Dexfenfluramine and Pergolide Cause Heart Valve Disease via Valve Metabolic Reprogramming and Ongoing Matrix Remodeling

 , , and
, , and 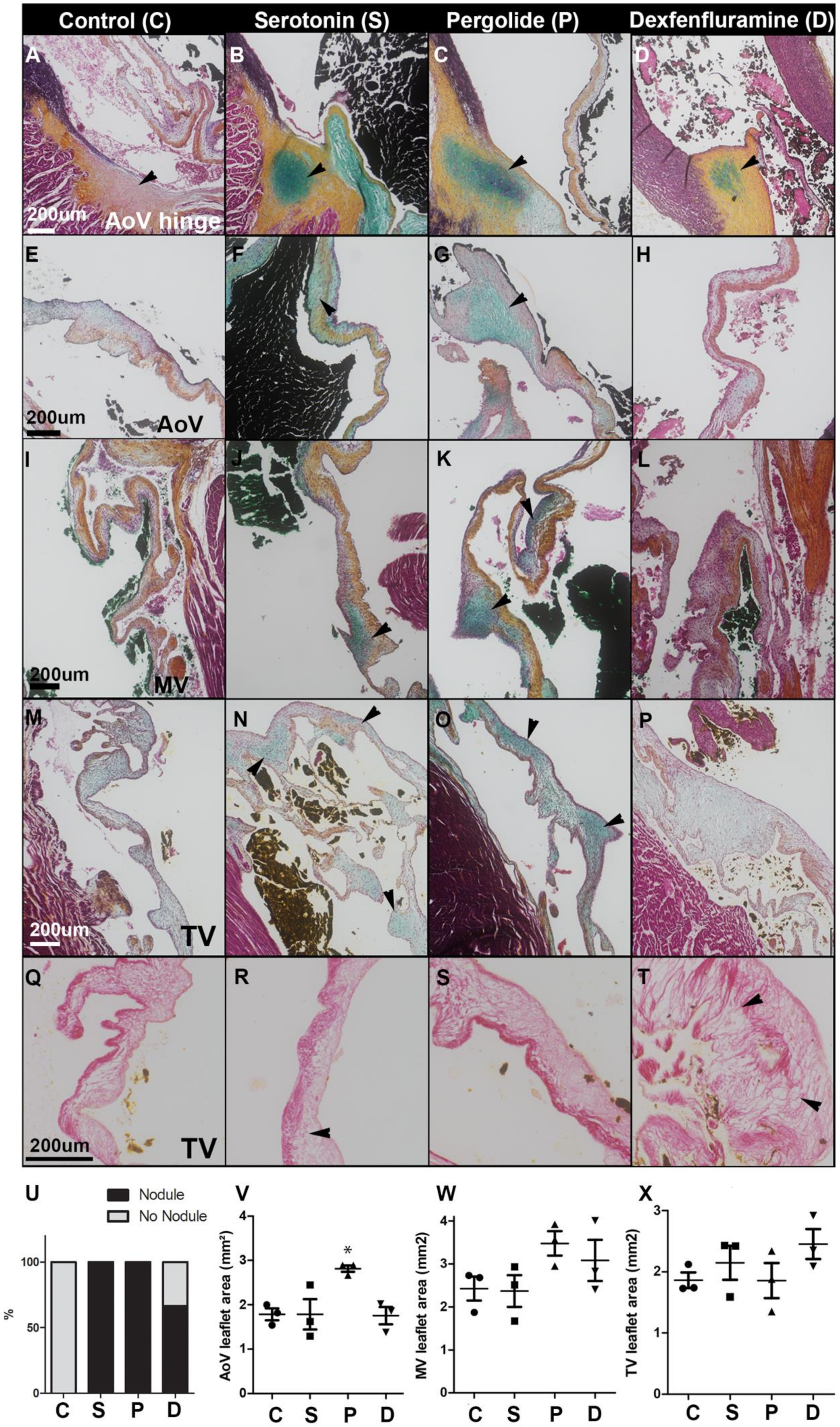{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Oral Serotonin, Pergolide, and Dexfenfluramine Cause Valvular Lesions with Distinct Histological Features
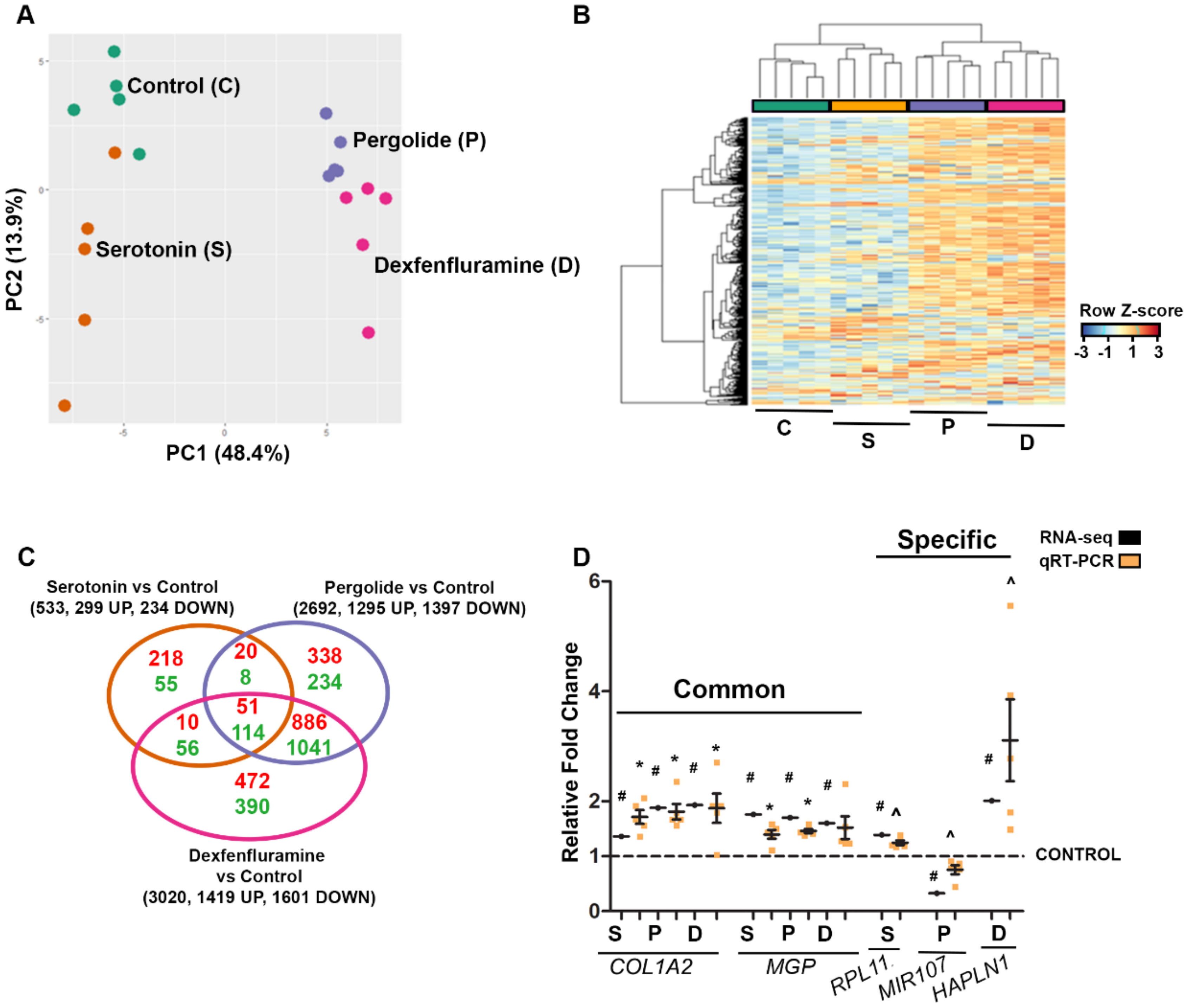
2.2. RNA Profiling Identifies Gene Expression Patterns of Drug-Induced Valvular Alterations
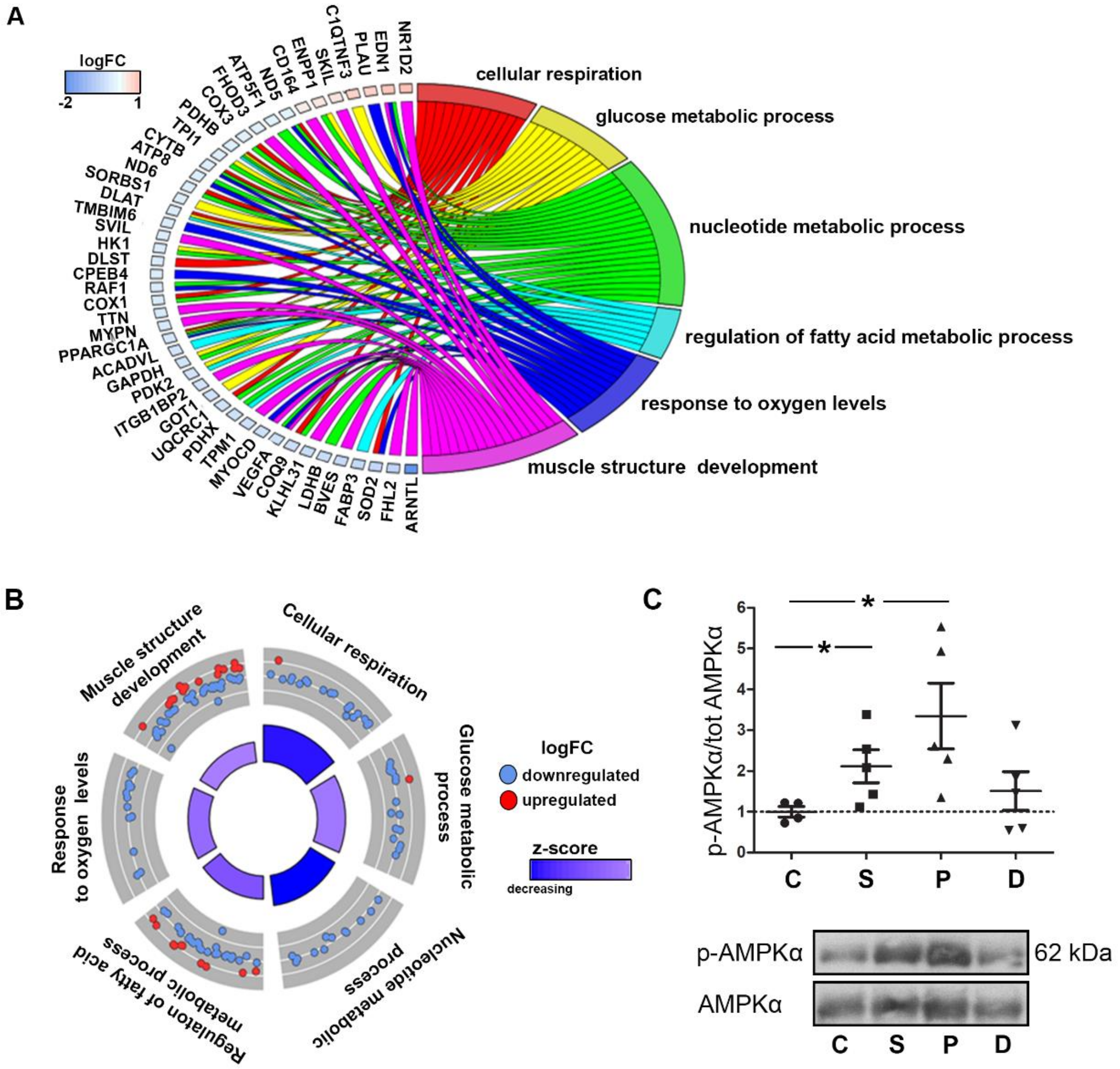
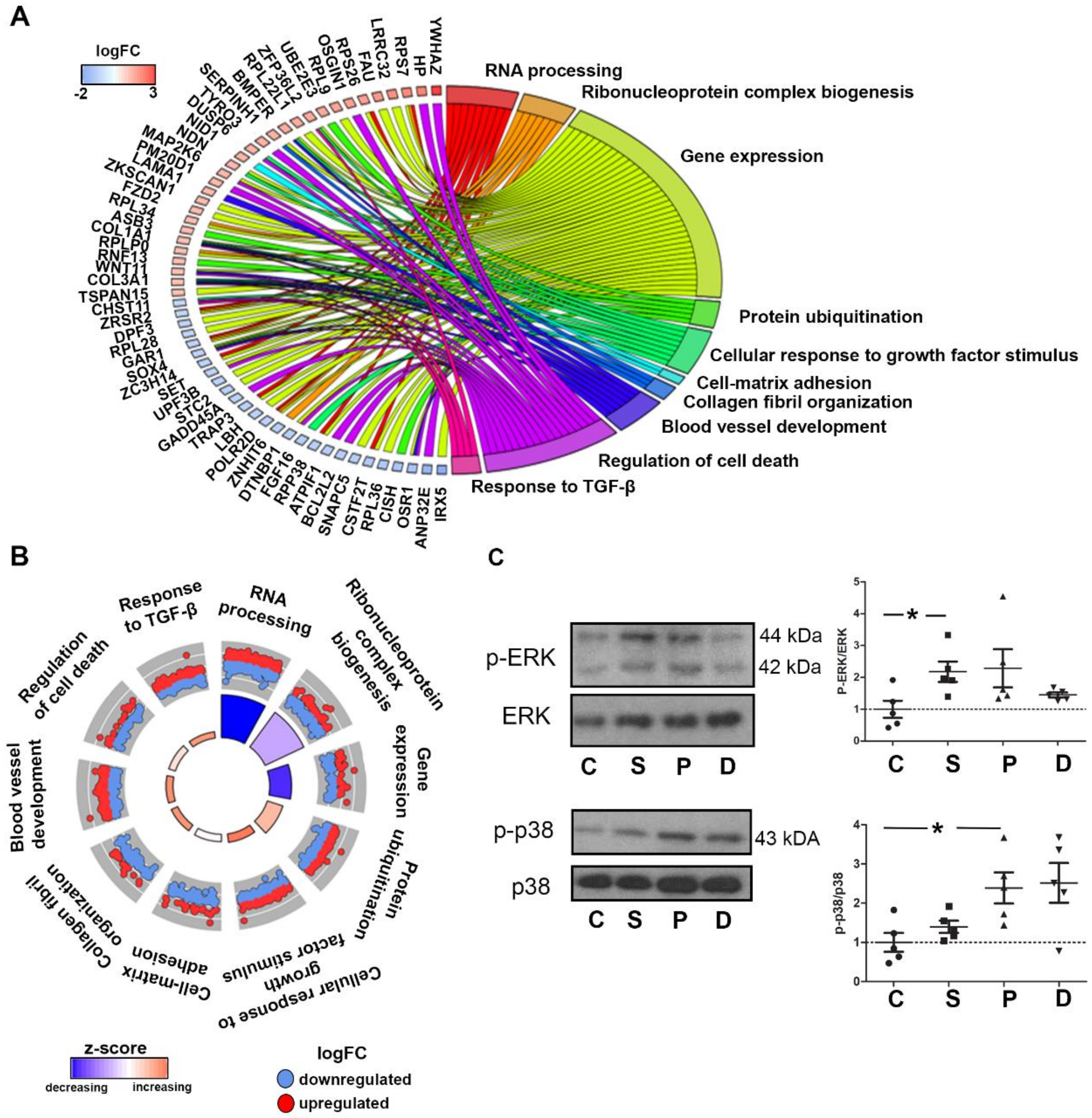
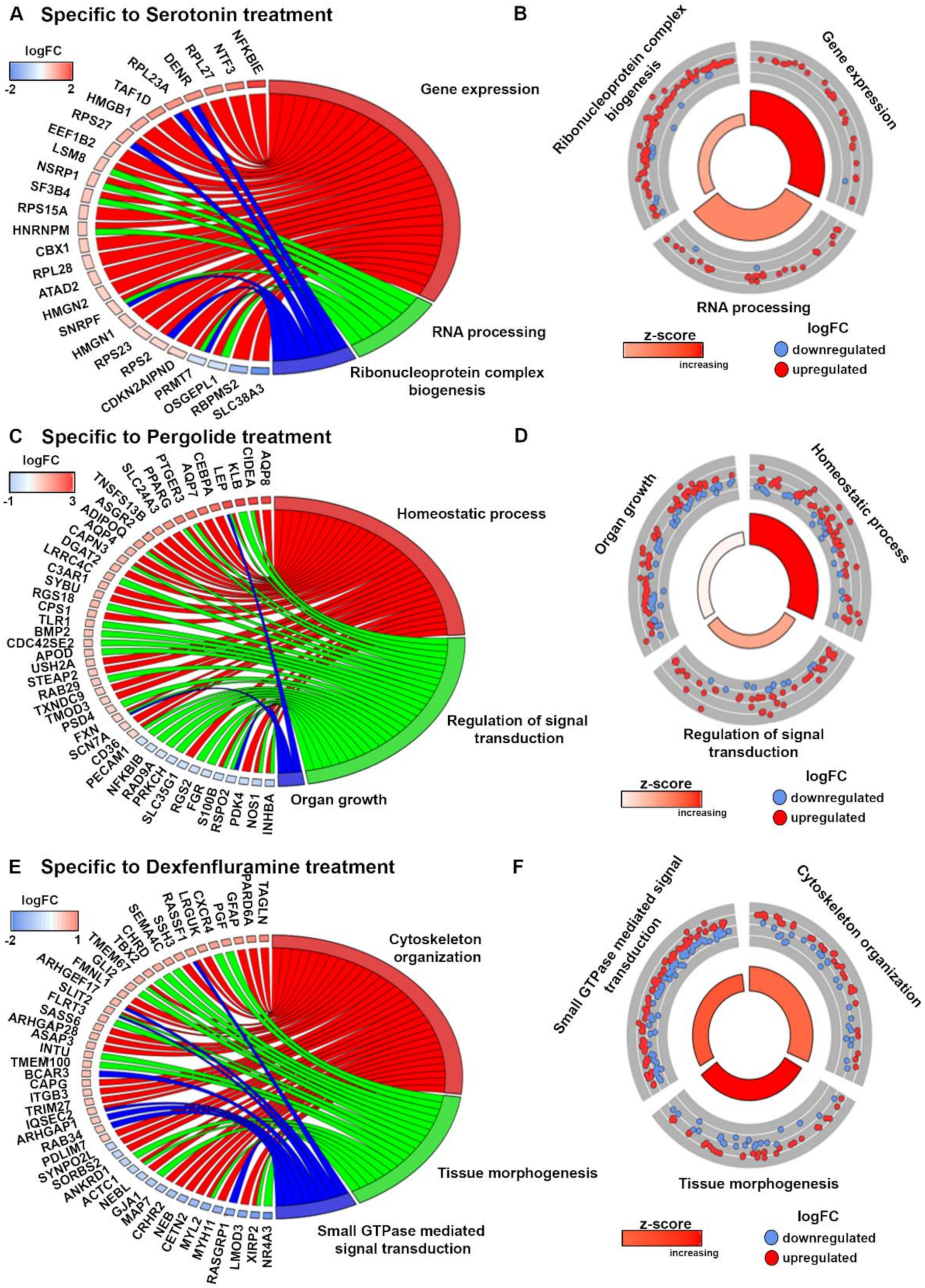
2.3. Gene Ontology Analyses Depict Enrichment of Distinct Biological Processes Leading to Valve Remodeling
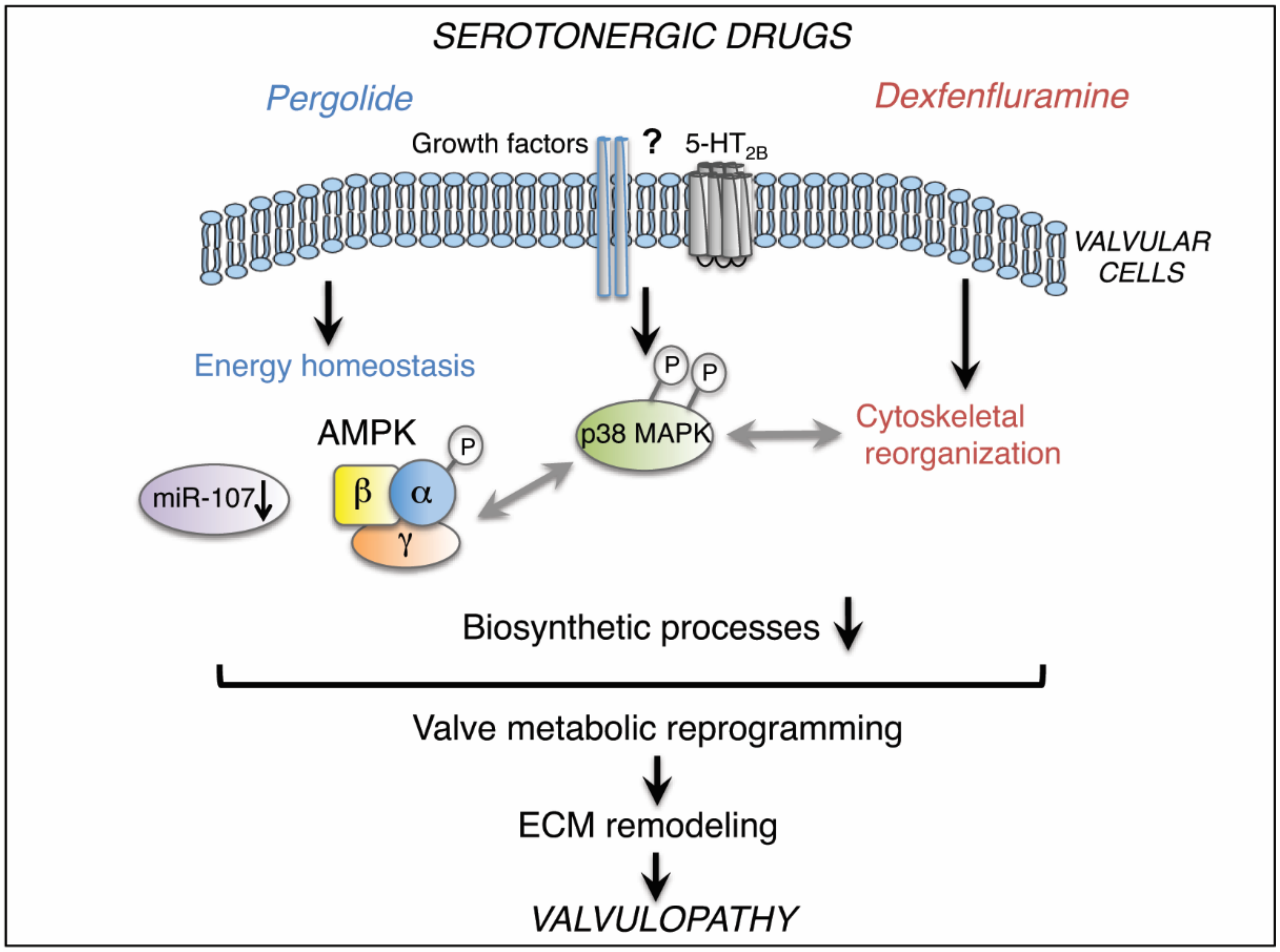
3. Discussion
4. Materials and Methods
4.1. Rabbits
4.2. Measurements of Pergolide and Dexfenfluramine in Rabbit Plasma
4.3. Histology and Immunohistochemistry
4.4. RNA Isolation
4.5. RNA Sequencing, Data Processing, and Analysis
4.6. Gene Enrichment Analysis
4.7. Real-Time Quantitative PCR
4.8. Protein Extraction and Western Blotting
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPK DEG | AMP-activated protein kinase Differentially expressed gene |
| ECM VHD VIC | Extracellular matrix Valvular heart disease Valvular interstitial cell |
| TGF-β | Transforming growth factor-β |
| MAPK | Mitogen-activated protein kinase |
References
- Bhattacharyya, S.; Schapira, A.H.; Mikhailidis, D.P.; Davar, J. Drug-induced fibrotic valvular heart disease. Lancet 2009, 374, 577–585. [Google Scholar] [CrossRef]
- Tribouilloy, C.; Marechaux, S.; Jobic, Y.; Jeu, A.; Ederhy, S.; Donal, E.; Reant, P.; Arnalsteen, E.; Boulanger, J.; Garban, T.; et al. Frequency of drug-induced valvular heart disease in patients previously exposd to benfluorex: A multicentre prospective study. Eur. Heart J. 2013, 34, 3580–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosyns, B.; Droogmans, S.; Rosenhek, R.; Lancellotti, P. Drug-induced valvular heart disease. Heart 2013, 99, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, B.I.; Tommeras, K.; Nordrum, I.; Loennechen, J.P.; Brunsvik, A.; Solligard, E.; Fossmark, R.; Bakke, I.; Syversen, U.; Waldum, H. Long-term serotonin administration induces heart valve disease in rats. Circulation 2005, 111, 1517–1522. [Google Scholar] [CrossRef]
- Lancellotti, P.; Nchimi, A.; Hego, A.; Dulgheru, R.; Delvenne, P.; Drion, P.; Oury, C. High-dose oral intake of serotonin induces valvular heart disease in rabbits. Int. J. Cardiol. 2015, 197, 72–75. [Google Scholar] [CrossRef]
- Droogmans, S.; Roosens, B.; Cosyns, B.; Degaillier, C.; Hernot, S.; Weytjens, C.; Garbar, C.; Caveliers, V.; Pipeleers-Marichal, M.; Franken, P.R.; et al. Cyproheptadine prevents pergolide-induced valvulopathy in rats: An echocardiographic and histopathological study. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1940–H1948. [Google Scholar] [CrossRef]
- Ayme-Dietrich, E.; Lawson, R.; Cote, F.; de Tapia, C.; Da Silva, S.; Ebel, C.; Hechler, B.; Gachet, C.; Guyonnet, J.; Rouillard, H.; et al. The role of 5-ht2b receptors in mitral valvulopathy: Bone marrow mobilization of endothelial progenitors. Br. J. Pharm. 2017, 174, 4123–4139. [Google Scholar] [CrossRef] [Green Version]
- Rothman, R.B.; Baumann, M.H.; Savage, J.E.; Rauser, L.; McBride, A.; Hufeisen, S.J.; Roth, B.L. Evidence for possible involvement of 5-ht(2b) receptors in the cardiac valvulopathy associated with fenfluramine and other serotonergic medications. Circulation 2000, 102, 2836–2841. [Google Scholar] [CrossRef] [Green Version]
- McDonald, P.C.; Wilson, J.E.; Gao, M.; McNeill, S.; Spinelli, J.J.; Williams, O.D.; Harji, S.; Kenyon, J.; McManus, B.M. Quantitative analysis of human heart valves: Does anorexigen exposure produce a distinctive morphological lesion? Cardiovasc. Pathol. Off. J. Soc. Cardiovasc. Pathol. 2002, 11, 251–262. [Google Scholar] [CrossRef]
- Hutcheson, J.D.; Ryzhova, L.M.; Setola, V.; Merryman, W.D. 5-ht(2b) antagonism arrests non-canonical tgf-beta1-induced valvular myofibroblast differentiation. J. Mol. Cell. Cardiol. 2012, 53, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Walker, G.A.; Masters, K.S.; Shah, D.N.; Anseth, K.S.; Leinwand, L.A. Valvular myofibroblast activation by transforming growth factor-beta: Implications for pathological extracellular matrix remodeling in heart valve disease. Circ. Res. 2004, 95, 253–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabkin, E.; Aikawa, M.; Stone, J.R.; Fukumoto, Y.; Libby, P.; Schoen, F.J. Activated interstitial myofibroblasts express catabolic enzymes and mediate matrix remodeling in myxomatous heart valves. Circulation 2001, 104, 2525–2532. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Leinwand, L.A.; Anseth, K.S. Cardiac valve cells and their microenvironment--insights from in vitro studies. Nat. Rev. Cardiol. 2014, 11, 715–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grande-Allen, K. Fibrotic vs. Myxomatous remodeling of mitral valves. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2004, 2004, 3737–3740. [Google Scholar]
- Liu, A.C.; Gotlieb, A.I. Transforming growth factor-beta regulates in vitro heart valve repair by activated valve interstitial cells. Am. J. Pathol. 2008, 173, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, P.; Despres, J.P.; Pibarot, P. The ‘valvulo-metabolic’ risk in calcific aortic valve disease. Can. J. Cardiol. 2007, 23 (Suppl. B), 32B–39B. [Google Scholar] [CrossRef]
- Drolet, M.C.; Roussel, E.; Deshaies, Y.; Couet, J.; Arsenault, M. A high fat/high carbohydrate diet induces aortic valve disease in c57bl/6j mice. J. Am. Coll. Cardiol. 2006, 47, 850–855. [Google Scholar] [CrossRef] [Green Version]
- Kamel, P.I.; Qu, X.; Geiszler, A.M.; Nagrath, D.; Harmancey, R.; Taegtmeyer, H.; Grande-Allen, K.J. Metabolic regulation of collagen gel contraction by porcine aortic valvular interstitial cells. J. R. Soc. Interface 2014, 11, 20140852. [Google Scholar] [CrossRef]
- Yip, C.Y.; Chen, J.H.; Zhao, R.; Simmons, C.A. Calcification by valve interstitial cells is regulated by the stiffness of the extracellular matrix. Arter. Thromb. Vasc. Biol. 2009, 29, 936–942. [Google Scholar] [CrossRef] [Green Version]
- Sunyer, R.; Ritort, F.; Farre, R.; Navajas, D. Thermal activation and atp dependence of the cytoskeleton remodeling dynamics. Phys. Review. Estat. Nonlinearand Soft Matter Phys. 2009, 79, 051920. [Google Scholar] [CrossRef] [Green Version]
- Sung, D.C.; Bowen, C.J.; Vaidya, K.A.; Zhou, J.; Chapurin, N.; Recknagel, A.; Zhou, B.; Chen, J.; Kotlikoff, M.; Butcher, J.T. Cadherin-11 overexpression induces extracellular matrix remodeling and calcification in mature aortic valves. Arter. Thromb. Vasc. Biol. 2016, 36, 1627–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyndt, F.; Gueffet, J.P.; Probst, V.; Jaafar, P.; Legendre, A.; Le Bouffant, F.; Toquet, C.; Roy, E.; McGregor, L.; Lynch, S.A.; et al. Mutations in the gene encoding filamin a as a cause for familial cardiac valvular dystrophy. Circulation 2007, 115, 40–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durst, R.; Sauls, K.; Peal, D.S.; de Vlaming, A.; Toomer, K.; Leyne, M.; Salani, M.; Talkowski, M.E.; Brand, H.; Perrocheau, M.; et al. Mutations in dchs1 cause mitral valve prolapse. Nature 2015, 525, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Lam, N.T.; Muldoon, T.J.; Quinn, K.P.; Rajaram, N.; Balachandran, K. Valve interstitial cell contractile strength and metabolic state are dependent on its shape. Integr. Biol. Quant. Biosci. Nano Macro 2016, 8, 1079–1089. [Google Scholar] [CrossRef]
- Hardie, D.G.; Schaffer, B.E.; Brunet, A. Ampk: An energy-sensing pathway with multiple inputs and outputs. Trends Cell Biol. 2016, 26, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Day, E.A.; Ford, R.J.; Steinberg, G.R. Ampk as a therapeutic target for treating metabolic diseases. Trends Endocrinol. Metab. 2017, 28, 545–560. [Google Scholar] [CrossRef]
- Trajkovski, M.; Hausser, J.; Soutschek, J.; Bhat, B.; Akin, A.; Zavolan, M.; Heim, M.H.; Stoffel, M. Micrornas 103 and 107 regulate insulin sensitivity. Nature 2011, 474, 649–653. [Google Scholar] [CrossRef] [Green Version]
- Padang, R.; Bagnall, R.D.; Tsoutsman, T.; Bannon, P.G.; Semsarian, C. Comparative transcriptome profiling in human bicuspid aortic valve disease using RNA sequencing. Physiol. Genom. 2015, 47, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Guauque-Olarte, S.; Droit, A.; Tremblay-Marchand, J.; Gaudreault, N.; Kalavrouziotis, D.; Dagenais, F.; Seidman, J.G.; Body, S.C.; Pibarot, P.; Mathieu, P.; et al. RNA expression profile of calcified bicuspid, tricuspid, and normal human aortic valves by RNA sequencing. Physiol. Genom. 2016, 48, 749–761. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. Tophat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. Htseq--a python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Bourgon, R.; Gentleman, R.; Huber, W. Independent filtering increases detection power for high-throughput experiments. Proc. Natl. Acad. Sci. USA 2010, 107, 9546–9551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risso, D.; Ngai, J.; Speed, T.P.; Dudoit, S. Normalization of rna-seq data using factor analysis of control genes or samples. Nat. Biotechnol. 2014, 32, 896–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oury, C.; Maréchal, P.; Donis, N.; Hulin, A.; Hego, A.; Tridetti, J.; Nguyen, M.-L.; Dulgheru, R.; Fillet, M.; Nchimi, A.; et al. Dexfenfluramine and Pergolide Cause Heart Valve Disease via Valve Metabolic Reprogramming and Ongoing Matrix Remodeling. Int. J. Mol. Sci. 2020, 21, 4003. https://doi.org/10.3390/ijms21114003
Oury C, Maréchal P, Donis N, Hulin A, Hego A, Tridetti J, Nguyen M-L, Dulgheru R, Fillet M, Nchimi A, et al. Dexfenfluramine and Pergolide Cause Heart Valve Disease via Valve Metabolic Reprogramming and Ongoing Matrix Remodeling. International Journal of Molecular Sciences. 2020; 21(11):4003. https://doi.org/10.3390/ijms21114003
Chicago/Turabian StyleOury, Cécile, Patrick Maréchal, Nathalie Donis, Alexia Hulin, Alexandre Hego, Julien Tridetti, Mai-Linh Nguyen, Raluca Dulgheru, Marianne Fillet, Alain Nchimi, and et al. 2020. "Dexfenfluramine and Pergolide Cause Heart Valve Disease via Valve Metabolic Reprogramming and Ongoing Matrix Remodeling" International Journal of Molecular Sciences 21, no. 11: 4003. https://doi.org/10.3390/ijms21114003
APA StyleOury, C., Maréchal, P., Donis, N., Hulin, A., Hego, A., Tridetti, J., Nguyen, M. -L., Dulgheru, R., Fillet, M., Nchimi, A., & Lancellotti, P. (2020). Dexfenfluramine and Pergolide Cause Heart Valve Disease via Valve Metabolic Reprogramming and Ongoing Matrix Remodeling. International Journal of Molecular Sciences, 21(11), 4003. https://doi.org/10.3390/ijms21114003