Vernonia calvoana Shows Promise towards the Treatment of Ovarian Cancer

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
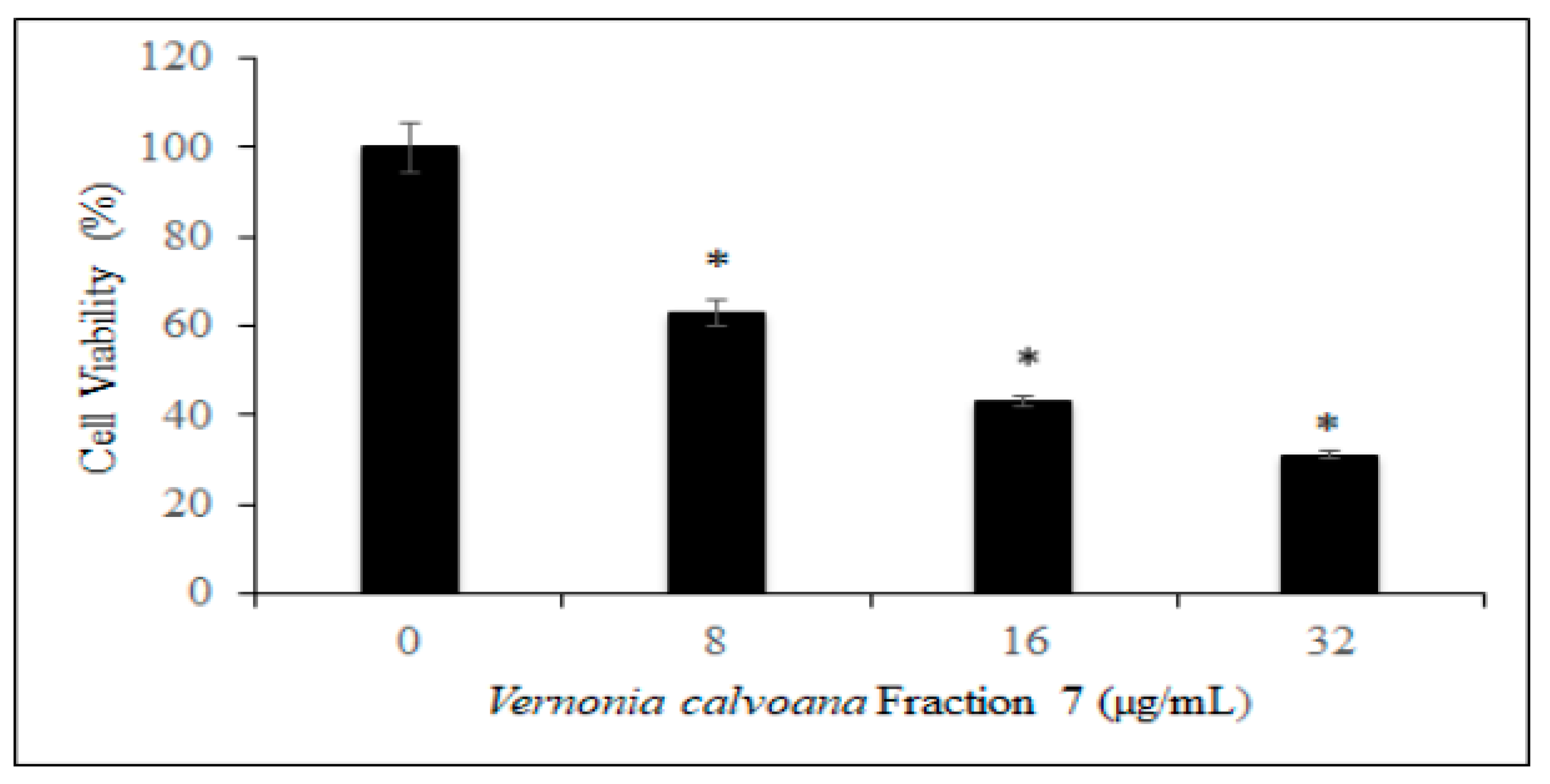
2.1. Antiproliferative Effect
2.2. Morphological Changes
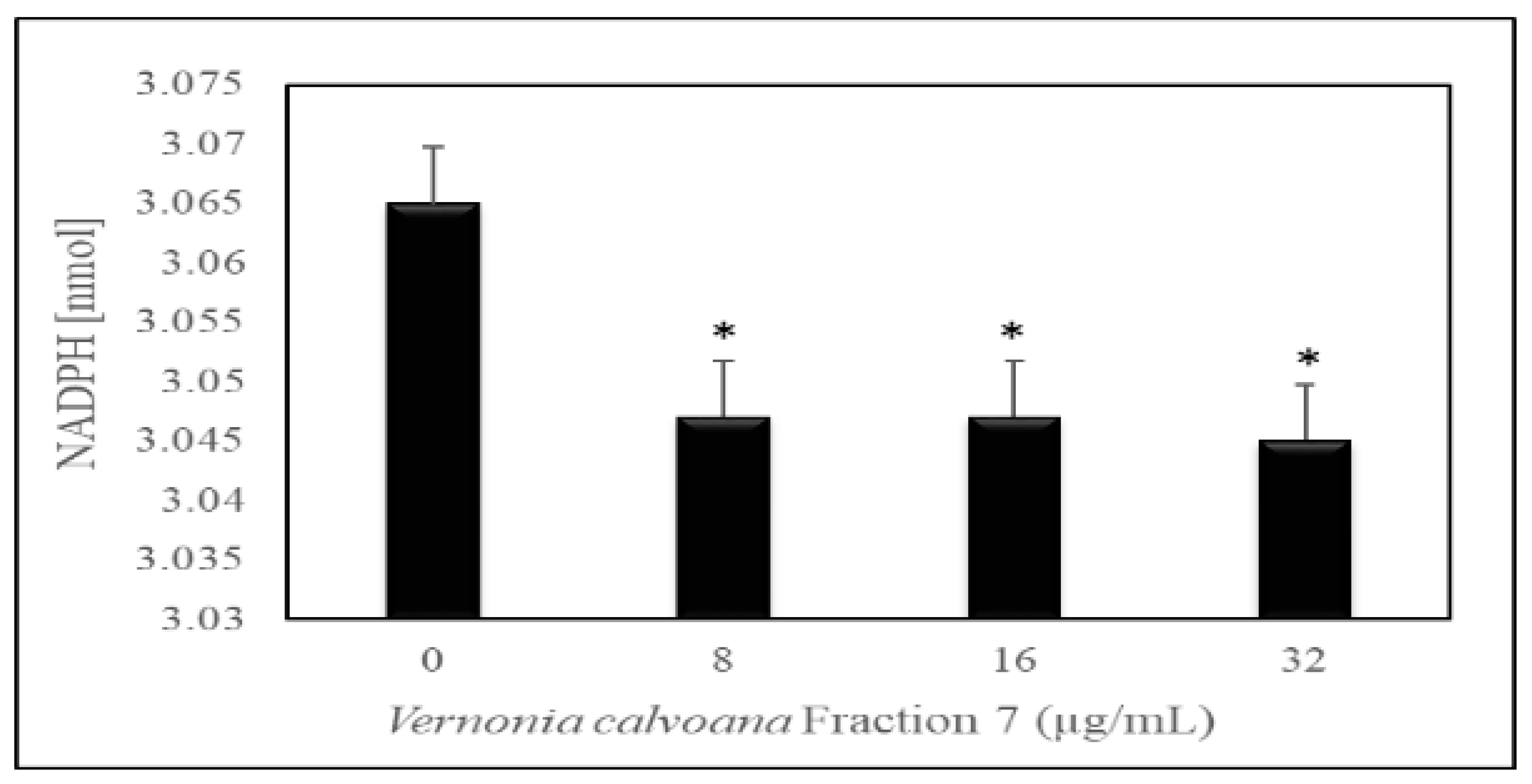
2.3. Induction of Oxidative Stress
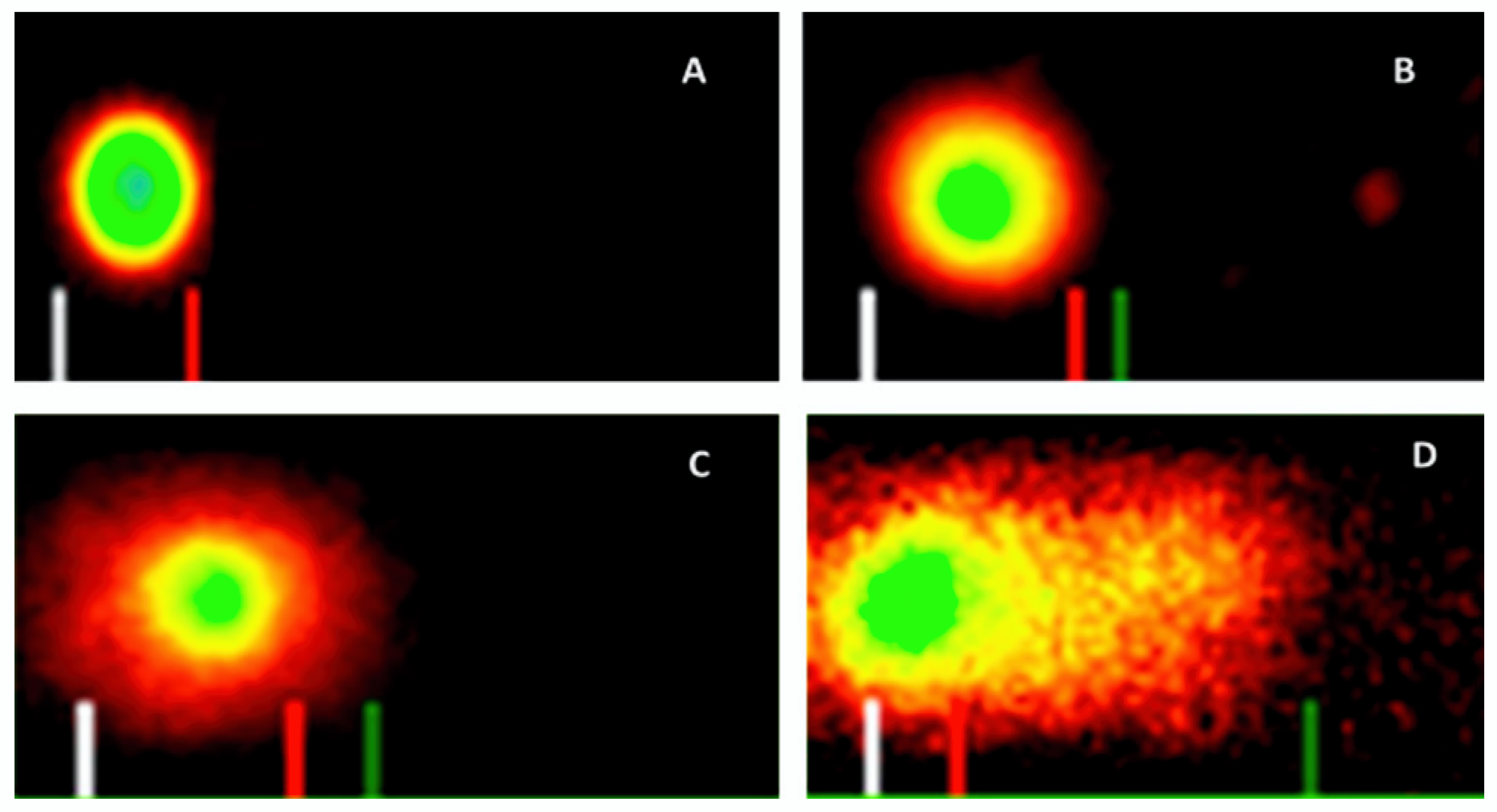
2.4. Induction of DNA Damage
2.5. Induction of Cell Cycle Arrest
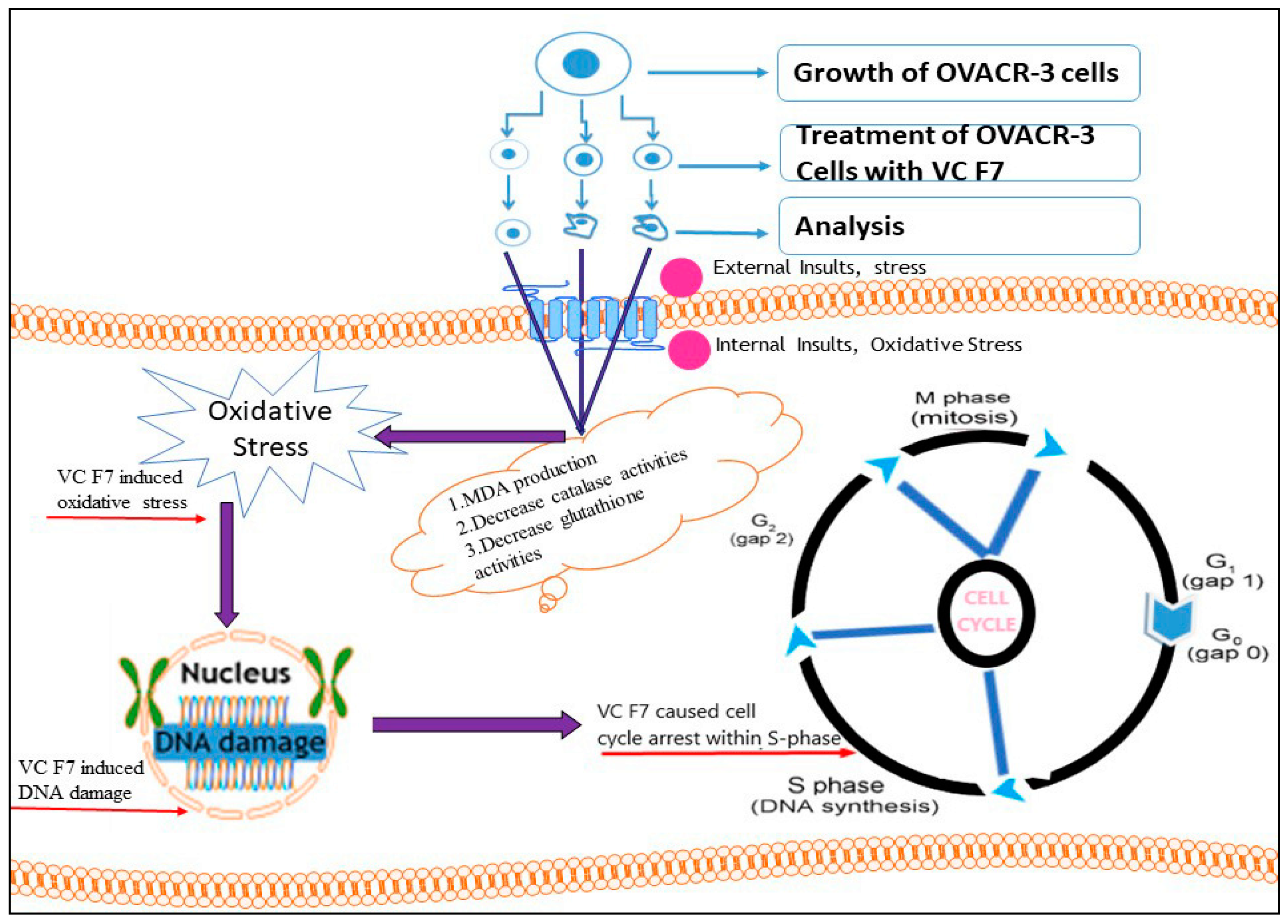
3. Discussion
3.1. Antiproliferative Effect
3.2. Induction of Oxidative Stress
3.3. Induction of DNA Damage
3.4. Induction of Cell Cycle Arrest
4. Materials and Methods
4.1. Chemicals and Media
4.2. Vernonia calvoana Preparation and Fractionation
4.3. Cell Culture
4.4. Cell Treatment and Determination of Cell Viability
4.5. Morphological Changes
4.6. Measurement of Lipid Peroxidation/Malondiadehyde
4.7. Measurement of Catalase Activity
4.8. Measurement of Glutathione Peroxidase Activity
4.9. Assessment of DNA Damage
4.10. Assessment of Cell Cycle Distribution
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA. Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Cancer Society. Breast Cancer Facts & Figures 2017–2018; American Cancer Society, Inc.: Atlanta, GA, USA, 2017. [Google Scholar]
- Jones, M.R.; Kamara, D.; Karlan, B.Y.; Pharoah, P.D.P.; Gayther, S.A. Genetic epidemiology of ovarian cancer and prospects for polygenic risk prediction. Gynecol. Oncol. 2017, 147, 705–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Committee on Practice Bulletins–Gynecology, Committee on Genetics, Society of Gynecologic Oncology. Practice Bulletin No 182: Hereditary Breast and Ovarian Cancer Syndrome. Obstet. Gynecol. 2017, 130, e110–e126. [Google Scholar] [CrossRef] [PubMed]
- Friebel, T.M.; Domchek, S.M.; Rebbeck, T.R. Modifiers of cancer risk in BRCA1 and BRCA2 mutation carriers: Systematic review and meta-analysis. J. Natl. Cancer Inst. 2014, 106, 235. [Google Scholar] [CrossRef] [Green Version]
- Moorman, P.G.; Havrilesky, L.J.; Gierisch, J.; Coeytaux, R.R.; Lowery, W.J.; Urrutia, R.P.; Dinan, M.; McBroom, A.J.; Hasselblad, V.; Sanders, G.D.; et al. Oral contraceptives and risk of ovarian cancer and breast cancer among high-risk women: A systematic review and meta-analysis. J. Clin. Oncol. 2013, 31, 4188–4198. [Google Scholar] [CrossRef]
- Cibula, D.; Zikan, M.; Dusek, L.; Majek, O. Oral contraceptives and risk of ovarian and breast cancers in BRCA mutation carriers: A meta-analysis. Expert Rev. Anticancer Ther. 2011, 11, 1197–1207. [Google Scholar] [CrossRef]
- Miller, K.D.; Havrilesky, L.J.; Gierisch, J.M.; Coeytaux, R.R.; Lowery, W.J.; Dinan, R.P.U.M.; McBroom, A.J.; Hasselblad, V.; Sanders, G.D.; Myers, E.R. Cancer treatment and survivorship statistics, 2016. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [Green Version]
- Jasen, P. From the ‘silent killer’ to the ‘whispering disease’: Ovarian cancer and the uses of metaphor. Med. Hist. 2009, 53, 489–512. [Google Scholar] [CrossRef] [Green Version]
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef]
- Howlader, N.; Noone, A.M.; Krapcho, M.; Miller, D.; Bishop, K.; Kosary, C.L.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; et al. (Eds.) SEER Cancer Statistics Review, 1975–2014; National Cancer Institute: Bethesda, MD, USA, 2017. Available online: http://Seer.Cancer.Gov./Csr/1975_2014/ (accessed on 1 April 2017).
- Matsuo, K.; Lin, Y.G.; Roman, L.D.; Sood, A.K. Overcoming platinum resistance in ovarian carcinoma. Expert Opin. Investig. Drugs 2010, 19, 1339–1354. [Google Scholar] [CrossRef]
- Kim, M.; Suh, D.H.; Lee, K.-H.; Eom, K.-Y.; Toftdahl, N.G.; Mirza, M.R.; Kim, J.-W. Major clinical research advances in gynecologic cancer in 2018. J. Gynecol. Oncol. 2019, 30, e18. [Google Scholar] [CrossRef]
- Wright, A.A.; Bohlke, K.; Armstrong, D.K.; Bookman, M.A.; Cliby, W.A.; Coleman, R.L.; Dizon, D.S.; Kash, J.J.; Meyer, L.A.; Moore, K.N.; et al. Neoadjuvant chemotherapy for newly diagnosed, advanced ovarian cancer: Society of Gynecologic Oncology and American Society of Clinical Oncology Clinical Practice Guideline. Gynecol. Oncol. 2016, 143, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ejoh, R.A.; Nkonga, D.V.; Inocent, G.; Moses, M.C. Nutritional components of some non-conventional leafy vegetables consumed in Cameroon. Pak. J. Nutr. 2007, 6, 712–717. [Google Scholar] [CrossRef] [Green Version]
- Igile, G.O.; Iwara, I.A.; Mgbeje, B.I.A.; Uboh, F.E.; Ebong, P.E. Phytochemical, proximate and nutrient composition of Vernonia calvaona Hook (Asterecea): A green-leafy vegetable in Nigeria. J. Food Res. 2013, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Neurol. 2014, 4, 177. [Google Scholar] [CrossRef] [Green Version]
- Toyang, N.J.; Verpoorte, R. A review of the medicinal potentials of plants of the genus Vernonia (Asteraceae). J. Ethnopharmacol. 2013, 146, 681–723. [Google Scholar] [CrossRef]
- Egbung, G.E.; Atangwho, I.J.; Kiasira, Z.B.; Iwara, I.A.; Igile, G.O. Antioxidant activity of the inflorescents of Vernonia calvoana growing in Yakurr Local Government Area of Cross River State, Nigeria. Glob. J. Pure Appl. Sci. 2016, 22, 2. [Google Scholar] [CrossRef] [Green Version]
- Iwara, I.; Igile, G.; Uboh, F.; Eyong, E.; Ebong, P. Hypoglycemic and Hypolipidemic Potentials of Extract of Vernonia calvoana on Alloxan-Induced Diabetic Albino Wistar Rats. Eur. J. Med. Plants 2015, 8, 78–86. [Google Scholar] [CrossRef]
- Yedjou, C.; Izevbigie, E.; Tchounwou, P. Preclinical assessment of Vernonia amygdalina leaf extracts as DNA damaging anti-cancer agent in the management of breast cancer. Int. J. Environ. Res. Public Health 2008, 5, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Yedjou, C.G.; Izevbigie, E.B.; Tchounwou, P.B. Induced Growth Arrest and Apoptosis of Breast Cancer (MCF-7) Cells. Pharmacol. Pharm. 2013, 4, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Lowe, H.; Daley-Beckford, D.; Toyang, N.; Watson, C.; Hartley, S.; Bryant, J. The anti-cancer activity of Vernonia divaricata Sw against leukaemia, breast and prostate cancers in vitro. West. Indian Med. J. 2014, 63, 285–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, W.; Tchounwou, P.B.; Yedjou, C.G. Therapeutic mechanisms of vernonia amygdalina delile in the treatment of prostate cancer. Molecules 2017, 22, 1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yedjou, C.G.; Tchounwou, S.S.; Williams, K.; Tchounwou, P.B. Novel Cellular Staining Protocol and Antiproliferative effect of vernonia amygdalina delile on lung and prostate cancer cells. Int. J. Eng. Sci Res. Technol. 2018, 7, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Yedjou, C.G.; Tchounwou, P.B.; Miele, L.; Ogungbe, I.V.; Brown, R.J.; Payton, M. Abstract 5313: The anticancer effect of Vernonia amygdalina in the MMTV-PyVT transgenic mouse model. Exp. Mol. Ther. 2015, 75, 5313. [Google Scholar] [CrossRef]
- Tugume, P.; Kakudidi, E.K.; Buyinza, M.; Namaalwa, J.; Kamatenesi, M.; Mucunguzi, P.; Kalema, J. Ethnobotanical survey of medicinal plant species used by communities around Mabira Central Forest Reserve, Uganda. J. Ethnobiol. Ethnomed. 2016, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- Adedapo, A.A.; Adeoye, B.O.; Sofidiya, M.O.; Oyagbemi, A.A. Antioxidant, antinociceptive and anti-inflammatory properties of the aqueous and ethanolic leaf extracts of Andrographis paniculata in some laboratory animals. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 327–334. [Google Scholar] [CrossRef]
- Kankara, S.S.; Ibrahim, M.H.; Mustafa, M.; Go, R. Ethnobotanical survey of medicinal plants used for traditional maternal healthcare in Katsina state, Nigeria. South Afr. J. Bot. 2015, 97, 165–175. [Google Scholar] [CrossRef]
- Egbung, G.E.; Odey, O.D.; Atangwho, I.J. Effect of Vernonia calvoana Extract on Selected Serum Kidney Function Biomarkers of Acetaminophen Treated Wistar Rats. Asian J. Biochem. 2017, 12, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Asante, D.-B.; Effah-Yeboah, E.; Barnes, P.; Abban, H.A.; Ameyaw, E.O.; Boampong, J.; Ofori, E.G.; Dadzie, J.B. Corrigendum to ‘Antidiabetic Effect of Young and Old Ethanolic Leaf Extracts of Vernonia amygdalina: A Comparative Study’. J. Diabetes Res. 2017, 2017, 5618548. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Pradheep, K.; Sharma, N. Potential introduced medicinal plant african bitter leaf (Vernonia amygdalina delile) in India: Botany, propagation and uses. Med. Plants 2014, 6, 272. [Google Scholar] [CrossRef]
- Bodeker, G. WHO global atlas on traditional, complementary and alternative medicine (TCAM). Iran. J. Pharm. Res. 2010, 3, 25. [Google Scholar]
- Barnes, P.M.; Bloom, B.; Nahin, R.L. Complementary and Alternative Medicine use among Adults and Children: United States, 2007. Natl. Health Stat. Rep. 2010, 10, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Medicine, A. The Use of Complementary and Alternative Medicine in the United States: Cost Data. Integr. Med. 2008. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwjM7MWT7JTqAhVB_KQKHVwGDa0QFjAAegQIBBAB&url=http%3A%2F%2Fwww.orientalmedicine.edu%2Fdownloads%2FNHIS_costdata.pdf&usg=AOvVaw0dHslsA9VjzuftKRtrOBL_ (accessed on 1 July 2009).
- Sun, X.D.; Liu, X.E.; Huang, D.S. Curcumin induces apoptosis of triple-negative breast cancer cells by inhibition of EGFR expression. Mol. Med. Rep. 2012, 6, 1267–1270. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Lee, Y.-H.; Sharma, A.R.; Park, J.-B.; Jagga, S.; Sharma, G.; Lee, S.-S.; Nam, J.-S. Quercetin induces apoptosis and cell cycle arrest in triple-negative breast cancer cells through modulation of Foxo3a activity. Korean J. Physiol. Pharmacol. 2017, 21, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Mileo, A.M.; Miccadei, S. Polyphenols as Modulator of Oxidative Stress in Cancer Disease: New Therapeutic Strategies. Oxid. Med. Cell. Longev. 2016, 2016, 17. [Google Scholar] [CrossRef] [Green Version]
- Yedjou, C.G.; Tchounwou, P.B. In vitro assessment of oxidative stress and apoptotic mechanisms of garlic extract in the treatment of acute promyelocytic leukemia. J. Cancer Sci. Ther. 2012, 2012, 6. [Google Scholar] [CrossRef]
- Podsedek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT-Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Mei, F.; You, J.; Liu, B.; Zhang, M.; Liu, J.; Zhang, B.; Pei, F. LASS2/TMSG1 inhibits growth and invasion of breast cancer cell in vitro through regulation of vacuolar ATPase activity. Tumor Biol. 2015, 36, 2831–2844. [Google Scholar] [CrossRef]
- Zhu, M.; Chang, Q.; Wong, L.K.; Chong, F.S.; Li, R.C. Triterpene antioxidants from Ganoderma lucidum. Phyther. Res. 1999, 13, 529–531. [Google Scholar] [CrossRef]
- Forcados, G.E.; Chinyere, C.N.; Shu, M.L. Acalypha wilkesiana: Therapeutic and Toxic Potential. J. Med. Surg. Pathol. 2016, 1, 1000122. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Asp. Med. 2009, 30, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesen, C.; Kiess, Y.; Debatin, K.M. A critical role of glutathione in determining apoptosis sensitivity and resistance in leukemia cells. Cell Death Differ. 2004, 11, S73–S85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheweita, S.A.; Mashaly, S.; Newairy, A.A.; Abdou, H.M.; Eweda, S.M. Changes in oxidative stress and antioxidant enzyme activities in streptozotocin-induced diabetes mellitus in rats: Role of alhagi maurorum extracts. Oxid. Med. Cell. Longev. 2016, 2016, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Zhang, J.; Mizuno, T.; Zhuang, C.; Ito, H.; Mayuzumi, H.; Okamoto, H.; Li, J. Antitumor Active Polysaccharides from the Chinese Mushroom Songshan Lingzhi, the Fruiting Body of Ganoderma Tsugae. Biosci. Biotechnol. Biochem. 1993, 57, 894–900. [Google Scholar] [CrossRef]
- Gao, L.-L.; Feng, L.; Yao, S.-T.; Jiao, P.; Qin, S.; Zhang, W.; Zhang, Y.-B.; Li, F.-R. Molecular mechanisms of celery seed extract induced apoptosis via S phase cell cycle arrest in the BGC-823 human stomach cancer cell line. Asian Pac. J. Cancer Prev. 2011, 12, 2601–2606. [Google Scholar]
- Watts, M.J.; Galpin, I.J.; Collin, H.A. The Effect of Growth Regulators, Light and Temperature on Flavour Production in Celery Tissue Cultures. New Phytol. 1984, 98, 583–591. [Google Scholar] [CrossRef]
- Bunpo, P.; Kataoka, K.; Arimochi, H.; Nakayama, H.; Kuwahara, T.; Bando, Y.; Izumi, K.; Vinitketkumnuen, U.; Ohnishi, Y. Inhibitory effects of Centella asiatica on azoxymethane-induced aberrant crypt focus formation and carcinogenesis in the intestines of F344 rats. Food Chem. Toxicol. 2004, 42, 1987–1997. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, J.; Zhao, C.; Zhao, L.; Feng, B. Antiproliferative, cell-cycle dysregulation effects of novel asiatic acid derivatives on human non-small cell lung cancer cells. Chem. Pharm. Bull. 2013, 61, 1015–1023. [Google Scholar] [CrossRef] [Green Version]
- Sa, G.; Das, T. Anti cancer effects of curcumin: Cycle of life and death. Cell Div. 2008, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-W.; Lee, J.-Y.; Huang, J.-Y.; Wang, C.-C.; Chen, W.J.; Su, S.-F.; Huang, C.-W.; Ho, C.-C.; Chen, J.J.; Tsai, M.-F.; et al. Curcumin inhibits lung cancer cell invasion and metastasis through the tumor suppressor HLJ1. Cancer Res. 2008, 68, 7428–7438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogungbe, I.V.; Erwin, W.R.; Setzer, W.N. Antileishmanial phytochemical phenolics: Molecular docking to potential protein targets. J. Mol. Graph. Model. 2014, 48, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Abugri, D.A.; Witola, W.H.; Jaynes, J.M.; Toufic, N. In vitro activity of Sorghum bicolor extracts, 3-deoxyanthocyanidins, against Toxoplasma gondii. Exp. Parasitol. 2016, 164, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Janero, D.R. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med. 1990, 9, 515–540. [Google Scholar] [CrossRef]
- Johansson, L.H.; Håkan Borg, L.A. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem. 1988, 174, 331–336. [Google Scholar] [CrossRef]
- Forstrom, J.W.; Zakowski, J.J.; Tappel, A.L. Identification of the Catalytic Site of Rat Liver Glutathione Peroxidase as Selenocysteine†. Biochemistry 1978, 17, 2639–2644. [Google Scholar] [CrossRef]
- Surh, Y.J. Anti-tumor promoting potential of selected spice ingredients with antioxidative and anti-inflammatory activities: A short review. Food Chem. Toxicol. 2002, 40, 1091–1097. [Google Scholar] [CrossRef]
- Sznarkowska, A.; Kostecka, A.; Meller, K.; Bielawski, K.P. Inhibition of cancer antioxidant defense by natural compounds. Oncotarget 2017, 8, 15996–16016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willcox, J.K.; Ash, S.L.; Catignani, G.L. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef] [PubMed]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mbemi, A.T.; Sims, J.N.; Yedjou, C.G.; Noubissi, F.K.; Gomez, C.R.; Tchounwou, P.B. Vernonia calvoana Shows Promise towards the Treatment of Ovarian Cancer. Int. J. Mol. Sci. 2020, 21, 4429. https://doi.org/10.3390/ijms21124429
Mbemi AT, Sims JN, Yedjou CG, Noubissi FK, Gomez CR, Tchounwou PB. Vernonia calvoana Shows Promise towards the Treatment of Ovarian Cancer. International Journal of Molecular Sciences. 2020; 21(12):4429. https://doi.org/10.3390/ijms21124429
Chicago/Turabian StyleMbemi, Ariane T., Jennifer N. Sims, Clement G. Yedjou, Felicite K. Noubissi, Christian R. Gomez, and Paul B. Tchounwou. 2020. "Vernonia calvoana Shows Promise towards the Treatment of Ovarian Cancer" International Journal of Molecular Sciences 21, no. 12: 4429. https://doi.org/10.3390/ijms21124429
APA StyleMbemi, A. T., Sims, J. N., Yedjou, C. G., Noubissi, F. K., Gomez, C. R., & Tchounwou, P. B. (2020). Vernonia calvoana Shows Promise towards the Treatment of Ovarian Cancer. International Journal of Molecular Sciences, 21(12), 4429. https://doi.org/10.3390/ijms21124429