Ethylene Is Not Essential for R-Gene Mediated Resistance but Negatively Regulates Moderate Resistance to Some Aphids in Medicago truncatula

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
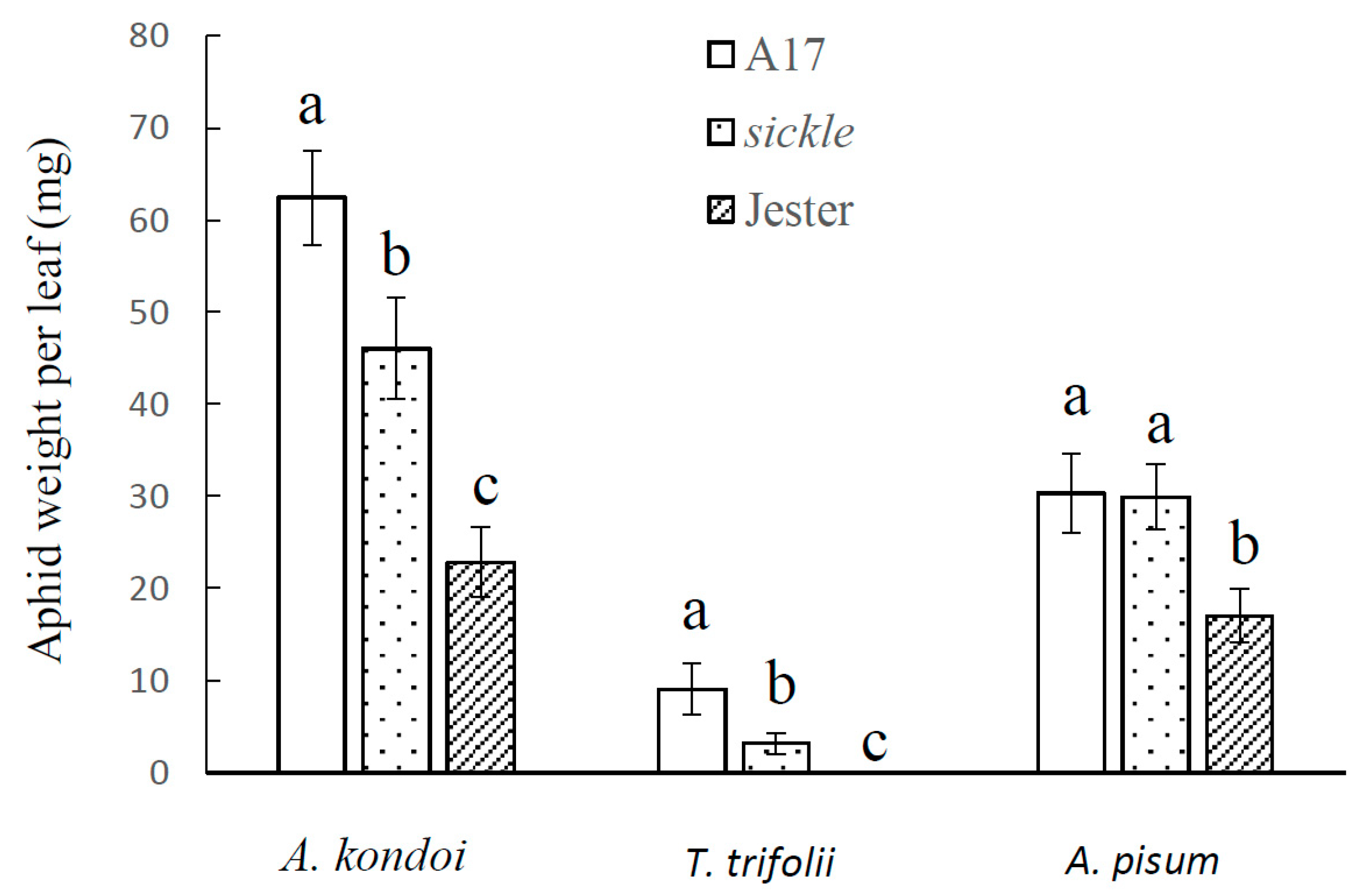
2.1. The M. truncatula Sickle Mutant Modulates Aphid Resistance
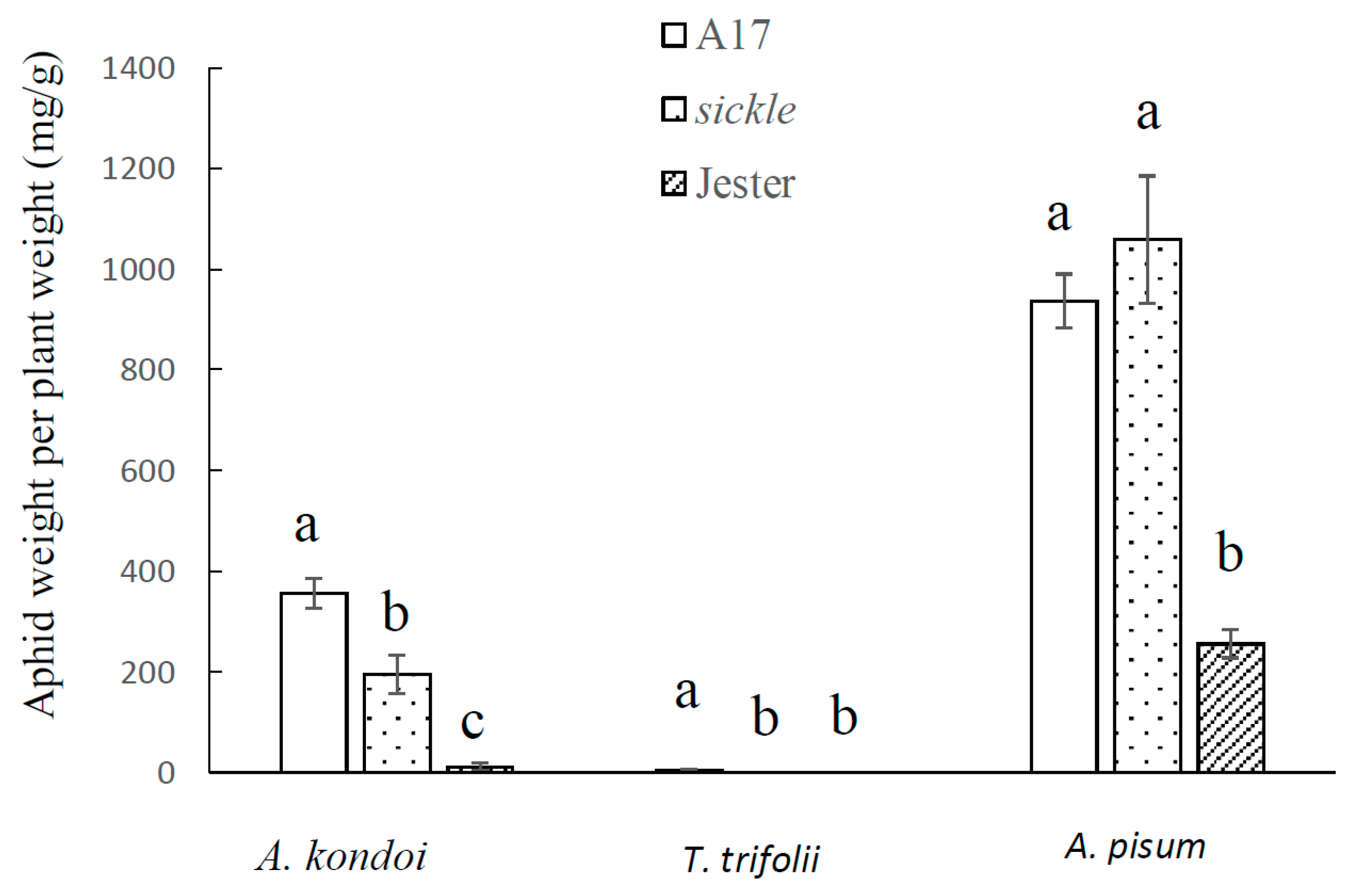
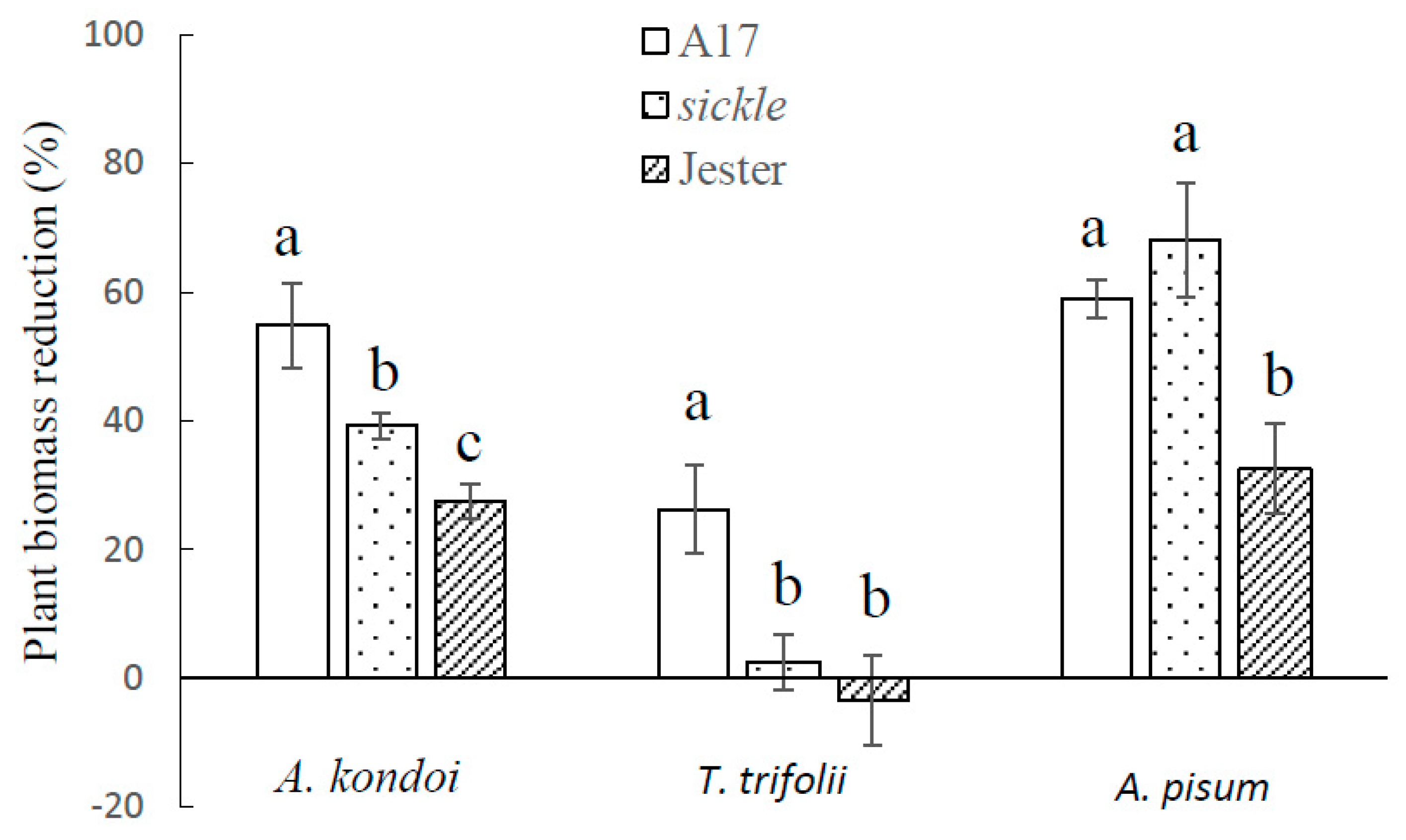
2.2. Effect of M. truncatula Sickle Mutant on Plant Tolerance to Aphid Feeding
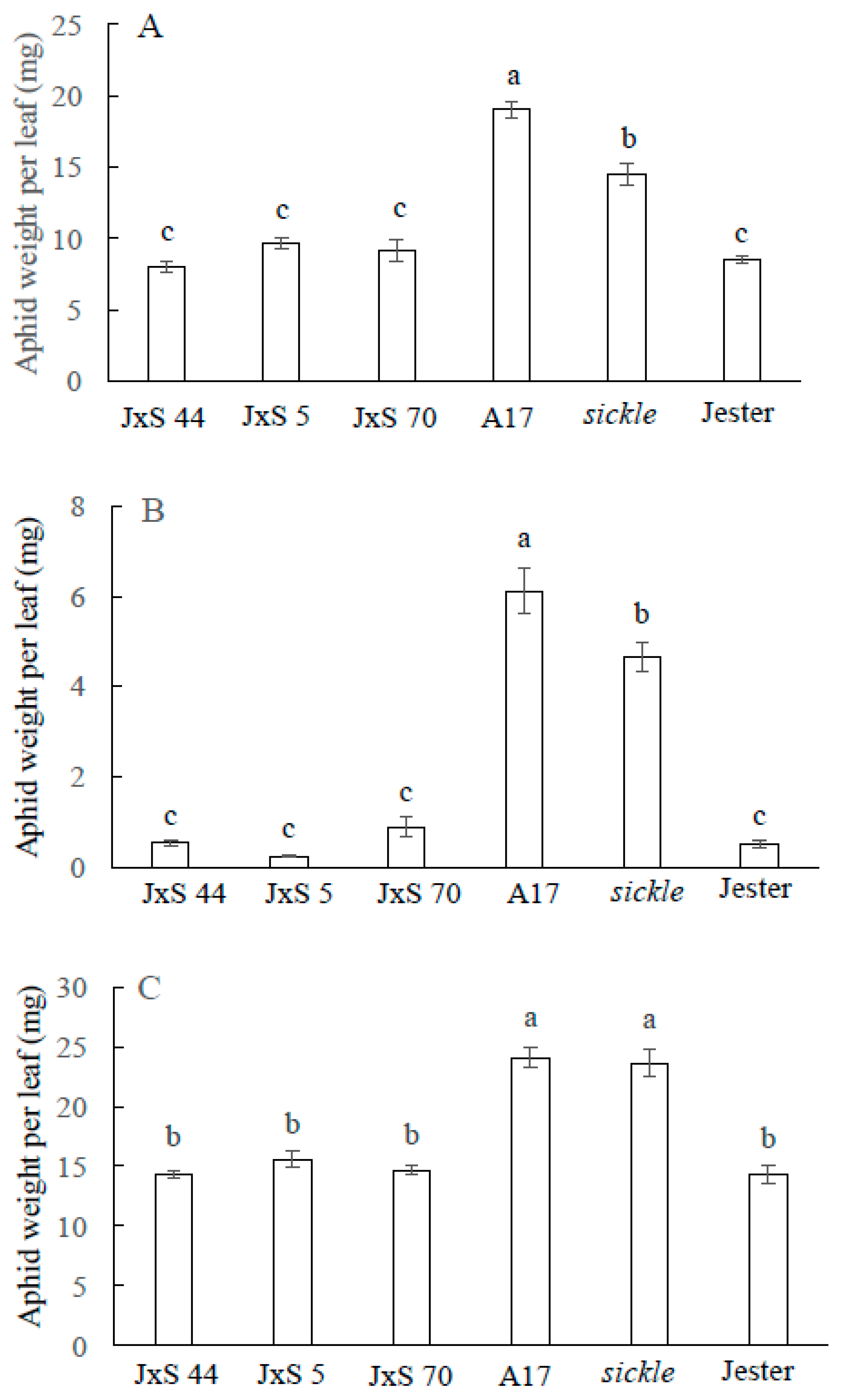
2.3. Effect of M. truncatula Sickle Mutant on R Gene Mediated Resistance to Aphids
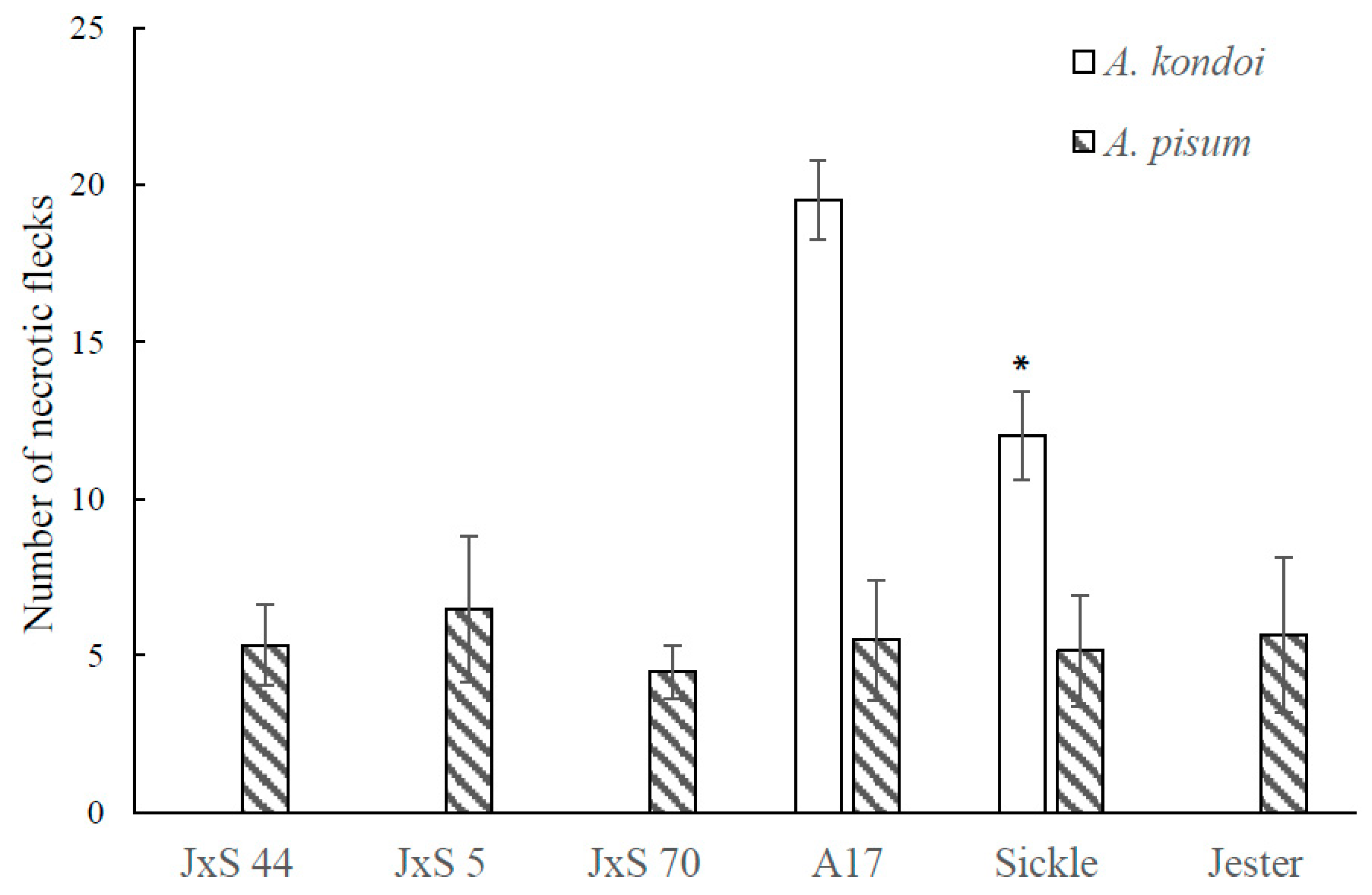
2.4. The Role of Ethylene Insensitivity in the AIN-Mediated Hypersensitive Response to A. kondoi and A. pisum Infestation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Aphid Species and Rearing Conditions
4.3. Screening for F2 Plants Containing the Sickle Homozygous Allele
4.4. Genotyping F2 Plants with AKR, APR and TTR Loci
4.5. Aphid Performance and Plant Damage on Single Trifoliate Leaves
4.6. Aphid Performance and Plant Tolerance Experiments on Whole Plants
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ng, J.C.K.; Perry, K.L. Transmission of plant viruses by aphid vectors. Mol. Plant Pathol. 2004, 5, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect vector-mediated transmission of plant viruses. Virology 2015, 479–480, 278–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, J.P.; Murphy, A.M.; Tungadi, T.; Yoon, J.Y. Plant defense signals: Players and pawns in plant-virus-vector interactions. Plant Sci. 2019, 279, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Meldau, S.; Howe, G.A. Role of phytohormones in insect-specific plant reactions. Trends Plant Sci. 2012, 17, 250–259. [Google Scholar] [CrossRef] [Green Version]
- Divol, F.; Vilaine, F.; Thibivilliers, S.; Amselem, J.; Palauqui, J.C.; Kusiak, C.; Dinant, S. Systemic response to aphid infestation by Myzus persicae in the phloem of Apium graveolens. Plant Mol. Biol. 2005, 57, 517–540. [Google Scholar] [CrossRef]
- Moran, P.J.; Cheng, Y.F.; Cassell, J.L.; Thompson, G.A. Gene expression profiling of Arabidopsis thaliana in compatible plant-aphid interactions. Arch. Insect Biochem. Physiol. 2002, 51, 182–203. [Google Scholar] [CrossRef]
- Heidel, A.J.; Baldwin, I.T. Microarray analysis of salicylic acid- and jasmonic acid-signalling in responses of Nicotiana attenuata to attack by insects from multiple feeding guilds. Plant Cell Environ. 2004, 27, 1362–1373. [Google Scholar] [CrossRef]
- De Vos, M.; Van Oosten, V.R.; Van Poecke, R.M.P.; Van Pelt, J.A.; Pozo, M.J.; Mueller, M.J.; Buchala, A.J.; Metraux, J.P.; Van Loon, L.C.; Dicke, M.; et al. Signal signature and transcriptome changes of Arabidopsis during pathogen and insect attack. Mol. Plant-Microbe Interact. 2005, 18, 923–937. [Google Scholar] [CrossRef] [Green Version]
- Kettles, G.J.; Drurey, C.; Schoonbeek, H.-j.; Maule, A.J.; Hogenhout, S.A. Resistance of Arabidopsis thaliana to the green peach aphid, Myzus persicae, involves camalexin and is regulated by microRNAs. New Phytol. 2013, 198, 1178–1190. [Google Scholar] [CrossRef] [Green Version]
- Mewis, I.; Tokuhisa, J.G.; Schultz, J.C.; Appel, H.M.; Ulrichs, C.; Gershenzon, J. Gene expression and glucosinolate accumulation in Arabidopsis thaliana in response to generalist and specialist herbivores of different feeding guilds and the role of defense signaling pathways. Phytochemistry 2006, 67, 2450–2462. [Google Scholar] [CrossRef]
- Miller, H.L.; Neese, P.A.; Ketring, D.L.; Dillwith, J.W. Involvement of ethylene in aphid infestation of barley. J. Plant Growth Regul. 1994, 13, 167–171. [Google Scholar] [CrossRef]
- Argandona, V.H.; Chaman, M.; Cardemil, L.; Munoz, O.; Zuniga, G.E.; Corcuera, L.J. Ethylene production and peroxidase activity in aphid-infested barley. J. Chem. Ecol. 2001, 27, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Boyko, E.V.; Smith, C.M.; Thara, V.K.; Bruno, J.M.; Deng, Y.P.; Starkey, S.R.; Klaahsen, D.L. Molecular basis of plant gene expression during aphid invasion: Wheat Pto- and Pti-like sequences are involved in interactions between wheat and Russian wheat aphid (Homoptera: Aphididae). J. Econon. Entomol. 2006, 99, 1430–1445. [Google Scholar] [CrossRef] [PubMed]
- Anstead, J.; Samuel, P.; Song, N.; Wu, C.J.; Thompson, G.A.; Goggin, F. Activation of ethylene-related genes in response to aphid feeding on resistant and susceptible melon and tomato plants. Entomol. Exp. Appl. 2010, 134, 170–181. [Google Scholar] [CrossRef]
- Kamphuis, L.; Gao, L.; Turnbull, C.; Karam, S. Medicago—Aphid interactions. In The Model Legume Medicago Truncatula; de Bruijn, F.J., Ed.; Wiley & Sons, Inc.: New York, NY, USA, 2019; pp. 363–368. [Google Scholar]
- Kamphuis, L.G.; Zulak, K.; Gao, L.-L.; Anderson, J.; Singh, K.B. Plant–aphid interactions with a focus on legumes. Funct. Plant Biol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.L.; Klingler, J.P.; Anderson, J.P.; Edwards, O.R.; Singh, K.B. Characterization of pea aphid resistance in Medicago truncatula. Plant Physiol. 2008, 146, 996–1009. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.M.; Kamphuis, L.G.; Gao, L.L.; Klingler, J.P.; Lichtenzveig, J.; Edwards, O.; Singh, K.B. Identification of distinct quantitative trait loci associated with defence against the closely related aphids Acyrthosiphon pisum and A. kondoi in Medicago truncatula. J. Exp. Bot. 2012, 63, 3913–3922. [Google Scholar] [CrossRef] [Green Version]
- Kamphuis, L.G.; Guo, S.M.; Gao, L.L.; Singh, K.B. Genetic Mapping of a Major Resistance Gene to Pea Aphid (Acyrthosipon pisum) in the Model Legume Medicago truncatula. Int. J. Mol. Sci. 2016, 17, 1224. [Google Scholar] [CrossRef] [Green Version]
- Kamphuis, L.G.; Lichtenzveig, J.; Peng, K.F.; Guo, S.M.; Klingler, J.P.; Siddique, K.H.M.; Gao, L.L.; Singh, K.B. Characterization and genetic dissection of resistance to spotted alfalfa aphid (Therioaphis trifolii) in Medicago truncatula. J. Exp. Bot. 2013, 64, 5157–5172. [Google Scholar] [CrossRef]
- Klingler, J.; Creasy, R.; Gao, L.L.; Nair, R.M.; Calix, A.S.; Jacob, H.S.; Edwards, O.R.; Singh, K.B. Aphid resistance in Medicago truncatula involves antixenosis and phloem-specific, inducible antibiosis, and maps to a single locus flanked by NBS-LRR resistance gene analogs. Plant Physiol. 2005, 137, 1445–1455. [Google Scholar] [CrossRef] [Green Version]
- Klingler, J.P.; Edwards, O.R.; Singh, K.B. Independent action and contrasting phenotypes of resistance genes against spotted alfalfa aphid and bluegreen aphid in Medicago truncatula. New Phytol. 2007, 173, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Klingler, J.P.; Nair, R.M.; Edwards, O.R.; Singh, K.B. A single gene, AIN, in Medicago truncatula mediates a hypersensitive response to both bluegreen aphid and pea aphid, but confers resistance only to bluegreen aphid. J. Exp. Bot. 2009, 60, 4115–4127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, S.A.; Hodge, S.; Ismail, N.; Mansfield, J.W.; Feys, B.J.; Prosperi, J.M.; Huguet, T.; Ben, C.; Gentzbittel, L.; Powell, G. The RAP1 Gene Confers Effective, Race-Specific Resistance to the Pea Aphid in Medicago truncatula Independent of the Hypersensitive Reaction. Mol. Plant-Microbe Interact. 2009, 22, 1645–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamphuis, L.G.; Klingler, J.P.; Jacques, S.; Gao, L.-L.; Edwards, O.R.; Singh, K.B. Additive and epistatic interactions between AKR and AIN loci conferring bluegreen aphid resistance and hypersensitivity in Medicago truncatula. J. Exp. Bot. 2019, 70, 4887–4902. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.L.; Anderson, J.P.; Klingler, J.P.; Nair, R.M.; Edwards, O.R.; Singh, K.B. Involvement of the octadecanoid pathway in bluegreen aphid resistance in Medicago truncatula. Mol. Plant-Microbe Interact. 2007, 20, 82–93. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.L.; Kamphuis, L.G.; Kakar, K.; Edwards, O.R.; Udvardi, M.K.; Singh, K.B. Identification of potential early regulators of aphid resistance in Medicago truncatula via transcription factor expression profiling. New Phytol. 2010, 186, 980–994. [Google Scholar] [CrossRef]
- Stewart, S.A.; Hodge, S.; Bennett, M.; Mansfield, J.W.; Powell, G. Aphid induction of phytohormones in Medicago truncatula is dependent upon time post-infestation, aphid density and the genotypes of both plant and insect. Arthropod-Plant Interact. 2016, 10, 41–53. [Google Scholar] [CrossRef]
- Penmetsa, R.V.; Cook, D.R. A legume ethylene-insensitive mutant hyperinfected by its rhizobial symbiont. Science 1997, 275, 527–530. [Google Scholar] [CrossRef]
- Penmetsa, R.V.; Uribe, P.; Anderson, J.; Lichtenzveig, J.; Gish, J.C.; Nam, Y.W.; Engstrom, E.; Xu, K.; Sckisel, G.; Pereira, M.; et al. The Medicago truncatula ortholog of Arabidopsis EIN2, sickle, is a negative regulator of symbiotic and pathogenic microbial associations. Plant J. 2008, 55, 580–595. [Google Scholar] [CrossRef]
- Gao, L.L.; Horbury, R.; Nair, R.M.; Singh, K.B.; Edwards, O.R. Characterization of resistance to multiple aphid species (Hemiptera: Aphididae) in Medicago truncatula. Bull. Entomol. Res. 2007, 97, 41–48. [Google Scholar] [CrossRef]
- Guo, S.M.; Kamphuis, L.G.; Gao, L.L.; Edwards, O.R.; Singh, K.B. Two independent resistance genes in the Medicago truncatula cultivar jester confer resistance to two different aphid species of the genus Acyrthosiphon. Plant Signal. Behav. 2009, 4, 328–331. [Google Scholar] [CrossRef] [Green Version]
- Klingler, J.; Creasy, R.; Gao, L.L.; Nair, R.M.; Jacob, H.S.; Edwards, O.R.; Singh, K.B. Genetics of phloem-aphid interactions in Medicago truncatula. Comp. Biochem. Physiol. A-Mol. Integr. Physiol. 2005, 141, S230. [Google Scholar]
- Lu, J.; Li, J.; Ju, H.; Liu, X.; Erb, M.; Wang, X.; Lou, Y. Contrasting Effects of Ethylene Biosynthesis on Induced Plant Resistance against a Chewing and a Piercing-Sucking Herbivore in Rice. Mol. Plant 2014, 7, 1670–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mewis, I.; Appel, H.M.; Hom, A.; Raina, R.; Schultz, J.C. Major signaling pathways modulate Arabidopsis glucosinolate accumulation and response to both phloem-feeding and chewing insects. Plant Physiol. 2005, 138, 1149–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Guo, J.; Ma, Z.; Wang, J.; Zhou, C. Arabidopsis Transcription Factor MYB102 Increases Plant Susceptibility to Aphids by Substantial Activation of Ethylene Biosynthesis. Biomolecules 2018, 8, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantelin, S.; Bhattarai, K.K.; Kaloshian, I. Ethylene contributes to potato aphid susceptibility in a compatible tomato host. New Phytol. 2009, 183, 444–456. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.J.; Sun, Y.C.; Li, Y.F.; Liu, X.H.; Zhang, W.H.; Ge, F. Elevated CO2 decreases the response of the ethylene signaling pathway in Medicago truncatula and increases the abundance of the pea aphid. New Phytol. 2014, 201, 279–291. [Google Scholar] [CrossRef]
- Sun, Y.; Guo, H.; Zhu-Salzman, K.; Ge, F. Elevated CO2 increases the abundance of the peach aphid on Arabidopsis by reducing jasmonic acid defenses. Plant Sci. 2013, 210, 128–140. [Google Scholar] [CrossRef]
- Sun, Y.C.; Guo, H.J.; Ge, F. Plant-Aphid Interactions Under Elevated CO2: Some Cues from Aphid Feeding Behavior. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Buscaill, P.; Rivas, S. Transcriptional control of plant defence responses. Curr. Opin. Plant Biol. 2014, 20, 35–46. [Google Scholar] [CrossRef]
- Faulkner, C.; Robatzek, S. Plants and pathogens: Putting infection strategies and defence mechanisms on the map. Curr. Opin. Plant Biol. 2012, 15, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Shah, J. Plants under attack: Systemic signals in defence. Curr. Opin. Plant Biol. 2009, 12, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Girousse, C.; Moulia, B.; Silk, W.; Bonnemain, J.-L. Aphid Infestation Causes Different Changes in Carbon and Nitrogen Allocation in Alfalfa Stems as Well as Different Inhibitions of Longitudinal and Radial Expansion. Plant Physiol. 2005, 137, 1474–1484. [Google Scholar] [CrossRef] [Green Version]
- Pegadaraju, V.; Knepper, C.; Reese, J.; Shah, J. Premature leaf senescence modulated by the Arabidopsis PHYTOALEXIN DEFICIENT4 gene is associated with defense against the phloem-feeding green peach aphid. Plant Physiol. 2005, 139, 1927–1934. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.P.; Lichtenzveig, J.; Gleason, C.; Oliver, R.P.; Singh, K.B. The B-3 Ethylene Response Factor MtERF1-1 Mediates Resistance to a Subset of Root Pathogens in Medicago truncatula without Adversely Affecting Symbiosis with Rhizobia. Plant Physiol. 2010, 154, 861–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Hassan, S.; Kidd, B.N.; Garg, G.; Mathesius, U.; Singh, K.B.; Anderson, J.P. Ethylene Signaling Is Important for Isoflavonoid-Mediated Resistance to Rhizoctonia solani in Roots of Medicago truncatula. Mol. Plant-Microbe Interact. 2017, 30, 691–700. [Google Scholar] [CrossRef] [Green Version]
- Veerappan, V.; Kadel, K.; Alexis, N.; Scott, A.; Kryvoruchko, I.; Sinharoy, S.; Taylor, M.; Udvardi, M.; Dickstein, R. Keel petal incision: A simple and efficient method for genetic crossing in Medicago truncatula. Plant Methods 2014, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Ellwood, S.R.; D’Souza, N.K.; Kamphuis, L.G.; Burgess, T.I.; Nair, R.M.; Oliver, R.P. SSR analysis of the Medicago truncatula SARDI core collection reveals substantial diversity and unusual genotype dispersal throughout the Mediterranean basin. Theor. Appl. Genet. 2006, 112, 977–983. [Google Scholar] [CrossRef]
- Hayden, M.J.; Nguyen, T.M.; Waterman, A.; Chalmers, K.J. Multiplex-Ready PCR: A new method for multiplexed SSR and SNP genotyping. BMC Genom. 2008, 9, 80. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Kamphuis, L.G.; Guo, Y.; Jacques, S.; Singh, K.B.; Gao, L.-L. Ethylene Is Not Essential for R-Gene Mediated Resistance but Negatively Regulates Moderate Resistance to Some Aphids in Medicago truncatula. Int. J. Mol. Sci. 2020, 21, 4657. https://doi.org/10.3390/ijms21134657
Zhang L, Kamphuis LG, Guo Y, Jacques S, Singh KB, Gao L-L. Ethylene Is Not Essential for R-Gene Mediated Resistance but Negatively Regulates Moderate Resistance to Some Aphids in Medicago truncatula. International Journal of Molecular Sciences. 2020; 21(13):4657. https://doi.org/10.3390/ijms21134657
Chicago/Turabian StyleZhang, Lijun, Lars G. Kamphuis, Yanqiong Guo, Silke Jacques, Karam B. Singh, and Ling-Ling Gao. 2020. "Ethylene Is Not Essential for R-Gene Mediated Resistance but Negatively Regulates Moderate Resistance to Some Aphids in Medicago truncatula" International Journal of Molecular Sciences 21, no. 13: 4657. https://doi.org/10.3390/ijms21134657
APA StyleZhang, L., Kamphuis, L. G., Guo, Y., Jacques, S., Singh, K. B., & Gao, L. -L. (2020). Ethylene Is Not Essential for R-Gene Mediated Resistance but Negatively Regulates Moderate Resistance to Some Aphids in Medicago truncatula. International Journal of Molecular Sciences, 21(13), 4657. https://doi.org/10.3390/ijms21134657