Development of Specific Thinopyrum Cytogenetic Markers for Wheat-Wheatgrass Hybrids Using Sequencing and qPCR Data

 , , and
, , and
Abstract
:1. Introduction
2. Results
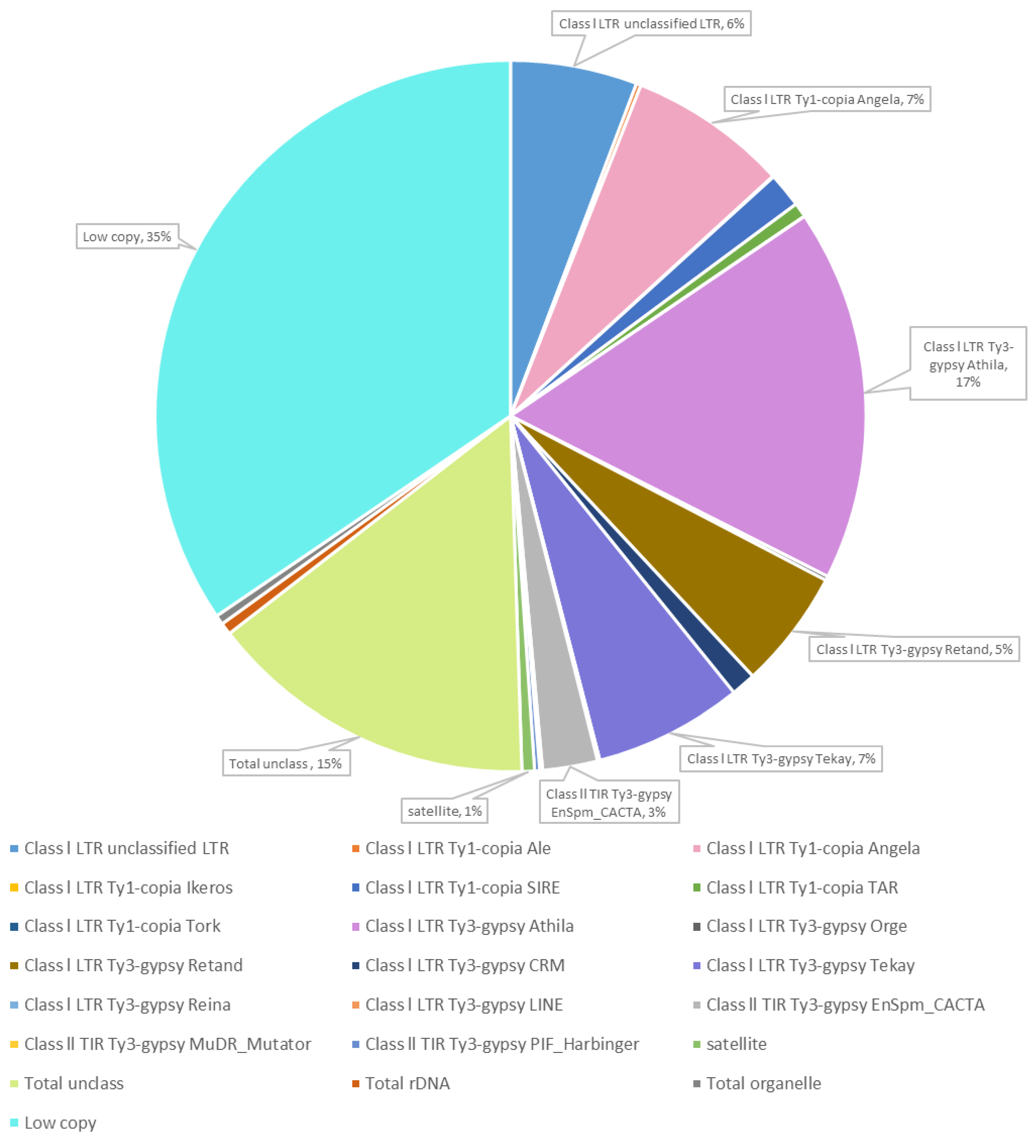
2.1. Bioinformatic Analysis of the Thinopyrum Ponticum Genome. The Search of the TRs and Primer Design
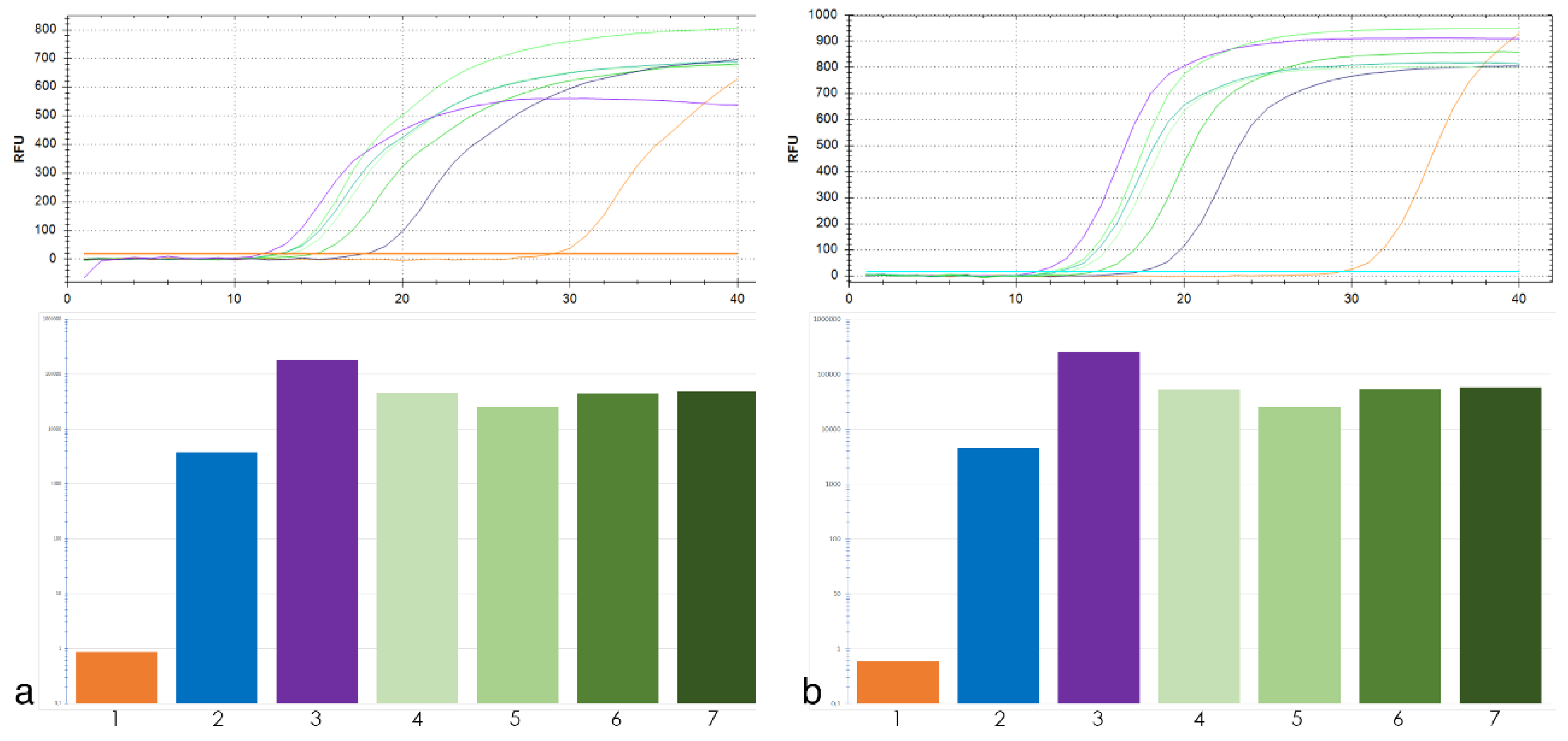
2.2. qPCR Analysis of the TANDEM REPEATs
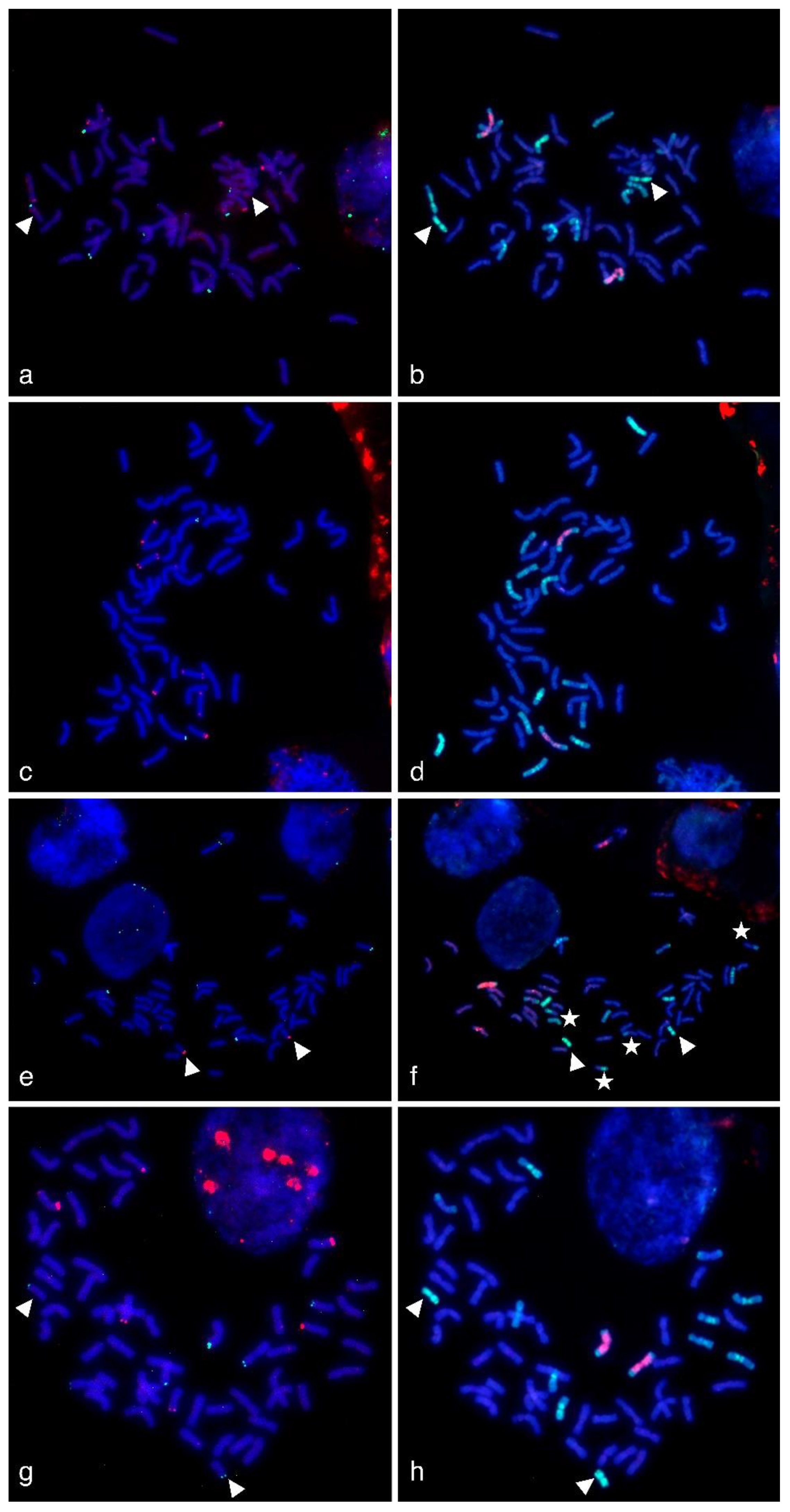
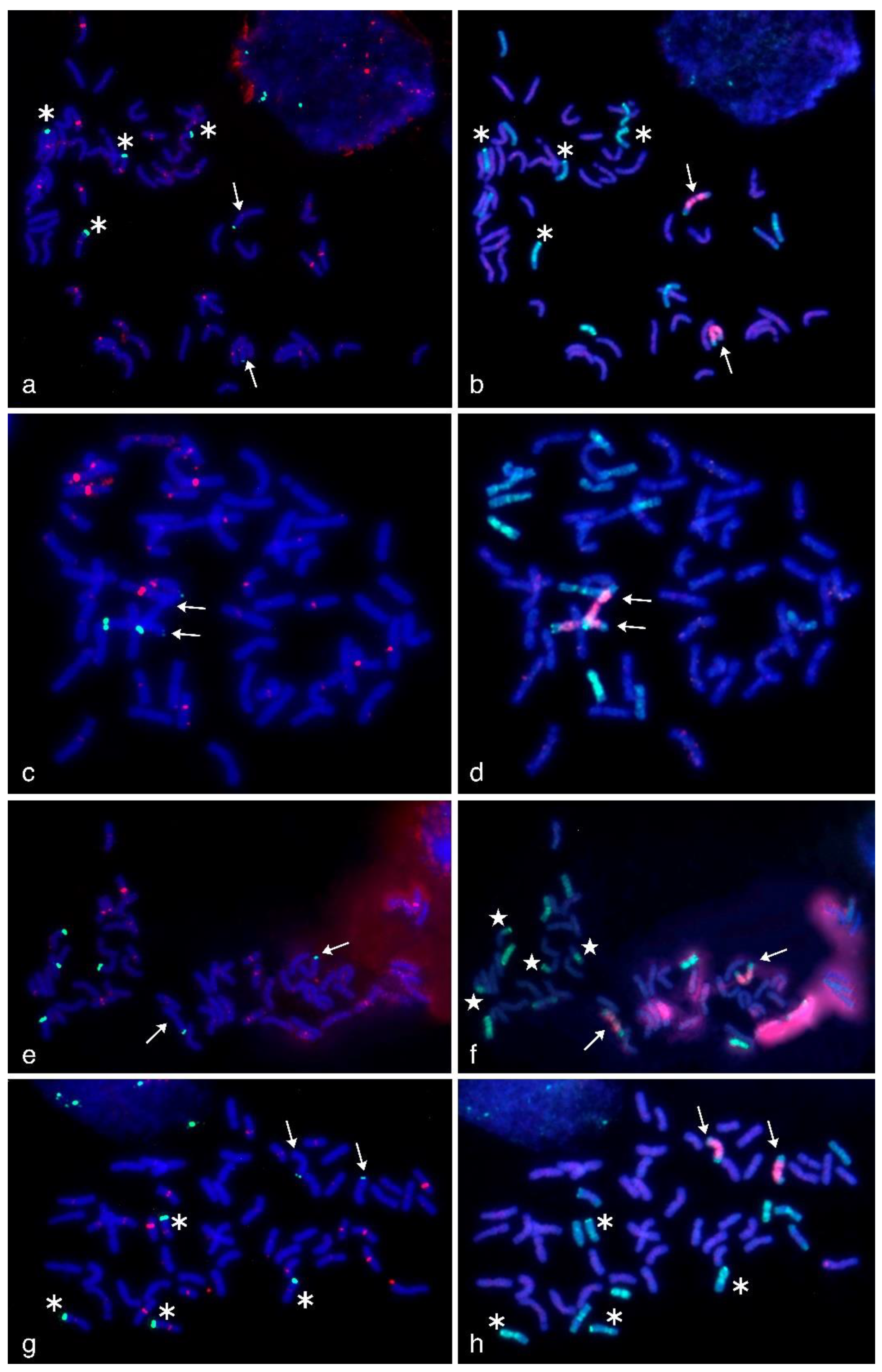
2.3. FISH/GISH Experiments
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Sequencing and Bioinformatics Analysis of Thinopyrum Ponticum 1158A/19
4.3. Real-Time qPCR
4.4. Fluorescence In Situ Hybridization (FISH) and Genome In Situ Hybridization (GISH)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| WWGH | wheat–wheatgrass hybrid |
| qPCR | quantitative polymerase chain reaction |
| BAC | bacterial artificial chromosome |
| FISH | fluorescence in situ hybridization |
| GISH | genomic in situ hybridization |
| mcGISH | multicolor genomic in situ hybridization |
References
- Lopes, M.; El-Basyoni, I.; Baenziger, P.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A.; et al. Exploiting genetic diversity from landraces in wheat breeding for adaptation to climate change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef]
- Liu, J.; Rasheed, A.; He, Z.; Imtiaz, M.; Arif, A.; Mahmood, T.; Ghafoor, A.; Siddiqui, S.; Ilyas, M.; Wen, W.; et al. Genome-wide variation patterns between landraces and cultivars uncover divergent selection during modern wheat breeding. Theor. Appl. Genet. 2019, 132, 2509–2523. [Google Scholar] [CrossRef] [PubMed]
- Dobrotvorskaya, T.; Martynov, S.; Pukhalskyi, V. Trends in genetic diversity change of spring bread wheat cultivars released in Russia in 1929–2003. Russ. J. Genet. 2004, 40, 1245–1257. [Google Scholar] [CrossRef]
- Petrovic, S.; Dimitrijevic, M. Genetic erosion of diversity in cereals. Genetika 2012, 44, 217–226. [Google Scholar] [CrossRef]
- Balfourier, F.; Bouchet, S.; Robert, S.; De Oliveira, R.; Rimbert, H.; Kitt, J.; Choulet, F.; Paux, E. Worldwide phylogeography and history of wheat genetic diversity. Sci. Adv. 2019, 5, eaav0536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.; Bubeck, D.; Nelson, B.; Stanek, J.; Gerke, J. Genetic diversity and modern plant breeding. In Genetic Diversity and Erosion in Plants; Springer: Cham, Switzerland, 2015; pp. 55–88. [Google Scholar]
- Frankin, S.; Kunta, S.; Abbo, S.; Sela, H.; Goldberg, B.; Bonfil, D.; Levy, A.; Avivi-Ragolsky, N.; Nashef, K.; Roychowdhury, R.; et al. The Israeli–Palestinian wheat landraces collection: Restoration and characterization of lost genetic diversity. J. Sci. Food Agric. 2019. [Google Scholar] [CrossRef]
- Wulff, B.; Moscou, M. Strategies for transferring resistance into wheat: From wide crosses to GM cassettes. Front. Plant Sci. 2014, 5, 692. [Google Scholar] [CrossRef] [Green Version]
- Motsnyj, I.; Nargan, T.; Jeryniak, M.; Lyfenko, S. Application of derivatives of incomplete wheat-wild-rye amphiploid (WWRA) Elytricum fertile in selection of winter soft wheat. Visnyk Agrar. Nauk. 2017, 95, 45–50. [Google Scholar] [CrossRef]
- Friebe, B.; Jiang, J.; Gill, B.; Dyck, P. Radiation-induced nonhomoeologous wheat-Agropyron intermedium chromosomal translocations conferring resistance to leaf rust. Theor. Appl. Genet. 1993, 86, 141–149. [Google Scholar] [CrossRef]
- Dyck, P.; Friebe, B. Evaluation of leaf rust resistance from wheat chromosomal translocation lines. Crop Sci. 1993, 33, 687–690. [Google Scholar] [CrossRef]
- Friebe, B.; Qi, L.; Wilson, D.; Chang, Z.; Seifers, D.; Martin, T.; Fritz, A.; Gill, B. Wheat-Thinopyrum intermedium recombinants resistant to wheat streak mosaic virus and Triticum mosaic virus. Crop Sci. 2009, 49, 1221–1226. [Google Scholar] [CrossRef]
- Li, H.; Wang, X. Thinopyrum ponticum and Th. intermedium: The promising source of resistance to fungal and viral diseases of wheat. J. Genet. Genom. 2009, 36, 557–565. [Google Scholar] [CrossRef]
- Nie, L.; Yang, Y.; Zhang, J.; Fu, T. Disomic chromosome addition from Thinopyrum intermedium to bread wheat appears to confer stripe rust resistance. Euphytica 2019, 215, 56. [Google Scholar] [CrossRef] [Green Version]
- Salina, E.; Adonina, I.; Badaeva, E.; Kroupin, P.; Stasyuk, A.; Leonova, I.; Shishkina, A.; Divashuk, M.; Starikova, E.; Khuat, T.; et al. A Thinopyrum intermedium chromosome in bread wheat cultivars as a source of genes conferring resistance to fungal diseases. Euphytica 2015, 204, 91–101. [Google Scholar] [CrossRef]
- Kroupin, P.; Divashuk, M.; Karlov, G. Gene resources of perennial wild cereals involved in breeding o improve wheat crop. Sel’skokhozyaistvennaya Biol. 2019, 54, 409–425. [Google Scholar] [CrossRef]
- Wang, H.; Sun, S.; Ge, W.; Zhao, L.; Hou, B.; Wang, K.; Lyu, Z.; Chen, L.; Xu, S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar] [CrossRef]
- Tang, S.; Li, Z.; Jia, X.; Larkin, P. Genomic in situ hybridization (GISH) analyses of Thinopyrum intermedium, its partial amphiploid Zhong 5, and disease-resistant derivatives in wheat. Theor. Appl. Genet. 2000, 100, 344–352. [Google Scholar] [CrossRef]
- Fedak, G.; Han, F. Characterization of derivatives from wheat-Thinopyrum wide crosses. Cytogenet. Genome Res. 2005, 109, 360–367. [Google Scholar] [CrossRef]
- Zhang, P.; Dundas, I.; Xu, S.; Friebe, B.; McIntosh, R.; Raupp, W. Chromosome engineering techniques for targeted introgression of rust resistance from wild wheat relatives. Methods Mol. Biol. 2017, 1659, 163–172. [Google Scholar]
- Li, J.; Chen, Q.; Zhang, P.; Lang, T.; Hoxha, S.; Li, G.; Yang, Z. Comparative FISH and molecular identification of new stripe rust resistant wheat-Thinopyrum intermedium ssp. trichophorum introgression lines. Crop J. 2019, 7, 819–829. [Google Scholar] [CrossRef]
- Singh, A.; Zhang, P.; Dong, C.; Li, J.; Singh, S.; Trethowan, R.; Sharp, P. Development and molecular cytogenetic characterization of Thinopyrum bessarabicum introgression lines in hexaploid and tetraploid wheats. Theor. Appl. Genet. 2020, 133, 2117–2130. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, C.; Wang, Y.; Wang, Y.; Chen, C.; Ji, W. Molecular cytogenetic identification of two wheat–Thinopyrum ponticum substitution lines conferring stripe rust resistance. Mol. Breed. 2019, 39, 143. [Google Scholar] [CrossRef]
- Turner, M.; DeHaan, L.; Jin, Y.; Anderson, J. Wheatgrass-wheat partial amphiploids as a novel source of stem rust and fusarium head blight resistance. Crop Sci. 2013, 53, 1994–2005. [Google Scholar] [CrossRef] [Green Version]
- Krupin, P.; Divashuk, M.; Belov, V.; Zhemchuzhina, A.; Kovalenko, E.; Upelniek, V.; Karlov, G. Investigation of intermediary wheat-Agropyron hybrids on resistance to leaf rust. Sel’skokhozyaistvennaya Biol. 2013, 1, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Kocheshkova, A.; Kroupin, P.; Bazhenov, M.; Karlov, G.; Pochtovyy, A.; Upelniek, V.; Belov, V.; Divashuk, M. Pre-harvest sprouting resistance and haplotype variation of THVP-1 gene in the collection of wheat-wheatgrass hybrids. PLoS ONE 2017, 12, e0188049. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Ren, Y.; Murray, T.; Yan, W.; Guo, Q.; Niu, Y.; Sun, Y.; Li, H. Development of perennial wheat through hybridization between wheat and wheatgrasses: A Review. Engineering 2018, 4, 507–513. [Google Scholar] [CrossRef]
- Kroupin, P.; Divashuk, M.; Belov, V.; Glukhova, L.; Aleksandrov, O.; Karlov, G. Comparative molecular cytogenetic characterization of partial wheat-wheatgrass hybrids. Russ. J. Genet. 2011, 47, 432–437. [Google Scholar] [CrossRef]
- Trifonova, A.; Boris, K.; Dedova, L.; Melnik, V.; Ivanova, L.; Kuzmina, N.; Zavgorodniy, S.; Upelniek, V. genome polymorphism of the synthetic species X Trititrigia cziczinii Tsvel. inferred from AFLP analysis. Vavilov J. Genet. Breed. 2018, 22, 648–653. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, Y.; Qi, J.; Wang, H.; Wang, R.; Bao, Y.; Li, X. Identification of chromosomes in Thinopyrum intermedium and wheat-Th.intermedium amphiploids based on multiplex oligonucleotide probes. Genome 2018, 61, 515–521. [Google Scholar] [CrossRef]
- Chen, Q.; Conner, R.L.; Laroche, A.; Ahmad, F. Molecular cytogenetic evidence for a high level of chromosome pairing among different genomes in Triticum aestivum–Thinopyrum intermedium hybrids. Theor. Appl. Genet. 2001, 102, 847–852. [Google Scholar] [CrossRef]
- Brasileiro-Vidal, A.; Cuadrado, A.; Brammer, S.; Benko-Iseppon, A.; Guerra, M. Molecular cytogenetic characterization of parental genomes in the partial amphidiploid Triticum aestivum x Thinopyrum ponticum. Genet. Mol. Biol. 2005, 28, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Piaskowski, J.; Murphy, K.; Kisha, T.; Jones, S. Perennial wheat lines have highly admixed population structure and elevated rates of outcrossing. Euphytica 2017, 213, 171. [Google Scholar] [CrossRef]
- Curwen-McAdams, C.; Arterburn, M.; Murphy, K.; Cai, X.; Jones, S. Toward a taxonomic definition of perennial wheat: A new species ×Tritipyrum aaseae described. Genet. Resour. Crop Evol. 2016, 64, 1651–1659. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Xing, P.; Bao, Y.; Ren, M.; Liu, S.; Wang, Y.; Li, X.; Wang, H. Chromosome pairing in hybrid progeny between Triticum aestivum and Elytrigia elongata. Front. Plant Sci. 2017, 8, 2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Hao, W.; Tang, C.; Yao, H.; Li, B.; Zheng, Q.; Li, Z.; Zhang, X. Plasticity in Triticeae centromere DNA sequences: A wheat × tall wheatgrass (decaploid) model. Plant J. 2019, 100, 314–327. [Google Scholar] [CrossRef]
- Georgieva, M.; Sepsi, A.; Tyankova, N.; Molnár-Láng, M. Molecular cytogenetic characterization of two high protein wheat-Thinopyrum intermedium partial amphiploids. J. Appl. Genet. 2011, 52, 269–277. [Google Scholar] [CrossRef]
- Kruppa, K.; Türkösi, E.; Mayer, M.; Tóth, V.; Vida, G.; Szakács, É.; Molnár-Láng, M. Mcgish identification and phenotypic description of leaf rust and yellow rust resistant partial amphiploids originating from a wheat × Thinopyrum synthetic hybrid cross. J. Appl. Genet. 2016, 57, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, M.; Kruppa, K.; Tyankova, N.; MOLNÁR-LÁNG, M. Molecular cytogenetic identification of a novel hexaploid wheat–Thinopyrum intermedium partial amphiploid with high protein content. Turk. J. Biol. 2016, 40, 554–560. [Google Scholar] [CrossRef]
- Liu, L.; Luo, Q.; Teng, W.; Li, B.; Li, H.; Li, Y.; Li, Z.; Zheng, Q. Development of Thinopyrum ponticum-specific molecular markers and FISH probes based on SLAF-seq technology. Planta 2018, 247, 1099–1108. [Google Scholar] [CrossRef]
- Cseh, A.; Yang, C.; Hubbart-Edwards, S.; Scholefield, D.; Ashling, S.S.; Burridge, A.J.; Wilkinson, P.A.; King, I.P.; King, J.; Grewal, S. Development and validation of an exome-based SNP marker set for identification of the St, J r and J vs. genomes of Thinopyrym intermedium in a wheat background. Theor. Appl. Genet. 2019, 132, 1555–1570. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Wang, H.; Xu, Y.; Li, Y.; Lang, T.; Yang, Z.; Li, G. Characterization of chromosomal rearrangement in new wheat—Thinopyrum intermedium addition lines carrying Thinopyrum—Specific grain hardness genes. Agronomy 2019, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Kroupin, P.; Kuznetsova, V.; Romanov, D.; Kocheshkova, A.; Karlov, G.; Dang, T.; Khuat, T.; Kirov, I.; Alexandrov, O.; Polkhovskiy, A.; et al. Pipeline for the rapid development of cytogenetic markers using genomic data of related species. Genes 2019, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- Kroupin, P.; Kuznetsova, V.; Nikitina, E.; Martirosyan, Y.; Karlov, G.; Divashuk, M. Development of new cytogenetic markers for Thinopyrum ponticum (Podp.) Z.-W. Liu &Amp; R.-C. Wang. Comp. Cytogenet. 2019, 13, 231–243. [Google Scholar]
- Novák, P.; Neumann, P.; Macas, J. Graph-based clustering and characterization of repetitive sequences in next-generation sequencing data. BMC Bioinform. 2010, 11, 378. [Google Scholar] [CrossRef] [Green Version]
- Salina, E.; Adonina, I.; Vatolina, T.; Kurata, N. A Comparative analysis of the composition and organization of two subtelomeric repeat families in Aegilops speltoides Tausch. and related species. Genetica 2004, 122, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Pollak, Y.; Zelinger, E.; Raskina, O. Repetitive DNA in the architecture, repatterning, and diversification of the genome of Aegilops speltoides Tausch (Poaceae, Triticeae). Front. Plant Sci. 2018, 9, 1779. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; La, S.; Li, B.; Yu, Z.; Chen, Q.; Li, J.; Yang, E.; Li, G.; Yang, Z. Precise identification of wheat—Thinopyrum intermedium translocation chromosomes carrying resistance to wheat stripe rust in line Z4 and its derived progenies. Genome 2018, 61, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Xi, W.; Tang, Z.; Tang, S.; Yang, Z.; Luo, J.; Fu, S. New ND-FISH-positive oligo probes for identifying Thinopyrum chromosomes in wheat backgrounds. Int. J. Mol. Sci. 2019, 20, 2031. [Google Scholar] [CrossRef] [Green Version]
- Baruch, O.; Kashkush, K. Analysis of copy-number variation, insertional polymorphism, and methylation status of the tiniest Class I (TRIM) and Class II (MITE) transposable element families in various rice strains. Plant Cell Rep. 2011, 31, 885–893. [Google Scholar] [CrossRef]
- Said, M.; Hřibová, E.; Danilova, T.; Karafiátová, M.; Čížková, J.; Friebe, B.; Doležel, J.; Gill, B.; Vrána, J. The Agropyron cristatum karyotype, chromosome structure and cross-genome homoeology as revealed by fluorescence in situ hybridization with tandem repeats and wheat single-gene probes. Theor. Appl. Genet. 2018, 131, 2213–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Wang, H.; Lang, T.; Li, J.; La, S.; Yang, E.; Yang, Z. New molecular markers and cytogenetic probes enable chromosome identification of wheat-Thinopyrum intermedium introgression lines for improving protein and gluten contents. Planta 2016, 244, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Luo, Q.; Li, H.; Li, B.; Li, Z.; Zheng, Q. Physical mapping of the blue-grained gene from Thinopyrum ponticum chromosome 4Ag and development of blue-grain-related molecular markers and a FISH probe based on SLAF-Seq technology. Theor. Appl. Genet. 2018, 131, 2359–2370. [Google Scholar] [CrossRef]
- Upelniek, V.; Belov, V.; Ivanova, l.; Dolgova, S.; Demidov, A. Heritage of academician N.V. Tsitsin: State-of-the-art and potential of the collection of intermediate wheat × couch-grass hybrids. Vavilov J. Genet. Breed. Vavilovskij Žurnal Genet. I Sel. 2020, 6, 85–89. (In Russian) [Google Scholar]
- Divashuk, M.; Krupin, P.; Bazhenov, M.; Klimushina, M.; Belov, V.; Semyonova, E.; Karlov, G. Molecular-genetic characterization of seed storage protein composition in partial wheat-wheatgrass hybrids Izv. TSKHA 2012, 5, 29–37. (In Russian) [Google Scholar]
- Krupin, P.; Divashuk, M.; Bazhenov, M.; Gritsenko, L.; Tarakanov, I.; Upelniek, V.; Belov, V.; Post, A.; Starikova, E.; Khuat Thi Mai, L.; et al. Polymorphism of the reaction of wheat-wheatgrass hybrids to salinity. Agric. Biol. 2013, 5, 44–53. [Google Scholar]
- Rogers, S.; Bendich, A. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol. 1985, 5, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Novák, P.; Ávila Robledillo, L.; Koblížková, A.; Vrbová, I.; Neumann, P.; Macas, J. TAREAN: A computational tool for identification and characterization of satellite DNA from unassembled short reads. Nucleic Acids Res. 2017, 45, e111. [Google Scholar] [CrossRef]
- Divashuk, M.; Khuat, T.; Kroupin, P.; Kirov, I.; Romanov, D.; Kiseleva, A.; Khrustaleva, L.; Alexeev, D.; Zelenin, A.; Klimushina, M.; et al. Variation in copy number of Ty3/Gypsy centromeric retrotransposons in the genomes of Thinopyrum intermedium and its diploid progenitors. PLoS ONE 2016, 11, e0154241. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsova, V.; Razumova, O.; Karlov, G.; Dang, T.; Kroupin, P.; Divashuk, M. Some peculiarities in application of denaturating and non-denaturating in situ hybridization on chromosomes of cereals. Mosc. Univ. Biol. Sci. Bull. 2019, 74, 75–80. [Google Scholar] [CrossRef]
- Komuro, S.; Endo, R.; Shikata, K.; Kato, A. Genomic and chromosomal distribution patterns of various repeated DNA sequences in wheat revealed by a fluorescence in situ hybridization procedure. Genome 2013, 56, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Kishii, M.; Wang, R.; Tsujimoto, H. GISH analysis revealed new aspect of genomic constitution of Thinopyrum intermedium. Czech J. Genet. Plant Breed. 2012, 41, 92–95. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster Name | Proportion, % | Length, bp | Consensus Cluster Sequence | Homology to the Known Repeats |
|---|---|---|---|---|
| Putative satellites (high confidence) | ||||
| 19-202 | 0.034 | 380 | GATTTTACATACGTGCACACACAGGATCACATGCGCGGAAAATATCGAGCCCAAAAAGGGCCGTCTGGGCCCTCAAAAATGGCCTGCAGGATTGGAAAAAATGAAAGTTATCGCAATTACAGCTCAAATTTCGATGAGCCGGCACATCCCTTTGGTGTTCAGGTCCTGGGCGCCCCACCCATGTATGGATACAATAGGGTCTTAGGCCAACTCTCGCAAAGAAACGGTGATCAAAGAAAATACAAAAATCAACCTAGAGTCTGAATTGATCGAGACTCTCAAAGCAAGTGAGAACAAGCTCCGATTTCATCGTTTTCATAGGCGTTGGAAAGCTATTGGGGAATTGAGCTGATAGGACTTCTAGTTTTTGTGATTTGGAG | MG323511.1 Agropyron cristatum clone ACRI_TR_CL20 satellite sequence sequence |
| KP231286.1 Agropyron cristatum isolate Z559 TRT1 repeat sequence sequence | ||||
| 17-251 | 0.013 | 376 | GATGAAAAACAAAAGTTTTGGCAGTTAAAGTTTGAGATTCGATGGTGCCGTAGATCCGTTTGGTCATCAGGTGTTACGAGCCACACCCATGGATAGCTAGAATAGTGTGCTAGGCTCAGACTAGAGAAGAAACGGGGAACAAACTGATGAAAGAATTATCATACCGAGGGAATTCACTCAGAACTAACAAGAATGATAACGTGCTCCGACTGGAGCGGTTTAGGAACTGCTGGAAGGCTCTAGGGGAATAAAGATGGAATGACTTCTAATTTTTAGGTCCGTATGATTTGAAATACGTGCACACAAACCCCGAGAAAGCGTTCCTGGAAATCTCGGCCCAAAAAAGGACGTTTAGTAATTCTAAAAGGGACTACAG | AY117401.1 Aegilops speltoides subtelomeric repeat Spelt52.2 sequence |
| Putative satellites (low confidence) | ||||
| 18-158 | 0.097 | 333 | GCCACACCCCGGAGACGCGTGCCGCCTCTTCGCACATGCCACCACACTTCCAGACATGTATGAGGGCCCGGTGCGACGCTCCGGTGGCATTGCTACCCCCCAGGGCCCCCGTCCCGCCTAACCCTGGAACGGTTGACCACGAGATCTAGCCCTTTGACTTTCGCCGGACGGGCTTTGACCAGTGGACCTTTCCACCTGGTTGTGATAGGTCAGCCCATAGGAACACCTCGGAGCAACGTCCGGGCCAAACCCACAGCAAGATTCCCTCCGTGTCGACCCGACGCGTCCGTTTCCCCCCTCCAGGTGCCGGCGGCGGCGCCGTCCGTGAGGGGG | KC290905.1 Triticum aestivum clone pTa-465 FISH-positive repetitive sequence |
| 17-172 | 0.08 | 662 | ATTGGAAAACCTTCGCATTGTGTCATTATATGTGACCAAGTTACCAGGAAAAATAATAAACTTGTAATACGGCAATTATTTTAAAAAAGTGTTCTCAGAAATGAGCTATCATGTGTGAAGATTCATGGCTTTCAAGCCAAATGATCAATCTTATGGCCACATTCATGGCATAGTTTGTTCAAATGATCTCATATTGTGCACAAGGGTGCATCTTGGAATGGCAAACAATGTTGCCTAAGGAAGTTTTCATTTTCTTTGGACGAAAAATTCATTTTCCATTTTTCGAGTGCCCAAAATGAGTTTTTTTGTGAAGGACCTACCATATATTTGTTGCAAAATTGGACCAAATCAATTTTCTAAAATATTAGGCCATATTTAATGCACAATTGACCAAATGGTTGGGTGTCAAAAGTTTTGATCCACCTCTCGTGAAAAAGACAAATTTCCGCCGATTCAGTAGGAAGCGGGTCAAATTTGAACTGCAGCTGCCTCATAGTTTGCTCTTTATTTTTTCCAAAAATCATTTCTAGGTACATAAGTATCTATTTAATCAGAGAAACACCAAAAGTTTTCCAAGATTCAACCACTAGCTAGGAACGGTCATGCCCGCCGTTTTGACCGCATTTTGAAACGGGCATAAAAAATTCAAAAAAAATCAAAAA | MG323513.1 Agropyron cristatum clone ACRI_TR_CL80 satellite sequence |
| 17-202 | 0.032 | 553 | TTTGTAATGGAAGGATGGTGCATTGTTCTATATGTTATTGTCCATATATCAGTCCGTAGGTGAGCTCACGGAAGGGTGGTAGAGGGTGGCAGAGTATACTTCAAACATAAAATCATCCGAAACTCAATTTTACAAGCCGGATCTTGCCTCCGAAATGTTGTCGAAGCCGGCGAGTGGGTTACGGACGCATACAACTTTTCGTTGTGATCGTTTTGGCGGGTCATGGAGCTCCAATGGAGTTTTTATGGCCAAATTGTGGCCGTTTTATGGATACAACATCGCGGGACAGACCGTGAATACAACTTTCAGGGTAAGTTGATCGCACCGACGAGCCATCTTGCACCATTCGGAATGACCTATAATTTTTCGTGTGCATAAACAGAATGAGGATGAGCTGTTATGTACTTTATGATCCAAGAATAATGCATCCGCTGGTGAAAACGTGAGGGTGGTAGCGGGATGGGTGATAGACCATGTAAAGCATGAATTCTTGGGTTTCGATGCAACGAAGAGCCTCCTCCTTCTGTCGTGACTGAACCTGTAGTCTTTCTAG | MN161206.1 Triticum aestivum clone CentT550 satellite sequence |
| KT724936.1 Secale cereale clone BAC 19H13l pSc250 retrotransposon TREP231, complete sequence; and satellite pSc250 sequence | ||||
| Putative LTR elements | ||||
| 17-62 | 0.48 | 534 | TCTCAAAATTTCGTTTCCCGCCCAAAGTTTCGTCTCCCGCTCGAAATATCGTCTCGCGCCCGTTCATCATATTTTTATCCCTCCCTCCCATTTTTGTATTTATTATTTATTTATTTTCCGGGGAGACGCGGTGGCAATGGTGGACAACAACACACCCTACTTTGGCATGACGGCGAAATTTTCCCGCCCAAAATGACGAAAAAATGACAACGGCCGCGAGTGTCTCAAAGCGCTCCCGGAGGTGTAAAAACGGGTATAGGAACGTATCACGGAGTTTGGTGGCCCCCAATCCCGGGATGGTGTCGAAAATTAGGGGCAATACGGGCATTACTTTTGTACCGGGCAACGTAGGATGGCCTCGGGGATGACAACGCAACCGGGGCAAAGGGGCTCCGACCCGGGCGGCCACGGGCCCGTCGGAGAGGCCTCGTGGAGACGGCGACCCGGGAAGACTACCTTCCGCGTCCCGGGCGCGTCCGCACTACGTGCCCTCTCAAGCAAAACCGCAAACGACCGCATCTCTCTCTCTCTCTC | MG323514.1 Agropyron cristatum clone ACRI_TR_CL85 satellite sequence |
| Tandem repeat | Primer sequence |
|---|---|
| 17-62 | F: 5′-TTGCCCCTATTTTTCGACAC-3′ |
| R: 5′-GTGGCAATGGTGAACAACAA-3′ | |
| 17-251 | F: 5′-CAGTTCCTAAACCGCTCCAG-3′ |
| R: 5′-AGATCCGTTTGGTCATCAGG-3′ | |
| 17-202 | F: 5′-TCTATCACCCATCCCGCTAC-3′ |
| R: 5′-AATTGTGGCCGTTTTATGGA-3′ | |
| 19-202_1 | F: 5′-CAGCTCAAATTTCGATGAGC-3′ |
| R: 5′-TTCCAACGCCTATGAAAACG-3′ | |
| 19-202_2 | F: 5′-CATCCCTTTGGTGTTCAGGT-3′ |
| R: 5′-CCAACGCCTATGAAAACGAT-3′ | |
| 17-172 | F:5′-TGCAAAATTGGACCAAATCA-3′ |
| R:5′-GAGGCAGCTGCAGTTCAAAT-3′ | |
| 18-158 | F:5′-GGAAAGGTCCACTGGTCAAA-3′ |
| R:5′-ACATGCCACAACACTTCCAC-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikitina, E.; Kuznetsova, V.; Kroupin, P.; Karlov, G.I.; Divashuk, M.G. Development of Specific Thinopyrum Cytogenetic Markers for Wheat-Wheatgrass Hybrids Using Sequencing and qPCR Data. Int. J. Mol. Sci. 2020, 21, 4495. https://doi.org/10.3390/ijms21124495
Nikitina E, Kuznetsova V, Kroupin P, Karlov GI, Divashuk MG. Development of Specific Thinopyrum Cytogenetic Markers for Wheat-Wheatgrass Hybrids Using Sequencing and qPCR Data. International Journal of Molecular Sciences. 2020; 21(12):4495. https://doi.org/10.3390/ijms21124495
Chicago/Turabian StyleNikitina, Ekaterina, Victoria Kuznetsova, Pavel Kroupin, Gennady I. Karlov, and Mikhail G. Divashuk. 2020. "Development of Specific Thinopyrum Cytogenetic Markers for Wheat-Wheatgrass Hybrids Using Sequencing and qPCR Data" International Journal of Molecular Sciences 21, no. 12: 4495. https://doi.org/10.3390/ijms21124495
APA StyleNikitina, E., Kuznetsova, V., Kroupin, P., Karlov, G. I., & Divashuk, M. G. (2020). Development of Specific Thinopyrum Cytogenetic Markers for Wheat-Wheatgrass Hybrids Using Sequencing and qPCR Data. International Journal of Molecular Sciences, 21(12), 4495. https://doi.org/10.3390/ijms21124495