New Determinants of Aminoglycoside Resistance and Their Association with the Class 1 Integron Gene Cassettes in Trueperella pyogenes


Abstract
:1. Introduction
2. Results
2.1. Aminoglycoside Susceptibility
2.2. Distribution of Integrons and Gene Cassettes in Studied T. pyogenes Isolates
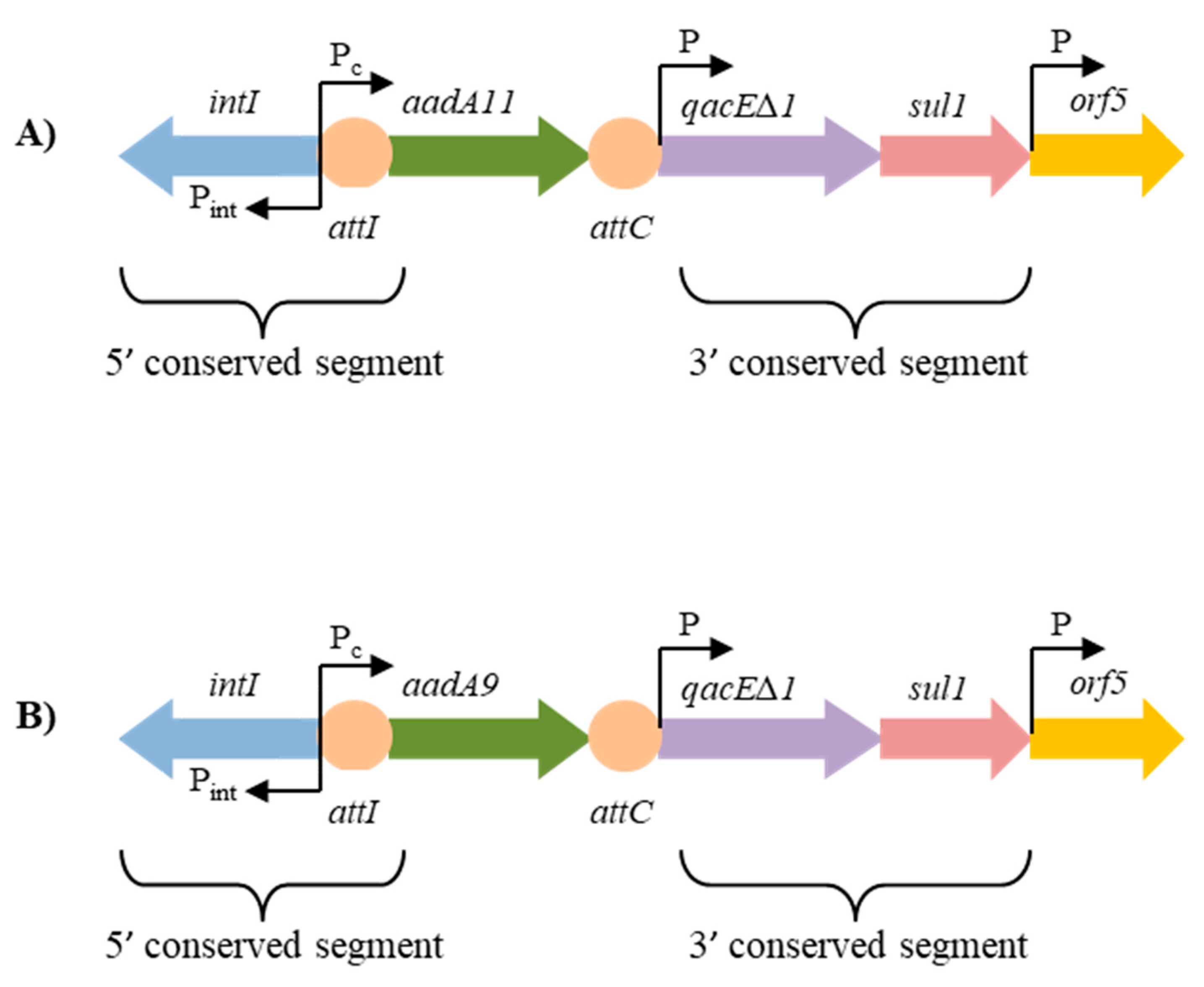
2.3. Structure of the Class 1 Integron Gene Cassettes Associated with Aminoglycoside Resistance
2.4. Prevalence of the Aminoglycoside Resistance Genes in Studied T. pyogenes Isolates
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Antimicrobial Susceptibility Testing
4.3. Detection of Integrons and Gene Cassettes
4.4. Detection of Aminoglycoside Resistance Genes
4.5. Nucleotide Sequence Accession Numbers
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jost, B.H.; Billington, S.J. Arcanobacterium pyogenes: Molecular pathogenesis of an animal opportunist. Antonie Van Leeuwenhoek 2005, 88, 87–102. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, Y.; Wang, H.; Ma, B.; Xu, L.; Wang, J.; Zhang, W. Identification of B-cell linear epitopes in domains 1–3 of pyolysin of Trueperella pyogenes using polyclonal antibodies. Vet. Microbiol. 2017, 210, 24–31. [Google Scholar] [CrossRef]
- Rzewuska, M.; Kwiecień, E.; Chrobak-Chmiel, D.; Kizerwetter-Świda, M.; Stefańska, I.; Gieryńska, M. Pathogenicity and Virulence of Trueperella pyogenes: A Review. Int. J. Mol. Sci. 2019, 20, 2737. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, H.; Kojima, A.; Ishimaru, M. Antimicrobial susceptibility of Arcanobacterium pyogenes isolated from cattle and pigs. J. Vet. Med. B Infect. Dis. Vet. Public Health 2000, 47, 139–143. [Google Scholar] [CrossRef]
- Liu, M.; Wu, C.; Liu, Y.; Zhao, J.; Yang, Y.; Shen, J. Identification, susceptibility, and detection of integron-gene cassettes of Arcanobacterium pyogenes in bovine endometritis. J. Dairy Sci. 2009, 92, 3659–3666. [Google Scholar] [CrossRef]
- Santos, T.M.; Caixeta, L.S.; Machado, V.S.; Rauf, A.K.; Gilbert, R.O.; Bicalho, R.C. Antimicrobial resistance and presence of virulence factor genes in Arcanobacterium pyogenes isolated from the uterus of postpartum dairy cows. Vet. Microbiol. 2010, 145, 84–89. [Google Scholar] [CrossRef]
- Zhao, K.; Liu, Y.; Zhang, X.; Palahati, P.; Wang, H.; Yue, B. Detection and characterization of antibiotic resistance genes in Arcanobacterium pyogenes strains from abscesses of forest musk deer. J. Med. Microbiol. 2011, 60, 1820–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risseti, R.M.; Zastempowska, E.; Twarużek, M.; Lassa, H.; Pantoja, J.C.F.; De Vargas, A.P.C.; Guerra, S.T.; Bolanõs, C.A.D.; de Paula, C.L.; Alves, A.C.; et al. Virulence markers associated with Trueperella pyogenes infections in livestock and companion animals. Lett. Appl. Microbiol. 2017, 65, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, K.; Latha, R.; Udayashankar, C.; Jayanthi, K.; Oudeacoumar, P. Three cases of Arcanobacterium pyogenes-associated soft tissue infection. J. Med. Microbiol. 2010, 59, 736–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plamondon, M.; Martinez, G.; Raynal, L.; Touchette, M.; Valiguette, L. A fatal case of Arcanobacterium pyogenes endocarditis in a man with no identified animal contact: Case report and review of the literature. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Rzewuska, M.; Czopowicz, M.; Gawryś, M.; Markowska-Daniel, I.; Bielecki, W. Relationship between antimicrobial resistance, distribution of virulence factor genes and the origin of Trueperella pyogenes isolated from domestic animals and European bison (Bison bonasus). Microb. Pathog. 2016, 96, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Billington, S.J.; Jost, B.H. Multiple genetic elements carry the tetracycline resistance gene tet(W) in the animal pathogen Arcanobacterium pyogenes. Antimicrob. Ag. Chemother. 2006, 50, 3580–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jost, B.H.; Field, A.C.; Trinh, H.T.; Songer, J.G.; Billington, S.J. Tylosin resistance in Arcanobacterium pyogenes is encoded by an Erm X determinant. Antimicrob. Ag. Chemother. 2003, 47, 3519–3524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jost, B.H.; Trinh, H.T.; Songer, J.G.; Billington, S.J. A second tylosin resistance determinant, Erm B, in Arcanobacterium pyogenes. Antimicrob. Ag. Chemother. 2004, 48, 721–727. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.L.; Kong, L.C.; Wang, Y.; Gou, C.; Xu, B.; Ma, H.; Gao, Y.H. Aminoglicoside resistance of Trueperella pyogenes isolated from pigs in China. J. Vet. Med. Sci. 2017, 79, 1836–1839. [Google Scholar] [CrossRef] [Green Version]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef]
- Barraud, O.; Ploy, M.C. Diversity of Class 1 Integron Gene Cassette Rearrangements Selected under Antibiotic Resistance Pressure. J. Bacteriol. 2015, 197, 2171–2178. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y.; Yu, G. Resistance integrons: Class 1, 2 and 3 integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- Böhm, M.-E.; Razavi, M.; Marathe, N.P.; Flach, C.-F.; Larsson, D.G.J. Discovery of a novel integron-borne aminoglycoside resistance gene present in clinical pathogens by screening environmental bacterial communities. Microbiome 2020, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Palma, E.; Tilocca, B.; Roncada, P. Antimicrobial Resistance in Veterinary Medicine: An Overview. Int. J. Mol. Sci. 2020, 21, 1914. [Google Scholar] [CrossRef] [Green Version]
- Alduina, R.; Gambino, D.; Presentato, A.; Gentile, A.; Sucato, A.; Savoca, D.; Filippello, S.; Visconti, G.; Caracappa, G.; Vicari, D.; et al. Is Caretta Caretta a Carrier of Antibiotic Resistance in the Mediterranean Sea? Antibiotics 2020, 9, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.; Wang, F.; Sun, J.; Wang, Q.; Chen, Q.; Yu, S.; Rui, Y. Class 1 integron gene cassettes in multidrug-resistant Gram-negative bacteria in southern China. Int. J. Antimicrob. Ag. 2012, 40, 264–2687. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhao, X. Characterization of the resistance class 1 integrons in Staphylococcus aureus isolates from milk of lactating dairy cattle in Northwestern China. BMC Vet. Res. 2018, 14, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, N.C.; Olsvik, O.; Swenson, J.M.; Spiegel, C.A.; Tenover, F.C. Detection of a streptomycin/spectinomycin adenylyltransferase gene (aadA) in Enterococcus faecalis. Antimicrob. Ag. Chemother. 1999, 43, 157–160. [Google Scholar] [CrossRef] [Green Version]
- Tauch, A.; Götker, S.; Pühler, A.; Kalinowski, J.; Thierbach, G. The 27.8-kb R-plasmid pTET3 from Corynebacterium glutamicum encodes the aminoglycoside adenyltransferase gene cassette aadA9 and the regulated tetracycline efflux system Tet 33 flanked by active copies of the widespread insertion sequence IS6100. Plasmid 2002, 48, 117–129. [Google Scholar] [CrossRef]
- Xu, Z.; Li, L.; Shirtliff, M.E.; Peters, B.M.; Li, B.; Peng, Y.; Alam, M.J.; Yamasaki, S.; Shi, L. Resistance class 1 integron in clinical methicillin-resistant Staphylococcus aureus strains in southern China, 2001–2006. Clin. Microbiol. Infect. 2011, 17, 714–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Update 2010, 13, 151–171. [Google Scholar] [CrossRef] [Green Version]
- Larouche, A.; Roy, P.H. Effect of attC structure on cassette excision by integron integrases. Mobile DNA 2011, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Partridge, S.R.; Recchia, G.D.; Scaramuzzi, C.; Collis, C.M.; Stokes, H.W.; Hall, R.M. Definition of the attI site of class 1 integrons. Microbiology 2000, 146, 2855–2864. [Google Scholar] [CrossRef] [Green Version]
- Anonymous. FDA Annual Summary Report on Antimicrobials Sold or Distributed in 2018 for Use in Food-Producing Animals; FDA Center for Veterinary Medicine: Rockville, MD, USA, 2019. [Google Scholar]
- Tamai, I.A.; Mohammadzadeh, A.; Salehi, T.Z.; Mahmoodi, P. Genomic characterisation, detection of genes encoding virulence factors and evaluation of antibiotic resistance of Trueperella pyogenes isolated from cattle with clinical metritis. Antonie van Leeuwenhoek 2018, 111, 2441–2453. [Google Scholar] [CrossRef]
- Pohl, A.; Lübke-Becker, A.; Heuwieser, W. Minimum inhibitory concentrations of frequently used antibiotics against Escherichia coli and Trueperella pyogenes isolated from uteri of postpartum dairy cows. J. Dairy Sci. 2018, 101, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- Brodzki, P.; Bochniarz, M.; Brodzki, A.; Wrona, Z.; Wawron, W. Trueperella pyogenes and Escherichia coli as an etiological factor of endometritis in cows and the susceptibility of these bacteria to selected antibiotics. Pol. J. Vet. Sci. 2014, 17, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, D.; Turutoglu, H.; Pehlivanoglu, F.; Guler, L. Virulence genes, biofilm production and antibiotic susceptibility in Trueperella pyogenes isolated from cattle. Isr. J. Vet. Med. 2016, 71, 36–42. [Google Scholar]
- Rezanejad, M.; Karimi, S.; Momtaz, H. Phenotypic and molecular characterization of antimicrobial resistance in Trueperella pyogenes strains isolated from bovine mastitis and metritis. BMC Microbiol. 2019, 19, 305. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.L.; Liu, L.; Odah, K.A.; Atiah, L.A.; Gao, Y.H.; Kong, L.C.; Ma, H.X. Antimicrobial resistance and presence of virulence factor genes in Trueperella pyogenes isolated from pig lungs with pneumonia. Trop. Anim. Health Prod. 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Galán-Relaño, Á.; Gómez-Gascón, L.; Luque, I.; Barrero-Domínguez, B.; Casamayor, A.; Cardoso-Toset, F.; Vela, A.I.; Fernández-Garayzábal, J.F.; Tarradas, C. Antimicrobial susceptibility and genetic characterization of Trueperella pyogenes isolates from pigs reared under intensive and extensive farming practices. Vet. Microbiol. 2019, 232, 89–95. [Google Scholar] [CrossRef]
- Moreno, L.Z.; Matajira, C.E.C.; da Costa, B.L.P.; Ferreira, T.S.P.; Silva, G.F.R.; Dutra, M.C.; Gomes, V.T.M.; Silva, A.P.S.; Christ, A.P.G.; Sato, M.I.Z.; et al. Characterization of porcine Trueperella pyogenes by matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS), molecular typing and antimicrobial susceptibility profiling in Sao Paulo State. Comp. Immunol. Microbiol. Infect. Dis. 2017, 51, 49–53. [Google Scholar] [CrossRef]
- Tell, L.A.; Brooks, J.W.; Lintner, V.; Matthews, T.; Kariyawasam, S. Antimicrobial susceptibility of Arcanobacterium pyogenes isolated from the lungs of white-tailed deer (Odocoileus virginianus) with pneumonia. J. Vet. Diagn. Investig. 2011, 23, 1009–1013. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Methods for Antimicrobial Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria Isolated from Animals, 1st ed.; CLSI Supplement VET06; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Byrne-Bailey, K.G.; Gaze, W.H.; Zhang, L.; Kay, P.; Boxall, A.; Hawkey, P.M.; Wellington, E.M.H. Integron Prevalence and Diversity in Manured Soil. Appl. Environ. Microbiol. 2011, 77, 684–687. [Google Scholar] [CrossRef] [Green Version]
- Llanes, C.; Neuwirth, C.; El Garch, F.; Hocquet, D.; Plésiat, P. Genetic analysis of a multiresistant strain of Pseudomonas aeruginosa producing PER-1 β-lactamase. Clin. Microbiol. Infect. 2006, 12, 270–278. [Google Scholar] [CrossRef]
- Agersø, Y.; Sandvang, D. Class 1 Integrons and Tetracycline Resistance Genes in Alcaligenes, Arthrobacter, and Pseudomonas spp. Isolated from Pigsties and Manured Soil. Appl. Environ. Microbiol. 2005, 71, 7941–7947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinué, L.; Jové, T.; Torres, C.; Ploy, M.C. Diversity of class 1 integron gene cassette Pc promoter variants in clinical Escherichia coli strains and description of a new P2 promoter variant. Int. J. Antimicrob. Agents. 2011, 38, 526–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiou, C.S.; Jones, A.L. Expression and Identification of the strA-strB Gene Pair from Streptomycin-Resistant Erwinia Amylovora. Gene 1995, 152, 47–51. [Google Scholar] [CrossRef]
- Sunde, M.; Norström, M. The genetic background for streptomycin resistance in Escherichia coli influences the distribution of MICs. J. Antimicrob. Chemother. 2005, 56, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Pyatov, V.; Vrtková, I.; Knoll, A. Detection of selected antibiotic resistance genes using multiplex PCR assay in mastitis pathogens in the Czech Republic. Acta Vet. Brno 2017, 86, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Song, X.; Ma, X.; Zhang, S.; Zhang, J. Molecular characterization of multidrug-resistant Klebsiella pneumoniae isolates. Braz. J. Microbiol. 2015, 46, 759–768. [Google Scholar] [CrossRef] [Green Version]
- L’Abée-Lund, T.M.; Sørum, H. Functional Tn5393-like transposon in the R plasmid pRAS2 from the fish pathogen Aeromonas salmonicida subspecies salmonicida isolated in Norway. Appl. Environ. Microbiol. 2000, 66, 5533–5535. [Google Scholar] [CrossRef] [Green Version]
- Dyall-Smith, M.L.; Liu, Y.; Billman-Jacobe, H. Genome Sequence of an Australian Monophasic Salmonella enterica subsp. enterica Typhimurium Isolate (TW-Stm6) Carrying a Large Plasmid with Multiple Antimicrobial Resistance Genes. Genome Announc. 2017, 5, e00793-17. [Google Scholar] [CrossRef] [Green Version]
- Pezzella, C.; Ricci, A.; DiGiannatale, E.; Luzzi, I.; Carattoli, A. Tetracycline and Streptomycin Resistance Genes, Transposons, and Plasmids in Salmonella enterica Isolates from Animals in Italy. Antimicrob. Ag. Chemother. 2004, 48, 903–908. [Google Scholar] [CrossRef] [Green Version]
- Navas, J.; Fernández-Martínez, M.; Salas, C.; Cano, M.E.; Martínez-Martínez, L. Susceptibility to Aminoglycosides and Distribution of aph and aac(3)-XI Genes among Corynebacterium striatum Clinical Isolates. PLoS ONE 2016, 11, e0167856. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, L.; Kromann, S.; Chen, M.; Shi, L.; Meng, H. Antibiotic Resistance of Lactobacillus spp. and Streptococcus thermophilus Isolated from Chinese Fermented Milk Products. Foodborne Pathog. Dis. 2019, 16, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Woegerbauer, M.; Zeinzinger, J.; Springer, B.; Hufnagl, P.; Indra, A.; Korschineck, I.; Hofrichter, J.; Kopacka, I.; Fuchs, R.; Steinwider, J.; et al. Prevalence of the aminoglycoside phosphotransferase genes aph(3’)-IIIa and aph(3’)-IIa in Escherichia coli, Enterococcus faecalis, Enterococcus faecium, Pseudomonas aeruginosa, Salmonella enterica subsp. enterica and Staphylococcus aureus isolates in Austria. J. Med. Microbiol. 2014, 63, 210–217. [Google Scholar] [PubMed] [Green Version]
- Lapierre, L.; Cornejo, J.; Borie, C.; Toro, C.; San Martín, B. Genetic characterization of antibiotic resistance genes linked to class 1 and class 2 integrons in commensal strains of Escherichia coli isolated from poultry and swine. Microb. Drug. Resist. 2008, 14, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thu, W.P.; Sinwat, N.; Bitrus, A.A.; Angkittitrakul, S.; Prathan, R.; Chuanchuen, R. Prevalence, antimicrobial resistance, virulence gene, and class 1 integrons of Enterococcus faecium and Enterococcus faecalis from pigs, pork and humans in Thai-Laos border provinces. J. Glob. Antimicrob. Resist. 2019, 18, 130–138. [Google Scholar] [CrossRef]
- Rzewuska, M.; Stefańska, I.; Osińska, B.; Kizerwetter-Świda, M.; Chrobak, D.; Kaba, J.; Bielecki, W. Phenotypic characteristics and virulence genotypes of Trueperella (Arcanobacterium) pyogenes strains isolated from European bison (Bison bonasus). Vet. Microbiol. 2012, 160, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vakulenko, S.B.; Donabedian, S.M.; Voskresenskiy, A.M.; Zervos, M.J.; Lerner, S.A.; Chow, J.W. Multiplex PCR for Detection of Aminoglycoside Resistance Genes in Enterococci. Antimicrob. Ag. Chemother. 2003, 47, 1423–1426. [Google Scholar] [CrossRef] [Green Version]
- Ouoba, L.I.; Lei, V.; Jensen, L.B. Resistance of potential probiotic lactic acid bacteria and bifidobacteria of African and European origin to antimicrobials: Determination and transferability of the resistance genes to other bacteria. Int. J. Food Microbiol. 2008, 121, 217–224. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antibiotic | Number of Isolates with MIC (µg/mL)* | MIC50 | MIC90 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ≤0.125 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | ≥128 | |||
| Gentamicin | 9 | 12 | 35 | 18 | 12 | │ | 0.5 | 2 | |||||
| Streptomycin | 41 | 17│ | 8 | 4 | 4 | 9 | 3 | 4 | 64 | ||||
| Kanamycin | 2 | 6 | 12 | 23 | 20 | 13│ | 5 | 2 | 3 | 1 | 8 | ||
| Antibiotic | Total (n = 86) | Resistant Isolates From [% (n)] | |||||
|---|---|---|---|---|---|---|---|
| Cattle (n = 26) | Pigs (n = 21) | European bison (n = 25) | Goats (n = 8) | Sheep (n = 5) | Antelope (n = 1) | ||
| Gentamicin | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Streptomycin | 32.6 (28) | 53.8 (14) | 57.1 (12) | 0 (0) | 12.5 (1) | 20.0 (1) | 0 (0) |
| Kanamycin | 11.6 (10) | 11.5 (3) | 23.8 (5) | 8,0 (2) | 0 (0) | 0 (0) | 0 (0) |
| Resistance Gene | Total (n = 86) | Isolates From [% (n)] | |||||
|---|---|---|---|---|---|---|---|
| Cattle (n = 26) | Pigs (n = 21) | European Bison (n = 25) | Goats (n = 8) | Sheep (n = 5) | Antelope (n = 1) | ||
| aadA1 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| aadA9 | 9.3 (8) | 27.0 (7) | 0 (0) | 0 (0) | 12.5 (1) | 0 (0) | 0 (0) |
| aadA11 | 8.1 (7) | 0 (0) | 33.3 (7) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| aacC | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| strA-strB | 1.2 (1) | 3.8 (1) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| aph(3’)-IIIa | 5.8 (5) | 0 (0) | 19.0 (4) | 0 (0) | 12.5 (1) | 0 (0) | 0 (0) |
| aac(6’)-aph(2’’) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Amplification Target | Primers Sequence (5’-3’) | Annealing Temperature (°C) | PCR Product size (bp) | Reference |
|---|---|---|---|---|
| Class 1 intI | F: CCTCCCGCACGATGATC | 57 | 280 | [7] |
| R: TCCACGCATCGTCAGGC | ||||
| Class 2 intI | F: TTATTGCTGGGATTAGGC | 50 | 233 | [7] |
| R: ACGGCTACCCTCTGTTATC | ||||
| Class 1 gene cassette | F: GGCATCCAAGCAGCAAG | 58 | unpredictable | [15] |
| R: AAGCAGACTTGACCTGA | ||||
| aadA1 | F: CGGTGACCGTAAGGCTTGAT | 52 | 193 | This study |
| R: ATGTCATTGCGCTGCCATTC | ||||
| aadA9 | F: ACGCCGACCTTGCAATTCT | 52 | 373 | This study |
| R: TAGCCAATGAACGCCGAAGT | ||||
| aadA11 | F: CGTGCATTTGTACGGCTCTG | 53 | 352 | This study |
| R: ACCTGCCAATGCAAGGCTAT | ||||
| aacC | F: TTGCTGCCTTCGACCAAGAA | 53 | 256 | This study |
| R: TCCCGTATGCCCAACTTTGT | ||||
| strA-strB | F: TATCTGCGATTGGACCCTCTG | 60 | 538 | [46] |
| R: CATTGCTCATCATTTGATCGGCT | ||||
| aph(3’)-IIIa | F: GGCTAAAATGAGAATATCACCGG | 55 | 523 | [59] |
| R: CTTTAAAAAATCATACAGCTCGCG | ||||
| aac(6’)-aph(2’’) | F: CCAAGAGCAATAAGGGCATA | 48 | 220 | [60] |
| R: CACTATCATAACCACTACCG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwiecień, E.; Stefańska, I.; Chrobak-Chmiel, D.; Sałamaszyńska-Guz, A.; Rzewuska, M. New Determinants of Aminoglycoside Resistance and Their Association with the Class 1 Integron Gene Cassettes in Trueperella pyogenes. Int. J. Mol. Sci. 2020, 21, 4230. https://doi.org/10.3390/ijms21124230
Kwiecień E, Stefańska I, Chrobak-Chmiel D, Sałamaszyńska-Guz A, Rzewuska M. New Determinants of Aminoglycoside Resistance and Their Association with the Class 1 Integron Gene Cassettes in Trueperella pyogenes. International Journal of Molecular Sciences. 2020; 21(12):4230. https://doi.org/10.3390/ijms21124230
Chicago/Turabian StyleKwiecień, Ewelina, Ilona Stefańska, Dorota Chrobak-Chmiel, Agnieszka Sałamaszyńska-Guz, and Magdalena Rzewuska. 2020. "New Determinants of Aminoglycoside Resistance and Their Association with the Class 1 Integron Gene Cassettes in Trueperella pyogenes" International Journal of Molecular Sciences 21, no. 12: 4230. https://doi.org/10.3390/ijms21124230
APA StyleKwiecień, E., Stefańska, I., Chrobak-Chmiel, D., Sałamaszyńska-Guz, A., & Rzewuska, M. (2020). New Determinants of Aminoglycoside Resistance and Their Association with the Class 1 Integron Gene Cassettes in Trueperella pyogenes. International Journal of Molecular Sciences, 21(12), 4230. https://doi.org/10.3390/ijms21124230