Metabolic Characterisation of the Midgut of Bombyx mori Varieties after BmNPV Infection Using GC-MS-Based Metabolite Profiling

Abstract
:1. Introduction
2. Results
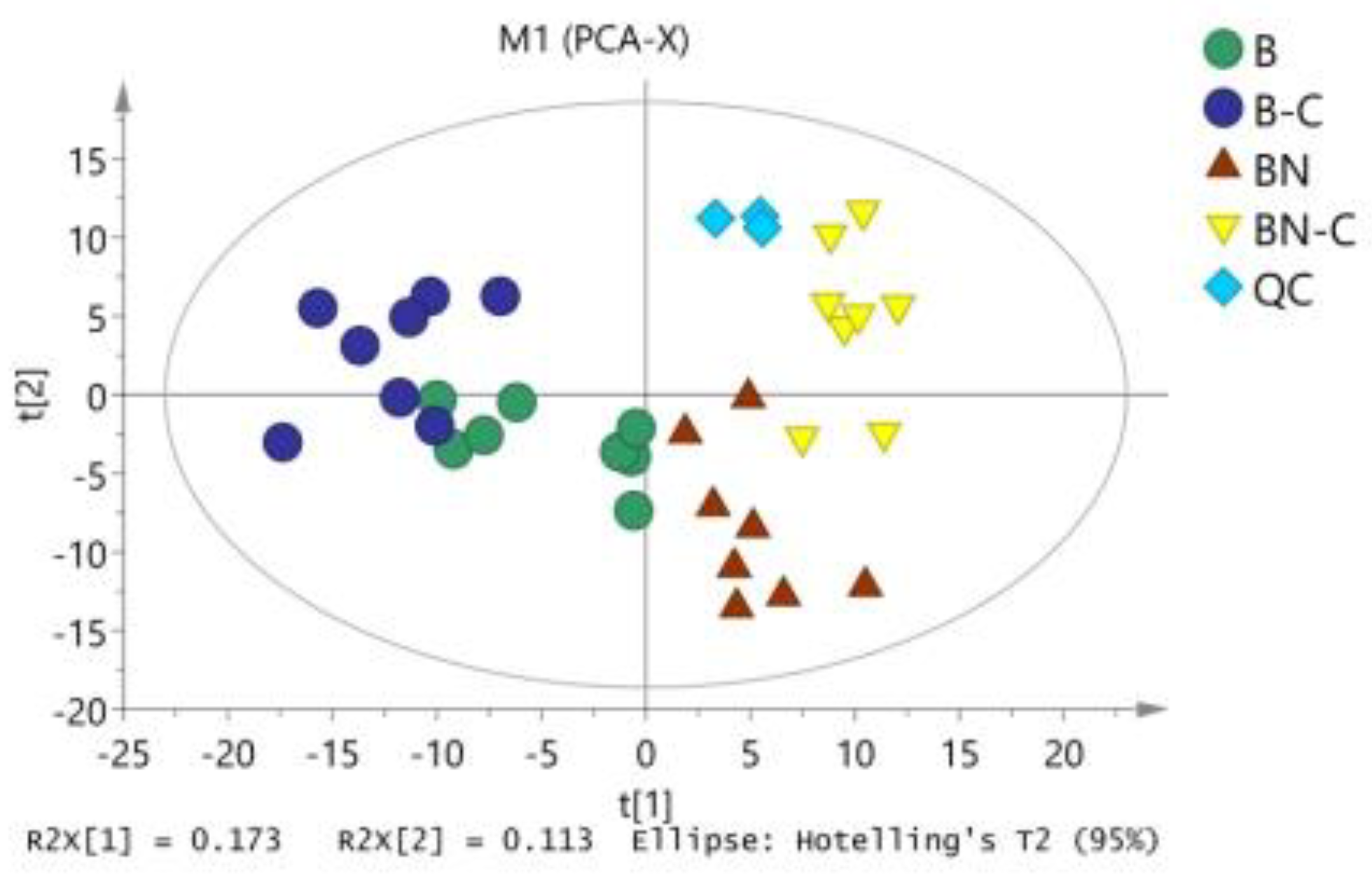
2.1. Inter-Sample Atlas Detection
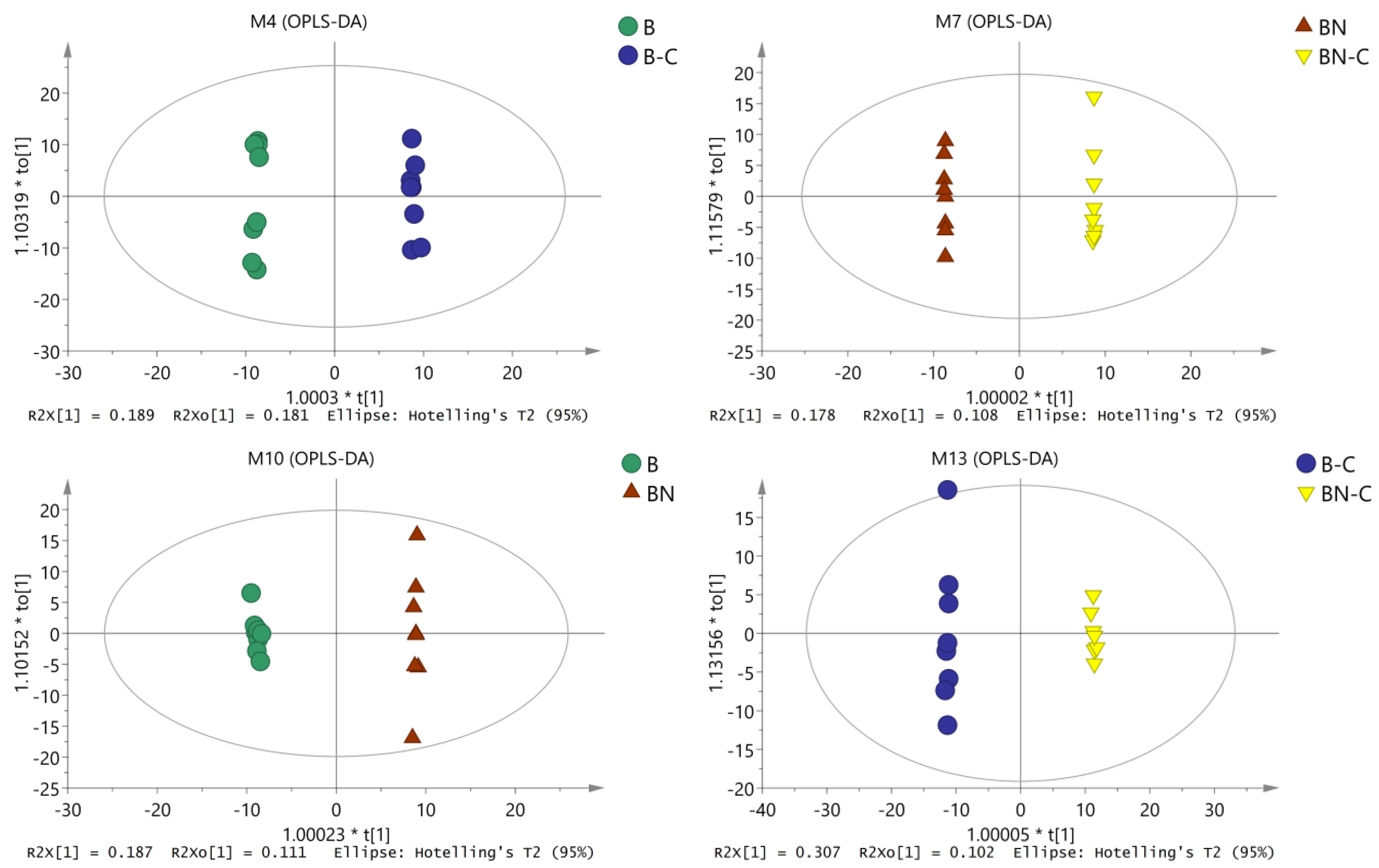
2.2. OPLS-DA Analysis
2.3. Screening of Differential Metabolites
2.3.1. Metabonomic Comparison between Different Silkworm Varieties Pre- and Post BmNPV Infection
2.3.2. Metabonomic Comparison between the Same Silkworm Variety before and after Adding BmNPV
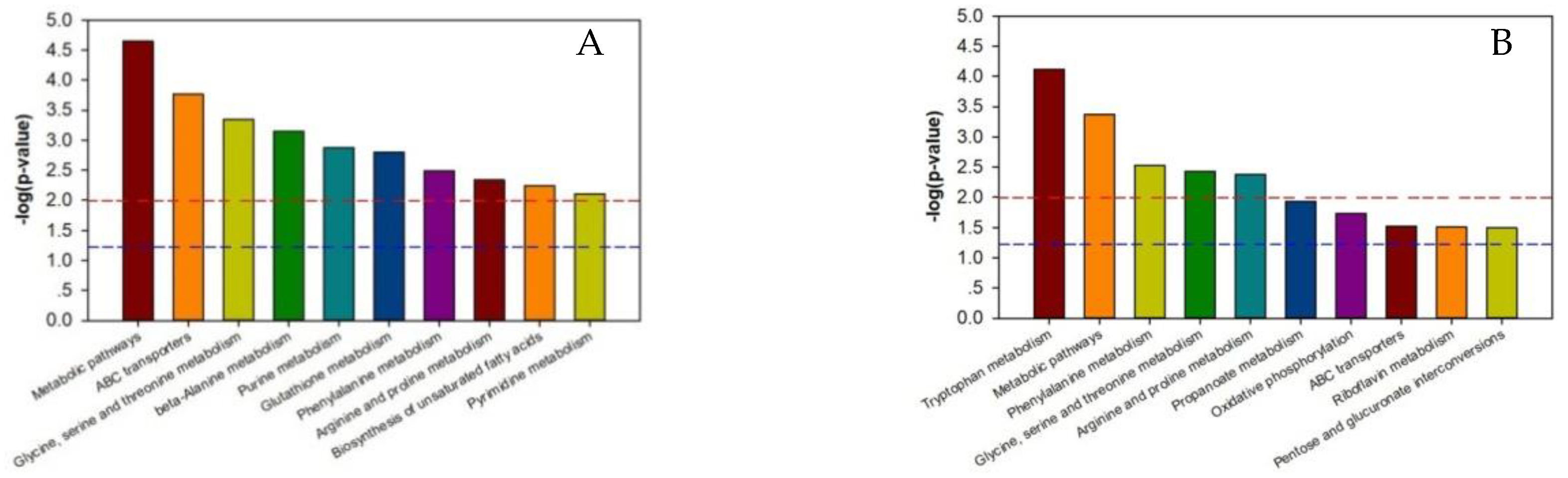
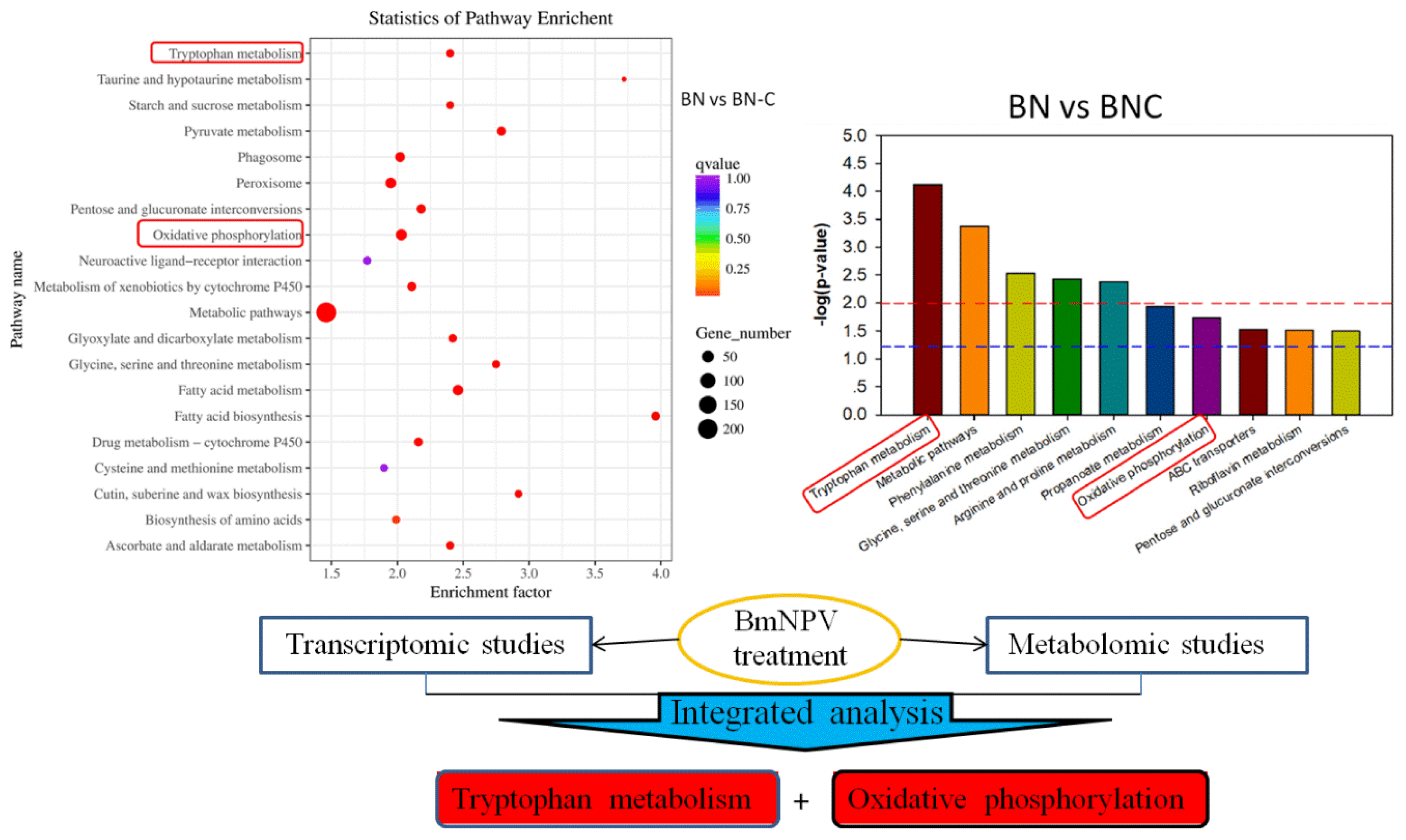
2.4. Differential Metabolites Metabolic Pathways Analysis
2.5. Analysis and Mining of Transcriptome Data
2.5.1. Analysis of Differentially Expressed Genes
2.5.2. KEGG Annotation of Differentially Expressed Genes
3. Discussion
4. Materials and Methods
4.1. Test Materials and Sample Preparation
4.2. Sample Preprocessing
4.3. GC-MS Analysis
4.4. Screening Differential Metabolites
4.5. Analysis of Metabolic Pathways of Differential Metabolites
4.6. Analysis and Screening of Differentially Expressed Genes
4.7. KEGG Annotation of Differentially Expressed Genes
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- China Silk Yearbook; Zhejiang University of Science and Technology Press: Hangzhou, China, 2014; pp. 254–256.
- Li, J.Q.; Gu, G.D. Silkworm-rearing willingness, sericulture risk and prevention measures—A questionnaire survey based on 1782 rural households in 91 counties of 14 provinces. Seric. Sci. 2013, 39, 355–364. [Google Scholar]
- Qian, H.Y.; Xu, A.Y.; Lin, C.Q.; Zhao, Y.P.; Sun, P.J.; Zhang, Y.H. Studies on the resistance to nuclear polyhedrosis virus (NPV) and its inheritance law in silkworm Bombyx Mori. J. Agric. Univ. Hebei 2006, 29, 77–79, 91. [Google Scholar]
- Xu, A.Y.; Lin, C.Q.; Qian, H.Y.; Sun, P.J.; Zhang, Y.H.; Liu, M.Z.; Li, L. Breeding of a new silkworm variety "Huakang 2" with tolerance to Bombyx mori nucleopolyhedrovirus disease. Seric. Sci. 2013, 39, 275–282. [Google Scholar]
- Mao, F.; Lei, J.; Enoch, O.; Wei, M.; Zhao, C.; Quan, Y.; Yu, W. Quantitative proteomics of Bombyx mori after BmNPV challenge. J. Proteom. 2018, 181, 142–151. [Google Scholar] [CrossRef]
- Singh, C.P.; Singh, J.; Nagaraju, J. bmnpv-miR-3 facilitates BmNPV infection by modulating the expression of viral P6.9 and other late genes in Bombyx mori. Insect. Biochem. Mol. Biol. 2014, 49, 59–69. [Google Scholar] [CrossRef]
- Yu, H.; Wang, X.; Xu, J.; Ma, Y.; Zhang, S.; Yu, D.; Fei, D.; Muhammad, A. iTRAQ-based quantitative proteomics analysis of molecular mechanisms associated with Bombyx mori (Lepidoptera) larval midgut response to BmNPV in susceptible and near-isogenic strains. J. Proteom. 2017, 165, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xia, D.; Zhao, Q.; Zhang, G.; Zhang, Y.; Qiu, Z.; Shen, D.; Lu, C. Label-free proteomic analysis of silkworm midgut infected by Bombyx mori nuclear polyhedrosis virus. J. Proteom. 2019, 200, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.S. Principles of Insect Immunolog; Shanghai Science and Technology Press: Shanghai, China, 2018; pp. 218–220. [Google Scholar]
- Qin, J.; Yuan, L.W. The latest research progress on domesticated silkworm genome in china—The reading of fine genome map of domesticated silkworm and 40 genetic variation maps of domesticated and wild silkworms. Seric. Sci. 2010, 36, 115–119. [Google Scholar]
- Fang, S.M.; Hu, B.L.; Zhou, Q.Z.; Yu, Q.Y.; Zhang, Z. Comparative analysis of the silk gland transcriptomes between the domestic and wild silkworms. BMC Genom. 2015, 16, 60. [Google Scholar] [CrossRef] [Green Version]
- Moghaddam, S.H.H.; Xin, D.; Li, J.; Cao, J.; Zhong, B.; Chen, Y.Y. Proteome analysis on differentially expressedproteins of the fat body of two silkworm breeds, Bombyx Mori, Exposed to heat shock exposure. Biotechnol. Bioprocess Eng. 2008, 13, 624–631. [Google Scholar] [CrossRef]
- Li, G.; Zhang, X.; Qian, H.Y.; Liu, M.Z.; Zhao, G.D.; Xu, A.Y. Gas chromatography-mass spectrometry based midgut metabolomics reveals the metabolic perturbations under NaF stress in Bombyx mori. Insects 2020, 11, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. MetPA: A web-based metabolomics tool for pathway analysis and visualization. Bioinformatics 2010, 26, 2342–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.L.; Zhang, S.X.; Tao, H.; Chen, Z.H.; Li, X.; Qiu, J.F.; Cui, W.Z.; SI Ma, Y.H.; Cui, W.Z.; Xu, S.Q. Metabolomics differences between silkworms (Bombyx mori) reared on fresh mulberry (Morus) leaves or artificial diets. Sci. Rep. 2017, 7, 10972. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, L.; Kettanehwold, N.; Trygg, J.; Wikström, C.; Wold, S. Multi- and Megavariate Data Analysis: Part I: Basic Principles and Applications; Umetrics Academy: Umea, Sweden, 2006; pp. 63–98. [Google Scholar]
- Peng, M.K.; Pallassana, N.; Bowden, R.; Cunningham, B.; Joy, A.; Kohn, J.; Babensee, J.E. Predicting biomaterial property-dendritic cell phenotype relationships from the multivariate analysis of responses to polymethacrylates. Biomaterials 2012, 33, 1699–1713. [Google Scholar]
- Tanaka, H.; Ishibashi, J.; Fujita, K.; Nakajima, Y.; Sagisaka, A.; Tomimoto, K.; Suzuki, N.; Yoshiyama, M.; Kaneko, Y.; Iwasaki, T.; et al. A genome-wide analysis of genes and gene families involved in innate immunity of Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1087–1110. [Google Scholar] [CrossRef]
- Angata, T.; Hayakawa, T.; Yamanaka, M.; Varki, A.; Nakamura, M. Discovery of Siglec-14, a novel sialic acid receptor undergoing concerted evolution with Siglec-5 in primates. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2006, 20, 1964–1973. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Kanost, M.R. The clip-domain family of serine proteases in arthropods. Insect Biochem. Mol. Biol. 2000, 30, 95–105. [Google Scholar] [CrossRef]
- Selot, R.; Kumar, V.; Shukla, S.; Chandrakuntal, K.; Kumar, P.G. Identification of a Soluble NADPH Oxidoreductase (BmNOX) with antiviral activites in the gut juice of Bombyx mori. J. Agric. Chem. Soc. Jpn. 2007, 71, 200–205. [Google Scholar]
- Ponnuvel, K.M.; Nakazawa, H.; Furukawa, S.; Asaoka, A.; Ishibashi, J.; Tanaka, H.; Yamakawa, M. A lipase isolated from the silkworm Bombyx mori shows antiviral activity against nucleopolyhedrovirus. J. Virol. 2003, 77, 10725–10729. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Wang, X.Y.; Du, C.; Gao, J.; Xu, J.P. Expression analysis of several antiviral related genes to BmNPV in different resistant strains of silkworm, Bombyx mori. J. Insect Sci. 2014, 14, 76. [Google Scholar] [CrossRef]
- Gowda, M.K.; Peter, A.; Naika, M.B.; Niranjana, P.; Shamprasad, P. Identification of in-vitro red fluorescent protein with antipathogenic activity from the midgut of the silkworm (Bombyx Mori L.). Protein Peptide Lett. 2018, 25, 302–313. [Google Scholar]
- Liu, W.; Liu, J.; Lu, Y.; Gong, Y.; Zhu, M.; Chen, F.; Liang, Z.; Zhu, L.; Kuang, S.; Hu, X. Immune signaling pathways activated in response to different pathogenic micro-organisms in Bombyx mori. Mol. Immunol. 2015, 65, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the gut microbiota on intestinal immunity mediated by tryptophan metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 2–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papa, S.; Martino, P.L.; Capitanio, G.; Gaballo, A.; Rasmo, D.D.; Signorile, A.; Petruzzella, V. The oxidative phosphorylation system in mammalian mitochondria. Adv. Exp. Med. Biol. 2012, 942, 3. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, 1344–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotech. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Alexa, A.; Rahnenfuhrer, J. topGO: Enrichment analysis for gene ontology. R Package Version 2010, 2, 8. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Model | Type | A | N | R2X(cum) | R2Y(cum) | Q2(cum) | R2 | Q2 |
|---|---|---|---|---|---|---|---|---|---|
| All | M1 | PCA-X | 6 | 35 | 0.553 | 0.221 | |||
| B vs. B-C | M2 | PCA-X | 3 | 16 | 0.51 | 0.145 | |||
| BN vs. BN-C | M5 | PCA-X | 3 | 16 | 0.488 | 0.0336 | |||
| BN vs. B | M8 | PCA-X | 4 | 16 | 0.586 | 0.0885 | |||
| BN-C vs. B-C | M11 | PCA-X | 4 | 16 | 0.653 | 0.208 | |||
| B vs. B-C | M4 | OPLS-DA | 1+2+0 | 16 | 0.426 | 0.999 | 0.894 | 0.978 | −0.216 |
| BN vs. BN-C | M7 | OPLS-DA | 1+3+0 | 16 | 0.461 | 1 | 0.835 | 0.742 | −1.080 |
| BN vs. B | M10 | OPLS-DA | 1+2+0 | 16 | 0.409 | 0.999 | 0.854 | 0.904 | −0.196 |
| BN-C vs. B-C | M13 | OPLS-DA | 1+2+0 | 16 | 0.499 | 1 | 0.966 | 0.813 | −0.471 |
| Tissue | Group | Metabolites | Up | Down | QuantMass | RT(min) | VIP | p-Value | FC |
|---|---|---|---|---|---|---|---|---|---|
| Midgut | B vs. B-C | 124 | 73 | 51 | 56–451 | 5.1286–35.1984 | 1.0065–2.1926 | 2.4055 × 10−8–0.0494 | 7.9322 × 10−6–310452.02 |
| BN vs. BN-C | 117 | 56 | 61 | 58–451 | 5.1766–36.1096 | 1.0555–2.3677 | 9.5441 × 10−8–0.0499 | 2.6607 × 10−6–685403.10 | |
| BN vs. B | 117 | 75 | 42 | 71–468 | 5.1955–34.9107 | 1.0277–2.2455 | 3.4469 × 10−10–0.0484 | 4.3374 × 10−6–58324.76 | |
| BN-C vs. B-C | 178 | 101 | 77 | 56–397 | 5.2509–38.0193 | 1.0014–1.8053 | 3.1763 × 10−12–0.0499 | 3.5462 × 10−7–596800.06 |
| Tissue | Group | Metabolites | QuantMass | RT(min) | VIP | p Value | FC | Average(B) | Average(BC) |
|---|---|---|---|---|---|---|---|---|---|
| Midgut | B vs. B-C | 2-Amino-2-norbornanecarboxylic acid | 89 | 8.3346 | 1.5773 | 0.0044 | 51086.82 | 0.051087 | 0.000001 |
| Benzoylformic acid | 255 | 34.9107 | 1.5502 | 0.0045 | 67933.12 | 0.067933 | 0.000001 | ||
| Adipamide | 215 | 25.6626 | 1.5467 | 0.0045 | 310452.02 | 0.310452 | 0.000001 | ||
| 2,4-diaminobutyric acid | 176 | 14.0483 | 2.0223 | 1.22 × 10−5 | 7.93 × 10−6 | 0.000001 | 0.126068 | ||
| BN vs. BN-C | Phenylacetic acid | 83 | 5.2971 | 2.3675 | 1.31 × 10−11 | 685403.1 | 0.685403 | 0.000001 | |
| Indolelactate | 202 | 34.8854 | 1.5517 | 0.0436 | 37819.53 | 0.037820 | 0.000001 | ||
| 1,3-Cyclohexanedione | 216 | 13.7762 | 1.5538 | 0.0204 | 25094.28 | 0.025094 | 0.000001 | ||
| Analyte 173 | 327 | 7.7791 | 1.5601 | 0.01957 | 12816.88 | 0.012817 | 0.000001 | ||
| octanal | 96 | 22.5501 | 2.3677 | 9.54 × 10−14 | 2.66 × 10−6 | 0.000001 | 0.375837 | ||
| 1-Hexadecanol | 184 | 10.4910 | 2.0596 | 0.0008 | 5.33 × 10−6 | 0.000001 | 0.187692 | ||
| BN vs. B | 4-Hydroxymandelonitrile | 205 | 25.4390 | 1.5148 | 0.0362 | 58324.76 | 0.058325 | 0.000001 | |
| octanal | 96 | 22.5501 | 2.0407 | 1.07 × 10−5 | 4.34 × 10−6 | 0.000001 | 0.230551 | ||
| 6-deoxy-D-glucose | 156 | 19.7094 | 1.7947 | 0.001 | 6.80 × 10−6 | 0.000001 | 0.147048 | ||
| Benzoylformic acid | 255 | 34.9107 | 1.5556 | 0.0045 | 1.47 × 10−5 | 0.000001 | 0.067933 | ||
| BN-C vs. B-C | 2-Amino-2-norbornanecarboxylic acid | 89 | 8.3346 | 1.8053 | 3.18 × 10−12 | 88556.14 | 0.088556 | 0.000001 | |
| 1-Hexadecanol | 184 | 10.4910 | 1.5828 | 0.0008 | 187691.72 | 0.187692 | 0.000001 | ||
| N-Acetyl-5-hydroxytryptamine | 231 | 8.0323 | 1.3072 | 0.0077 | 495947.17 | 0.495947 | 0.000001 | ||
| Analyte 963 | 72 | 24.1345 | 1.0548 | 0.0205 | 596800.07 | 0.596800 | 0.000001 | ||
| Phenylacetic acid | 83 | 5.2971 | 1.8047 | 4.94 × 10−9 | 1.02 × 10−6 | 0.000001 | 0.981171 | ||
| Farnesal | 217 | 27.0758 | 1.7906 | 1.21 × 10−5 | 3.55 × 10−7 | 0.000001 | 2.819911 | ||
| leucine | 158 | 5.9749 | 1.5922 | 0.0001 | 1.52 × 10−5 | 0.000001 | 0.065595 | ||
| Benzylsuccinic acid | 131 | 12.3266 | 1.4033 | 0.0006 | 3.93 × 10−6 | 0.000001 | 0.254367 | ||
| 1,3-Cyclohexanedione | 216 | 13.7762 | 1.3965 | 0.0008 | 2.72 × 10−5 | 0.000001 | 0.036744 | ||
| 4-Hydroxybenzyl cyanide | 221 | 10.1969 | 1.0416 | 0.0226 | 3.22 × 10−5 | 0.000001 | 0.031097 | ||
| Indolelactate | 202 | 34.8854 | 1.0462 | 0.0234 | 1.46 × 10−5 | 0.000001 | 0.068349 |
| DEG Set | DEG Number | Up-Regulated | Down-Regulated | KEGG Pathway |
|---|---|---|---|---|
| B vs. B-C | 2651 | 1406 | 1245 | 561 |
| BN vs. B | 2622 | 1577 | 1045 | 460 |
| BN-C vs. B-C | 1522 | 1023 | 499 | 276 |
| BN vs. BN-C | 434 | 209 | 225 | 96 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, H.; Li, G.; Zhao, G.; Liu, M.; Xu, A. Metabolic Characterisation of the Midgut of Bombyx mori Varieties after BmNPV Infection Using GC-MS-Based Metabolite Profiling. Int. J. Mol. Sci. 2020, 21, 4707. https://doi.org/10.3390/ijms21134707
Qian H, Li G, Zhao G, Liu M, Xu A. Metabolic Characterisation of the Midgut of Bombyx mori Varieties after BmNPV Infection Using GC-MS-Based Metabolite Profiling. International Journal of Molecular Sciences. 2020; 21(13):4707. https://doi.org/10.3390/ijms21134707
Chicago/Turabian StyleQian, Heying, Gang Li, Guodong Zhao, Mingzhu Liu, and Anying Xu. 2020. "Metabolic Characterisation of the Midgut of Bombyx mori Varieties after BmNPV Infection Using GC-MS-Based Metabolite Profiling" International Journal of Molecular Sciences 21, no. 13: 4707. https://doi.org/10.3390/ijms21134707
APA StyleQian, H., Li, G., Zhao, G., Liu, M., & Xu, A. (2020). Metabolic Characterisation of the Midgut of Bombyx mori Varieties after BmNPV Infection Using GC-MS-Based Metabolite Profiling. International Journal of Molecular Sciences, 21(13), 4707. https://doi.org/10.3390/ijms21134707