Mediterranean Diet: Lipids, Inflammation, and Malaria Infection

 ,
,  and
and
Abstract
:1. Introduction
2. Malaria and Inflammation
2.1. Malaria Cycle, Vaccine, and Treatment
2.2. Malaria Physiopathology
2.3. The Role of Monocytes in Malaria
2.4. The Role of T Cells in Malaria
2.5. Cerebral Malaria and Inflammation
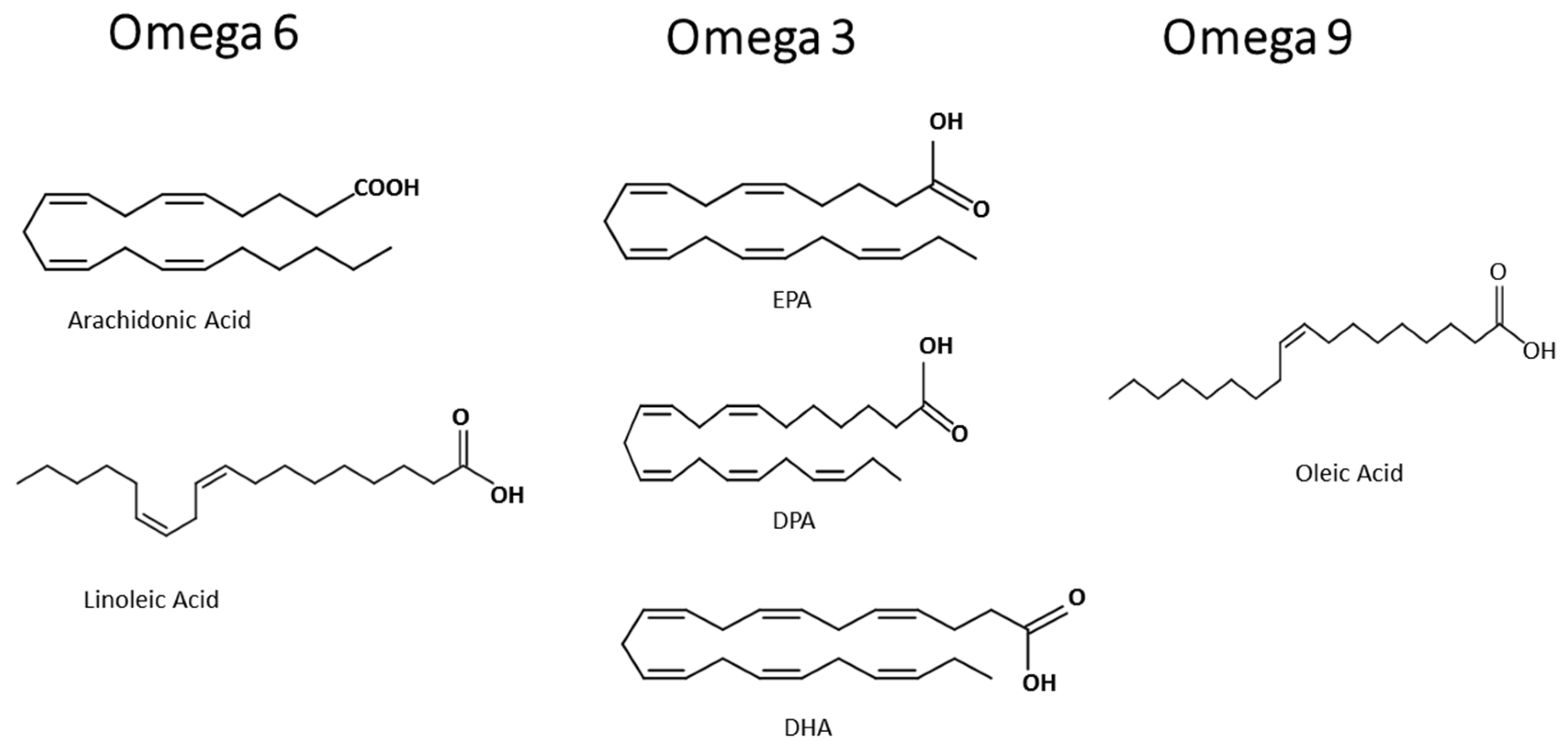
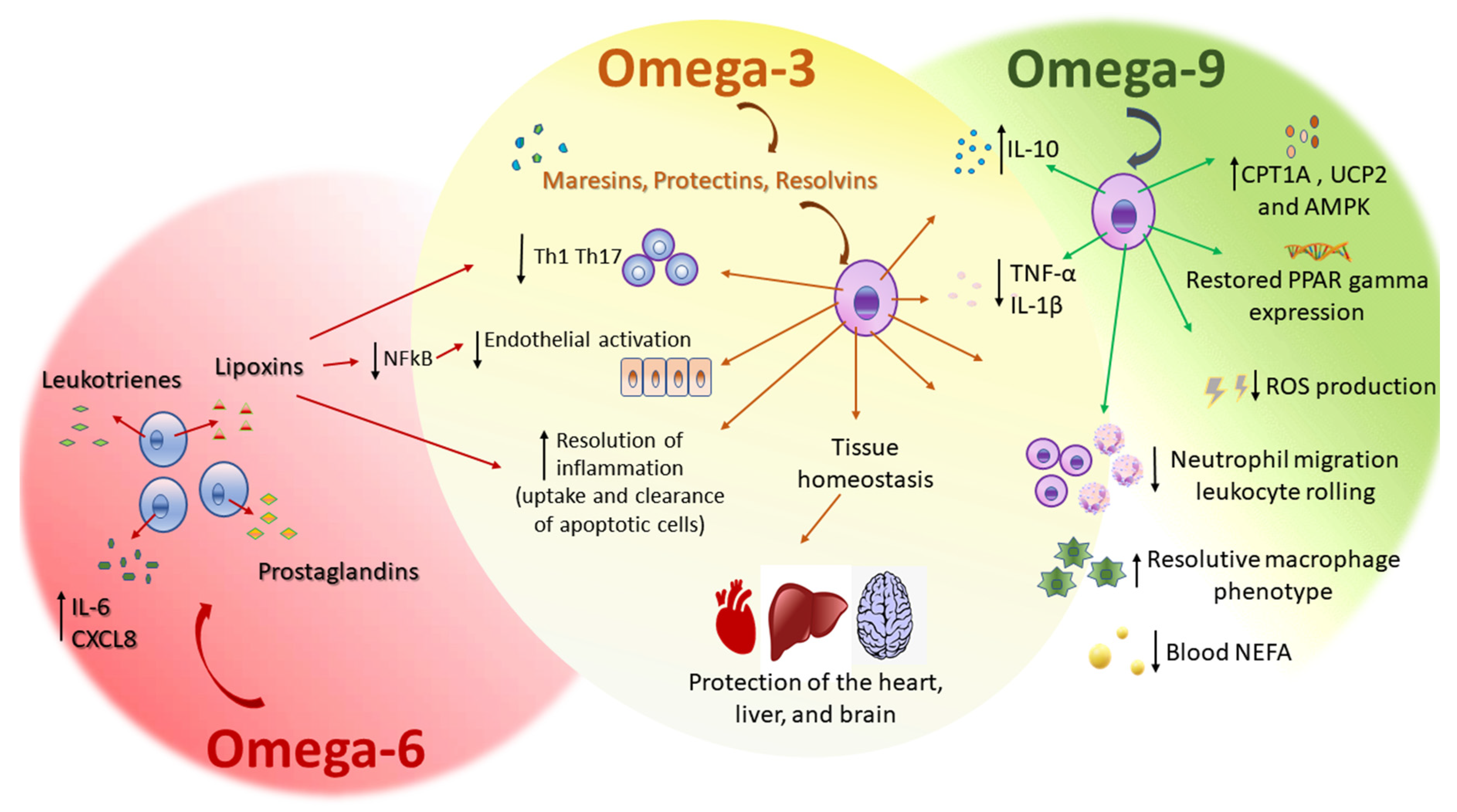
3. Inflammation and Fatty Acids
3.1. Omega-9
3.2. Omega-3
3.3. Omega-6
3.4. Omega-3 Versus Omega-6
4. Fatty Acids and Malaria
4.1. Fatty Acid Metabolism of the Parasite as a Target of Antimalarial Compounds
4.2. Omega-3, Omega-6, and Malaria
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Tuttolomondo, A.; Simonetta, I.; Daidone, M.; Mogavero, A.; Ortello, A.; Pinto, A. Metabolic and Vascular Effect of the Mediterranean Diet. Int. J. Mol. Sci. 2019, 20, 4716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, J.; Fialho, M.; Santos, R.; Peixoto-Placido, C.; Madeira, T.; Sousa-Santos, N.; Virgolino, A.; Santos, O.; Vaz Carneiro, A. Is olive oil good for you? A systematic review and meta-analysis on anti-inflammatory benefits from regular dietary intake. Nutrition 2020, 69, 110559. [Google Scholar] [CrossRef] [PubMed]
- Newman, T.M.; Vitolins, M.Z.; Cook, K.L. From the Table to the Tumor: The Role of Mediterranean and Western Dietary Patterns in Shifting Microbial-Mediated Signaling to Impact Breast Cancer Risk. Nutrients 2019, 11, 2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidharan, J.; Galie, S.; Hernandez-Alonso, P.; Bullo, M.; Salas-Salvado, J. Plant-Based Fat, Dietary Patterns Rich in Vegetable Fat and Gut Microbiota Modulation. Front. Nutr. 2019, 6, 157. [Google Scholar] [CrossRef] [Green Version]
- Kargin, D.; Tomaino, L.; Serra-Majem, L. Experimental Outcomes of the Mediterranean Diet: Lessons Learned from the Predimed Randomized Controlled Trial. Nutrients 2019, 11, 2991. [Google Scholar] [CrossRef] [Green Version]
- Ferro-Luzzi, A.; Branca, F. Mediterranean diet, Italian-style: Prototype of a healthy diet. Am. J. Clin. Nutr. 1995, 61 (Suppl. 6), 1338S–1345S. [Google Scholar] [CrossRef]
- Mastorakou, D.; Rabaeus, M.; Salen, P.; Pounis, G.; de Lorgeril, M. Mediterranean Diet: A Health-Protective Dietary Pattern for Modern Times. In Analysis in Nutrition Research, 1st ed.; Pounis, G., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 233–258. [Google Scholar]
- Simopoulos, A.P. Fish and Fish Oil in the Mediterranean Diets. In Fish and Fish Oil in Health and Disease Prevention; Susan, K., Raatz, D.M.B., Eds.; Academic Press—Elsevier: Amsterdam, The Netherlands, 2016; p. 380. [Google Scholar]
- Willett, W.C.; Sacks, F.; Trichopoulou, A.; Drescher, G.; Ferro-Luzzi, A.; Helsing, E.; Trichopoulos, D. Mediterranean diet pyramid: A cultural model for healthy eating. Am. J. Clin. Nutr. 1995, 61 (Suppl. 6), 1402S–1406S. [Google Scholar] [CrossRef]
- Galli, C.; Marangoni, F. N-3 fatty acids in the Mediterranean diet. Prostaglandins Leukot Essent Fatty Acids 2006, 75, 129–133. [Google Scholar] [CrossRef]
- Mazzocchi, A.; Leone, L.; Agostoni, C.; Pali-Scholl, I. The Secrets of the Mediterranean Diet. Does [Only] Olive Oil Matter? Nutrients 2019, 11, 2941. [Google Scholar] [CrossRef] [Green Version]
- Castiglione, D.; Platania, A.; Conti, A.; Falla, M.; D’Urso, M.; Marranzano, M. Dietary Micronutrient and Mineral Intake in the Mediterranean Healthy Eating, Ageing, and Lifestyle (MEAL) Study. Antioxidants 2018, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Bedard, A.; Riverin, M.; Dodin, S.; Corneau, L.; Lemieux, S. Sex differences in the impact of the Mediterranean diet on cardiovascular risk profile. Br. J. Nutr. 2012, 108, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Stamm, O.; Latscha, U.; Janecek, P.; Campana, A. Development of a special electrode for continuous subcutaneous pH measurement in the infant scalp. Am. J. Obstet. Gynecol. 1976, 124, 193–195. [Google Scholar] [CrossRef]
- Bedard, A.; Dodin, S.; Corneau, L.; Lemieux, S. The impact of abdominal obesity status on cardiovascular response to the mediterranean diet. J. Obes. 2012, 2012, 969124. [Google Scholar] [CrossRef] [PubMed]
- Mayr, H.L.; Thomas, C.J.; Tierney, A.C.; Kucianski, T.; George, E.S.; Ruiz-Canela, M.; Hebert, J.R.; Shivappa, N.; Itsiopoulos, C. Randomization to 6-month Mediterranean diet compared with a low-fat diet leads to improvement in Dietary Inflammatory Index scores in patients with coronary heart disease: The AUSMED Heart Trial. Nutr. Res. 2018, 55, 94–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkhatib, A.; Tsang, C.; Tuomilehto, J. Olive Oil Nutraceuticals in the Prevention and Management of Diabetes: From Molecules to Lifestyle. Int. J. Mol. Sci. 2018, 19, 2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yubero-Serrano, E.M.; Lopez-Moreno, J.; Gomez-Delgado, F.; Lopez-Miranda, J. Extra virgin olive oil: More than a healthy fat. Eur. J. Clin. Nutr. 2019, 72 (Suppl. 1), 8–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casas, R.; Estruch, R.; Sacanella, E. Influence of Bioactive Nutrients on the Atherosclerotic Process: A Review. Nutrients 2018, 10, 1630. [Google Scholar] [CrossRef] [Green Version]
- World-Health-Organization. World Malaria Report. 2015. Available online: https://apps.who.int/iris/bitstream/handle/10665/200018/9789241565158_eng.pdf;jsessionid=0F94417AFADC9C6AEA7DD8DDC7B6624E?sequence=1 (accessed on 30 December 2019).
- Cowman, A.F.; Healer, J.; Marapana, D.; Marsh, K. Malaria: Biology and Disease. Cell 2016, 167, 610–624. [Google Scholar] [CrossRef] [Green Version]
- World-Health-Organization. World Malaria Report. 2018. Available online: http://apps.who.int/iris/bitstream/handle/10665/275867/9789241565653-eng.pdf?ua=1 (accessed on 30 December 2019).
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Kabaria, C.W.; et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: Occurrence data, distribution maps and bionomic precis. Parasit Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [Green Version]
- Ashley, E.A.; Pyae Phyo, A.; Woodrow, C.J. Malaria. Lancet 2018, 391, 1608–1621. [Google Scholar] [CrossRef]
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josling, G.A.; Llinas, M. Sexual development in Plasmodium parasites: Knowing when it’s time to commit. Nat. Rev. Microbiol. 2015, 13, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Pajares, A.; Rogerson, S.J. The Rough Guide to Monocytes in Malaria Infection. Front. Immunol. 2018, 9, 2888. [Google Scholar] [CrossRef] [PubMed]
- Olotu, A.; Fegan, G.; Wambua, J.; Nyangweso, G.; Leach, A.; Lievens, M.; Kaslow, D.C.; Njuguna, P.; Marsh, K.; Bejon, P. Seven-Year Efficacy of RTS,S/AS01 Malaria Vaccine among Young African Children. N. Engl. J. Med. 2016, 374, 2519–2529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walk, J.; de Bree, L.C.J.; Graumans, W.; Stoter, R.; van Gemert, G.J.; van de Vegte-Bolmer, M.; Teelen, K.; Hermsen, C.C.; Arts, R.J.W.; Behet, M.C.; et al. Outcomes of controlled human malaria infection after BCG vaccination. Nat. Commun. 2019, 10, 874. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.C.; Kumari, V.; Dixit, R.; Tiwari, R.; Katyal, A. Metacaspases: Potential Drug Target against Protozoan Parasites. Front. Pharmacol. 2019, 10, 790. [Google Scholar] [CrossRef]
- Lacerda, M.V.G.; Llanos-Cuentas, A.; Krudsood, S.; Lon, C.; Saunders, D.L.; Mohammed, R.; Yilma, D.; Batista Pereira, D.; Espino, F.E.J.; Mia, R.Z.; et al. Single-Dose Tafenoquine to Prevent Relapse of Plasmodium vivax Malaria. N. Engl. J. Med. 2019, 380, 215–228. [Google Scholar] [CrossRef]
- Milner, D.A., Jr. Malaria Pathogenesis. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Anstey, N.M.; Douglas, N.M.; Poespoprodjo, J.R.; Price, R.N. Plasmodium vivax: Clinical spectrum, risk factors and pathogenesis. Adv. Parasitol. 2012, 80, 151–201. [Google Scholar]
- Goncalves-de-Albuquerque, C.F.; Rohwedder, I.; Silva, A.R.; Ferreira, A.S.; Kurz, A.R.M.; Cougoule, C.; Klapproth, S.; Eggersmann, T.; Silva, J.D.; de Oliveira, G.P.; et al. The Yin and Yang of Tyrosine Kinase Inhibition during Experimental Polymicrobial Sepsis. Front. Immunol. 2018, 9, 901. [Google Scholar] [CrossRef] [Green Version]
- Penha-Goncalves, C. Genetics of Malaria Inflammatory Responses: A Pathogenesis Perspective. Front. Immunol. 2019, 10, 1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shryock, N.; McBerry, C.; Salazar Gonzalez, R.M.; Janes, S.; Costa, F.T.; Aliberti, J. Lipoxin A(4) and 15-epi-lipoxin A(4) protect against experimental cerebral malaria by inhibiting IL-12/IFN-gamma in the brain. PLoS ONE 2013, 8, e61882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunnington, A.J.; Walther, M.; Riley, E.M. Piecing together the puzzle of severe malaria. Sci. Transl. Med. 2013, 5, 211ps18. [Google Scholar] [CrossRef] [PubMed]
- Shastri, A.; Bonifati, D.M.; Kishore, U. Innate immunity and neuroinflammation. Mediat. Inflamm. 2013, 2013, 342931. [Google Scholar] [CrossRef] [PubMed]
- Sypniewska, P.; Duda, J.F.; Locatelli, I.; Althaus, C.R.; Althaus, F.; Genton, B. Clinical and laboratory predictors of death in African children with features of severe malaria: A systematic review and meta-analysis. BMC Med. 2017, 15, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schofield, L.; Hackett, F. Signal transduction in host cells by a glycosylphosphatidylinositol toxin of malaria parasites. J. Exp. Med. 1993, 177, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Tachado, S.D.; Gerold, P.; McConville, M.J.; Baldwin, T.; Quilici, D.; Schwarz, R.T.; Schofield, L. Glycosylphosphatidylinositol toxin of Plasmodium induces nitric oxide synthase expression in macrophages and vascular endothelial cells by a protein tyrosine kinase-dependent and protein kinase C-dependent signaling pathway. J. Immunol. 1996, 156, 1897–1907. [Google Scholar]
- Schofield, L.; Novakovic, S.; Gerold, P.; Schwarz, R.T.; McConville, M.J.; Tachado, S.D. Glycosylphosphatidylinositol toxin of Plasmodium up-regulates intercellular adhesion molecule-1, vascular cell adhesion molecule-1, and E-selectin expression in vascular endothelial cells and increases leukocyte and parasite cytoadherence via tyrosine kinase-dependent signal transduction. J. Immunol. 1996, 156, 1886–1896. [Google Scholar]
- De Menezes, M.N.; Salles, E.M.; Vieira, F.; Amaral, E.P.; Zuzarte-Luis, V.; Cassado, A.; Epiphanio, S.; Alvarez, J.M.; Alves-Filho, J.C.; Mota, M.M.; et al. IL-1alpha promotes liver inflammation and necrosis during blood-stage Plasmodium chabaudi malaria. Sci. Rep. 2019, 9, 7575. [Google Scholar] [CrossRef]
- Ty, M.C.; Zuniga, M.; Gotz, A.; Kayal, S.; Sahu, P.K.; Mohanty, A.; Mohanty, S.; Wassmer, S.C.; Rodriguez, A. Malaria inflammation by xanthine oxidase-produced reactive oxygen species. EMBO Mol. Med. 2019, 11, e9903. [Google Scholar] [CrossRef]
- Sanchez-Arcila, J.C.; Perce-da-Silva, D.S.; Vasconcelos, M.P.; Rodrigues-da-Silva, R.N.; Pereira, V.A.; Aprigio, C.J.; Lima, C.A.; Fonseca e Fonseca, B.P.; Banic, D.M.; Lima-Junior, J.C.; et al. Intestinal parasites coinfection does not alter plasma cytokines profile elicited in acute malaria in subjects from endemic area of Brazil. Mediat. Inflamm. 2014, 2014, 857245. [Google Scholar] [CrossRef] [PubMed]
- Davenport, G.C.; Hittner, J.B.; Otieno, V.; Karim, Z.; Mukundan, H.; Fenimore, P.W.; Hengartner, N.W.; McMahon, B.H.; Kempaiah, P.; Ong’echa, J.M.; et al. Reduced Parasite Burden in Children with Falciparum Malaria and Bacteremia Coinfections: Role of Mediators of Inflammation. Mediat. Inflamm. 2016, 2016, 4286576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassmer, S.C.; Grau, G.E. Severe malaria: what’s new on the pathogenesis front? Int. J. Parasitol. 2017, 47, 145–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deroost, K.; Pham, T.T.; Opdenakker, G.; Van den Steen, P.E. The immunological balance between host and parasite in malaria. FEMS Microbiol. Rev. 2016, 40, 208–257. [Google Scholar] [CrossRef]
- Souza, M.C.; Padua, T.A.; Henriques, M.G. Endothelial-Leukocyte Interaction in Severe Malaria: Beyond the Brain. Mediat. Inflamm. 2015, 2015, 168937. [Google Scholar] [CrossRef] [Green Version]
- Bansal, G.P.; Weinstein, C.S.; Kumar, N. Insight into phagocytosis of mature sexual (gametocyte) stages of Plasmodium falciparum using a human monocyte cell line. Acta Trop. 2016, 157, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.A.; Stanisic, D.I.; Duffy, M.F.; Robinson, L.J.; Lin, E.; Kazura, J.W.; King, C.L.; Siba, P.M.; Fowkes, F.J.; Mueller, I.; et al. Patterns of protective associations differ for antibodies to P. falciparum-infected erythrocytes and merozoites in immunity against malaria in children. Eur. J. Immunol. 2017, 47, 2124–2136. [Google Scholar] [CrossRef] [Green Version]
- Lambert, L.H.; Bullock, J.L.; Cook, S.T.; Miura, K.; Garboczi, D.N.; Diakite, M.; Fairhurst, R.M.; Singh, K.; Long, C.A. Antigen reversal identifies targets of opsonizing IgGs against pregnancy-associated malaria. Infect. Immun. 2014, 82, 4842–4853. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Feng, G.; Beeson, J.; Hogarth, P.M.; Rogerson, S.J.; Yan, Y.; Jaworowski, A. CD14(hi)CD16+ monocytes phagocytose antibody-opsonised Plasmodium falciparum infected erythrocytes more efficiently than other monocyte subsets, and require CD16 and complement to do so. BMC Med. 2015, 13, 154. [Google Scholar] [CrossRef] [Green Version]
- Hommel, M.; Chan, J.A.; Umbers, A.J.; Langer, C.; Rogerson, S.J.; Smith, J.D.; Beeson, J.G. Evaluating antibody functional activity and strain-specificity of vaccine candidates for malaria in pregnancy using in vitro phagocytosis assays. Parasit Vectors 2018, 11, 69. [Google Scholar] [CrossRef] [Green Version]
- Kumsiri, R.; Troye-Blomberg, M.; Pattanapanyasat, K.; Krudsood, S.; Maneerat, Y. IgE low affinity receptor (CD23) expression, Plasmodium falciparum specific IgE and tumor necrosis factor-alpha production in Thai uncomplicated and severe falciparum malaria patients. Acta Trop. 2016, 154, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Pai, S.; Qin, J.; Cavanagh, L.; Mitchell, A.; El-Assaad, F.; Jain, R.; Combes, V.; Hunt, N.H.; Grau, G.E.; Weninger, W. Real-time imaging reveals the dynamics of leukocyte behaviour during experimental cerebral malaria pathogenesis. PLoS Pathog. 2014, 10, e1004236. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, L.J.; Nie, C.Q.; Ly, A.; Ryg-Cornejo, V.; Chiu, C.Y.; Hansen, D.S. Monocyte- and Neutrophil-Derived CXCL10 Impairs Efficient Control of Blood-Stage Malaria Infection and Promotes Severe Disease. J. Immunol. 2016, 196, 1227–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiadou, A.; Lee, H.J.; Walther, M.; van Beek, A.E.; Fitriani, F.; Wouters, D.; Kuijpers, T.W.; Nwakanma, D.; D’Alessandro, U.; Riley, E.M.; et al. Modelling pathogen load dynamics to elucidate mechanistic determinants of host-Plasmodium falciparum interactions. Nat. Microbiol. 2019, 4, 1592–1602. [Google Scholar] [CrossRef] [PubMed]
- Besnard, A.G.; Guabiraba, R.; Niedbala, W.; Palomo, J.; Reverchon, F.; Shaw, T.N.; Couper, K.N.; Ryffel, B.; Liew, F.Y. IL-33-mediated protection against experimental cerebral malaria is linked to induction of type 2 innate lymphoid cells, M2 macrophages and regulatory T cells. PLoS Pathog. 2015, 11, e1004607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomo, J.; Reverchon, F.; Piotet, J.; Besnard, A.G.; Couturier-Maillard, A.; Maillet, I.; Tefit, M.; Erard, F.; Mazier, D.; Ryffel, B.; et al. Critical role of IL-33 receptor ST2 in experimental cerebral malaria development. Eur. J. Immunol. 2015, 45, 1354–1365. [Google Scholar] [CrossRef] [Green Version]
- Wykes, M.N.; Lewin, S.R. Immune checkpoint blockade in infectious diseases. Nat. Rev. Immunol. 2018, 18, 91–104. [Google Scholar] [CrossRef]
- Hafalla, J.C.; Claser, C.; Couper, K.N.; Grau, G.E.; Renia, L.; de Souza, J.B.; Riley, E.M. The CTLA-4 and PD-1/PD-L1 inhibitory pathways independently regulate host resistance to Plasmodium-induced acute immune pathology. PLoS Pathog. 2012, 8, e1002504. [Google Scholar] [CrossRef]
- Karunarathne, D.S.; Horne-Debets, J.M.; Huang, J.X.; Faleiro, R.; Leow, C.Y.; Amante, F.; Watkins, T.S.; Miles, J.J.; Dwyer, P.J.; Stacey, K.J.; et al. Programmed Death-1 Ligand 2-Mediated Regulation of the PD-L1 to PD-1 Axis Is Essential for Establishing CD4(+) T Cell Immunity. Immunity 2016, 45, 333–345. [Google Scholar] [CrossRef] [Green Version]
- Butler, N.S.; Moebius, J.; Pewe, L.L.; Traore, B.; Doumbo, O.K.; Tygrett, L.T.; Waldschmidt, T.J.; Crompton, P.D.; Harty, J.T. Therapeutic blockade of PD-L1 and LAG-3 rapidly clears established blood-stage Plasmodium infection. Nat. Immunol. 2011, 13, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Zander, R.A.; Obeng-Adjei, N.; Guthmiller, J.J.; Kulu, D.I.; Li, J.; Ongoiba, A.; Traore, B.; Crompton, P.D.; Butler, N.S. PD-1 Co-inhibitory and OX40 Co-stimulatory Crosstalk Regulates Helper T Cell Differentiation and Anti-Plasmodium Humoral Immunity. Cell Host Microbe 2015, 17, 628–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, N.; Zou, Y.; Piao, X.; Liu, S.; Wang, L.; Li, S.; Chen, Q. T-Cell Immunoglobulin- and Mucin-Domain-Containing Molecule 3 Signaling Blockade Improves Cell-Mediated Immunity Against Malaria. J. Infect. Dis. 2016, 214, 1547–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tubo, N.J.; Jenkins, M.K. CD4+ T Cells: Guardians of the phagosome. Clin. Microbiol. Rev. 2014, 27, 200–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef]
- Freitas do Rosario, A.P.; Langhorne, J. T cell-derived IL-10 and its impact on the regulation of host responses during malaria. Int. J. Parasitol. 2012, 42, 549–555. [Google Scholar] [CrossRef]
- Kobayashi, F.; Ishida, H.; Matsui, T.; Tsuji, M. Effects of in vivo administration of anti-IL-10 or anti-IFN-gamma monoclonal antibody on the host defense mechanism against Plasmodium yoelii yoelii infection. J. Vet. Med. Sci. 2000, 62, 583–587. [Google Scholar] [CrossRef] [Green Version]
- Omer, F.M.; de Souza, J.B.; Riley, E.M. Differential induction of TGF-beta regulates proinflammatory cytokine production and determines the outcome of lethal and nonlethal Plasmodium yoelii infections. J. Immunol. 2003, 171, 5430–5436. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Ng, S.; Engwerda, C. The Role of IL-10 in Malaria: A Double Edged Sword. Front. Immunol. 2019, 10, 229. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, A.; Keswani, T.; Sarkar, S.; Ghosh, S.; Mukherjee, S.; Bhattacharyya, A. Autophagic induction modulates splenic plasmacytoid dendritic cell mediated immune response in cerebral malarial infection model. Microbes Infect. 2019, 21, 475–484. [Google Scholar] [CrossRef]
- Nakamae, S.; Kimura, D.; Miyakoda, M.; Sukhbaatar, O.; Inoue, S.I.; Yui, K. Role of IL-10 in inhibiting protective immune responses against infection with heterologous Plasmodium parasites. Parasitol. Int. 2019, 70, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Grau, G.E.; Heremans, H.; Piguet, P.F.; Pointaire, P.; Lambert, P.H.; Billiau, A.; Vassalli, P. Monoclonal antibody against interferon gamma can prevent experimental cerebral malaria and its associated overproduction of tumor necrosis factor. Proc. Natl. Acad. Sci. USA 1989, 86, 5572–5574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanez, D.M.; Manning, D.D.; Cooley, A.J.; Weidanz, W.P.; van der Heyde, H.C. Participation of lymphocyte subpopulations in the pathogenesis of experimental murine cerebral malaria. J. Immunol. 1996, 157, 1620–1624. [Google Scholar] [PubMed]
- Toda, A.; Terawaki, K.; Yamazaki, S.; Saeki, K.; Shimizu, T.; Yokomizo, T. Attenuated Th1 induction by dendritic cells from mice deficient in the leukotriene B4 receptor 1. Biochimie 2010, 92, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Arcoleo, F.; Milano, S.; D’Agostino, P.; Cillari, E. Effect of exogenous leukotriene B4 (LTB4) on BALB/c mice splenocyte production of Th1 and Th2 lymphokines. Int. J. Immunopharmacol. 1995, 17, 457–463. [Google Scholar] [CrossRef]
- Babu, K.S.; Salvi, S.S. Aspirin and asthma. Chest 2000, 118, 1470–1476. [Google Scholar] [CrossRef]
- Rogerio, A.P.; Anibal, F.F. Role of leukotrienes on protozoan and helminth infections. Mediat. Inflamm. 2012, 2012, 595694. [Google Scholar] [CrossRef] [Green Version]
- Foller, M.; Mahmud, H.; Gu, S.; Wang, K.; Floride, E.; Kucherenko, Y.; Luik, S.; Laufer, S.; Lang, F. Participation of leukotriene C(4) in the regulation of suicidal erythrocyte death. J. Physiol. Pharmacol. 2009, 60, 135–143. [Google Scholar]
- Yeo, K.T.; Embury, P.; Anderson, T.; Mungai, P.; Malhotra, I.; King, C.; Kazura, J.; Dent, A. HIV, Cytomegalovirus, and Malaria Infections during Pregnancy Lead to Inflammation and Shifts in Memory B Cell Subsets in Kenyan Neonates. J. Immunol. 2019, 202, 1465–1478. [Google Scholar] [CrossRef]
- Ly, A.; Hansen, D.S. Development of B Cell Memory in Malaria. Front. Immunol. 2019, 10, 559. [Google Scholar] [CrossRef]
- Storm, J.; Jespersen, J.S.; Seydel, K.B.; Szestak, T.; Mbewe, M.; Chisala, N.V.; Phula, P.; Wang, C.W.; Taylor, T.E.; Moxon, C.A.; et al. Cerebral malaria is associated with differential cytoadherence to brain endothelial cells. EMBO Mol. Med. 2019, 11, e9164. [Google Scholar] [CrossRef]
- Barrera, V.; Hiscott, P.S.; Craig, A.G.; White, V.A.; Milner, D.A.; Beare, N.A.; MacCormick, I.J.; Kamiza, S.; Taylor, T.E.; Molyneux, M.E.; et al. Severity of retinopathy parallels the degree of parasite sequestration in the eyes and brains of malawian children with fatal cerebral malaria. J. Infect. Dis. 2015, 211, 1977–1986. [Google Scholar] [CrossRef]
- Gillrie, M.R.; Avril, M.; Brazier, A.J.; Davis, S.P.; Stins, M.F.; Smith, J.D.; Ho, M. Diverse functional outcomes of Plasmodium falciparum ligation of EPCR: Potential implications for malarial pathogenesis. Cell. Microbiol. 2015, 17, 1883–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampath, S.; Brazier, A.J.; Avril, M.; Bernabeu, M.; Vigdorovich, V.; Mascarenhas, A.; Gomes, E.; Sather, D.N.; Esmon, C.T.; Smith, J.D. Plasmodium falciparum adhesion domains linked to severe malaria differ in blockade of endothelial protein C receptor. Cell. Microbiol. 2015, 17, 1868–1882. [Google Scholar] [CrossRef] [Green Version]
- Bernabeu, M.; Danziger, S.A.; Avril, M.; Vaz, M.; Babar, P.H.; Brazier, A.J.; Herricks, T.; Maki, J.N.; Pereira, L.; Mascarenhas, A.; et al. Severe adult malaria is associated with specific PfEMP1 adhesion types and high parasite biomass. Proc. Natl. Acad. Sci. USA 2016, 113, E3270–E3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, A.; Dankwa, S.; Bernabeu, M.; Harawa, V.; Danziger, S.A.; Duffy, F.; Kampondeni, S.D.; Potchen, M.J.; Dambrauskas, N.; Vigdorovich, V.; et al. Linking EPCR-Binding PfEMP1 to Brain Swelling in Pediatric Cerebral Malaria. Cell Host Microbe 2017, 22, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Brito, C.X.; Teixeira, S.A.; Barboza, R.; dos Reis, A.S.; Azevedo-Santos, A.P.; Muscara, M.; Costa, S.K.; Marinho, C.R.; Brain, S.D.; et al. TRPV1 antagonism by capsazepine modulates innate immune response in mice infected with Plasmodium berghei ANKA. Mediat. Inflamm. 2014, 2014, 506450. [Google Scholar] [CrossRef] [Green Version]
- Pereira, D.M.S.; Teixeira, S.A.; Murillo, O.; Peixoto, E.P.M.; Araujo, M.C.; Sousa, N.C.F.; Monteiro-Neto, V.; Calixto, J.B.; Cunha, T.M.; Marinho, C.R.F.; et al. TRPV1 Contributes to Cerebral Malaria Severity and Mortality by Regulating Brain Inflammation. Oxidative Med. Cell. Longev. 2019, 2019, 9451671. [Google Scholar] [CrossRef] [Green Version]
- Goncalves de Albuquerque, C.F.; Burth, P.; Younes Ibrahim, M.; Garcia, D.G.; Bozza, P.T.; Castro Faria Neto, H.C.; Castro Faria, M.V. Reduced plasma nonesterified fatty acid levels and the advent of an acute lung injury in mice after intravenous or enteral oleic acid administration. Mediat. Inflamm. 2012, 2012, 601032. [Google Scholar] [CrossRef] [Green Version]
- Goncalves-de-Albuquerque, C.F.; Burth, P.; Silva, A.R.; de Moraes, I.M.; de Jesus Oliveira, F.M.; Santelli, R.E.; Freire, A.S.; Bozza, P.T.; Younes-Ibrahim, M.; de Castro-Faria-Neto, H.C.; et al. Oleic acid inhibits lung Na/K-ATPase in mice and induces injury with lipid body formation in leukocytes and eicosanoid production. J. Inflamm. 2013, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Goncalves-de-Albuquerque, C.F.; Silva, A.R.; Burth, P.; de Moraes, I.M.; Oliveira, F.M.; Younes-Ibrahim, M.; dos Santos Mda, C.; D’Avila, H.; Bozza, P.T.; Faria Neto, H.C.; et al. Oleic acid induces lung injury in mice through activation of the ERK pathway. Mediat. Inflamm. 2012, 2012, 956509. [Google Scholar] [CrossRef] [Green Version]
- Goncalves-de-Albuquerque, C.F.; Medeiros-de-Moraes, I.M.; Oliveira, F.M.; Burth, P.; Bozza, P.T.; Castro Faria, M.V.; Silva, A.R.; Castro-Faria-Neto, H.C. Omega-9 Oleic Acid Induces Fatty Acid Oxidation and Decreases Organ Dysfunction and Mortality in Experimental Sepsis. PLoS ONE 2016, 11, e0153607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, C.V.; Campbell, C.; Goncalves-de-Albuquerque, C.F.; Molinaro, R.; Cody, M.J.; Yost, C.C.; Bozza, P.T.; Zimmerman, G.A.; Weyrich, A.S.; Castro-Faria-Neto, H.C.; et al. A PPARgamma agonist enhances bacterial clearance through neutrophil extracellular trap formation and improves survival in sepsis. Shock 2016, 45, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, C.V.; Estato, V.; Tibirica, E.; Bozza, P.T.; Castro-Faria-Neto, H.C.; Silva, A.R. PPAR gamma activation protects the brain against microvascular dysfunction in sepsis. Microvasc. Res. 2012, 84, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Medeiros-de-Moraes, I.M.; Goncalves-de-Albuquerque, C.F.; Kurz, A.R.M.; Oliveira, F.M.J.; de Abreu, V.H.P.; Torres, R.C.; Carvalho, V.F.; Estato, V.; Bozza, P.T.; Sperandio, M.; et al. Omega-9 Oleic Acid, the Main Compound of Olive Oil, Mitigates Inflammation during Experimental Sepsis. Oxidative Med. Cell. Longev. 2018, 2018, 6053492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pararasa, C.; Ikwuobe, J.; Shigdar, S.; Boukouvalas, A.; Nabney, I.T.; Brown, J.E.; Devitt, A.; Bailey, C.J.; Bennett, S.J.; Griffiths, H.R. Age-associated changes in long-chain fatty acid profile during healthy aging promote pro-inflammatory monocyte polarization via PPARgamma. Aging Cell 2016, 15, 128–139. [Google Scholar] [CrossRef]
- Ye, J.; Ghosh, S. Omega-3 PUFA vs. NSAIDs for Preventing Cardiac Inflammation. Front. Cardiovasc. Med. 2018, 5, 146. [Google Scholar] [CrossRef]
- Dessi, M.; Noce, A.; Bertucci, P.; Manca di Villahermosa, S.; Zenobi, R.; Castagnola, V.; Addessi, E.; Di Daniele, N. Atherosclerosis, dyslipidemia, and inflammation: The significant role of polyunsaturated Fatty acids. ISRN Inflamm. 2013, 2013, 191823. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, T.; Yoshida, M.; Arita, M. Omega-3 fatty acid-derived mediators that control inflammation and tissue homeostasis. Int. Immunol. 2019, 9, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Kendall, A.C.; Pilkington, S.M.; Murphy, S.A.; Del Carratore, F.; Sunarwidhi, A.L.; Kiezel-Tsugunova, M.; Urquhart, P.; Watson, R.E.B.; Breitling, R.; Rhodes, L.E.; et al. Dynamics of the human skin mediator lipidome in response to dietary omega-3 fatty acid supplementation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 13014–13027. [Google Scholar]
- Dalli, J.; Serhan, C.N. Pro-Resolving Mediators in Regulating and Conferring Macrophage Function. Front. Immunol. 2017, 8, 1400. [Google Scholar] [CrossRef] [Green Version]
- Weylandt, K.H. Docosapentaenoic acid derived metabolites and mediators—The new world of lipid mediator medicine in a nutshell. Eur. J. Pharmacol. 2016, 785, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Guo, X.F.; Sinclair, A.J. Short update on docosapentaenoic acid: A bioactive long-chain n-3 fatty acid. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.; Hiram, R.; Rousseau, E.; Blier, P.U.; Fortin, S. Docosapentaenoic acid monoacylglyceride reduces inflammation and vascular remodeling in experimental pulmonary hypertension. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H574–H586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalli, J.; Colas, R.A.; Serhan, C.N. Novel n-3 immunoresolvents: Structures and actions. Sci. Rep. 2013, 3, 1940. [Google Scholar] [CrossRef]
- Aursnes, M.; Tungen, J.E.; Vik, A.; Colas, R.; Cheng, C.Y.; Dalli, J.; Serhan, C.N.; Hansen, T.V. Total synthesis of the lipid mediator PD1n-3 DPA: Configurational assignments and anti-inflammatory and pro-resolving actions. J. Nat. Prod. 2014, 77, 910–916. [Google Scholar] [CrossRef]
- Pistorius, K.; Souza, P.R.; De Matteis, R.; Austin-Williams, S.; Primdahl, K.G.; Vik, A.; Mazzacuva, F.; Colas, R.A.; Marques, R.M.; Hansen, T.V.; et al. PDn-3 DPA Pathway Regulates Human Monocyte Differentiation and Macrophage Function. Cell Chem. Biol. 2018, 25, 749–760. [Google Scholar] [CrossRef]
- Tungen, J.E.; Aursnes, M.; Dalli, J.; Arnardottir, H.; Serhan, C.N.; Hansen, T.V. Total synthesis of the anti-inflammatory and pro-resolving lipid mediator MaR1n-3 DPA utilizing an sp(3)-sp(3) Negishi cross-coupling reaction. Chemistry 2014, 20, 14575–14578. [Google Scholar] [CrossRef] [Green Version]
- Skulas-Ray, A.C.; Flock, M.R.; Richter, C.K.; Harris, W.S.; West, S.G.; Kris-Etherton, P.M. Red Blood Cell Docosapentaenoic Acid (DPA n-3) is Inversely Associated with Triglycerides and C-reactive Protein (CRP) in Healthy Adults and Dose-Dependently Increases Following n-3 Fatty Acid Supplementation. Nutrients 2015, 7, 6390–6404. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Dai, Z.; Cao, Y.; Shen, Q.; Zhang, Y. Docosapentaenoic acid (DPA, 22:5n-3) ameliorates inflammation in an ulcerative colitis model. Food Funct. 2019, 10, 4199–4209. [Google Scholar] [CrossRef]
- Chiurchiu, V.; Leuti, A.; Saracini, S.; Fontana, D.; Finamore, P.; Giua, R.; Padovini, L.; Incalzi, R.A.; Maccarrone, M. Resolution of inflammation is altered in chronic heart failure and entails a dysfunctional responsiveness of T lymphocytes. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 909–916. [Google Scholar] [CrossRef]
- Lopez-Munoz, R.A.; Molina-Berrios, A.; Campos-Estrada, C.; Abarca-Sanhueza, P.; Urrutia-Llancaqueo, L.; Pena-Espinoza, M.; Maya, J.D. Inflammatory and Pro-resolving Lipids in Trypanosomatid Infections: A Key to Understanding Parasite Control. Front. Microbiol. 2018, 9, 1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, C.D.; Schwarze, J. The role of pro-resolution lipid mediators in infectious disease. Immunology 2014, 141, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot Essent Fatty Acids 2018, 132, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutting, S.; Zakarya, R.; Bozier, J.; Xenaki, D.; Horvat, J.C.; Wood, L.G.; Hansbro, P.M.; Oliver, B.G. Dietary Fatty Acids Amplify Inflammatory Responses to Infection through p38 MAPK Signaling. Am. J. Respir. Cell Mol. Biol. 2019, 60, 554–568. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.; Godson, C. Lipoxins: Regulators of resolution. Curr. Opin. Pharmacol. 2010, 10, 166–172. [Google Scholar] [CrossRef]
- Wu, B.; Walker, J.; Spur, B.; Rodriguez, A.; Yin, K. Effects of Lipoxin A4 on antimicrobial actions of neutrophils in sepsis. Prostaglandins Leukot Essent Fatty Acids 2015, 94, 55–64. [Google Scholar] [CrossRef]
- Romano, M.; Cianci, E.; Simiele, F.; Recchiuti, A. Lipoxins and aspirin-triggered lipoxins in resolution of inflammation. Eur. J. Pharmacol. 2015, 760, 49–63. [Google Scholar] [CrossRef]
- Brennan, E.P.; Mohan, M.; McClelland, A.; de Gaetano, M.; Tikellis, C.; Marai, M.; Crean, D.; Dai, A.; Beuscart, O.; Derouiche, S.; et al. Lipoxins Protect Against Inflammation in Diabetes-Associated Atherosclerosis. Diabetes 2018, 67, 2657–2667. [Google Scholar] [CrossRef] [Green Version]
- Brennan, E.P.; Cacace, A.; Godson, C. Specialized pro-resolving mediators in renal fibrosis. Mol. Asp. Med. 2017, 58, 102–113. [Google Scholar] [CrossRef]
- Svensson, C.I.; Zattoni, M.; Serhan, C.N. Lipoxins and aspirin-triggered lipoxin inhibit inflammatory pain processing. J. Exp. Med. 2007, 204, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basil, M.C.; Levy, B.D. Specialized pro-resolving mediators: Endogenous regulators of infection and inflammation. Nat. Rev. Immunol. 2016, 16, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.M.; Roca, F.J.; Oh, S.F.; McFarland, R.; Vickery, T.W.; Ray, J.P.; Ko, D.C.; Zou, Y.; Bang, N.D.; Chau, T.T.; et al. Host genotype-specific therapies can optimize the inflammatory response to mycobacterial infections. Cell 2012, 148, 434–446. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Divangahi, M.; Gan, H.; Shin, D.S.; Hong, S.; Lee, D.M.; Serhan, C.N.; Behar, S.M.; Remold, H.G. Lipid mediators in innate immunity against tuberculosis: Opposing roles of PGE2 and LXA4 in the induction of macrophage death. J. Exp. Med. 2008, 205, 2791–2801. [Google Scholar] [CrossRef] [Green Version]
- Molina-Berrios, A.; Campos-Estrada, C.; Henriquez, N.; Faundez, M.; Torres, G.; Castillo, C.; Escanilla, S.; Kemmerling, U.; Morello, A.; Lopez-Munoz, R.A.; et al. Protective role of acetylsalicylic acid in experimental Trypanosoma cruzi infection: Evidence of a 15-epi-lipoxin A(4)-mediated effect. PLoS Negl. Trop. Dis. 2013, 7, e2173. [Google Scholar] [CrossRef]
- Aliberti, J.; Serhan, C.; Sher, A. Parasite-induced lipoxin A4 is an endogenous regulator of IL-12 production and immunopathology in Toxoplasma gondii infection. J. Exp. Med. 2002, 196, 1253–1262. [Google Scholar] [CrossRef]
- Walker, J.; Dichter, E.; Lacorte, G.; Kerner, D.; Spur, B.; Rodriguez, A.; Yin, K. Lipoxin a4 increases survival by decreasing systemic inflammation and bacterial load in sepsis. Shock 2011, 36, 410–416. [Google Scholar] [CrossRef]
- Merched, A.J.; Serhan, C.N.; Chan, L. Nutrigenetic disruption of inflammation-resolution homeostasis and atherogenesis. J. Nutr. Nutr. 2011, 4, 12–24. [Google Scholar] [CrossRef] [Green Version]
- Campos-Estrada, C.; Liempi, A.; Gonzalez-Herrera, F.; Lapier, M.; Kemmerling, U.; Pesce, B.; Ferreira, J.; Lopez-Munoz, R.; Maya, J.D. Simvastatin and Benznidazole-Mediated Prevention of Trypanosoma cruzi-Induced Endothelial Activation: Role of 15-epi-lipoxin A4 in the Action of Simvastatin. PLoS Negl. Trop. Dis. 2015, 9, e0003770. [Google Scholar] [CrossRef]
- Gonzalez-Herrera, F.; Cramer, A.; Pimentel, P.; Castillo, C.; Liempi, A.; Kemmerling, U.; Machado, F.S.; Maya, J.D. Simvastatin Attenuates Endothelial Activation through 15-Epi-Lipoxin A4 Production in Murine Chronic Chagas Cardiomyopathy. Antimicrob. Agents Chemother. 2017, 61, e02137-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiNicolantonio, J.J.; O’Keefe, J.H. Importance of maintaining a low omega-6/omega-3 ratio for reducing inflammation. Open Heart 2018, 5, e000946. [Google Scholar] [CrossRef] [PubMed]
- Uchi, S.H.; Yanai, R.; Kobayashi, M.; Hatano, M.; Kobayashi, Y.; Yamashiro, C.; Nagai, T.; Tokuda, K.; Connor, K.M.; Sonoda, K.H.; et al. Dendritic cells mediate the anti-inflammatory action of omega-3 long-chain polyunsaturated fatty acids in experimental autoimmune uveitis. PLoS ONE 2019, 14, e0219405. [Google Scholar] [CrossRef]
- Shoda, H.; Yanai, R.; Yoshimura, T.; Nagai, T.; Kimura, K.; Sobrin, L.; Connor, K.M.; Sakoda, Y.; Tamada, K.; Ikeda, T.; et al. Dietary Omega-3 Fatty Acids Suppress Experimental Autoimmune Uveitis in Association with Inhibition of Th1 and Th17 Cell Function. PLoS ONE 2015, 10, e0138241. [Google Scholar] [CrossRef]
- Tarun, A.S.; Vaughan, A.M.; Kappe, S.H. Redefining the role of de novo fatty acid synthesis in Plasmodium parasites. Trends Parasitol. 2009, 25, 545–550. [Google Scholar] [CrossRef] [PubMed]
- van Schaijk, B.C.; Kumar, T.R.; Vos, M.W.; Richman, A.; van Gemert, G.J.; Li, T.; Eappen, A.G.; Williamson, K.C.; Morahan, B.J.; Fishbaugher, M.; et al. Type II fatty acid biosynthesis is essential for Plasmodium falciparum sporozoite development in the midgut of Anopheles mosquitoes. Eukaryot. Cell 2014, 13, 550–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Kumar, T.R.; Nkrumah, L.J.; Coppi, A.; Retzlaff, S.; Li, C.D.; Kelly, B.J.; Moura, P.A.; Lakshmanan, V.; Freundlich, J.S.; et al. The fatty acid biosynthesis enzyme FabI plays a key role in the development of liver-stage malarial parasites. Cell Host Microbe 2008, 4, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, A.M.; O’Neill, M.T.; Tarun, A.S.; Camargo, N.; Phuong, T.M.; Aly, A.S.; Cowman, A.F.; Kappe, S.H. Type II fatty acid synthesis is essential only for malaria parasite late liver stage development. Cell. Microbiol. 2009, 11, 506–520. [Google Scholar] [CrossRef] [Green Version]
- Tasdemir, D.; Sanabria, D.; Lauinger, I.L.; Tarun, A.; Herman, R.; Perozzo, R.; Zloh, M.; Kappe, S.H.; Brun, R.; Carballeira, N.M. 2-Hexadecynoic acid inhibits plasmodial FAS-II enzymes and arrests erythrocytic and liver stage Plasmodium infections. Bioorg. Med. Chem. 2010, 18, 7475–7485. [Google Scholar] [CrossRef] [Green Version]
- Carballeira, N.M.; Bwalya, A.G.; Itoe, M.A.; Andricopulo, A.D.; Cordero-Maldonado, M.L.; Kaiser, M.; Mota, M.M.; Crawford, A.D.; Guido, R.V.; Tasdemir, D. 2-Octadecynoic acid as a dual life stage inhibitor of Plasmodium infections and plasmodial FAS-II enzymes. Bioorg. Med. Chem. Lett. 2014, 24, 4151–4157. [Google Scholar] [CrossRef]
- Lauinger, I.L.; Vivas, L.; Perozzo, R.; Stairiker, C.; Tarun, A.; Zloh, M.; Zhang, X.; Xu, H.; Tonge, P.J.; Franzblau, S.G.; et al. Potential of lichen secondary metabolites against Plasmodium liver stage parasites with FAS-II as the potential target. J. Nat. Prod. 2013, 76, 1064–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freundlich, J.S.; Anderson, J.W.; Sarantakis, D.; Shieh, H.M.; Yu, M.; Valderramos, J.C.; Lucumi, E.; Kuo, M.; Jacobs, W.R., Jr.; Fidock, D.A.; et al. Synthesis, biological activity, and X-ray crystal structural analysis of diaryl ether inhibitors of malarial enoyl acyl carrier protein reductase. Part 1: 4’-substituted triclosan derivatives. Bioorg. Med. Chem. Lett. 2005, 15, 5247–5252. [Google Scholar] [CrossRef] [PubMed]
- Surolia, A.; Ramya, T.N.; Ramya, V.; Surolia, N. ‘FAS’t inhibition of malaria. Biochem. J. 2004, 383, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Sauleau, P.; Moriou, C.; Al Mourabit, A. Metabolomics approach to chemical diversity of the Mediterranean marine sponge Agelas oroides. Nat. Prod. Res. 2017, 31, 1625–1632. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, D.; Topaloglu, B.; Perozzo, R.; Brun, R.; O’Neill, R.; Carballeira, N.M.; Zhang, X.; Tonge, P.J.; Linden, A.; Ruedi, P. Marine natural products from the Turkish sponge Agelas oroides that inhibit the enoyl reductases from Plasmodium falciparum, Mycobacterium tuberculosis and Escherichia coli. Bioorg. Med. Chem. 2007, 15, 6834–6845. [Google Scholar] [CrossRef]
- Kumar, G.; Parasuraman, P.; Sharma, S.K.; Banerjee, T.; Karmodiya, K.; Surolia, N.; Surolia, A. Discovery of a rhodanine class of compounds as inhibitors of Plasmodium falciparum enoyl-acyl carrier protein reductase. J. Med. Chem. 2007, 50, 2665–2675. [Google Scholar] [CrossRef]
- Kumar, G.; Banerjee, T.; Kapoor, N.; Surolia, N.; Surolia, A. SAR and pharmacophore models for the rhodanine inhibitors of Plasmodium falciparum enoyl-acyl carrier protein reductase. IUBMB Life 2010, 62, 204–213. [Google Scholar] [CrossRef]
- Samuel, M.; Oliver, S.V.; Wood, O.R.; Coetzee, M.; Brooke, B.D. Evaluation of the toxicity and repellence of an organic fatty acids mixture (C8910) against insecticide susceptible and resistant strains of the major malaria vector Anopheles funestus Giles (Diptera: Culicidae). Parasit Vectors 2015, 8, 321. [Google Scholar] [CrossRef] [Green Version]
- Delhaye, J.; Glaizot, O.; Christe, P. The effect of dietary antioxidant supplementation in a vertebrate host on the infection dynamics and transmission of avian malaria to the vector. Parasitol. Res. 2018, 117, 2043–2052. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, D.G. Antiparasitic action of dietary cod liver oil upon Plasmodium berghei and its reversal by vitamin E. Exp. Parasitol. 1957, 6, 555–565. [Google Scholar] [CrossRef]
- Levander, O.A.; Ager, A.L., Jr.; Morris, V.C.; May, R.G. Plasmodium yoelii: Comparative antimalarial activities of dietary fish oils and fish oil concentrates in vitamin E-deficient mice. Exp. Parasitol. 1990, 70, 323–329. [Google Scholar] [CrossRef]
- Fujikawa, M.; Kamitani, T.; Tunru, I.; Yamazaki, K.; Hamazaki, T. Antimalarial effects of purified and α-tocopherol—Fortified n-3 polyunsaturated fatty acids. J. Nutr. Biochem. 1993, 4, 153–157. [Google Scholar] [CrossRef]
- Endres, S.; Ghorbani, R.; Kelley, V.E.; Georgilis, K.; Lonnemann, G.; van der Meer, J.W.; Cannon, J.G.; Rogers, T.S.; Klempner, M.S.; Weber, P.C.; et al. The effect of dietary supplementation with n-3 polyunsaturated fatty acids on the synthesis of interleukin-1 and tumor necrosis factor by mononuclear cells. N. Engl. J. Med. 1989, 320, 265–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fevang, P.; Saav, H.; Hostmark, A.T. Dietary fish oils and long-term malaria protection in mice. Lipids 1995, 30, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Krugliak, M.; Deharo, E.; Shalmiev, G.; Sauvain, M.; Moretti, C.; Ginsburg, H. Antimalarial effects of C18 fatty acids on Plasmodium falciparum in culture and on Plasmodium vinckei petteri and Plasmodium yoelii nigeriensis in vivo. Exp. Parasitol. 1995, 81, 97–105. [Google Scholar] [CrossRef]
- Kumaratilake, L.M.; Ferrante, A.; Robinson, B.S.; Jaeger, T.; Poulos, A. Enhancement of neutrophil-mediated killing of Plasmodium falciparum asexual blood forms by fatty acids: Importance of fatty acid structure. Infect. Immun. 1997, 65, 4152–4157. [Google Scholar] [CrossRef] [Green Version]
- Arun Kumar, C.; Das, U.N. Lipid peroxides, nitric oxide and essential fatty acids in patients with Plasmodium falciparum malaria. Prostaglandins Leukot Essent Fatty Acids 1999, 61, 255–258. [Google Scholar] [CrossRef]
- Hamazaki, K.; Syafruddin, D.; Tunru, I.S.; Azwir, M.F.; Asih, P.B.; Sawazaki, S.; Hamazaki, T. The effects of docosahexaenoic acid-rich fish oil on behavior, school attendance rate and malaria infection in school children—a double-blind, randomized, placebo-controlled trial in Lampung, Indonesia. Asia Pac. J. Clin. Nutr. 2008, 17, 258–263. [Google Scholar]
- Melariri, P.; Campbell, W.; Etusim, P.; Smith, P. In Vitro and in Vivo Antimalarial Activity of Linolenic and Linoleic Acids and their Methyl Esters. Adv. Stud. Biol. 2012, 4, 333–349. [Google Scholar]
- Villegas-Mendez, A.; Greig, R.; Shaw, T.N.; de Souza, J.B.; Gwyer Findlay, E.; Stumhofer, J.S.; Hafalla, J.C.; Blount, D.G.; Hunter, C.A.; Riley, E.M.; et al. IFN-gamma-producing CD4+ T cells promote experimental cerebral malaria by modulating CD8+ T cell accumulation within the brain. J. Immunol. 2012, 189, 968–979. [Google Scholar] [CrossRef] [Green Version]
- Ademolue, T.W.; Aniweh, Y.; Kusi, K.A.; Awandare, G.A. Patterns of inflammatory responses and parasite tolerance vary with malaria transmission intensity. Malar. J. 2017, 16, 145. [Google Scholar] [CrossRef] [PubMed]
- Clark, I.A.; Alleva, L.M.; Budd, A.C.; Cowden, W.B. Understanding the role of inflammatory cytokines in malaria and related diseases. Travel Med. Infect. Dis. 2008, 6, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Gozzelino, R.; Jeney, V.; Soares, M.P. Mechanisms of cell protection by heme oxygenase-1. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 323–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Z.; Zhang, Y.C.; Cheng, J.S.; Ni, Q.; Li, P.J.; Wang, S.W.; Han, W.; Zhang, Y.L. BML-111, a lipoxin receptor agonist, ameliorates ‘two-hit’-induced acute pancreatitis-associated lung injury in mice by the upregulation of heme oxygenase-1. Artif. Cells Nanomed. Biotechnol. 2014, 42, 110–120. [Google Scholar] [CrossRef]
- Lv, W.; Lv, C.; Yu, S.; Yang, Y.; Kong, H.; Xie, J.; Sun, H.; Andersson, R.; Xu, D.; Chen, B.; et al. Lipoxin A4 attenuation of endothelial inflammation response mimicking pancreatitis-induced lung injury. Exp. Biol. Med. 2013, 238, 1388–1395. [Google Scholar] [CrossRef]
- Belhaj, A.; Dewachter, L.; Kerbaul, F.; Brimioulle, S.; Dewachter, C.; Naeije, R.; Rondelet, B. Heme oxygenase-1 and inflammation in experimental right ventricular failure on prolonged overcirculation-induced pulmonary hypertension. PLoS ONE 2013, 8, e69470. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, P.; Zhang, Z.; Hu, Q.; He, J.; Liu, H.; Zhao, J.; Liang, Y.; He, Z.; Li, X.; et al. LXA4 ameliorates cerebrovascular endothelial dysfunction by reducing acute inflammation after subarachnoid hemorrhage in rats. Neuroscience 2019, 408, 105–114. [Google Scholar] [CrossRef]
- Pamplona, A.; Ferreira, A.; Balla, J.; Jeney, V.; Balla, G.; Epiphanio, S.; Chora, A.; Rodrigues, C.D.; Gregoire, I.P.; Cunha-Rodrigues, M.; et al. Heme oxygenase-1 and carbon monoxide suppress the pathogenesis of experimental cerebral malaria. Nat. Med. 2007, 13, 703–710. [Google Scholar] [CrossRef]
- Padua, T.A.; Torres, N.D.; Candea, A.L.P.; Costa, M.F.S.; Silva, J.D.; Silva-Filho, J.L.; Costa, F.T.M.; Rocco, P.R.M.; Souza, M.C.; Henriques, M.G. Therapeutic effect of Lipoxin A4 in malaria-induced acute lung injury. J. Leukoc. Biol. 2018, 103, 657–670. [Google Scholar] [CrossRef]
- Zuzarte-Luis, V.; Mello-Vieira, J.; Marreiros, I.M.; Liehl, P.; Chora, A.F.; Carret, C.K.; Carvalho, T.; Mota, M.M. Dietary alterations modulate susceptibility to Plasmodium infection. Nat. Microbiol. 2017, 2, 1600–1607. [Google Scholar] [CrossRef]
- Luisi, M.L.E.; Lucarini, L.; Biffi, B.; Rafanelli, E.; Pietramellara, G.; Durante, M.; Vidali, S.; Provensi, G.; Madiai, S.; Gheri, C.F.; et al. Effect of Mediterranean Diet Enriched in High Quality Extra Virgin Olive Oil on Oxidative Stress, Inflammation and Gut Microbiota in Obese and Normal Weight Adult Subjects. Front. Pharmacol. 2019, 10, 1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra-Majem, L.; Roman-Vinas, B.; Sanchez-Villegas, A.; Guasch-Ferre, M.; Corella, D.; La Vecchia, C. Benefits of the Mediterranean diet: Epidemiological and molecular aspects. Mol. Asp. Med. 2019, 67, 1–55. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Morze, J.; Hoffmann, G. Mediterranean diet and health status: Active ingredients and pharmacological mechanisms. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Nutrients | Amount |
|---|---|
| Carbohydrate (g) | 194.0 |
| Fiber (g) | 41.1 |
| Protein (g) | 87.3 |
| Fat (g) | 106.0 |
| SFA (g) | 20.9 |
| MUFA (g) | 52.5 |
| PUFA (g) | 24.8 |
| Cholesterol (mg) | 289.7 |
| Alcohol (g) | 11.1 |
| MUFA to SFA ratio | 2.7 |
| Omega-3 (g) | 5.9 |
| Omega-6 (g) | 18.7 |
| Cholesterol (mg) | 255.8 |
| Beta carotene (μg) | 3039 |
| Vitamin A (RE) | 2161.6 |
| Vitamin D (μg) | 3.2 |
| Vitamin E (mg) | 29.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.R.; Moraes, B.P.T.; Gonçalves-de-Albuquerque, C.F. Mediterranean Diet: Lipids, Inflammation, and Malaria Infection. Int. J. Mol. Sci. 2020, 21, 4489. https://doi.org/10.3390/ijms21124489
Silva AR, Moraes BPT, Gonçalves-de-Albuquerque CF. Mediterranean Diet: Lipids, Inflammation, and Malaria Infection. International Journal of Molecular Sciences. 2020; 21(12):4489. https://doi.org/10.3390/ijms21124489
Chicago/Turabian StyleSilva, Adriana R., Bianca P. T. Moraes, and Cassiano F. Gonçalves-de-Albuquerque. 2020. "Mediterranean Diet: Lipids, Inflammation, and Malaria Infection" International Journal of Molecular Sciences 21, no. 12: 4489. https://doi.org/10.3390/ijms21124489
APA StyleSilva, A. R., Moraes, B. P. T., & Gonçalves-de-Albuquerque, C. F. (2020). Mediterranean Diet: Lipids, Inflammation, and Malaria Infection. International Journal of Molecular Sciences, 21(12), 4489. https://doi.org/10.3390/ijms21124489