G13D) amplification promotes EMT-chemoresistance. Nat. Commun. 2019, 10, 2030. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.S.; Jalgaonkar, S.P.; Middleton, J.D.; Hai, T. Stress-inducible gene Atf3 in the noncancer host cells contributes to chemotherapy-exacerbated breast cancer metastasis. Proc. Natl. Acad. Sci. USA 2017, 114, E7159–E7168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Palma, M.; Lewis, C.E. Macrophage regulation of tumor responses to anticancer therapies. Cancer Cell 2013, 23, 277–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagiannis, G.S.; Pastoriza, J.M.; Wang, Y.; Harney, A.S.; Entenberg, D.; Pignatelli, J.; Sharma, V.P.; Xue, E.A.; Cheng, E.; D’Alfonso, T.M.; et al. Neoadjuvant chemotherapy induces breast cancer metastasis through a TMEM-mediated mechanism. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffell, B.; Coussens, L.M. Macrophages and therapeutic resistance in cancer. Cancer Cell 2015, 27, 462–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.; Pluddemann, A.; Martinez Estrada, F. Macrophage heterogeneity in tissues: Phenotypic diversity and functions. Immunol. Rev. 2014, 262, 36–55. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, X.; Li, X.; Fan, Y.; Li, G.; Guo, C.; Zhu, F.; Zhang, L.; Shi, Y. CD68+HLA-DR+ M1-like macrophages promote motility of HCC cells via NF-κB/FAK pathway. Cancer Lett. 2014, 345, 91–99. [Google Scholar] [CrossRef]
- Cho, U.; Kim, B.; Kim, S.; Han, Y.; Song, Y.S. Pro-inflammatory M1 macrophage enhances metastatic potential of ovarian cancer cells through NF-κB activation. Mol. Carcinog. 2018, 57, 235–242. [Google Scholar] [CrossRef]
- Xiao, M.; Zhang, J.; Chen, W.; Chen, W. M1-like tumor-associated macrophages activated by exosome-transferred THBS1 promote malignant migration in oral squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 143. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wang, W.; Wang, X.; Xu, C.; Zhang, N.; Di, W. Cisplatin-stimulated macrophages promote ovarian cancer migration via the CCL20-CCR6 axis. Cancer Lett. 2020, 472, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.J.; Lin, S.Z.; Zhou, L.; Xie, H.Y.; Zhou, W.H.; Taki-Eldin, A.; Zheng, S.S. Selective recruitment of regulatory T cell through CCR6-CCL20 in hepatocellular carcinoma fosters tumor progression and predicts poor prognosis. PLoS ONE 2011, 6, e24671. [Google Scholar] [CrossRef]
- Kryczek, I.; Lin, Y.; Nagarsheth, N.; Peng, D.; Zhao, L.; Zhao, E.; Vatan, L.; Szeliga, W.; Dou, Y.; Owens, S.; et al. IL-22+CD4+ T Cells Promote Colorectal Cancer Stemness via STAT3 Transcription Factor Activation and Induction of the Methyltransferase DOT1L. Immunity 2014, 40, 772–784. [Google Scholar] [CrossRef] [Green Version]
- Walch-Ruckheim, B.; Mavrova, R.; Henning, M.; Vicinus, B.; Kim, Y.J.; Bohle, R.M.; Juhasz-Boss, I.; Solomayer, E.F.; Smola, S. Stromal Fibroblasts Induce CCL20 through IL6/C/EBPbeta to Support the Recruitment of Th17 Cells during Cervical Cancer Progression. Cancer Res. 2015, 75, 5248–5259. [Google Scholar] [CrossRef] [Green Version]
- Ranasinghe, R.; Eri, R. Modulation of the CCR6-CCL20 Axis: A Potential Therapeutic Target in Inflammation and Cancer. Medicina 2018, 54, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, J.L.; Group, W.S.C.; Concannon, P. ATM, radiation, and the risk of second primary breast cancer. Int. J. Radiat. Biol. 2017, 93, 1121–1127. [Google Scholar] [CrossRef] [PubMed]
- Nanda, N.; Roberts, N.J. ATM Serine/Threonine Kinase and its Role in Pancreatic Risk. Genes (Basel) 2020, 11, 108. [Google Scholar] [CrossRef] [Green Version]
- Murai, J.; Thomas, A.; Miettinen, M.; Pommier, Y. Schlafen 11 (SLFN11), a restriction factor for replicative stress induced by DNA-targeting anti-cancer therapies. Pharmacol. Ther. 2019, 201, 94–102. [Google Scholar] [CrossRef]
- Allison Stewart, C.; Tong, P.; Cardnell, R.J.; Sen, T.; Li, L.; Gay, C.M.; Masrorpour, F.; Fan, Y.; Bara, R.O.; Feng, Y.; et al. Dynamic variations in epithelial-to-mesenchymal transition (EMT), ATM, and SLFN11 govern response to PARP inhibitors and cisplatin in small cell lung cancer. Oncotarget 2017, 8, 28575–28587. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Yu, L.; Mei, H.; Yang, J.; Gao, T.; Cheng, A.; Guo, W.; Xia, K.; Liu, G. Cisplatin promotes mesenchymal-like characteristics in osteosarcoma through Snail. Oncol. Lett. 2016, 12, 5007–5014. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Zhang, G.A.; Zhang, B.C.; Wang, Y.; Liu, Z.; Jiao, Y.L.; Liu, N.; Zhao, Y.R. Short low concentration cisplatin treatment leads to an epithelial mesenchymal transition-like response in DU145 prostate cancer cells. Asian Pac. J. Cancer Prev. APJCP 2015, 16, 1025–1028. [Google Scholar] [CrossRef] [PubMed]
- Miow, Q.H.; Tan, T.Z.; Ye, J.; Lau, J.A.; Yokomizo, T.; Thiery, J.P.; Mori, S. Epithelial-mesenchymal status renders differential responses to cisplatin in ovarian cancer. Oncogene 2015, 34, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, M.; Kang, R.; Klionsky, D.J.; Tang, D. Autophagic degradation of the circadian clock regulator promotes ferroptosis. Autophagy 2019, 15, 2033–2035. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jin, M.; Yao, Z.; Bernard, A.; Klionsky, D.J. Bidirectional roles of Dhh1 in regulating autophagy. Autophagy 2019, 15, 1838–1839. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J. Autophagy participates in, well, just about everything. Cell Death Differ. 2020. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Klionsky, D.J. Autophagy and disease: Unanswered questions. Cell Death Differ. 2020. [Google Scholar] [CrossRef]
- Tyutyunyk-Massey, L.; Gewirtz, D.A. Roles of autophagy in breast cancer treatment: Target, bystander or benefactor. Semin. Cancer Biol. 2019. [Google Scholar] [CrossRef]
- Towers, C.G.; Fitzwalter, B.E.; Regan, D.; Goodspeed, A.; Morgan, M.J.; Liu, C.-W.; Gustafson, D.L.; Thorburn, A. Cancer Cells Upregulate NRF2 Signaling to Adapt to Autophagy Inhibition. Dev. Cell 2019, 50, 690–703.e696. [Google Scholar] [CrossRef]
- Kocaturk, N.M.; Akkoc, Y.; Kig, C.; Bayraktar, O.; Gozuacik, D.; Kutlu, O. Autophagy as a molecular target for cancer treatment. Eur. J. Pharm. Sci. 2019, 134, 116–137. [Google Scholar] [CrossRef]
- Quan, Y.; Lei, H.; Wahafu, W.; Liu, Y.; Ping, H.; Zhang, X. Inhibition of autophagy enhances the anticancer effect of enzalutamide on bladder cancer. Biomed. Pharmacother. 2019, 120, 109490. [Google Scholar] [CrossRef]
- Lei, Y.; Tang, L.; Hu, J.; Wang, S.; Liu, Y.; Yang, M.; Zhang, J.; Tang, B. Inhibition of MGMT-mediated autophagy suppression decreases cisplatin chemosensitivity in gastric cancer. Biomed. Pharmacother. 2020, 125, 109896. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; McWilliams, T.G. Monitoring autophagy in cancer: From bench to bedside. Semin. Cancer Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Li, G.; Liu, C.; Ren, S.; Deng, T.; Zhang, S.; Tian, Y.; Liu, Y.; Qiu, Y. Autophagy inhibition impairs the epithelial-mesenchymal transition and enhances cisplatin sensitivity in nasopharyngeal carcinoma. Oncol. Lett. 2017, 13, 4147–4154. [Google Scholar] [CrossRef] [Green Version]
- Mina-Osorio, P. The moonlighting enzyme CD13: Old and new functions to target. Trends Mol. Med. 2008, 14, 361–371. [Google Scholar] [CrossRef]
- Carl-McGrath, S.; Lendeckel, U.; Ebert, M.; Wolter, A.-B.; Roessner, A.; Röcken, C. The ectopeptidases CD10, CD13, CD26, and CD143 are upregulated in gastric cancer. Int. J. Oncol. 2004, 25, 1223–1232. [Google Scholar] [CrossRef]
- Ben-Porath, I.; Kozak, C.A.; Benvenisty, N. Chromosomal Mapping ofTmp (Emp1), Xmp (Emp2), andYmp (Emp3), Genes Encoding Membrane Proteins Related toPmp22. Genomics 1998, 49, 443–447. [Google Scholar] [CrossRef]
- Mikata, R.; Yokosuka, O.; Fukai, K.; Imazeki, F.; Arai, M.; Tada, M.; Kurihara, T.; Zhang, K.; Kanda, T.; Saisho, H. Analysis of genes upregulated by the demethylating agent 5-aza-2′-deoxycytidine in gastric cancer cell lines. Int. J. Cancer 2006, 119, 1616–1622. [Google Scholar] [CrossRef]
- Guo, Q.; Jing, F.J.; Xu, W.; Li, X.; Li, X.; Sun, J.L.; Xing, X.M.; Zhou, C.K.; Jing, F.B. Ubenimex induces autophagy inhibition and EMT suppression to overcome cisplatin resistance in GC cells by perturbing the CD13/EMP3/PI3K/AKT/NF-kappaB axis. Aging 2019, 12, 80. [Google Scholar] [CrossRef]
- Cocucci, E.; Meldolesi, J. Ectosomes and exosomes: Shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015, 25, 364–372. [Google Scholar] [CrossRef]
- Kahlert, C.; Kalluri, R. Exosomes in tumor microenvironment influence cancer progression and metastasis. J. Mol. Med. 2013, 91, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Qu, L.; Ding, J.; Chen, C.; Wu, Z.-J.; Liu, B.; Gao, Y.; Chen, W.; Liu, F.; Sun, W.; Li, X.-F. Exosome-transmitted lncARSR promotes sunitinib resistance in renal cancer by acting as a competing endogenous RNA. Cancer Cell 2016, 29, 653–668. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Yang, M.; Ren, Y.; Wu, C.; Wang, L. Exosomes mediated transfer of lncRNA UCA1 results in increased tamoxifen resistance in breast cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4362–4368. [Google Scholar] [PubMed]
- Wang, J.; Lv, B.; Su, Y.; Wang, X.; Bu, J.; Yao, L. Exosome-Mediated Transfer of lncRNA HOTTIP Promotes Cisplatin Resistance in Gastric Cancer Cells by Regulating HMGA1/miR-218 Axis. OncoTargets Ther. 2019, 12, 11325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.; Wang, Z.; Mijiti, M.; Du, G.; Li, Y.; Dangmurenjiafu, G. MiR-28-5p promotes human glioblastoma cell growth through inactivation of FOXO1. Int. J. Clin. Exp. Pathol. 2019, 12, 2972. [Google Scholar]
- Xie, C.; Guo, Y.; Lou, S. LncRNA ANCR Promotes Invasion and Migration of Gastric Cancer by Regulating FoxO1 Expression to Inhibit Macrophage M1 Polarization. Dig. Dis. Sci. 2020, 1–10. [Google Scholar] [CrossRef]
- Zhao, M.; Luo, R.; Liu, Y.; Gao, L.; Fu, Z.; Fu, Q.; Luo, X.; Chen, Y.; Deng, X.; Liang, Z. miR-3188 regulates nasopharyngeal carcinoma proliferation and chemosensitivity through a FOXO1-modulated positive feedback loop with mTOR–p-PI3K/AKT-c-JUN. Nat. Commun. 2016, 7, 11309. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, X.; Lin, X.; Zhao, M.; Xiao, Y.; Liu, C.; Liang, Z.; Lin, Z.; Yi, R.; Tang, Z.; et al. Chemical compound cinobufotalin potently induces FOXO1-stimulated cisplatin sensitivity by antagonizing its binding partner MYH9. Signal Transduct. Target. Ther. 2019, 4, 48. [Google Scholar] [CrossRef]
- Zhang, F.; Yin, Y.; Xu, T. Cinobufotalin injection combined with chemotherapy for the treatment of advanced NSCLC in China: A PRISMA-compliant meta-analysis of 29 randomized controlled trials. Medicine 2019, 98, e16969. [Google Scholar] [CrossRef]
- Mirzaei, H.; Momeni, F.; Saadatpour, L.; Sahebkar, A.; Goodarzi, M.; Masoudifar, A.; Kouhpayeh, S.; Salehi, H.; Mirzaei, H.R.; Jaafari, M.R. MicroRNA: Relevance to stroke diagnosis, prognosis, and therapy. J. Cell. Physiol. 2018, 233, 856–865. [Google Scholar] [CrossRef]
- Moridikia, A.; Mirzaei, H.; Sahebkar, A.; Salimian, J. MicroRNAs: Potential candidates for diagnosis and treatment of colorectal cancer. J. Cell. Physiol. 2018, 233, 901–913. [Google Scholar] [CrossRef]
- Hermann, H.; Runnel, T.; Aab, A.; Baurecht, H.; Rodriguez, E.; Magilnick, N.; Urgard, E.; Šahmatova, L.; Prans, E.; Maslovskaja, J. miR-146b probably assists miRNA-146a in the suppression of keratinocyte proliferation and inflammatory responses in psoriasis. J. Investig. Dermatol. 2017, 137, 1945–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, X.; Wu, B.; Xiao, K.; Kang, J.; Xie, J.; Zhang, X.; Fan, Y. MiR-146b-5p promotes metastasis and induces epithelial-mesenchymal transition in thyroid cancer by targeting ZNRF3. Cell. Physiol. Biochem. 2015, 35, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Balducci, E.; Leroyer, A.S.; Lacroix, R.; Robert, S.; Todorova, D.; Simoncini, S.; Lyonnet, L.; Chareyre, C.; Zaegel-Faucher, O.; Micallef, J. Microvesicles from T cells overexpress miR-146b-5p in HIV-1 infection and repress endothelial activation. J. Extracell. Vesicles 2018, 7, 71–72. [Google Scholar]
- Cho, S.; Lee, H.-M.; Yu, I.-S.; Choi, Y.S.; Huang, H.-Y.; Hashemifar, S.S.; Lin, L.-L.; Chen, M.-C.; Afanasiev, N.D.; Khan, A.A. Differential cell-intrinsic regulations of germinal center B and T cells by miR-146a and miR-146b. Nat. Commun. 2018, 9, 2757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labbé, D.P.; Tremblay, M.L. PTP1B: From Metabolism to Cancer. In Protein Tyrosine Phosphatases in Cancer; Springer: Berlin/Heidelberg, Germany, 2016; pp. 169–199. [Google Scholar]
- Wang, N.; She, J.; Liu, W.; Shi, J.; Yang, Q.; Shi, B.; Hou, P. Frequent amplification of PTP1B is associated with poor survival of gastric cancer patients. Cell Cycle 2015, 14, 732–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Q.; Cheng, P.; Yang, H.; Liang, H.; Lin, F. miR-146b Reverses epithelial-mesenchymal transition via targeting PTP1B in cisplatin-resistance human lung adenocarcinoma cells. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef]
- Bian, W.G.; Zhou, X.N.; Song, S.; Chen, H.T.; Shen, Y.; Chen, P. Reduced miR-363-3p expression in non-small cell lung cancer is associated with gemcitabine resistance via targeting of CUL4A. Eur. Rev. Med Pharmacol. Sci. 2019, 23, 649–659. [Google Scholar] [CrossRef]
- Wang, X.; Ren, M.; Li, Y.; Hu, J.; Lu, G.; Ma, W.; Guo, D.; Lu, X.; He, S. Long noncoding RNA NNT-AS1 promotes gastric cancer proliferation and invasion by regulating microRNA-363 expression. J. Cell. Biochem. 2019, 120, 5704–5712. [Google Scholar] [CrossRef]
- Cao, L.; Wan, Q.; Li, F.; Tang, C.E. MiR-363 inhibits cisplatin chemoresistance of epithelial ovarian cancer by regulating snail-induced epithelial-mesenchymal transition. BMB Rep. 2018, 51, 456. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, H.; Takahashi, R.U.; Prieto-Vila, M.; Kawamura, Y.; Kondo, S.; Shirota, T.; Ochiya, T. CD44 exerts a functional role during EMT induction in cisplatin-resistant head and neck cancer cells. Oncotarget 2018, 9, 10029–10041. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Wu, H.; Fan, M.; Yu, R.; Zhang, Y.; Liu, J.; Zhou, X.; Cai, Y.; Huang, S.; Hu, Z.; et al. Sodium butyrate inhibits migration and induces AMPK-mTOR pathway-dependent autophagy and ROS-mediated apoptosis via the miR-139-5p/Bmi-1 axis in human bladder cancer cells. FASEB J. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Li, Y.; Wen, J.; Qi, T.; Wang, Y. Long non-coding RNA TTN-AS1 promotes tumorigenesis of ovarian cancer through modulating the miR-139-5p/ROCK2 axis. Biomed. Pharmacother. Biomed. Pharmacother. 2020, 125, 109882. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Wen, D.Y.; Que, Q.; Zhou, C.Y.; Wang, X.D.; Peng, Y.T.; He, Y.; Yang, H.; Liao, B.M. Reduced expression of microRNA-139-5p in hepatocellular carcinoma results in a poor outcome: An exploration the roles of microRNA-139-5p in tumorigenesis, advancement and prognosis at the molecular biological level using an integrated meta-analysis and bioinformatic investigation. Oncol. Lett. 2019, 18, 6704–6724. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Guo, W.; Ren, K.; Li, W.; Jiang, Y.; Sun, J.; Dai, W.; Zhao, W. LncRNA AFAP1-AS1 Supresses miR-139-5p and Promotes Cell Proliferation and Chemotherapy Resistance of Non-small Cell Lung Cancer by Competitively Upregulating RRM2. Front. Oncol. 2019, 9, 1103. [Google Scholar] [CrossRef] [Green Version]
- Shao, Q.; Zhang, P.; Ma, Y.; Lu, Z.; Meng, J.; Li, H.; Wang, X.; Chen, D.; Zhang, M.; Han, Y.; et al. MicroRNA-139-5p affects cisplatin sensitivity in human nasopharyngeal carcinoma cells by regulating the epithelial-to-mesenchymal transition. Gene 2018, 652, 48–58. [Google Scholar] [CrossRef]
- Nieto, M.A. Epithelial plasticity: A common theme in embryonic and cancer cells. Science 2013, 342, 1234850. [Google Scholar] [CrossRef] [Green Version]
- Meacham, C.E.; Morrison, S.J. Tumour heterogeneity and cancer cell plasticity. Nature 2013, 501, 328. [Google Scholar] [CrossRef] [Green Version]
- Najafi, M.; Mortezaee, K.; Majidpoor, J. Cancer stem cell (CSC) resistance drivers. Life Sci. 2019, 234, 116781. [Google Scholar] [CrossRef]
- Najafi, M.; Farhood, B.; Mortezaee, K. Cancer stem cells (CSCs) in cancer progression and therapy. J. Cell. Physiol. 2019, 234, 8381–8395. [Google Scholar] [CrossRef]
- Saha, S.; Mukherjee, S.; Khan, P.; Kajal, K.; Mazumdar, M.; Manna, A.; Mukherjee, S.; De, S.; Jana, D.; Sarkar, D.K. Aspirin suppresses the acquisition of chemoresistance in breast cancer by disrupting an NFκB–IL6 signaling axis responsible for the generation of cancer stem cells. Cancer Res. 2016, 76, 2000–2012. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.F.; Hunter, R.W.; Hers, I. mTORC2 protein-mediated protein kinase B (Akt) serine 473 phosphorylation is not required for Akt1 activity in human platelets. J. Biol. Chem. 2011, 286, 24553–24560. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.S.; Yap, W.N.; Arfuso, F.; Kar, S.; Wang, C.; Cai, W.; Dharmarajan, A.M.; Sethi, G.; Kumar, A.P. Targeting the PI3K/Akt signaling pathway in gastric carcinoma: A reality for personalized medicine? World J. Gastroenterol. 2015, 21, 12261–12273. [Google Scholar] [CrossRef] [PubMed]
- Siveen, K.S.; Ahn, K.S.; Ong, T.H.; Shanmugam, M.K.; Li, F.; Yap, W.N.; Kumar, A.P.; Fong, C.W.; Tergaonkar, V.; Hui, K.M.; et al. Y-tocotrienol inhibits angiogenesis-dependent growth of human hepatocellular carcinoma through abrogation of AKT/mTOR pathway in an orthotopic mouse model. Oncotarget 2014, 5, 1897–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, P.S.; Wang, L.Z.; Dai, X.; Tseng, S.H.; Loo, S.J.; Sethi, G. Judicious Toggling of mTOR Activity to Combat Insulin Resistance and Cancer: Current Evidence and Perspectives. Front. Pharmacol. 2016, 7, 395. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, C.; Um, J.Y.; Sethi, G.; Ahn, K.S. Casticin-Induced Inhibition of Cell Growth and Survival Are Mediated through the Dual Modulation of Akt/mTOR Signaling Cascade. Cancers 2019, 11, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, Y.; Ji, C.; Yang, B.; Yang, Z.; Gu, H.; Lu, C.-C.; Wang, R.; Su, Z.-L.; Chen, B.; Sun, W.-L. DNA-dependent protein kinase catalytic subunit (DNA-PKcs)-SIN1 association mediates ultraviolet B (UVB)-induced Akt Ser-473 phosphorylation and skin cell survival. Mol. Cancer 2013, 12, 172. [Google Scholar] [CrossRef] [Green Version]
- Sunayama, J.; Matsuda, K.I.; Sato, A.; Tachibana, K.; Suzuki, K.; Narita, Y.; Shibui, S.; Sakurada, K.; Kayama, T.; Tomiyama, A. Crosstalk between the PI3K/mTOR and MEK/ERK pathways involved in the maintenance of self-renewal and tumorigenicity of glioblastoma stem-like cells. Stem Cells 2010, 28, 1930–1939. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.-H.; Wang, J.-B.; Lin, M.-Q.; Zhang, P.-Y.; Liu, L.-C.; Lin, J.-X.; Lu, J.; Chen, Q.-Y.; Cao, L.-L.; Lin, M. CDK5RAP3 suppresses Wnt/β-catenin signaling by inhibiting AKT phosphorylation in gastric cancer. J. Exp. Clin. Cancer Res. 2018, 37, 59. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Li, W.; Wang, Y.; Liu, Z.; Cai, Y.; Xie, T.; Shi, M.; Wang, Z.; Jiang, B. Glycogen synthase kinase-3 beta regulates Snail and β-catenin expression during Fas-induced epithelial–mesenchymal transition in gastrointestinal cancer. Eur. J. Cancer 2013, 49, 2734–2746. [Google Scholar] [CrossRef]
- Yuan, L.; Zhang, K.; Zhou, M.M.; Wasan, H.S.; Tao, F.F.; Yan, Q.Y.; Feng, G.; Tang, Y.S.; Shen, M.H.; Ma, S.L.; et al. Jiedu Sangen Decoction Reverses Epithelial-to-mesenchymal Transition and Inhibits Invasion and Metastasis of Colon Cancer via AKT/GSK-3beta Signaling Pathway. J. Cancer 2019, 10, 6439–6456. [Google Scholar] [CrossRef]
- Shin, J.H.; Kim, K.M.; Jeong, J.U.; Shin, J.M.; Kang, J.H.; Bang, K.; Kim, J.H. Nrf2-Heme Oxygenase-1 Attenuates High-Glucose-Induced Epithelial-to-Mesenchymal Transition of Renal Tubule Cells by Inhibiting ROS-Mediated PI3K/Akt/GSK-3beta Signaling. J. Diabetes Res. 2019, 2019, 2510105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, P.; Bhattacharya, A.; Sengupta, D.; Banerjee, S.; Adhikary, A.; Das, T. Aspirin enhances cisplatin sensitivity of resistant non-small cell lung carcinoma stem-like cells by targeting mTOR-Akt axis to repress migration. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bordonaro, M. Hypothesis: Retinoblastoma protein inactivation mediates effects of histone deacetylase inhibitor-induced Wnt hyperactivation in colorectal cancer cells. J. Cancer 2020, 11, 668–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarman, E.J.; Boulter, L. Targeting the Wnt signalling pathway: The challenge of reducing scarring without affecting repair. Expert Opin. Investig. Drugs 2020. [Google Scholar] [CrossRef]
- Ong, M.S.; Cai, W.; Yuan, Y.; Leong, H.C.; Tan, T.Z.; Mohammad, A.; You, M.L.; Arfuso, F.; Goh, B.C.; Warrier, S.; et al. ’Lnc’-ing Wnt in female reproductive cancers: Therapeutic potential of long non-coding RNAs in Wnt signalling. Br. J. Pharmacol. 2017, 174, 4684–4700. [Google Scholar] [CrossRef] [Green Version]
- Bhuvanalakshmi, G.; Basappa; Rangappa, K.S.; Dharmarajan, A.; Sethi, G.; Kumar, A.P.; Warrier, S. Breast Cancer Stem-Like Cells Are Inhibited by Diosgenin, a Steroidal Saponin, by the Attenuation of the Wnt beta-Catenin Signaling via the Wnt Antagonist Secreted Frizzled Related Protein-4. Front. Pharmacol. 2017, 8, 124. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.; Pu, Y.; Gou, R.; Chen, Y.; Ren, X.; Liu, W.; Dong, P. Silencing of RIPK4 inhibits epithelialmesenchymal transition by inactivating the Wnt/betacatenin signaling pathway in osteosarcoma. Mol. Med. Rep. 2020. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Sun, W.; Zhang, L.; Li, J. Silencing Of MAGI1 Promotes The Proliferation And Inhibits Apoptosis Of Glioma Cells Via The Wnt/beta-Catenin And PTEN/AKT Signaling Pathways. OncoTargets Ther. 2019, 12, 9639–9650. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Xu, Y.; Yang, G.; Li, S.; Rui, Z. Knockdown Of Long Non-Coding RNA TP73-AS1 Inhibited Cell Proliferation And Metastasis Through Wnt/beta-Catenin Pathway In Lung Adenocarcinoma. OncoTargets Ther. 2019, 12, 9599–9610. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Huang, Z.; Zhou, R.; An, H.; Cao, G.; Ye, J.; Huang, C.; Wu, D. Numb negatively regulates the epithelial-to-mesenchymal transition in colorectal cancer through the Wnt signalling pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2020. [Google Scholar] [CrossRef]
- Wu, W.; Guo, L.; Liang, Z.; Liu, Y.; Yao, Z. Lnc-SNHG16/miR-128 axis modulates malignant phenotype through WNT/beta-catenin pathway in cervical cancer cells. J. Cancer 2020, 11, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhao, Z.; Sun, Z.; Liu, C.; Cheng, X.; Ruge, F.; Yang, Y.; Jiang, W.G.; Ye, L. Increased expression of Psoriasin is correlated with poor prognosis of bladder transitional cell carcinoma by promoting invasion and proliferation. Oncol. Rep. 2020, 43, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Fang, S.; Tian, H.; Zhou, C.; Zhao, X.; Tian, H.; He, J.; Shen, W.; Meng, X.; Jin, X.; et al. lncRNA JPX/miR-33a-5p/Twist1 axis regulates tumorigenesis and metastasis of lung cancer by activating Wnt/beta-catenin signaling. Mol. Cancer 2020, 19, 9. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Liu, Z.; Gao, J.; Wang, Q. MiR-338-3p Enhances Ovarian Cancer Cell Sensitivity to Cisplatin by Downregulating WNT2B. Yonsei Med. J. 2019, 60, 1146–1156. [Google Scholar] [CrossRef]
- Worthmuller, J.; Salicio, V.; Oberson, A.; Blum, W.; Schwaller, B. Modulation of Calretinin Expression in Human Mesothelioma Cells Reveals the Implication of the FAK and Wnt Signaling Pathways in Conferring Chemoresistance towards Cisplatin. Int. J. Mol. Sci. 2019, 20, 5391. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Jiang, Q.; Liu, X.; Lin, X.; Li, Y.; Tang, Z.; Liu, C.; Zhao, M.; Li, X.; Liu, Z. Cinobufotalin Powerfully Reversed EBV-miR-BART22-Induced Cisplatin Resistance via Stimulating MAP2K4 to Antagonize Non-Muscle Myosin Heavy Chain IIA/Glycogen Synthase 3β/β-Catenin Signaling Pathway. EBioMedicine 2019, 48, 386–404. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.H.; Grove, C.L.; Baker, R. Development of oculomotor circuitry independent of hox3 genes. Nat. Commun. 2014, 5, 4221. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Chen, J.; Qian, W.; Kang, J.; Wang, J.; Jiang, L.; Qiao, L.; Chen, W.; Zhang, J. Integrated long non-coding RNA analyses identify novel regulators of epithelial-mesenchymal transition in the mouse model of pulmonary fibrosis. J. Cell. Mol. Med. 2016, 20, 1234–1246. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Gu, X.; Wu, N.; Zhang, P.; Liu, Y.; Jiang, S. Human antigen R enhances the epithelial-mesenchymal transition via regulation of ZEB-1 in the human airway epithelium. Respir. Res. 2018, 19, 109. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Wang, M.; Dai, Y.; Bao, D.; Zhang, J.; Pan, H. LncRNA HOXA-AS3 Sponges miR-29c to Facilitate Cell Proliferation, Metastasis, and EMT Process and Activate the MEK/ERK Signaling Pathway in Hepatocellular Carcinoma. Hum. Gene Ther. Clin. Dev. 2019, 30, 129–141. [Google Scholar] [CrossRef]
- Gao, H.; Yang, J.Y.; Tong, L.X.; Jin, H.; Liu, C.Z. Long noncoding RNA UCA1 promotes proliferation and metastasis of thyroid cancer cells by sponging miR-497-3p. Eur. Rev. Med Pharmacol. Sci. 2020, 24, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Jayaganesh, R.; Pugalendhi, P.; Murali, R. Effect of citronellol on NF-kB inflammatory signaling molecules in chemical carcinogen-induced mammary cancer in the rat model. J. Biochem. Mol. Toxicol. 2020, e22441. [Google Scholar] [CrossRef] [PubMed]
- Zha, Z.; Han, Q.; Liu, W.; Huo, S. lncRNA GAS8-AS1 downregulates lncRNA UCA1 to inhibit osteosarcoma cell migration and invasion. J. Orthop. Surg. Res. 2020, 15, 38. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, Z.; Qian, W.; Zhang, Q.; Sun, J. Silence of lncRNA UCA1 rescues drug resistance of cisplatin to non-small-cell lung cancer cells. J. Cell. Biochem. 2019, 120, 9243–9249. [Google Scholar] [CrossRef]
- Puar, Y.R.; Shanmugam, M.K.; Fan, L.; Arfuso, F.; Sethi, G.; Tergaonkar, V. Evidence for the Involvement of the Master Transcription Factor NF-kappaB in Cancer Initiation and Progression. Biomedicines 2018, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Shin, E.M.; Hay, H.S.; Lee, M.H.; Goh, J.N.; Tan, T.Z.; Sen, Y.P.; Lim, S.W.; Yousef, E.M.; Ong, H.T.; Thike, A.A.; et al. DEAD-box helicase DP103 defines metastatic potential of human breast cancers. J. Clin. Investig. 2014, 124, 3807–3824. [Google Scholar] [CrossRef]
- Sethi, G.; Ahn, K.S.; Sung, B.; Aggarwal, B.B. Pinitol targets nuclear factor-κB activation pathway leading to inhibition of gene products associated with proliferation, apoptosis, invasion, and angiogenesis. Mol. Cancer Ther. 2008, 7, 1604–1614. [Google Scholar] [CrossRef] [Green Version]
- Sethi, G.; Sung, B.; Kunnumakkara, A.B.; Aggarwal, B.B. Targeting TNF for Treatment of Cancer and Autoimmunity. Adv. Exp. Med. Biol. 2009, 647, 37–51. [Google Scholar] [CrossRef]
- Ahn, K.S.; Sethi, G.; Jain, A.K.; Jaiswal, A.K.; Aggarwal, B.B. Genetic deletion of NAD(P)H:quinone oxidoreductase 1 abrogates activation of nuclear factor-kappaB, IkappaBalpha kinase, c-Jun N-terminal kinase, Akt, p38, and p44/42 mitogen-activated protein kinases and potentiates apoptosis. J. Biol. Chem. 2006, 281, 19798–19808. [Google Scholar] [CrossRef] [Green Version]
- Siveen, K.S.; Mustafa, N.; Li, F.; Kannaiyan, R.; Ahn, K.S.; Kumar, A.P.; Chng, W.J.; Sethi, G. Thymoquinone overcomes chemoresistance and enhances the anticancer effects of bortezomib through abrogation of NF-kappaB regulated gene products in multiple myeloma xenograft mouse model. Oncotarget 2014, 5, 634–648. [Google Scholar] [CrossRef] [Green Version]
- Nottingham, L.K.; Yan, C.H.; Yang, X.; Si, H.; Coupar, J.; Bian, Y.; Cheng, T.-F.; Allen, C.; Arun, P.; Gius, D. Aberrant IKKα and IKKβ cooperatively activate NF-κB and induce EGFR/AP1 signaling to promote survival and migration of head and neck cancer. Oncogene 2014, 33, 1135–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psyrri, A.; Seiwert, T.Y.; Jimeno, A. Molecular pathways in head and neck cancer: EGFR, PI3K, and more. Am. Soc. Clin. Oncol. Educ. Book 2013, 33, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Shostak, K.; Chariot, A. EGFR and NF-κB: Partners in cancer. Trends Mol. Med. 2015, 21, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Arun, P.; Friedman, J.; Chen, Z.; Van Waes, C. Current and potential inflammation targeted therapies in head and neck cancer. Curr. Opin. Pharmacol. 2009, 9, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Manu, K.A.; Shanmugam, M.K.; Li, F.; Chen, L.; Siveen, K.S.; Ahn, K.S.; Kumar, A.P.; Sethi, G. Simvastatin sensitizes human gastric cancer xenograft in nude mice to capecitabine by suppressing nuclear factor-kappa B-regulated gene products. J. Mol. Med. 2014, 92, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Shanmugam, M.K.; Chen, L.; Chatterjee, S.; Basha, J.; Kumar, A.P.; Kundu, T.K.; Sethi, G. Garcinol, a polyisoprenylated benzophenone modulates multiple proinflammatory signaling cascades leading to the suppression of growth and survival of head and neck carcinoma. Cancer Prev. Res. 2013, 6, 843–854. [Google Scholar] [CrossRef] [Green Version]
- Manu, K.A.; Shanmugam, M.K.; Ramachandran, L.; Li, F.; Siveen, K.S.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Arfuso, F.; Kumar, A.P.; et al. Isorhamnetin augments the anti-tumor effect of capecitabine through the negative regulation of NF-kappaB signaling cascade in gastric cancer. Cancer Lett. 2015, 363, 28–36. [Google Scholar] [CrossRef]
- Liao, J.; Yang, Z.; Carter-Cooper, B.; Chang, E.T.; Choi, E.Y.; Kallakury, B.; Liu, X.; Lapidus, R.G.; Cullen, K.J.; Dan, H. Suppression of migration, invasion, and metastasis of cisplatin-resistant head and neck squamous cell carcinoma through IKKβ inhibition. Clin. Exp. Metastasis 2020, 37, 283–292. [Google Scholar] [CrossRef]
- He, Y.; Xie, H.; Yu, P.; Jiang, S.; Wei, L. FOXC2 promotes epithelial-mesenchymal transition and cisplatin resistance of non-small cell lung cancer cells. Cancer Chemother. Pharmacol. 2018, 82, 1049–1059. [Google Scholar] [CrossRef]
- Soleimani, F.; Hajjari, M.; Mohammad Soltani, B.; Behmanesh, M. Up-Regulation of FOXC2 and FOXQ1 Is Associated with The Progression of Gastric-Type Adenocarcinoma. Cell J. 2017, 19, 66–71. [Google Scholar] [CrossRef]
- Song, L.; Tang, H.; Liao, W.; Luo, X.; Li, Y.; Chen, T.; Zhang, X. FOXC2 positively regulates YAP signaling and promotes the glycolysis of nasopharyngeal carcinoma. Exp. Cell Res. 2017, 357, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Hargadon, K.M.; Gyorffy, B.; Strong, E.W.; Tarnai, B.D.; Thompson, J.C.; Bushhouse, D.Z.; Johnson, C.E.; Williams, C.J. The FOXC2 Transcription Factor Promotes Melanoma Outgrowth and Regulates Expression of Genes Associated With Drug Resistance and Interferon Responsiveness. Cancer Genom. Proteom. 2019, 16, 491–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borretzen, A.; Gravdal, K.; Haukaas, S.A.; Beisland, C.; Akslen, L.A.; Halvorsen, O.J. FOXC2 expression and epithelial-mesenchymal phenotypes are associated with castration resistance, metastasis and survival in prostate cancer. J. Pathol. Clin. Res. 2019, 5, 272–286. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.L.; Lee, W.; Williams, E.D.; Lubik, A.A.; Stylianou, N.; Shokoohmand, A.; Lehman, M.L.; Hollier, B.G.; Gunter, J.H.; Nelson, C.C. Insulin Enhances Migration and Invasion in Prostate Cancer Cells by Up-Regulation of FOXC2. Front. Endocrinol. 2019, 10, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, T.N.D.; Perez White, B.E.; Zhao, H.; Mortazavi, F.; Tonetti, D.A. Protein kinase C alpha enhances migration of breast cancer cells through FOXC2-mediated repression of p120-catenin. BMC Cancer 2017, 17, 832. [Google Scholar] [CrossRef]
- Gao, F.; Wang, X.; Chen, S.; Xu, T.; Wang, X.; Shen, Y.; Dong, F.; Zhong, S.; Shen, Z. CIP2A depletion potentiates the chemosensitivity of cisplatin by inducing increased apoptosis in bladder cancer cells. Oncol. Rep. 2018, 40, 2445–2454. [Google Scholar] [CrossRef]
- Chanvorachote, P.; Chunhacha, P. Caveolin-1 regulates endothelial adhesion of lung cancer cells via reactive oxygen species-dependent mechanism. PLoS ONE 2013, 8, e57466. [Google Scholar] [CrossRef]
- Petpiroon, N.; Bhummaphan, N.; Tungsukruthai, S.; Pinkhien, T.; Maiuthed, A.; Sritularak, B.; Chanvorachote, P. Chrysotobibenzyl inhibition of lung cancer cell migration through Caveolin-1-dependent mediation of the integrin switch and the sensitization of lung cancer cells to cisplatin-mediated apoptosis. Phytomedicine 2019, 58, 152888. [Google Scholar] [CrossRef]
- Milone, M.R.; Lombardi, R.; Roca, M.S.; Bruzzese, F.; Addi, L.; Pucci, B.; Budillon, A. Novel pathways involved in cisplatin resistance identified by a proteomics approach in non-small-cell lung cancer cells. J. Cell. Physiol. 2019, 234, 9077–9092. [Google Scholar] [CrossRef]
- Sato, N.; Kumasawa, K.; Yamashita, M.; Miyake, T.; Nakamura, H.; Kimura, T. Therapeutic potential of combination therapy of soluble VEGF receptor 1 and conventional chemotherapy for ovarian cancer growth. J. Obstet. Gynaecol. Res. 2020. [Google Scholar] [CrossRef]
- Alghzzawy, Z.M.; Elmaghraby, T.K.; El-Hamid Hagag, S.A.; Awwad, M.H. Combretastatin A-4 disodium phosphate and low dose gamma irradiation suppress hepatocellular carcinoma by downregulating ROCK1 and VEGF gene expression. Mol. Biol. Rep. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C. Regulation of PD-L1 expression on cancer cells with ROS-modulating drugs. Life Sci. 2020, 117403. [Google Scholar] [CrossRef] [PubMed]
- Gadducci, A.; Lanfredini, N.; Sergiampietri, C. Antiangiogenic agents in gynecological cancer: State of art and perspectives of clinical research. Crit. Rev. Oncol. Hematol. 2015, 96, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Cereda, V.; Formica, V.; Roselli, M. Issues and promises of bevacizumab in prostate cancer treatment. Expert Opin. Biol. Ther. 2018, 18, 707–717. [Google Scholar] [CrossRef]
- Schmidt, E.V. Developing combination strategies using PD-1 checkpoint inhibitors to treat cancer. Semin. Immunopathol. 2019, 41, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Balar, A.V.; Weber, J.S. PD-1 and PD-L1 antibodies in cancer: Current status and future directions. Cancer Immunol. Immunother. 2017, 66, 551–564. [Google Scholar] [CrossRef]
- Kastenhuber, E.R.; Lowe, S.W. Putting p53 in context. Cell 2017, 170, 1062–1078. [Google Scholar] [CrossRef] [Green Version]
- Wasylishen, A.R.; Lozano, G. Attenuating the p53 pathway in human cancers: Many means to the same end. Cold Spring Harb. Perspect. Med. 2016, 6, a026211. [Google Scholar] [CrossRef]
- Honda, R.; Tanaka, H.; Yasuda, H. Oncoprotein MDM2 is a ubiquitin ligase E3 for tumor suppressor p53. FEBS Lett. 1997, 420, 25–27. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Laramy, J.K.; Gampa, G.; Parrish, K.E.; Brundage, R.; Sarkaria, J.N.; Elmquist, W.F. Brain Distributional Kinetics of a Novel MDM2 Inhibitor SAR405838: Implications for Use in Brain Tumor Therapy. Drug Metab. Dispos. 2019, 47, 1403–1414. [Google Scholar] [CrossRef]
- Ning, Y.; Hui, N.; Qing, B.; Zhuo, Y.; Sun, W.; Du, Y.; Liu, S.; Liu, K.; Zhou, J. ZCCHC10 suppresses lung cancer progression and cisplatin resistance by attenuating MDM2-mediated p53 ubiquitination and degradation. Cell Death Dis. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-S. Medical uses of mylabris in ancient China and recent studies. J. Ethnopharmacol. 1989, 26, 147–162. [Google Scholar] [CrossRef]
- Shi, X.; Chen, S.; Zhang, Y.; Xie, W.; Hu, Z.; Li, H.; Li, J.; Zhou, Z.; Tan, W. Norcantharidin inhibits the DDR of bladder cancer stem-like cells through cdc6 degradation. OncoTargets Ther. 2019, 12, 4403–4413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, D.; Wu, Y.; Shao, C.; Gao, Y.; Wang, D.; Guo, J. Norcantharidin reverses cisplatin resistance and inhibits the epithelial mesenchymal transition of human nonsmall lung cancer cells by regulating the YAP pathway. Oncol. Rep. 2018, 40, 609–620. [Google Scholar] [CrossRef]
- Yeo, C.; Han, D.S.; Lee, H.J.; Lee, E.O. Epigallocatechin-3-Gallate Suppresses Vasculogenic Mimicry through Inhibiting the Twist/VE-Cadherin/AKT Pathway in Human Prostate Cancer PC-3 Cells. Int. J. Mol. Sci. 2020, 21, 439. [Google Scholar] [CrossRef] [Green Version]
- Nagaishi, M.; Fujii, Y.; Sugiura, Y.; Takano, I.; Takigawa, T.; Yokoo, H.; Suzuki, K. Increased Twist and ZEB2 expression in a cutaneous metastasis of high-grade glioma. Neuropathology 2019. [Google Scholar] [CrossRef]
- Wang, L.; Tan, R.Z.; Zhang, Z.X.; Yin, R.; Zhang, Y.L.; Cui, W.J.; He, T. Association between Twist and multidrug resistance gene-associated proteins in Taxol((R))-resistant MCF-7 cells and a 293 cell model of Twist overexpression. Oncol. Lett. 2018, 15, 1058–1066. [Google Scholar] [CrossRef]
- Lai, Y.J.; Yu, W.N.; Kuo, S.C.; Ho, C.T.; Hung, C.M.; Way, T.D.; Chen, C.T. CSC-3436 inhibits TWIST-induced epithelial-mesenchymal transition via the suppression of Twist/Bmi1/Akt pathway in head and neck squamous cell carcinoma. J. Cell. Physiol. 2019, 234, 9118–9129. [Google Scholar] [CrossRef]
- Cheong, C.M.; Mrozik, K.M.; Hewett, D.R.; Bell, E.; Panagopoulos, V.; Noll, J.E.; Licht, J.D.; Gronthos, S.; Zannettino, A.C.W.; Vandyke, K. Twist-1 is upregulated by NSD2 and contributes to tumour dissemination and an epithelial-mesenchymal transition-like gene expression signature in t(4;14)-positive multiple myeloma. Cancer Lett. 2020. [Google Scholar] [CrossRef]
- Cho, Y.A.; Kim, E.K.; Cho, B.C.; Koh, Y.W.; Yoon, S.O. Twist and Snail/Slug Expression in Oropharyngeal Squamous Cell Carcinoma in Correlation With Lymph Node Metastasis. Anticancer Res. 2019, 39, 6307–6316. [Google Scholar] [CrossRef]
- Lin, C.R.; Chu, T.M.; Luo, A.; Huang, S.J.; Chou, H.Y.; Lu, M.W.; Wu, J.L. Omega-3 polyunsaturated fatty acids suppress metastatic features of human cholangiocarcinoma cells by suppressing twist. J. Nutr. Biochem. 2019, 74, 108245. [Google Scholar] [CrossRef] [PubMed]
- Shahin, S.A.; Wang, R.; Simargi, S.I.; Contreras, A.; Echavarria, L.P.; Qu, L.; Wen, W.; Dellinger, T.; Unternaehrer, J.; Tamanoi, F. Hyaluronic acid conjugated nanoparticle delivery of siRNA against TWIST reduces tumor burden and enhances sensitivity to cisplatin in ovarian cancer. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 1381–1394. [Google Scholar] [CrossRef] [PubMed]
- Manirujjaman, M.; Ozaki, I.; Murata, Y.; Guo, J.; Xia, J.; Nishioka, K.; Perveen, R.; Takahashi, H.; Anzai, K.; Matsuhashi, S. Degradation of the Tumor Suppressor PDCD4 Is Impaired by the Suppression of p62/SQSTM1 and Autophagy. Cells 2020, 9, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De, P.; Carlson, J.; Leyland-Jones, B.; Dey, N. Oncogenic nexus of cancerous inhibitor of protein phosphatase 2A (CIP2A): An oncoprotein with many hands. Oncotarget 2014, 5, 4581. [Google Scholar] [CrossRef] [Green Version]
- Junttila, M.R.; Puustinen, P.; Niemelä, M.; Ahola, R.; Arnold, H.; Böttzauw, T.; Ala-aho, R.; Nielsen, C.; Ivaska, J.; Taya, Y. CIP2A inhibits PP2A in human malignancies. Cell 2007, 130, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Gu, T.-T.; Zheng, P.-S. CIP2A cooperates with H-Ras to promote epithelial–mesenchymal transition in cervical-cancer progression. Cancer Lett. 2015, 356, 646–655. [Google Scholar] [CrossRef]
- Xu, P.; Yao, J.; He, J.; Zhao, L.; Wang, X.; Li, Z.; Qian, J. CIP2A down regulation enhances the sensitivity of pancreatic cancer cells to gemcitabine. Oncotarget 2016, 7, 14831. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.-Z.; Zhou, X.-D.; Qian, G.; Shi, X.; Fang, J.; Jiang, B.-H. AKT1 amplification regulates cisplatin resistance in human lung cancer cells through the mammalian target of rapamycin/p70S6K1 pathway. Cancer Res. 2007, 67, 6325–6332. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Gong, G.Y.; Ma, L.L.; Wang, Z.Q.; Song, D.; Fang, M.Y. Anti-cancer effects of Polyphyllin I: An update in 5 years. Chem. Biol. Interact. 2019, 316, 108936. [Google Scholar] [CrossRef]
- Cui, J.; Man, S.; Cui, N.; Yang, L.; Guo, Q.; Ma, L.; Gao, W. The synergistic anticancer effect of formosanin C and polyphyllin VII based on caspase-mediated cleavage of Beclin1 inhibiting autophagy and promoting apoptosis. Cell Prolif. 2019, 52, e12520. [Google Scholar] [CrossRef] [Green Version]
- Feng, F.; Cheng, P.; Wang, C.; Wang, Y.; Wang, W. Polyphyllin I and VII potentiate the chemosensitivity of A549/DDP cells to cisplatin by enhancing apoptosis, reversing EMT and suppressing the CIP2A/AKT/mTOR signaling axis. Oncol. Lett. 2019, 18, 5428–5436. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhai, R.; Zhao, J.; Kong, F.; Wang, J.; Jiang, W.; Xin, Q.; Xue, X.; Luan, Y. Programmed cell death 4 overexpression enhances sensitivity to cisplatin via the JNK/c-Jun signaling pathway in bladder cancer. Int. J. Oncol. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Chen, L.; Ge, J.; Yan, C.; Huang, Z.; Hu, J.; Wen, C.; Li, M.; Huang, D.; Qiu, Y. The ubiquitin-like protein FAT10 stabilizes eEF1A1 expression to promote tumor proliferation in a complex manner. Cancer Res. 2016, 76, 4897–4907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, D.-M.; Bian, X.-Y.; Qin, C.-D.; Wu, W.-Z. miR-106b-5p promotes stem cell-like properties of hepatocellular carcinoma cells by targeting PTEN via PI3K/Akt pathway. OncoTargets Ther. 2018, 11, 571. [Google Scholar] [CrossRef] [Green Version]
- Talesa, V.N.; Ferri, I.; Bellezza, G.; Love, H.D.; Sidoni, A.; Antognelli, C. Glyoxalase 2 is involved in human prostate cancer progression as part of a mechanism driven by PTEN/PI3K/AKT/mTOR signaling with involvement of PKM2 and ERα. Prostate 2017, 77, 196–210. [Google Scholar] [CrossRef]
- Zhao, X.; Dai, L.; Yue, Q.; Wang, H.; Wang, X.U.; Li, Y.; Chen, R. MiR-195 inhibits migration, invasion and epithelial-mesenchymal transition (EMT) of endometrial carcinoma cells by targeting SOX4. J. Biosci. 2019, 44, 146. [Google Scholar] [CrossRef]
- Liu, L.; Jung, S.N.; Oh, C.; Lee, K.; Won, H.R.; Chang, J.W.; Kim, J.M.; Koo, B.S. LAMB3 is associated with disease progression and cisplatin cytotoxic sensitivity in head and neck squamous cell carcinoma. Eur. J. Surg. Oncol. 2019, 45, 359–365. [Google Scholar] [CrossRef]
- Zhang, P.; Lu, X.; Shi, Z.; Li, X.; Zhang, Y.; Zhao, S.; Liu, H. miR-205-5p regulates epithelial-mesenchymal transition by targeting PTEN via PI3K/AKT signaling pathway in cisplatin-resistant nasopharyngeal carcinoma cells. Gene 2019, 710, 103–113. [Google Scholar] [CrossRef]
- Jung, S.N.; Lim, H.S.; Liu, L.; Chang, J.W.; Lim, Y.C.; Rha, K.S.; Koo, B.S. LAMB3 mediates metastatic tumor behavior in papillary thyroid cancer by regulating c-MET/Akt signals. Sci. Rep. 2018, 8, 2718. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jin, Y.; Bhandari, A.; Yao, Z.; Yang, F.; Pan, Y.; Zheng, Z.; Lv, S.; Wang, O. Upregulated LAMB3 increases proliferation and metastasis in thyroid cancer. OncoTargets Ther. 2018, 11, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Li, Y.; Fan, L.; Zhao, Q.; Tan, B.; Cui, G. Upregulation Of miR-153 Inhibits Triple-Negative Breast Cancer Progression By Targeting ZEB2-Mediated EMT And Contributes To Better Prognosis. OncoTargets Ther. 2019, 12, 9611–9625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, A.; Rahaman, A.; Biswas, I.; Mukherjee, G.; Chatterjee, S.; Bhattacharjee, S.; Mandal, D.P. TGFbeta mediated LINC00273 upregulation sponges mir200a-3p and promotes invasion and metastasis by activating ZEB1. J. Cell. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, X.; Yang, L.; Dong, X.; Zheng, Z.; Cheng, Y.; Chen, H.; Lan, B.; Li, D.; Zhou, J. Disruptor of telomeric silencing 1-like (DOT1L) is involved in breast cancer metastasis via transcriptional regulation of MALAT1 and ZEB2. J. Genet. Genom. Yi Chuan Xue Bao 2019, 46, 591–594. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Sun, S.; Zhou, X.; Zhang, Q.; Guo, W.; Zhang, L. Clinicopathological significance of ZEB-1 and E-cadherin proteins in patients with oral cavity squamous cell carcinoma. OncoTargets Ther. 2017, 10, 781. [Google Scholar] [CrossRef] [Green Version]
- Cleophas, M.; Joosten, L.; Stamp, L.K.; Dalbeth, N.; Woodward, O.M.; Merriman, T.R. ABCG2 polymorphisms in gout: Insights into disease susceptibility and treatment approaches. Pharm. Pers. Med. 2017, 10, 129. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, G.; Rath, B. Pharmacogenetics of platinum-based chemotherapy in non-small cell lung cancer: Predictive validity of polymorphisms of ERCC1. Expert Opin. Drug Metab. Toxicol. 2018, 14, 17–24. [Google Scholar] [CrossRef]
- Wu, Y.; Jin, D.; Wang, X.; Du, J.; Di, W.; An, J.; Shao, C.; Guo, J. UBE2C induces cisplatin resistance via ZEB1/2-dependent upregulation of ABCG2 and ERCC1 in NSCLC cells. J. Oncol. 2019, 2019, 8607859. [Google Scholar] [CrossRef] [Green Version]
- Guo, T.; Yuan, X.; Liu, D.F.; Peng, S.H.; Xu, A.M. LncRNA HOXA11-AS promotes migration and invasion through modulating miR-148a/WNT1/beta-catenin pathway in gastric cancer. Neoplasma 2020. [Google Scholar] [CrossRef]
- Bai, Y.; Lang, L.; Zhao, W.; Niu, R. Long Non-Coding RNA HOXA11-AS Promotes Non-Small Cell Lung Cancer Tumorigenesis Through microRNA-148a-3p/DNMT1 Regulatory Axis. OncoTargets Ther. 2019, 12, 11195–11206. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, Q.; Chen, H.; Wang, C. BCL9 promotes epithelial mesenchymal transition and invasion in cisplatin resistant NSCLC cells via β-catenin pathway. Life Sci. 2018, 208, 284–294. [Google Scholar] [CrossRef]
- Chen, N.; Kong, Y.; Wu, Y.; Gao, Q.; Fu, J.; Sun, X.; Geng, Q. CAC1 knockdown reverses drug resistance through the downregulation of P-gp and MRP-1 expression in colorectal cancer. PLoS ONE 2019, 14, e0222035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.Y.; Duan, G.L.; Xu, R.Y.; Li, X.R.; Xiao, L.; Zhao, L.; Ma, Z.X.; Xu, X.W.; Qiu, L.J.; Zhu, Z.M.; et al. DJ-1 overexpression confers the multidrug resistance phenotype to SGC7901cells by upregulating P-gp and Bcl-2. Biochem. Biophys. Res. Commun. 2019, 519, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Sachs, J.; Dohl, K.; Weber, A.; Bonus, M.; Ehlers, F.; Fleischer, E.; Klinger, A.; Gohlke, H.; Pietruszka, J.; Schmitt, L.; et al. Novel 3,4-Dihydroisocoumarins Inhibit Human P-gp and BCRP in Multidrug Resistant Tumors and Demonstrate Substrate Inhibition of Yeast Pdr5. Front. Pharmacol. 2019, 10, 400. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.C.; Honrado, M.; Armada, A.; Viveiros, M.; Rueff, J.; Rodrigues, A.S. ABC Efflux Transporters and the Circuitry of miRNAs: Kinetics of Expression in Cancer Drug Resistance. Int. J. Mol. Sci. 2020, 21, 2985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leopoldo, M.; Nardulli, P.; Contino, M.; Leonetti, F.; Luurtsema, G.; Colabufo, N.A. An updated patent review on P-glycoprotein inhibitors (2011–2018). Expert Opin. Ther. Pat. 2019, 29, 455–461. [Google Scholar] [CrossRef]
- Ayrton, A.; Morgan, P. Role of transport proteins in drug absorption, distribution and excretion. Xenobiotica 2001, 31, 469–497. [Google Scholar] [CrossRef]
- Dean, M.; Hamon, Y.; Chimini, G. The human ATP-binding cassette (ABC) transporter superfamily. J. Lipid Res. 2001, 42, 1007–1017. [Google Scholar] [CrossRef]
- Giacomini, K.M.; Huang, S.-M.; Tweedie, D.J.; Benet, L.Z.; Brouwer, K.L.; Chu, X.; Dahlin, A.; Evers, R.; Fischer, V.; Hillgren, K.M. Membrane transporters in drug development. Nat. Rev. Drug Discov. 2010, 9, 215. [Google Scholar]
- Lin, H.; Hu, B.; He, X.; Mao, J.; Wang, Y.; Wang, J.; Zhang, T.; Zheng, J.; Peng, Y.; Zhang, F. Overcoming Taxol-resistance in A549 cells: A comprehensive strategy of targeting P-gp transporter, AKT/ERK pathways, and cytochrome P450 enzyme CYP1B1 by 4-hydroxyemodin. Biochem. Pharmacol. 2020, 171, 113733. [Google Scholar] [CrossRef]
- Zhang, E.; Liu, J.; Shi, L.; Guo, X.; Liang, Z.; Zuo, J.; Xu, H.; Wang, H.; Shu, X.; Huang, S.; et al. 7-O-geranylquercetin contributes to reverse P-gp-mediated adriamycin resistance in breast cancer. Life Sci. 2019, 238, 116938. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Y.; Xiang, F.; Li, M.; Li, H.; Chi, J.; Ren, K. Suppression of TGF-beta1 enhances chemosensitivity of cisplatin-resistant lung cancer cells through the inhibition of drug-resistant proteins. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1505–1512. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Kim, Y.-K.; Yun, P.-Y. Upregulation of MDR-and EMT-Related Molecules in Cisplatin-Resistant Human Oral Squamous Cell Carcinoma Cell Lines. Int. J. Mol. Sci. 2019, 20, 3034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enerbäck, C.; Porter, D.A.; Seth, P.; Sgroi, D.; Gaudet, J.; Weremowicz, S.; Morton, C.C.; Schnitt, S.; Pitts, R.L.; Stampl, J. Psoriasin expression in mammary epithelial cells in vitro and in vivo. Cancer Res. 2002, 62, 43–47. [Google Scholar] [PubMed]
- Jinquan, T.; Vorum, H.; Larsen, C.G.; Madsen, P.; Rasmussen, H.H.; Gesser, B.; Etzerodt, M.; Honoré, B.; Celis, J.E.; Thestrup-Pedersen, K. Psoriasin: A novel chemotactic protein. J. Investig. Dermatol. 1996, 107, 5–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, P.; Rasmussen, H.H.; Leffers, H.; Honoré, B.; Dejgaard, K.; Olsen, E.; Kiil, J.; Walbum, E.; Andersen, A.H.; Basse, B. Molecular cloning, occurrence, and expression of a novel partially secreted protein “psoriasin” that is highly up-regulated in psoriatic skin. J. Investig. Dermatol. 1991, 97, 701–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, P.H.; Leygue, E.R.; Murphy, L.C. Psoriasin (S100A7). Int. J. Biochem. Cell Biol. 1998, 30, 567–571. [Google Scholar] [CrossRef]
- Liu, Y.; Bunston, C.; Hodson, N.; Resaul, J.; Sun, P.-H.; Cai, S.; Chen, G.; Gu, Y.; Satherley, L.K.; Bosanquet, D.C. Psoriasin promotes invasion, aggregation and survival of pancreatic cancer cells; association with disease progression. Int. J. Oncol. 2017, 50, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Yu, Z.; Ma, B. The increase of miR-27a affects the role of cisplatin on proliferation and migration capacities of liver cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5490–5498. [Google Scholar]
- Ren, J.; Kan, A.; Leong, S.H.; Ooi, L.L.; Jeang, K.-T.; Chong, S.S.; Kon, O.L.; Lee, C.G. FAT10 plays a role in the regulation of chromosomal stability. J. Biol. Chem. 2006, 281, 11413–11421. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.-Y.; Sham, J.S.; Tang, T.C.; Fang, Y.; Huo, K.-K.; Yang, J.-M. Isolation of a novel candidate oncogene within a frequently amplified region at 3q26 in ovarian cancer. Cancer Res. 2001, 61, 3806–3809. [Google Scholar]
- Guan, X.-Y.; Fung, J.M.; Ma, N.-F.; Lau, S.-H.; Tai, L.-S.; Xie, D.; Zhang, Y.; Hu, L.; Wu, Q.-L.; Fang, Y. Oncogenic role of eIF-5A2 in the development of ovarian cancer. Cancer Res. 2004, 64, 4197–4200. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.-F.; Xie, D.; Liu, J.-H.; Luo, J.-H.; Li, L.-J.; Hua, W.-F.; Wu, H.-M.; Kung, H.-F.; Zeng, Y.-X.; Guan, X.-Y. Expression and amplification of eIF-5A2 in human epithelial ovarian tumors and overexpression of EIF-5A2 is a new independent predictor of outcome in patients with ovarian carcinoma. Gynecol. Oncol. 2009, 112, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.J.; Dong, S.S.; Ma, N.F.; Xie, D.; Chen, L.; Fu, L.; Lau, S.H.; Li, Y.; Li, Y.; Guan, X.Y. Overexpression of eukaryotic initiation factor 5A2 enhances cell motility and promotes tumor metastasis in hepatocellular carcinoma. Hepatology 2010, 51, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Ma, N.-F.; Pan, Z.-Z.; Wu, H.-X.; Liu, Y.-D.; Wu, G.-Q.; Kung, H.-F.; Guan, X.-Y. Overexpression of EIF-5A2 is associated with metastasis of human colorectal carcinoma. Hum. Pathol. 2008, 39, 80–86. [Google Scholar] [CrossRef]
- Huang, P.-Y.; Zeng, T.-T.; Ban, X.; Li, M.-Q.; Zhang, B.-Z.; Zhu, Y.-H.; Hua, W.-F.; Mai, H.-Q.; Zhang, L.; Guan, X.-Y. Expression of EIF5A2 associates with poor survival of nasopharyngeal carcinoma patients treated with induction chemotherapy. BMC Cancer 2016, 16, 669. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Xu, Z.; Lv, H.; Wang, Y.; Wang, L.; Ni, Y.; Wang, X.; Hu, C.; Chen, S.; Teng, F.; et al. eIF5A2 regulates the resistance of gastric cancer cells to cisplatin via induction of EMT. Am. J. Transl. Res. 2018, 10, 4269–4279. [Google Scholar]
- Amerik, A.Y.; Hochstrasser, M. Mechanism and function of deubiquitinating enzymes. Biochim. Biophys. Acta BBA Mol. Cell Res. 2004, 1695, 189–207. [Google Scholar] [CrossRef] [Green Version]
- Alhosin, M.; Omran, Z.; Zamzami, M.A.; Al-Malki, A.L.; Choudhry, H.; Mousli, M.; Bronner, C. Signalling pathways in UHRF1-dependent regulation of tumor suppressor genes in cancer. J. Exp. Clin. Cancer Res. 2016, 35, 174. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Zhou, Z.; Wu, G.; Chen, Q.; Wan, Y. Emerging role of DUBs in tumor metastasis and apoptosis: Therapeutic implication. Pharmacol. Ther. 2017, 177, 96–107. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, X.; Chen, S.; Qin, X. Predictive value of deubiquitination enzymes USP37 in the prognosis of breast cancer. Zhonghua Yi Xue Za Zhi 2016, 96, 944–948. [Google Scholar]
- Tanno, H.; Shigematsu, T.; Nishikawa, S.; Hayakawa, A.; Denda, K.; Tanaka, T.; Komada, M. Ubiquitin-interacting motifs confer full catalytic activity, but not ubiquitin chain substrate specificity, to deubiquitinating enzyme USP37. J. Biol. Chem. 2014, 289, 2415–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Summers, M.K.; Pham, V.; Lill, J.R.; Liu, J.; Lee, G.; Kirkpatrick, D.S.; Jackson, P.K.; Fang, G.; Dixit, V.M. Deubiquitinase USP37 is activated by CDK2 to antagonize APCCDH1 and promote S phase entry. Mol. Cell 2011, 42, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Burrows, A.C.; Prokop, J.; Summers, M.K. Skp1-Cul1-F-box ubiquitin ligase (SCFβTrCP)-mediated destruction of the ubiquitin-specific protease USP37 during G2-phase promotes mitotic entry. J. Biol. Chem. 2012, 287, 39021–39029. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Li, M.; Wang, X.; Li, L.; Li, Q.; Hou, Z.; Jia, H.; Liu, S. USP37 promotes lung cancer cell migration by stabilizing Snail protein via deubiquitination. Front. Genet. 2020, 10, 1324. [Google Scholar] [CrossRef] [PubMed]
- Kasper, M.; Jaks, V.; Fiaschi, M.; Toftgård, R. Hedgehog signalling in breast cancer. Carcinogenesis 2009, 30, 903–911. [Google Scholar] [CrossRef] [Green Version]
- Qin, T.; Li, B.; Feng, X.; Fan, S.; Liu, L.; Liu, D.; Mao, J.; Lu, Y.; Yang, J.; Yu, X. Abnormally elevated USP37 expression in breast cancer stem cells regulates stemness, epithelial-mesenchymal transition and cisplatin sensitivity. J. Exp. Clin. Cancer Res. 2018, 37, 287. [Google Scholar] [CrossRef] [Green Version]
- Fischer, K.R.; Durrans, A.; Lee, S.; Sheng, J.; Li, F.; Wong, S.T.; Choi, H.; El Rayes, T.; Ryu, S.; Troeger, J. Epithelial-to-mesenchymal transition is not required for lung metastasis but contributes to chemoresistance. Nature 2015, 527, 472–476. [Google Scholar] [CrossRef]
- Yu, Y.-C.; Yang, P.-M.; Chuah, Q.-Y.; Huang, Y.-H.; Peng, C.-W.; Lee, Y.-J.; Chiu, S.-J. Radiation-induced senescence in securin-deficient cancer cells promotes cell invasion involving the IL-6/STAT3 and PDGF-BB/PDGFR pathways. Sci. Rep. 2013, 3, 1675. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.L.A.; Hirpara, J.L.; Pervaiz, S.; Eu, J.Q.; Sethi, G.; Goh, B.C. Do STAT3 inhibitors have potential in the future for cancer therapy? Expert Opin. Investig. Drugs 2017, 26, 883–887. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.M.; Li, F.; Rajendran, P.; Kumar, A.P.; Hui, K.M.; Sethi, G. Identification of beta-escin as a novel inhibitor of signal transducer and activator of transcription 3/Janus-activated kinase 2 signaling pathway that suppresses proliferation and induces apoptosis in human hepatocellular carcinoma cells. J. Pharmacol. Exp. Ther. 2010, 334, 285–293. [Google Scholar] [CrossRef]
- Mohan, C.D.; Bharathkumar, H.; Bulusu, K.C.; Pandey, V.; Rangappa, S.; Fuchs, J.E.; Shanmugam, M.K.; Dai, X.; Li, F.; Deivasigamani, A.; et al. Development of a novel azaspirane that targets the Janus kinase-signal transducer and activator of transcription (STAT) pathway in hepatocellular carcinoma in vitro and in vivo. J. Biol. Chem. 2014, 289, 34296–34307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajendran, P.; Li, F.; Shanmugam, M.K.; Vali, S.; Abbasi, T.; Kapoor, S.; Ahn, K.S.; Kumar, A.P.; Sethi, G. Honokiol inhibits signal transducer and activator of transcription-3 signaling, proliferation, and survival of hepatocellular carcinoma cells via the protein tyrosine phosphatase SHP-1. J. Cell. Physiol. 2012, 227, 2184–2195. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Koh, J.; Kim, M.-Y.; Kwon, D.; Go, H.; Kim, Y.A.; Jeon, Y.K.; Chung, D.H. PD-L1 expression is associated with epithelial-to-mesenchymal transition in adenocarcinoma of the lung. Hum. Pathol. 2016, 58, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, H.; Zhao, Q.; Xia, Y.; Hu, X.; Guo, J. PD-L1 induces epithelial-to-mesenchymal transition via activating SREBP-1c in renal cell carcinoma. Med. Oncol. 2015, 32, 212. [Google Scholar] [CrossRef]
- Shen, M.; Xu, Z.; Xu, W.; Jiang, K.; Zhang, F.; Ding, Q.; Xu, Z.; Chen, Y. Inhibition of ATM reverses EMT and decreases metastatic potential of cisplatin-resistant lung cancer cells through JAK/STAT3/PD-L1 pathway. J. Exp. Clin. Cancer Res. 2019, 38, 149. [Google Scholar] [CrossRef] [Green Version]
- Romacho, T.; Sánchez-Ferrer, C.F.; Peiró, C. Visfatin/Nampt: An adipokine with cardiovascular impact. Mediat. Inflamm. 2013, 2013, 946427. [Google Scholar] [CrossRef]
- Carbone, F.; Liberale, L.; Bonaventura, A.; Vecchiè, A.; Casula, M.; Cea, M.; Monacelli, F.; Caffa, I.; Bruzzone, S.; Montecucco, F. Regulation and function of extracellular nicotinamide phosphoribosyltransferase/visfatin. Compr. Physiol. 2011, 7, 603–621. [Google Scholar]
- Mohammadi, M.; Moradi, A.; Farhadi, J.; Akbari, A.; Pourmandi, S.; Mehrad-Majd, H. Prognostic value of visfatin in various human malignancies: A systematic review and meta-analysis. Cytokine 2020, 127, 154964. [Google Scholar] [CrossRef]
- Mohammadi, M.; Mianabadi, F.; Mehrad-Majd, H. Circulating visfatin levels and cancers risk: A systematic review and meta-analysis. J. Cell. Physiol. 2019, 234, 5011–5022. [Google Scholar] [CrossRef]
- Wang, D.; Qian, G.; Wang, J.; Wang, T.; Zhang, L.; Yang, P.; Lin, F. Visfatin is involved in the cisplatin resistance of osteosarcoma cells via upregulation of Snail and Zeb1. Cancer Boil. Ther. 2019, 20, 999–1006. [Google Scholar] [CrossRef]
- Wang, J.; Han, Y.; Wang, M.; Zhao, Q.; Chen, X.; Liu, X. Natural triterpenoid saponin Momordin Ic suppresses HepG2 cell invasion via COX-2 inhibition and PPARgamma activation. Toxicol. Vitro 2020, 65, 104784. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Hu, S.; Liu, C.F.; Jiang, L.; Yu, W.; Li, Z.L.; Guo, W.; Tang, R.; Dong, C.Y.; Wu, T.H.; et al. BPTF cooperates with p50 NF-kappaB to promote COX-2 expression and tumor cell growth in lung cancer. Am. J. Transl. Res. 2019, 11, 7398–7409. [Google Scholar] [PubMed]
- Jiang, G.B.; Fang, H.Y.; Tao, D.Y.; Chen, X.P.; Cao, F.L. COX-2 potentiates cisplatin resistance of non-small cell lung cancer cells by promoting EMT in an AKT signaling pathway-dependent manner. Eur. Rev. Med Pharmacol. Sci. 2019, 23, 3838–3846. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.S.; Nemeria, N.S.; Furey, W.; Jordan, F. The pyruvate dehydrogenase complexes: Structure-based function and regulation. J. Boil. Chem. 2014, 289, 16615–16623. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Su, J.; Xu, H.; Yu, S.; Liu, Y.; Zhang, Y.; Sun, L.; Yue, Y.; Zhou, X. Dicumarol inhibits PDK1 and targets multiple malignant behaviors of ovarian cancer cells. PLoS ONE 2017, 12, e0179672. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Yin, H. PDK1 promotes tumor cell proliferation and migration by enhancing the Warburg effect in non-small cell lung cancer. Oncol. Rep. 2017, 37, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.-L.; Hu, X.; Zhang, W.; Yao, H.; Tam, K.Y. Development of pyruvate dehydrogenase kinase inhibitors in medicinal chemistry with particular emphasis as anticancer agents. Drug Discov. Today 2015, 20, 1112–1119. [Google Scholar] [CrossRef]
- Sugiyama, E.; Togashi, Y.; Takeuchi, Y.; Shinya, S.; Tada, Y.; Kataoka, K.; Tane, K.; Sato, E.; Ishii, G.; Goto, K.; et al. Blockade of EGFR improves responsiveness to PD-1 blockade in EGFR-mutated non-small cell lung cancer. Sci. Immunol. 2020, 5. [Google Scholar] [CrossRef]
- Huang, L.; Jiang, X.; Kang, P.; Wang, Z.; Leng, K.; Ji, D.; Xu, Y.; Wang, H.; Cui, Y. Long non-coding RNA NNT-AS1 functions as an oncogenic gene through modulating miR-485/BCL9 in cholangiocarcinoma. Cancer Manag. Res. 2019, 11, 7739–7749. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Shen, T.; Zhang, C.; Zhang, Q.H.; Zhang, Z.Q. MiR-125a-5p inhibits EMT of ovarian cancer cells by regulating TAZ/EGFR signaling pathway. Eur. Rev. Med Pharmacol. Sci. 2019, 23, 8249–8256. [Google Scholar] [CrossRef]
- Xing, Y.; Jing, H.; Zhang, Y.; Suo, J.; Qian, M. MicroRNA-141-3p affected proliferation, chemosensitivity, migration and invasion of colorectal cancer cells by targeting EGFR. Int. J. Biochem. Cell Boil. 2020, 118, 105643. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.-J.; Hour, M.-J.; Jin, Y.-A.; Lu, C.-C.; Tsai, F.-J.; Chen, T.-L.; Ma, H.; Juan, Y.-N.; Yang, J.-S. Disruption of IGF-1R signaling by a novel quinazoline derivative, HMJ-30, inhibits invasiveness and reverses epithelial-mesenchymal transition in osteosarcoma U-2 OS cells. Int. J. Oncol. 2018, 52, 1465–1478. [Google Scholar] [CrossRef] [PubMed]
- Ernst, B.P.; Mikstas, C.; Stoever, T.; Stauber, R.; Strieth, S. Association of eIF4E and SPARC expression with lymphangiogenesis and lymph node metastasis in hypopharyngeal cancer. Anticancer Res. 2018, 38, 699–706. [Google Scholar] [PubMed]
- Liang, S.; Guo, R.; Zhang, Z.; Liu, D.; Xu, H.; Xu, Z.; Wang, X.; Yang, L. Upregulation of the eIF4E signaling pathway contributes to the progression of gastric cancer, and targeting eIF4E by perifosine inhibits cell growth. Oncol. Rep. 2013, 29, 2422–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oblinger, J.L.; Burns, S.S.; Huang, J.; Pan, L.; Ren, Y.; Shen, R.; Kinghorn, A.D.; Welling, D.B.; Chang, L.-S. Overexpression of eIF4F components in meningiomas and suppression of meningioma cell growth by inhibiting translation initiation. Exp. Neurol. 2018, 299, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Liu, Y.; Di, X.; Kang, H.; Zeng, H.; Zhao, Y.; Cai, K.; Pang, T.; Wang, S.; Yao, Y. EIF4E over-expresses and enhances cell proliferation and cell cycle progression in nasopharyngeal carcinoma. Med. Oncol. 2013, 30, 400. [Google Scholar] [CrossRef]
- Zhao, Y.; Pang, T.Y.; Wang, Y.; Wang, S.; Kang, H.X.; Ding, W.B.; Yong, W.W.; Bie, Y.H.; Cheng, X.G.; Zeng, C. LMP 1 stimulates the transcription of e IF 4E to promote the proliferation, migration and invasion of human nasopharyngeal carcinoma. FEBS J. 2014, 281, 3004–3018. [Google Scholar] [CrossRef]
- Yao, Y.; Pang, T.; Cheng, Y.; Yong, W.; Kang, H.; Zhao, Y.; Wang, S.; Hu, X. Positive Correlative over-Expression between eIF4E and Snail in Nasopharyngeal Carcinoma Promotes its Metastasis and Resistance to Cisplatin. Pathol. Oncol. Res. 2019, 1–11. [Google Scholar] [CrossRef]
- Giaimo, B.D.; Ferrante, F.; Herchenröther, A.; Hake, S.B.; Borggrefe, T. The histone variant H2A.Z in gene regulation. Epigenet. Chromatin 2019, 12, 37. [Google Scholar] [CrossRef] [Green Version]
- Rispal, J.; Baron, L.; Beaulieu, J.F.; Chevillard-Briet, M.; Trouche, D.; Escaffit, F. The H2A.Z histone variant integrates Wnt signaling in intestinal epithelial homeostasis. Nat. Commun. 2019, 10, 1827. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, M.; Cheema, M.S.; Dryhurst, D.; Eskiw, C.H.; Ausio, J. Metformin alters H2A.Z dynamics and regulates androgen dependent prostate cancer progression. Oncotarget 2018, 9, 37054–37068. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Kayukawa, N.; Ueda, T.; Taniguchi, H.; Morioka, Y.; Hongo, F.; Ukimura, O. MRGBP promotes AR-mediated transactivation of KLK3 and TMPRSS2 via acetylation of histone H2A.Z in prostate cancer cells. Biochim. Biophys. Acta Gene Regul. Mech. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Shi, J.; Yuan, C.; Zhao, D.; Jiang, S.; Lyu, J.; Wang, X.; Li, H.; Wen, H.; Li, W.; et al. Recognition of histone acetylation by the GAS41 YEATS domain promotes H2A.Z deposition in non-small cell lung cancer. Genes Dev. 2018, 32, 58–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Tong, R.; Liu, H.; Wu, J.; Chen, D.; Xue, Z.; Ding, C.; Zhou, L.; Xie, H.; Wu, J. H2A. Z regulates tumorigenesis, metastasis and sensitivity to cisplatin in intrahepatic cholangiocarcinoma. Int. J. Oncol. 2018, 52, 1235–1245. [Google Scholar] [PubMed]
- Wangpaichitr, M.; Kandemir, H.; Li, Y.Y.; Wu, C.; Nguyen, D.; Feun, L.G.; Kuo, M.T.; Savaraj, N. Relationship of Metabolic Alterations and PD-L1 Expression in Cisplatin Resistant Lung Cancer. Cell Dev. Biol. 2017, 6. [Google Scholar] [CrossRef]
- Xie, S.L.; Fan, S.; Zhang, S.Y.; Chen, W.X.; Li, Q.X.; Pan, G.K.; Zhang, H.Q.; Wang, W.W.; Weng, B.; Zhang, Z.; et al. SOX8 regulates cancer stem-like properties and cisplatin-induced EMT in tongue squamous cell carcinoma by acting on the Wnt/beta-catenin pathway. Int. J. Cancer 2018, 142, 1252–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Tian, L.; Khan, M.N.; Zhang, L.; Chen, Q.; Zhao, Y.; Yan, Q.; Fu, L.; Liu, J. Ginsenoside Rg3 sensitizes hypoxic lung cancer cells to cisplatin via blocking of NF-kappaB mediated epithelial-mesenchymal transition and stemness. Cancer Lett. 2018, 415, 73–85. [Google Scholar] [CrossRef]
- Hou, L.; Hou, X.; Wang, L.; Li, Z.; Xin, B.; Chen, J.; Gao, X.; Mu, H. PD98059 impairs the cisplatin-resistance of ovarian cancer cells by suppressing ERK pathway and epithelial mesenchymal transition process. Cancer Biomark. Sect. A Dis. Markers 2017, 21, 187–194. [Google Scholar] [CrossRef]
- Islam, S.S.; Al-Sharif, I.; Sultan, A.; Al-Mazrou, A.; Remmal, A.; Aboussekhra, A. Eugenol potentiates cisplatin anti-cancer activity through inhibition of ALDH-positive breast cancer stem cells and the NF-kappaB signaling pathway. Mol. Carcinog. 2018, 57, 333–346. [Google Scholar] [CrossRef]
- Mei, Y.; Peng, C.; Liu, Y.B.; Wang, J.; Zhou, H.H. Silencing RIF1 decreases cell growth, migration and increases cisplatin sensitivity of human cervical cancer cells. Oncotarget 2017, 8, 107044–107051. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Yao, J.F.; Deng, X.F.; Zheng, X.D.; Jia, M.; Wang, Y.Q.; Huang, Y.; Zhu, J.H. 14, 15-EET induces breast cancer cell EMT and cisplatin resistance by up-regulating integrin alphavbeta3 and activating FAK/PI3K/AKT signaling. J. Exp. Clin. Cancer Res. 2018, 37, 23. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Miao, C.; Jin, C.; Qiu, C.; Li, Y.; Sun, X.; Gao, M.; Lu, N.; Kong, B. SUSD2 promotes cancer metastasis and confers cisplatin resistance in high grade serous ovarian cancer. Exp. Cell Res. 2018, 363, 160–170. [Google Scholar] [CrossRef]
- Kim, S.K.; Park, J.A.; Zhang, D.; Cho, S.H.; Yi, H.; Cho, S.M.; Chang, B.J.; Kim, J.S.; Shim, J.H.; Abd El-Aty, A.M.; et al. Sustainability of CD24 expression, cell proliferation and migration, cisplatin-resistance, and caspase-3 expression during mesenchymal-epithelial transition induced by the removal of TGF-beta1 in A549 lung cancer cells. Oncol. Lett. 2017, 14, 2410–2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Han, S.; Li, Y.; Liu, Y.; Zhang, D.; Li, Y.; Zhang, J. MicroRNA-20a contributes to cisplatin-resistance and migration of OVCAR3 ovarian cancer cell line. Oncol. Lett. 2017, 14, 1780–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.M.; Liu, T.; Deng, S.H.; Han, R.; Xu, Y. SLC39A4 expression is associated with enhanced cell migration, cisplatin resistance, and poor survival in non-small cell lung cancer. Sci. Rep. 2017, 7, 7211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhang, X.; Zhang, R.; Liang, Z.; Liao, W.; Du, Z.; Gao, C.; Liu, F.; Fan, Y.; Hong, H. Hippo pathway contributes to cisplatin resistant-induced EMT in nasopharyngeal carcinoma cells. Cell Cycle 2017, 16, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, A. The role of hERG1 ion channels in epithelial-mesenchymal transition and the capacity of riluzole to reduce cisplatin resistance in colorectal cancer cells. Cell. Oncol. 2017, 40, 367–378. [Google Scholar] [CrossRef]
- Liu, S.L.; Lin, H.X.; Lin, C.Y.; Sun, X.Q.; Ye, L.P.; Qiu, F.; Wen, W.; Hua, X.; Wu, X.Q.; Li, J.; et al. TIMELESS confers cisplatin resistance in nasopharyngeal carcinoma by activating the Wnt/beta-catenin signaling pathway and promoting the epithelial mesenchymal transition. Cancer Lett. 2017, 402, 117–130. [Google Scholar] [CrossRef]
- Han, X.; Zhen, S.; Ye, Z.; Lu, J.; Wang, L.; Li, P.; Li, J.; Zheng, X.; Li, H.; Chen, W.; et al. A Feedback Loop Between miR-30a/c-5p and DNMT1 Mediates Cisplatin Resistance in Ovarian Cancer Cells. Cell. Physiol. Biochem. 2017, 41, 973–986. [Google Scholar] [CrossRef]
- Xiong, Y.; Sun, F.; Dong, P.; Watari, H.; Yue, J.; Yu, M.F.; Lan, C.Y.; Wang, Y.; Ma, Z.B. iASPP induces EMT and cisplatin resistance in human cervical cancer through miR-20a-FBXL5/BTG3 signaling. J. Exp. Clin. Cancer Res. 2017, 36, 48. [Google Scholar] [CrossRef]
- Feng, S.; Yang, G.; Yang, H.; Liang, Z.; Zhang, R.; Fan, Y.; Zhang, G. NEDD4 is involved in acquisition of epithelial-mesenchymal transition in cisplatin-resistant nasopharyngeal carcinoma cells. Cell Cycle 2017, 16, 869–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Guo, H.; Yao, B.; Helms, J. miR-15b inhibits cancer-initiating cell phenotypes and chemoresistance of cisplatin by targeting TRIM14 in oral tongue squamous cell cancer. Oncol. Rep. 2017, 37, 2720–2726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Qi, B.; Xiaoxiang, W.; Xu, J.; Liu, X. Baicalein increases cisplatin sensitivity of A549 lung adenocarcinoma cells via PI3K/Akt/NF-kappaB pathway. Biomed. Pharmacother. 2017, 90, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.J.; He, C.L.; Sun, T.; Duan, X.J.; Sun, Y.; Xiong, S.J. hsa-miR-485-5p reverses epithelial to mesenchymal transition and promotes cisplatin-induced cell death by targeting PAK1 in oral tongue squamous cell carcinoma. Int. J. Mol. Med. 2017, 40, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Ge, L.; Li, D.S.; Chen, F.; Feng, J.D.; Li, B.; Wang, T.J. TAZ overexpression is associated with epithelial-mesenchymal transition in cisplatin-resistant gastric cancer cells. Int. J. Oncol. 2017, 51, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zheng, C.; Xu, H.; He, W.; Ruan, Y.; Ma, J.; Zheng, J.; Ye, C.; Li, W. Inhibition of AMPK-related kinase 5 (ARK5) enhances cisplatin cytotoxicity in non-small cell lung cancer cells through regulation of epithelial-mesenchymal transition. Am. J. Transl. Res. 2017, 9, 1708–1719. [Google Scholar]
- Kuang, P.; Chen, Z.; Wang, J.; Liu, Z.; Wang, J.; Gao, J.; Shen, L. Characterization of Aurora A and Its Impact on the Effect of Cisplatin-Based Chemotherapy in Patients with Non-Small Cell Lung Cancer. Transl. Oncol. 2017, 10, 367–377. [Google Scholar] [CrossRef]
- Bugide, S.; Gonugunta, V.K.; Penugurti, V.; Malisetty, V.L.; Vadlamudi, R.K.; Manavathi, B. HPIP promotes epithelial-mesenchymal transition and cisplatin resistance in ovarian cancer cells through PI3K/AKT pathway activation. Cell. Oncol. 2017, 40, 133–144. [Google Scholar] [CrossRef]
- Roberts, C.M.; Tran, M.A.; Pitruzzello, M.C.; Wen, W.; Loeza, J.; Dellinger, T.H.; Mor, G.; Glackin, C.A. TWIST1 drives cisplatin resistance and cell survival in an ovarian cancer model, via upregulation of GAS6, L1CAM, and Akt signalling. Sci. Rep. 2016, 6, 37652. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Yin, S.; Su, Z.; Bai, M.; Zhang, H.; Hei, Z.; Cai, S. Downregulation of miR-101 contributes to epithelial-mesenchymal transition in cisplatin resistance of NSCLC cells by targeting ROCK2. Oncotarget 2016, 7, 37524–37535. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, F.; Wang, X.; Tsai, Y.; Chuang, K.H.; Keng, P.C.; Lee, S.O.; Chen, Y. A FASN-TGF-beta1-FASN regulatory loop contributes to high EMT/metastatic potential of cisplatin-resistant non-small cell lung cancer. Oncotarget 2016, 7, 55543. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Li, Y. miR-25-3p reverses epithelial-mesenchymal transition via targeting Sema4C in cisplatin-resistance cervical cancer cells. Cancer Sci. 2017, 108, 23–31. [Google Scholar] [CrossRef]
- Li, C.; Ding, H.; Tian, J.; Wu, L.; Wang, Y.; Xing, Y.; Chen, M. Forkhead Box Protein C2 Promotes Epithelial-Mesenchymal Transition, Migration and Invasion in Cisplatin-Resistant Human Ovarian Cancer Cell Line (SKOV3/CDDP). Cell. Physiol. Biochem. 2016, 39, 1098–1110. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Yang, H.; Chen, Y.; He, B.; Chen, Q. Kruppel-Like Factor 4 Enhances Sensitivity of Cisplatin to Lung Cancer Cells and Inhibits Regulating Epithelial-to-Mesenchymal Transition. Oncol. Res. 2016, 24, 81–87. [Google Scholar] [CrossRef]
- Zhen, Y.; Fang, W.; Zhao, M.; Luo, R.; Liu, Y.; Fu, Q.; Chen, Y.; Cheng, C.; Zhang, Y.; Liu, Z. miR-374a-CCND1-pPI3K/AKT-c-JUN feedback loop modulated by PDCD4 suppresses cell growth, metastasis, and sensitizes nasopharyngeal carcinoma to cisplatin. Oncogene 2017, 36, 275–285. [Google Scholar] [CrossRef]
- Piskareva, O.; Harvey, H.; Nolan, J.; Conlon, R.; Alcock, L.; Buckley, P.; Dowling, P.; Henry, M.; O’Sullivan, F.; Bray, I.; et al. The development of cisplatin resistance in neuroblastoma is accompanied by epithelial to mesenchymal transition in vitro. Cancer Lett. 2015, 364, 142–155. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Shen, H.; Yin, X.; Long, L.; Xie, C.; Liu, Y.; Hui, L.; Lin, X.; Fang, Y.; Cao, Y.; et al. miR-186 regulation of Twist1 and ovarian cancer sensitivity to cisplatin. Oncogene 2016, 35, 323–332. [Google Scholar] [CrossRef]
- Gu, J.; Liang, Y.; Qiao, L.; Lu, Y.; Hu, X.; Luo, D.; Li, N.; Zhang, L.; Chen, Y.; Du, J.; et al. URI expression in cervical cancer cells is associated with higher invasion capacity and resistance to cisplatin. Am. J. Cancer Res. 2015, 5, 1353–1367. [Google Scholar]
- Boac, B.M.; Xiong, Y.; Marchion, D.C.; Abbasi, F.; Bush, S.H.; Ramirez, I.J.; Khulpateea, B.R.; Clair McClung, E.; Berry, A.L.; Bou Zgheib, N.; et al. Micro-RNAs associated with the evolution of ovarian cancer cisplatin resistance. Gynecol. Oncol. 2016, 140, 259–263. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chen, X.; Ward, T.; Pegram, M.; Shen, K. Combined niclosamide with cisplatin inhibits epithelial-mesenchymal transition and tumor growth in cisplatin-resistant triple-negative breast cancer. Tumor Boil. 2016, 37, 9825–9835. [Google Scholar] [CrossRef]
- Huang, D.; Duan, H.; Huang, H.; Tong, X.; Han, Y.; Ru, G.; Qu, L.; Shou, C.; Zhao, Z. Cisplatin resistance in gastric cancer cells is associated with HER2 upregulation-induced epithelial-mesenchymal transition. Sci. Rep. 2016, 6, 20502. [Google Scholar] [CrossRef] [Green Version]
- Tieche, C.C.; Peng, R.W.; Dorn, P.; Froment, L.; Schmid, R.A.; Marti, T.M. Prolonged pemetrexed pretreatment augments persistence of cisplatin-induced DNA damage and eliminates resistant lung cancer stem-like cells associated with EMT. BMC Cancer 2016, 16, 125. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.; Liu, X.; Lin, F.; Li, P.; Liu, K.; Geng, R.; Dai, C.; Lin, Y.; Tang, W.; Wu, Z.; et al. MicroRNA-421 regulated by HIF-1alpha promotes metastasis, inhibits apoptosis, and induces cisplatin resistance by targeting E-cadherin and caspase-3 in gastric cancer. Oncotarget 2016, 7, 24466–24482. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Jiao, D.M.; Wang, J.; Hu, H.; Tang, X.; Chen, J.; Mou, H.; Lu, W. miR-206 regulates cisplatin resistance and EMT in human lung adenocarcinoma cells partly by targeting MET. Oncotarget 2016, 7, 24510–24526. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Hong, H.; Sun, X.; Jiang, H.; Ma, S.; Zhao, S.; Zhang, M.; Wang, Z.; Jiang, C.; Liu, H. MicroRNA-10b regulates epithelial-mesenchymal transition by modulating KLF4/Notch1/E-cadherin in cisplatin-resistant nasopharyngeal carcinoma cells. Am. J. Cancer Res. 2016, 6, 141–156. [Google Scholar]
- Wang, L.L.; Zhang, X.H.; Zhang, X.; Chu, J.K. MiR-30a increases cisplatin sensitivity of gastric cancer cells through suppressing epithelial-to-mesenchymal transition (EMT). Eur. Rev. Med Pharmacol. Sci. 2016, 20, 1733–1739. [Google Scholar]
- Xu, G.; Yu, H.; Shi, X.; Sun, L.; Zhou, Q.; Zheng, D.; Shi, H.; Li, N.; Zhang, X.; Shao, G. Cisplatin sensitivity is enhanced in non-small cell lung cancer cells by regulating epithelial-mesenchymal transition through inhibition of eukaryotic translation initiation factor 5A2. BMC Pulm. Med. 2014, 14, 174. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Yang, B.; Shi, S.; Jiang, X. RNA interference (RNAi) mediated stable knockdown of protein casein kinase 2-alpha (CK2alpha) inhibits migration and invasion and enhances cisplatin-induced apoptosis in HEp-2 laryngeal carcinoma cells. Acta Histochem. 2014, 116, 1000–1006. [Google Scholar] [CrossRef]
- Yu, M.; Zhang, C.; Li, L.; Dong, S.; Zhang, N.; Tong, X. Cx43 reverses the resistance of A549 lung adenocarcinoma cells to cisplatin by inhibiting EMT. Oncol. Rep. 2014, 31, 2751–2758. [Google Scholar] [CrossRef]
- Jiang, Z.; Yin, J.; Fu, W.; Mo, Y.; Pan, Y.; Dai, L.; Huang, H.; Li, S.; Zhao, J. MiRNA 17 family regulates cisplatin-resistant and metastasis by targeting TGFbetaR2 in NSCLC. PLoS ONE 2014, 9, e94639. [Google Scholar] [CrossRef] [Green Version]
- Sobral, L.M.; Sousa, L.O.; Coletta, R.D.; Cabral, H.; Greene, L.J.; Tajara, E.H.; Gutkind, J.S.; Curti, C.; Leopoldino, A.M. Stable SET knockdown in head and neck squamous cell carcinoma promotes cell invasion and the mesenchymal-like phenotype in vitro, as well as necrosis, cisplatin sensitivity and lymph node metastasis in xenograft tumor models. Mol. Cancer 2014, 13, 32. [Google Scholar] [CrossRef] [Green Version]
- Baribeau, S.; Chaudhry, P.; Parent, S.; Asselin, E. Resveratrol inhibits cisplatin-induced epithelial-to-mesenchymal transition in ovarian cancer cell lines. PLoS ONE 2014, 9, e86987. [Google Scholar] [CrossRef] [Green Version]
- Wolf, M.A.; Claudio, P.P. Benzyl isothiocyanate inhibits HNSCC cell migration and invasion, and sensitizes HNSCC cells to cisplatin. Nutr. Cancer 2014, 66, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.G.; Wei, W.; Xia, L.H.; Han, W.L.; Zhao, P.; Wu, S.J.; Li, W.D.; Chen, W. FBW7 upregulation enhances cisplatin cytotoxicity in non- small cell lung cancer cells. Asian Pac. J. Cancer Prev. 2013, 14, 6321–6326. [Google Scholar] [CrossRef] [Green Version]
- Pennati, M.; Lopergolo, A.; Profumo, V.; De Cesare, M.; Sbarra, S.; Valdagni, R.; Zaffaroni, N.; Gandellini, P.; Folini, M. miR-205 impairs the autophagic flux and enhances cisplatin cytotoxicity in castration-resistant prostate cancer cells. Biochem. Pharmacol. 2014, 87, 579–597. [Google Scholar] [CrossRef]
- Liu, H.; Gu, Y.; Yin, J.; Zheng, G.; Wang, C.; Zhang, Z.; Deng, M.; Liu, J.; Jia, X.; He, Z. SET-mediated NDRG1 inhibition is involved in acquisition of epithelial-to-mesenchymal transition phenotype and cisplatin resistance in human lung cancer cell. Cell. Signal. 2014, 26, 2710–2720. [Google Scholar] [CrossRef]
- Xiong, X.; Arvizo, R.R.; Saha, S.; Robertson, D.J.; McMeekin, S.; Bhattacharya, R.; Mukherjee, P. Sensitization of ovarian cancer cells to cisplatin by gold nanoparticles. Oncotarget 2014, 5, 6453–6465. [Google Scholar] [CrossRef]
- Fan, G.L.; Yao, Y.; Yao, L.; Li, Y. PDCD5 transfection increases cisplatin sensitivity and decreases invasion in hepatic cancer cells. Oncol. Lett. 2015, 9, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.H.; Chen, M.T.; Wang, M.L.; Lee, Y.Y.; Chiou, G.Y.; Chien, C.S.; Huang, P.I.; Chen, Y.W.; Huang, M.C.; Chiou, S.H.; et al. Cisplatin-selected resistance is associated with increased motility and stem-like properties via activation of STAT3/Snail axis in atypical teratoid/rhabdoid tumor cells. Oncotarget 2015, 6, 1750–1768. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Zhang, L.; Yao, Q.; Tao, Z. miR-15b regulates cisplatin resistance and metastasis by targeting PEBP4 in human lung adenocarcinoma cells. Cancer Gene Ther. 2015, 22, 108–114. [Google Scholar] [CrossRef]
- Chen, D.J.; Chen, W.; Jiang, H.; Yang, H.; Wang, Y.C.; Chen, J.H. Downregulation of DOCK1 sensitizes bladder cancer cells to cisplatin through preventing epithelial-mesenchymal transition. Drug Des. Dev. Ther. 2016, 10, 2845–2853. [Google Scholar] [CrossRef] [Green Version]
- Quiles, J.L.; Sánchez-González, C.; Vera-Ramirez, L.; Giampieri, F.; Navarro-Hortal, M.D.; Xiao, J.; Llopis, J.; Battino, M.; Varela-Lopez, A. Reductive Stress, Bioactive Compounds, Redox-Active Metals and Dormant Tumor Cell Biology to Develop Redox-Based Tools for the Treatment of Cancer. Antioxid. Redox Signal. 2020. [Google Scholar] [CrossRef]
- Flores-Guzmán, F.; Utikal, J.; Umansky, V. Dormant tumor cells interact with memory CD8+ T cells in RET transgenic mouse melanoma model. Cancer Lett. 2020, 474, 74–81. [Google Scholar] [CrossRef]
- Barney, L.E.; Hall, C.L.; Schwartz, A.D.; Parks, A.N.; Sparages, C.; Galarza, S.; Platt, M.O.; Mercurio, A.M.; Peyton, S.R. Tumor cell–organized fibronectin maintenance of a dormant breast cancer population. Sci. Adv. 2020, 6, eaaz4157. [Google Scholar] [CrossRef] [Green Version]
- Hen, O.; Barkan, D. Dormant disseminated tumor cells and cancer stem/progenitor-like cells: Similarities and opportunities. Semin. Cancer Boil. 2020, 60, 157–165. [Google Scholar] [CrossRef]



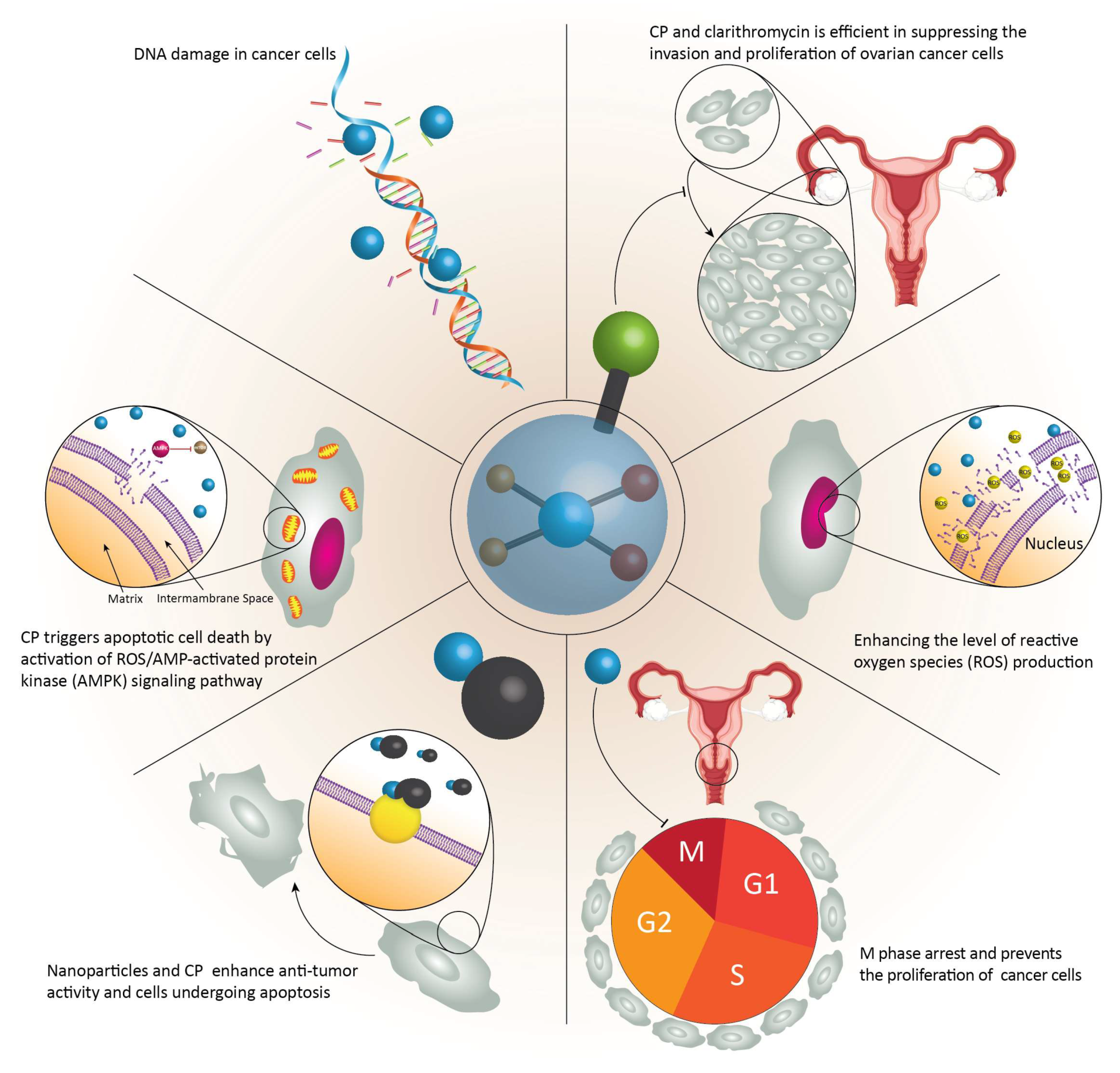{kind=link}
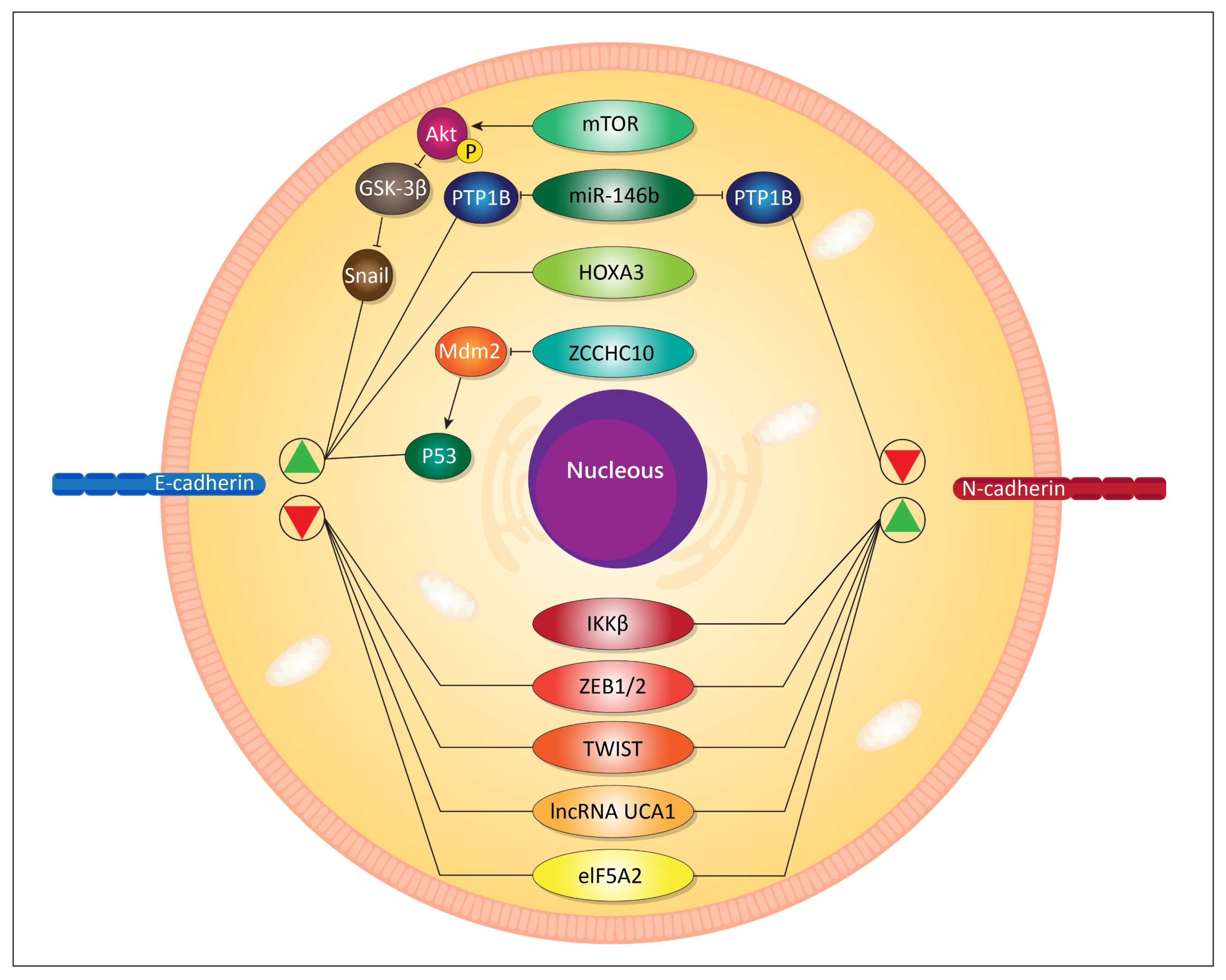{kind=link}
| Cancer Type | Signaling Axis | Effect on EMT | Results | Refs |
|---|---|---|---|---|
| Lung cancer | EMT/PD-L1/PD-1 | - | The CP-resistant lung cancer cells activate EMT to induce the expression of PD-L1/PD-1, ensuring their survival and resistance to CP therapy. Anti-PD-1 or anti-PD-L1 therapies may lead to the sensitivity of cancer cells in CP chemotherapy | [426] |
| Lung cancer | TGF-β1/EMT | Induction | TGF-β1 is able to stimulate EMT via enhancing N-cadherin and vimentin and decreasing E-cadherin. In addition, drug-resistant proteins such as ERCC1 and p-glycoprotein undergo upregulation. Inhibition of TGF-β1 enhances the sensitivity of cancer cells in CP therapy | [362] |
| Squamous cell carcinoma | SOX8/Wnt-β-catenin/EMT | Induction | SOX8 induces EMT through Frizzled-7-mediated Wnt signaling pathway to diminish the efficacy of CP chemotherapy | [427] |
| Lung cancer | MF-κB/EMT | Induction | The administration of ginsenoside Rg3 sensitizes cancer cells in CP chemotherapy by inhibition of NF-κB-induced EMT | [428] |
| Ovarian cancer | - | - | PD98059 induces EMT mechanism to reduce the sensitivity of cancer cells to CP chemotherapy | [429] |
| Breast cancer | NF-κB/EMT | Induction | Eugenol inhibits EMT through NF-κB downregulation, leading to the enhanced anti-tumor activity of CP | [430] |
| Cervical cancer | RIF1/EMT | Induction | RIF1 enhances the malignancy of cancer cells through EMT induction. Suppressing RIF1 sensitizes cancer cells in CP chemotherapy | [431] |
| Breast cancer | αvβ3 integrin/EMT FAK/PI3K/Akt/EMT | Induction | 14, 15-EET trigger EMT through activation of FAK/PI3K/Akt and αvβ3 integrin to ensure the resistance of cancer cells in CP chemotherapy | [432] |
| Ovarian cancer | Notch3/SUSD2/EMT | Induction | The downstream mediator of Notch3, SUSD4 induces EMT to trigger the resistance of cancer cells in CP chemotherapy | [433] |
| Lung cancer | TGF-β1/EMT | Induction | Inhibition of TGF-β1 is associated with EMT downregulation and consequently, the sensitivity of cancer cells in CP chemotherapy | [434] |
| Ovarian cancer | MiR-20a/EMT | Induction | The miR-20a induces CP resistance in cancer cells through EMT induction | [435] |
| Non-small cell lung cancer | SLC39A4/EMT | Induction | The inhibition of SLC39A4 suppresses EMT and sensitizes cancer cells in CP chemotherapy | [436] |
| Nasopharyngeal carcinoma | Hippo/TAZ/EMT | Induction | The TAZ activation as a key gene of Hippo pathway induces EMT and triggers the resistance of cancer cells in CP chemotherapy | [437] |
| Colorectal cancer | hERG1/EMT | Induction | The hERG1 ion channels induce EMT to diminish the sensitivity of cancer cells in CP chemotherapy | [438] |
| Nasopharyngeal carcinoma | TIMELESS/EMT | Induction | Suppressing TIMELESS expression inhibits EMT to enhance the efficacy of CP chemotherapy | [439] |
| Ovarian cancer | MiR-30a/c-5p/DNMT1/EMT | Inhibition | The overexpressed miR-30a/c-5p inhibits EMT through DNMT1 downregulation, leading to the sensitivity of cancer cells in CP chemotherapy | [440] |
| Cervical cancer | iASPP/miR-20a/FBXL5/BTG3/EMT | Induction | The iASPP inhibits FBXL5/BTG3 axis through miR-20a upregulation. As a consequence, EMT mechanism occurs to ensure the resistance of cancer cells in CP chemotherapy | [441] |
| Nasopharyngeal carcinoma | NEDD4/EMT | Induction | Knock-out of NEDD4 inhibits EMT and induces mesenchymal–epithelial transition, resulting in sensitivity of cancer cells in CP chemotherapy | [442] |
| Oral tongue squamous cell cancer | MiR-15b/TRIM14/EMT | Inhibition | The upregulated miR-15b suppresses EMT via TRIM14 downregulation to induce mesenchymal –epithelial transition and sensitize cancer cells in CP chemotherapy | [443] |
| Lung adenocarcinoma | PI3K/Akt/NF-κB/EMT | Induction | The administration of baicalein inhibits the PI3K/Akt/NF-κB-mediated EMT to sensitize cancer cells in CP chemotherapy and reduce their malignancy and migration | [444] |
| Oral tongue squamous cell carcinoma | has-miR-485-5p/PAK1/ERCC1-YAP/EMT | Inhibition | The downstream mediators of PAK1 including ERCC1 and YAP are involved in induction of EMT and resistance of cancer cells in CP chemotherapy. The hsa-miR-485-5p disrupts the aforementioned axis to suppresses EMT and malignancy, and restore the sensitivity to CP treatment | [445] |
| Gastric cancer | TAZ/EMT | Induction | The overexpression of TAZ stimulates EMT to ensure the malignancy and invasion of cancer cells, leading to resistance in CP therapy | [446] |
| Non-small cell lung cancer | ARK5/EMT | Induction | The ARK5 triggers EMT to enhance the malignancy and metastasis of cancer cells. The silencing of ARK5 sensitizes cancer cells in CP chemotherapy through EMT inhibition | [447] |
| Non-small cell lung cancer | Aurora A/EMT | Induction | The downregulation of Aurora A inhibits EMT, leading to the sensitivity of cancer cells in CP chemotherapy | [448] |
| Ovarian cancer | HPIP/PI3K/Akt/EMT | Induction | HPIP elevates the malignancy and invasion of cancer cells by EMT induction via PI3K/Akt axis. This enhanced malignancy leads to the resistance in CP chemotherapy | [449] |
| Ovarian cancer | EMT/Akt | - | The EMT induces Akt activation to drive the resistance in CP chemotherapy | [450] |
| Non-small cell lung cancer | MiR-101/ROCK2/EMT | Inhibition | The miR-101 has a reverse relationship with ROCK2 to inhibit EMT and promote the sensitivity of cancer cells in CP chemotherapy | [451] |
| Non-small cell lung cancer | FASN/TGF-β1/FASN | Induction | A positive loop between FASN and TGF-β1 induces EMT and triggers CP resistance | [452] |
| Cervical cancer | MiR-25-3p/Sema4C/EMT | Inhibition | By inhibition of Sema4C, miR-25-3p suppresses EMT to inhibit cancer metastasis and malignancy and sensitize them in CP chemotherapy | [453] |
| Ovarian cancer | FOXC2/ERK/EMT FOXC2/Akt/GSK-3β/EMT | Induction | The FOXC2 activates EMT mechanism via ERK and Akt/GSK-3β signaling pathways to induce the resistance of cancer cells in CP chemotherapy | [454] |
| Lung cancer | KLF4/EMT | Inhibition | The upregulation of KLF4 inhibits EMT mechanism through enhancing Slug, Twist and vimentin levels, and reducing E-cadherin levels, resulting in decreased malignancy and induction of apoptosis by CP chemotherapy | [455] |
| Nasopharyngeal carcinoma | MiR-374a/PDCD4/CCND1/ PI3K/Akt/c-Jun/EMT | Inhibition | The miR-374a downregulates the expression of CCND1 by induction of tumor suppressor PDCD4. Then, a decrease occurs in PI3K/Akt/c-Jun signaling pathway to inhibit EMT, leading to the sensitivity of cancer cells in CP chemotherapy | [456] |
| Neuroblastoma | MYH9 ACTN4 ROCK1 | Induction Induction Inhibition | Overexpressed MYH9 and ACTN4, and decreased expression of ROCK1, are involved in EMT and resistance of cancer cells in CP chemotherapy | [457] |
| Ovarian cancer | MiR-186/Twist1/EMT | Inhibition | The miR-186 inhibits EMT through Twist1 downregulation to drive the sensitivity of cancer cells in CP chemotherapy | [458] |
| Cervical cancer | URI/EMT | Induction | The silencing of URI inhibits EMT to sensitize cancer cells in CP chemotherapy | [459] |
| Ovarian cancer | MiR-496 MiR-485-5p MiR-152 Let-7g | Induction | These miRs are associated with induction of EMT and resistance of cancer cells in CP chemotherapy | [460] |
| Triple negative breast cancer | - | - | The administration of niclosamide inhibits EMT to enhance the induction of apoptotic cell death | [461] |
| Gastric cancer | HER2/Snail/EMT | Induction | The HER2 activates Snail/EMT axis to reduce the efficacy of CP chemotherapy | [462] |
| Lung cancer | - | - | Pemetrexed pretreatment inhibits EMT to sensitize cancer cells in CP chemotherapy | [463] |
| Gastric cancer | HIF-α/miR-421/EMT | Induction | The HIF-α-mediated miR-421 upregulation reduces E-cadherin levels, leading to the EMT stimulation and CP resistance | [464] |
| Lung adenocarcinoma | MiR-206/MET/PI3K/AKT/ mTOR/EMT | Inhibition | The inactivation of MET/PI3K/Akt/mTOR axis by miR-206 inhibits EMT and drives the sensitivity of cancer cells in CP chemotherapy | [465] |
| Nasopharyngeal carcinoma | MiR-10b/KLF4/Notch1/E-cadherin | Induction | The miR-10b upregulates the expression of Notch1 via KLF4 inhibition to reduce E-cadherin levels, leading to the stimulation of EMT and CP resistance | [466] |
| Gastric cancer | MiR-30a/EMT | Inhibition | The overexpressed miR-30a reduces EMT to sensitize cancer cells in CP chemotherapy | [467] |
| Non-small cell lung cancer | elF5A2/EMT | Induction | GC7 inhibits EMT through elF5A2 downregulation, resulting in sensitivity of cancer cells in CP chemotherapy | [468] |
| Laryngeal carcinoma cells | CK2α/EMT | Induction | Silencing of CK2α suppresses EMT to inhibit malignancy and resistance of cancer cells in CP chemotherapy | [469] |
| Lung adenocarcinoma | Cx43/EMT | Inhibition | Cx43 reduces the malignancy and invasion of cancer cells through EMT inhibition, leading to the sensitivity in CP chemotherapy | [470] |
| Non-small cell lung cancer | MiR-17 MiR-20a MiR-20b | Inhibition | These miRs are able to suppress EMT and sensitizing cancer cells in CP chemotherapy | [471] |
| Head and neck squamous cell carcinoma | SET/EMT | Induction | As an oncogenic factor, SET induces EMT to reduce the sensitivity of cancer cells in CP chemotherapy | [472] |
| Ovarian cancer | ERK/Snail/EMT | Induction | The administration of resveratrol diminishes the expression of ERK to inhibit Snail and EMT, leading to the sensitivity of cancer cells in CP chemotherapy | [473] |
| Head and neck squamous cell carcinoma | - | - | Benzyl isothiocyanate suppresses CP resistance through EMT downregulation | [474] |
| Non-small cell lung cancer | FBW7/EMT | Inhibition | The overexpression of FBW7 inhibits EMT to enhance the sensitivity of cancer cells in CP chemotherapy | [475] |
| Prostate cancer | MiR-205/autophagy/EMT | Inhibition | The overexpression of miR-205 inhibits autophagy to suppress EMT and sensitize cancer cells in CP chemotherapy | [476] |
| Lung cancer | SET/NDRG1/EMT | Induction | The inhibition of NDRG by SET triggers EMT, leading to the resistance of cancer cells in CP chemotherapy | [477] |
| Ovarian cancer | Akt/EMT NF-κB/EMT | Induction | Gold nanoparticles are able to inhibit Akt and NF-κB signaling pathways to suppress EMT and CP resistance | [478] |
| Hepatic cancer | PDCD5/TGF-β/EMT | Inhibition | By downregulation of TGF-β, PDCD5 inhibits EMT to sensitize cancer cells in CP chemotherapy | [479] |
| Atypical teratoid/rhabdoid tumor | STAT3/Snail/EMT | Induction | Inhibition of STAT3/Snail axis suppresses EMT and partially sensitizes cancer cells in CP chemotherapy | [480] |
| Lung adenocarcinoma | MiR-15b/PEBP4/EMT | Induction | By downregulation of PEBP4, miR-15b triggers EMT mechanism to mediate the resistance of cancer cells in CP chemotherapy | [481] |
| Bladder cancer | DOCK1/EMT | Induction | Silencing DOCK1 expression is associated with sensitivity of cancer cells to CP via EMT inhibition | [482] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Kalantari, M.; Mohammadinejad, R.; Javaheri, T.; Sethi, G. Association of the Epithelial–Mesenchymal Transition (EMT) with Cisplatin Resistance. Int. J. Mol. Sci. 2020, 21, 4002. https://doi.org/10.3390/ijms21114002
Ashrafizadeh M, Zarrabi A, Hushmandi K, Kalantari M, Mohammadinejad R, Javaheri T, Sethi G. Association of the Epithelial–Mesenchymal Transition (EMT) with Cisplatin Resistance. International Journal of Molecular Sciences. 2020; 21(11):4002. https://doi.org/10.3390/ijms21114002
Chicago/Turabian StyleAshrafizadeh, Milad, Ali Zarrabi, Kiavash Hushmandi, Mahshad Kalantari, Reza Mohammadinejad, Tahereh Javaheri, and Gautam Sethi. 2020. "Association of the Epithelial–Mesenchymal Transition (EMT) with Cisplatin Resistance" International Journal of Molecular Sciences 21, no. 11: 4002. https://doi.org/10.3390/ijms21114002
APA StyleAshrafizadeh, M., Zarrabi, A., Hushmandi, K., Kalantari, M., Mohammadinejad, R., Javaheri, T., & Sethi, G. (2020). Association of the Epithelial–Mesenchymal Transition (EMT) with Cisplatin Resistance. International Journal of Molecular Sciences, 21(11), 4002. https://doi.org/10.3390/ijms21114002