Functional Markers for Precision Plant Breeding



Abstract
:1. Introduction
2. Brief History of Molecular Marker Development
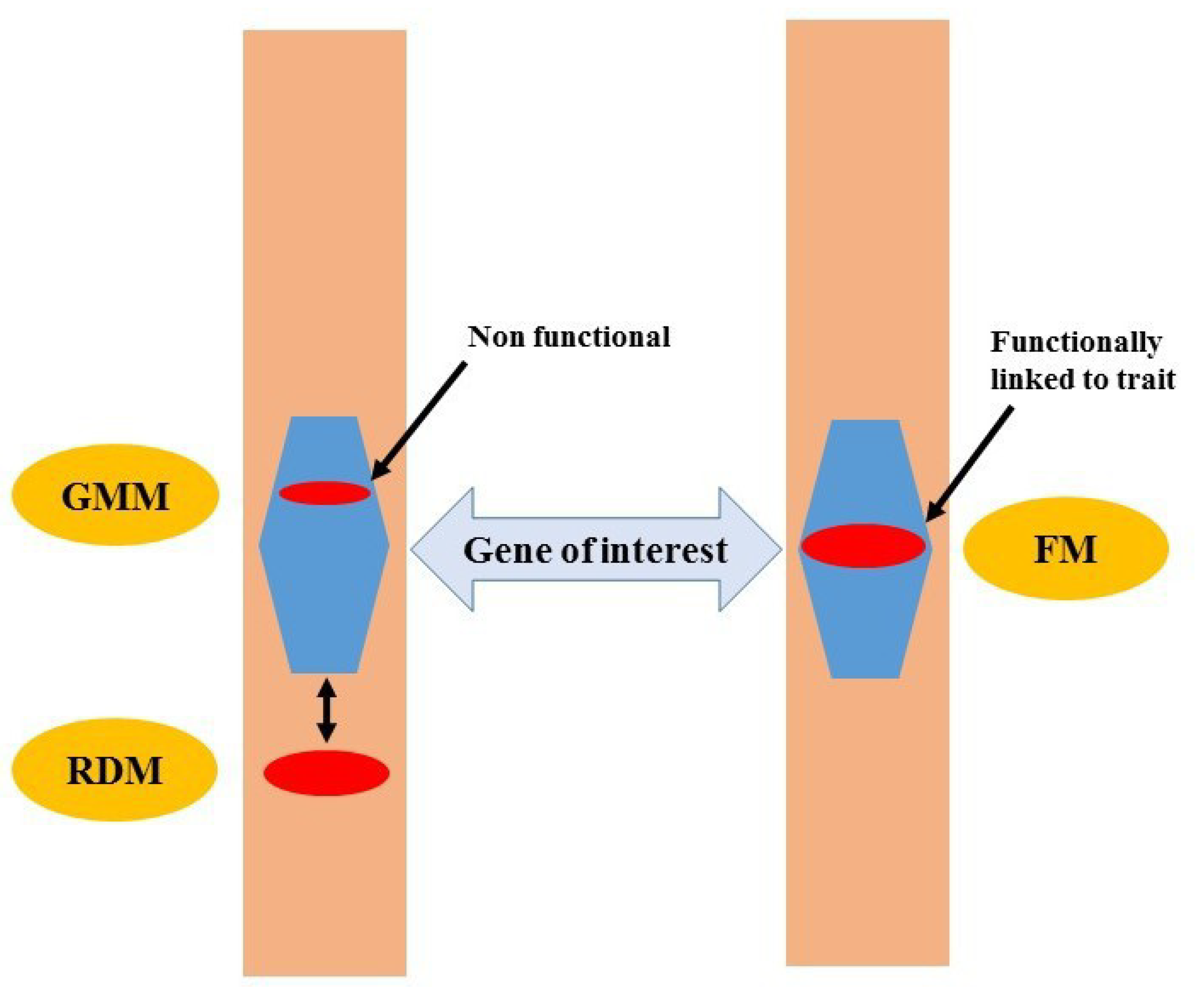
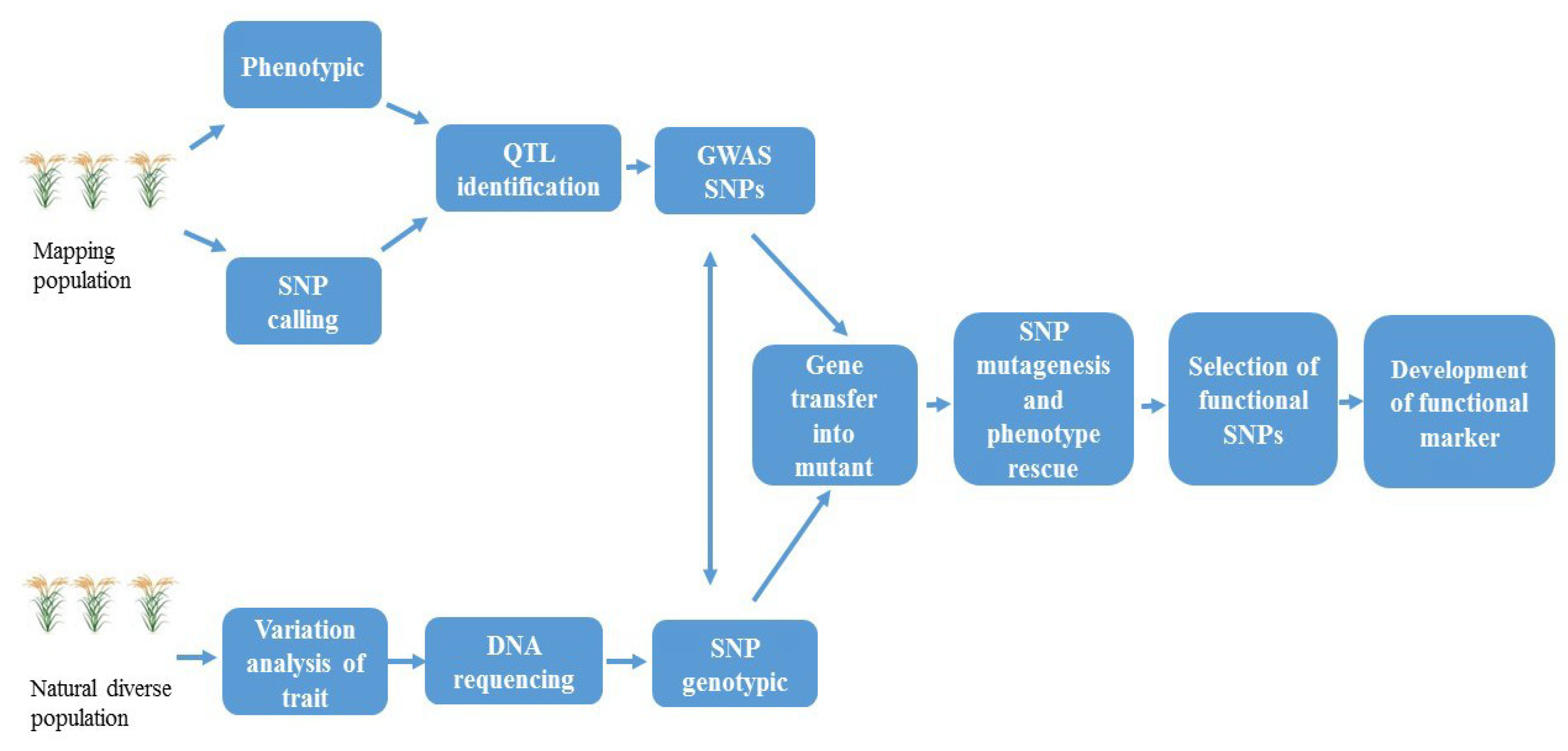
3. Functional Markers
4. Advantages of FMs over Other Markers
5. FMs in Precision Plant Breeding
5.1. Germplasm Evaluation and Genetic Diversity
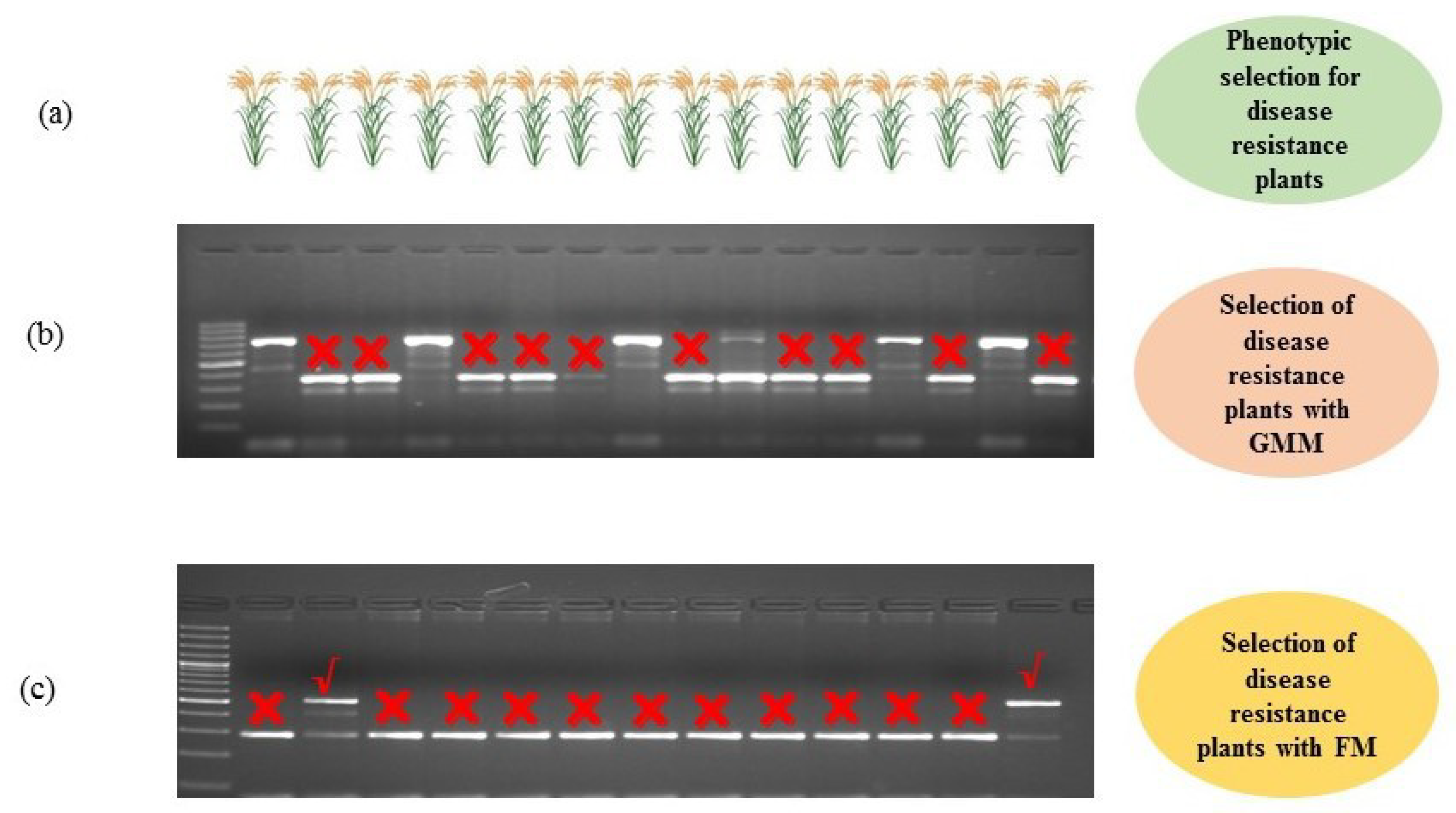
5.2. Marker Assisted Selection
5.3. Gene Pyramiding
5.4. Genomic Selection
6. FMs for the Improvement of Agronomic Traits, Quality Traits, and Stress Resistance
6.1. FMs for Agronomic Traits
6.2. FM for Quality Traits
6.3. FMs for Biotic Stress Resistance
6.4. FMs for Abiotic Stress Tolerance
7. Future Prospects and Conclusions
Author Contributions
Funding
Conflicts of Interest
Compliance with Ethical Standards
References
- Gupta, P.K.; Rustgi, S.; Kulwal, P.L. Linkage disequilibrium and association studies in higher plants: Present status and future prospects. Plant Mol. Biol. 2005, 57, 461–485. [Google Scholar] [CrossRef] [PubMed]
- Collard, B.C.; Jahufer, M.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Kumar, M.; Chaudhary, V.; Sharma, R.; Sirohi, U.; Singh, J. Advances in biochemical and molecular marker techniques and their applications in genetic studies of orchid: A review. Int. J. Chem. Stud. 2018, 6, 806–822. [Google Scholar]
- Winter, P.; Kahl, G. Molecular marker technologies for plant improvement. World J. Microbiol. Biotechnol. 1995, 11, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Frona, D.; Szenderak, J.; Harangi-Rakos, M. The challenge of feeding the World. Sustainability 2019, 11, 5816. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Xie, W.; He, H.; Yu, H.; Chen, W.; Li, J.; Yu, R.; Yao, Y.; Zhang, W.; He, Y.; et al. A high-density SNP genotyping array for rice biology and molecular breeding. Mol. Plant 2014, 7, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Oliver, M.J. Why we need GMO crops in agriculture. Mo. Med. 2014, 111, 493–507. [Google Scholar]
- Conner, T. Precision breeding: A new genetic technique providing international opportunities for crop improvement. Seed Quest 2004. Available online: https://www.seedquest.com (accessed on 15 April 2020).
- Bohra, A.; Bharadwaj, C.; Radhakrishnan, T.; Singh, N.P.; Varshney, R.K. Translational genomics and molecular breeding for enhancing precision and efficiency in crop improvement programs: Some examples in legumes. Indian J. Genet. 2019, 79, 227–240. [Google Scholar] [CrossRef]
- Schmutz, J.; Steven, B.; Cannon Schlueter, J.; Jianxin, M.; Therese, M.; William, N.; David, H.L.; Qijian, S.; Jay, T.J.; Jianlin, C. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhutmal, R.R.; Mundhe, A.G.; More, A.W. Molecular marker techniques: A Review. Int. J. Curr. Microbiol. Appl. Sci. 2018, 6, 816–825. [Google Scholar]
- Amom, T.; Nongdam, P. The use of molecular marker methods in plants: A review. Int. J. Curr. Res. Rev. 2017, 9, 1–9. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphism. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Williams, J.; Kubelik, A.; Livak, K.; Rafalski, J.; Tingey, S. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, J.; McClelland, M. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res. 1990, 18, 7213–7218. [Google Scholar] [CrossRef] [Green Version]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Tautz, D. Hypervariability of simple sequences as a general source of polymorphic DNA markers. Nucleic Acids Res. 1989, 17, 6463–6471. [Google Scholar] [CrossRef]
- Litt, M.; Luty, J.A. A hypervariable microsatellite revealed by in vitro amplification of a dinucleotide repeat within the cardiac muscle actin gene. Am. J. Hum. Genet. 1989, 44, 397–401. [Google Scholar]
- Paran, I.; Michelmore, R.W. Development of reliable PCR-based markers linked to downy mildew resistance genes in lettuce. Theor. Appl. Genet. 1993, 85, 985–993. [Google Scholar] [CrossRef]
- Konieczny, A.; Ausubel, F.M. A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specific PCR-based markers. Plant J. 1993, 4, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; Roy, J.K.; Prasad, M. Single nucleotide polymorphisms: A new paradigm for molecular marker technology and DNA polymorphism detection with emphasis on their use in plants. Curr. Sci. 2001, 80, 524–535. [Google Scholar]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity arrays: A solid state technology for sequence information independent genotyping. Nucleic Acids Res. 2001, 29, E25. [Google Scholar] [CrossRef] [Green Version]
- Sousa, T.V.; Caixeta, E.T.; Alkimim, E.R.; Oliveira, A.C.B.; Pereira, A.A.; Sakiyama, N.S.; Zambolim, L.; Resende, M.D.V. Early selection enabled by the implementation of genomic selection in Coffea arabica breeding. Front. Plant Sci. 2019, 9, 1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, J.R.; Lubberstedt, T. Functional markers in plants. Trends Plant Sci. 2003, 8, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genomics-assisted breeding for crop improvement. Trends Plant Sci. 2005, 10, 621–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagge, M.; Xia, X.; Lubberstedt, T. Functional markers in wheat. Curr. Opin. Plant Biol. 2007, 10, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Borevitz, J.O.; Nordborg, M. The impact of genomics on the study of natural variation in Arabidopsis. Plant Physiol. 2003, 132, 718–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgotra, R.K.; Gupta, B.B.; Stewart, C.N. From genomics to functional markers in the era of next-generation sequencing. Biotechnol. Lett. 2014, 36, 417–426. [Google Scholar] [CrossRef]
- Ingvardsen, C.; Schejbel, B.; Lubberstedt, T. Functional markers in resistance breeding. In Progress in Botany; Springer: Berlin/Heidelberg, Germany, 2008; Volume 69, pp. 61–87. [Google Scholar]
- Thornsberry, J.M.; Goodman, M.M.; Doebley, J.; Kresovich, S.; Nielsen, D.; Buckler, E.S. Dwarf8 polymorphisms associate with variation in flowering time. Nat. Gen. 2001, 28, 286–289. [Google Scholar] [CrossRef]
- McCouch, S.R.; Chen, X.; Panaud, O.; Temnykh, S.; Xu, Y.; Cho, Y.G.; Huang, N.; Ishii, T.; Blair, M. Microsatellite marker development, mapping and applications in rice genetics and breeding. Plant Mol. Biol. 1997, 35, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Soto-Cerda, B.J.; Cloutier, S. Association mapping in plant genomes. In Genetic Diversity in Plants; Mahmut, C., Ed.; IntechOpen Limited: London, UK, 2012; ISBN 978-953-51-0185-7. [Google Scholar]
- Rostoks, N.T.; Ramsay, L.; Mackenzie, K.; Cardle, L.; Bhat, P.R.; Roose, M.L.; Svensson, J.T.; Stein, N.; Varshney, R.K.; Marshall, D.F.; et al. Recent history of artificial outcrossing facilitates whole-genome association mapping in elite inbred crop varieties. Proc. Natl. Acad. Sci. USA 2006, 103, 18656–18661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breseghello, F.; Sorrels, M.E. Association analysis as a strategy for improvement of qualitative traits in plants. Crop Sci. 2006, 46, 1323–1330. [Google Scholar] [CrossRef]
- Simko, I.; Pechenick, D.A.; McHale, L.K.; Truco, M.J.; Ochoa, O.E.; Michelmore, R.W.; Scheffler, B.E. Association mapping and marker-assisted selection of the lettuce dieback resistance gene Tvr1. BMC Plant Biol. 2009, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Nie, X.; Shen, C.; You, C.; Li, W.; Zhao, W.; Zhang, X.; Lin, Z. Population structure and genetic basis of the agronomic traits of upland cotton in China revealed by a genome-wide association study using high-density SNPs. Plant Biotechnol. J. 2017, 15, 1374–1386. [Google Scholar] [CrossRef]
- Forcada, C.; Guajardo, V.; Chin-Wo, S.R.; Moreno, M.Á. Association mapping analysis for fruit quality traits in Prunus persica using SNP markers. Front. Plant Sci. 2019, 9, 2005. [Google Scholar] [CrossRef] [Green Version]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Sonah, H.; Bastien, M.; Iquira, E.; Tardivel, A.; Tardivel, A.; Legare, G.; Boyle, B.; Normandeau, E.; Laroche, J.; Larose, S.; et al. An improved genotyping by sequencing (GBS) approach offering increased versatility and efficiency of SNP discovery and genotyping. PLoS ONE 2013, 8, e54603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardivel, A.; Sonah, H.; Belzile, F.; O’Donoughue, L.S. Rapid identification of alleles at the soybean maturity gene E3 using genotyping by sequencing and a haplotype-based approach. Plant Genome 2014, 7, 1–9. [Google Scholar] [CrossRef]
- Garg, R.; Jain, M. RNA-Seq for transcriptome analysis in non-model plants. Methods Mol. Biol. 2013, 43–58. [Google Scholar] [CrossRef]
- Chen, X.R.; Xing, Y.P.; Li, Y.P.; Tong, Y.H.; Xu, J.Y. RNA-Seq reveals infection-related gene expression changes in Phytophthora capsici. PLoS ONE 2013, 8, e74588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novaes, E.; Drost, D.R.; Farmerie, W.G.; Pappas, G.J.; Grattapaglia, D.; Sederoff, R.R.; Kirst, M. High-throughput gene and SNP discovery in Eucalyptus grandis, an uncharacterized genome. BMC Genom. 2008, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Rupe, M.A.; Danilevskaya, O.N.; Yang, X.; Hu, Z. Genome-wide mRNA profiling reveals heterochronic allelic variation and a new imprinted gene in hybrid maize endosperm. Plant J. 2003, 36, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Duran, C.; Appleby, N.; Clark, T.; Wood, D.; Imelfort, M.; Batley, J.; David, E. AutoSNPdb: An annotated single nucleotide polymorphism database for crop plants. Nucleic Acids Res. 2009, 37, 951–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wang, L.; Zhan, Q.; Liu, Y.; Yang, X. Transcriptome characterization and functional marker development in Sorghum sudanense. PLoS ONE 2016, 11, e0154947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Son, S.; Shin, M.; Fujii, N.; Hoshino, T.; Park, S. Transcriptome-wide mining, characterization, and development of microsatellite markers in Lychnis kiusiana (Caryophyllaceae). BMC Plant Biol. 2019, 19, 14. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Chu, P.; Wu, Y.; Chan, W.R.; Wang, Y.H. Identification of functional SSR markers in freshwater ornamental Shrimps Neocaridina denticulata using transcriptome sequencing. Mar. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Abdurakhmonov, I.Y.; Ayubov, M.S.; Ubaydullaeva, K.A.; Buriev, Z.T.; Shermatov, S.E.; Ruziboev, H.S.; Shapulatov, U.M.; Saha, S.; Ulloa, M.; Yu, J.Z.; et al. RNA interference for functional genomics and improvement of cotton (Gossypium sp.). Front. Plant Sci. 2016, 7, 202. [Google Scholar] [CrossRef] [Green Version]
- Segal, G.; Song, R.; Messing, J. A new opaque variant of maize by a single dominant RNA-interference-inducing transgene. Genetics 2003, 165, 387–397. [Google Scholar]
- Angaji, S.A.; Hedayati, S.S.; Poor, R.H.; Poor, S.S.; Shiravi, S.; Madani, S. Application of RNA interference in plants. Plant Omics J. 2010, 3, 77–84. [Google Scholar]
- Machado, A.; Wu, Y.; Yang, Y.; Llewellyn, D.J.; Dennis, E.S. The MYB transcription factor GhMYB25 regulates early fiber and trichome development. Plant J. 2009, 59, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Que, Q.; Jorgensen, R.A. Homology-based control of gene expression patterns in transgenic petunia flowers. Dev. Genet. 1998, 22, 100–109. [Google Scholar] [CrossRef]
- Sigoillot, F.D.; Lyman, S.; Huckins, J.F.; Adamson, B.; Chung, E.; Quattrochi, B.; King, R.W. A bioinformatics method identifies prominent off targeted transcripts in RNAi screens. Nat. Methods 2012, 9, 363–366. [Google Scholar] [CrossRef] [Green Version]
- Schaeffer, S.M.; Nakata, P.A. CRISPR/Cas9-mediated genome editing and gene replacement in plants: Transitioning from lab to field. Plant Sci. 2015, 240. [Google Scholar] [CrossRef] [PubMed]
- Romay, G.; Bragard, C. Antiviral defenses in plants through genome editing. Front. Microbiol. 2017, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.; Liu, T.; Li, C.; Jiao, B.; Li, S.; Hou, Y.; Luo, K. Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Sci. Rep. 2015, 5, 12217. [Google Scholar] [CrossRef]
- Liu, D.; Hu, R.; Palla, K.J.; Tuskan, G.A.; Yang, X. Advances and perspectives on the use of CRISPR/Cas9 systems in plant genomics research. Curr. Opin. Plant Biol. 2016, 30, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.A.; Wang, T.L.; Welham, T.J.; Gardner, S.; Pike, J.M.; Yoshida, S.; Parniske, M. A TILLING reverse genetics tool and a web-accessible collection of mutants of the legume Lotus japonicus. Plant Physiol. 2003, 131, 866–871. [Google Scholar] [CrossRef] [Green Version]
- Till, B.J.; Reynolds, S.H.; Weil, C.; Springer, N.; Burtner, C.; Young, K.; Bowers, E.; Codomo, C.A.; Enns, L.C.; Odden, A.R.; et al. Discovery of induced point mutations in maize genes by TILLING. BMC Plant Biol. 2004, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- McCallum, C.M.; Comai, L.; Greene, E.A.; Henikoff, S. Targeted screening for induced mutations. Nat. Biotechnol. 2000, 18, 455–457. [Google Scholar] [CrossRef]
- Slade, A.J.; Fuerstenberg, S.I.; Loeffler, D.; Steine, M.N.; Facciotti, D. A reverse genetic, nontransgenic approach to wheat crop improvement by TILLING. Nat. Biotechnol. 2005, 23, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Kage, U.; Kumar, A.; Dhokane, D.; Karre, S.; Kushalappa, A.C. Functional molecular markers for crop improvement. Crit. Rev. Biotechnol. 2016, 36, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, R.J. The functional genomics laboratory: Functional validation of genetic variants. J. Inherit. Metab. Dis. 2018, 41, 297–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burch-Smith, T.M.; Anderson, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and advantages of virus—Induced gene silencing for gene function studies in plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef]
- Tadege, M.; Ratet, P.; Mysore, K.S. Insertional mutagenesis: A swiss army knife for functional genomics of Medicago truncatula. Trends Plant Sci. 2005, 10, 229–235. [Google Scholar] [CrossRef]
- Gupta, P.K. Beyond CRISPR: Single base editors for human health and crop improvement. Curr. Sci. 2019, 116, 386–397. [Google Scholar] [CrossRef]
- Robertson, D. VIGS vectors for gene silencing: Many targets, many tools. Annu. Rev. Plant Biol. 2004, 55, 495–519. [Google Scholar] [CrossRef]
- Cakir, C.; Gillespie, M.E.; Scofield, S.R. Rapid determination of gene function by virus-induced gene silencing in wheat and barley. Crop Sci. 2010, 50, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Kumar, J.; Mir, R.R.; Kumar, A. Marker-assisted selection as a component of conventional plant breeding. In Plant Breeding Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 145–217. [Google Scholar]
- Lau, W.C.P.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.; Latif, M.A.; Ramli, A. Review of functional markers for improving cooking, eating, and the nutritional qualities of rice. Front. Plant Sci. 2015, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.A.; Yang, S.H.; Rehman, H.M.; Baloch, F.S.; Lee, J.D.; Park, J.H.; Chung, G. Genetic diversity and population structure of Korean wild soybean (Glycine soja Sieb. and Zucc.) inferred from microsatellite markers. Biochem. Syst. Ecol. 2017, 71, 87–96. [Google Scholar] [CrossRef]
- Edmeades, G.O.; McMaster, G.S.; White, J.W.; Campos, H. Genomics and the physiologist: Bridging the gap between genes and crop response. Field Crops Res. 2004, 90, 5–18. [Google Scholar] [CrossRef]
- Yang, H.; Li, C.; Lam, H.M.; Clements, J.; Yan, G.; Zhao, S. Sequencing consolidates molecular markers with plant breeding practice. Theor. Appl. Genet. 2015, 128, 779–795. [Google Scholar] [CrossRef]
- Xu, Y.; McCouch, S.R.; Zhang, Q. How can we use genomics to improve cereals with rice as a reference genome? Plant Mol. Biol. 2005, 59, 7–26. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Than, K.; Vanavichit, A.; Chai-arree, W.; Toojinda, T. Marker assisted back cross breeding to improve cooking quality traits in Myanmar rice cultivar Manawthukha. Field Crop Res. 2009, 113, 178–186. [Google Scholar] [CrossRef]
- Jin, L.; Lu, Y.; Shao, Y.; Xiao, P.; Shen, S.; Corke, H.; Bao, J. Molecular marker assisted selection for improvement of the eating, cooking and sensory quality of rice (Oryza sativa L.). J. Cereal Sci. 2010, 51, 159–164. [Google Scholar] [CrossRef]
- Yu, J.; Hu, S.; Wang, J.; Wong, G.K.; Li, S.; Liu, B.; Deng, Y.; Dai, L.; Zhou, Y.; Zhang, X.; et al. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 2002, 296, 79–92. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Dogan, Y.; Comertpay, G.; Yildiz, M.; Hatipoglu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhang, Q. The Rice Genome: Implications for Breeding Rice and Other Cereals. In New Directions for a Diverse Planet, Proceedings of the 4th International Crop Science Congress, Brisbane, Australia, 26 September–1 October 2014; Fischer, R.A., Ed.; Crop Science Society of America: Madison WI, USA, 2014. [Google Scholar]
- Jamali, S.H.; Cockram, J.; Hickey, L.T. Insights into deployment of DNA markers in plant variety protection and registration. Theor. Appl. Genet. 2018, 132, 1911–1929. [Google Scholar] [CrossRef]
- Hodgkin, T.; Roviglioni, R.; De Vicente, M.C.; Dudnik, N. Molecular methods in the conservation and use of plant genetic resources. Acta Hort. 2001, 546, 107–118. [Google Scholar] [CrossRef]
- Kaisoon, O.; Siriamornpun, S.; Meeso, N. Distinction between cereal genotypes based on the protein and DNA composition of the grain by capillary electrophoresis. World Appl. Sci. J. 2008, 4, 384–395. [Google Scholar]
- Salgotra, R.K.; Gupta, B.B.; Monika, S. Biotechnological interventions and their role in sustainable hill agriculture. J. Plant Sci. Res. 2015, 2, 1–8. [Google Scholar]
- Jiang, G.L. Molecular markers and marker-assisted breeding in plants. In Plant Breeding from Laboratories to Fields; Andersen, S.B., Ed.; InTech: Rijeka, Croatia, 2013; pp. 45–83. [Google Scholar]
- Singh, K.; Kumar, S.; Kumar, S.R.; Singh, M.; Gupta, K. Plant genetic resources management and pre-breeding in genomics era. Indian J. Genet. 2019, 79, 117–130. [Google Scholar] [CrossRef]
- Rai, V.P.; Singh, A.K.; Jaiswal, H.K. Evaluation of molecular markers linked to fragrance and genetic diversity in Indian aromatic rice. Turk. J. Bot. 2015, 39, 209–217. [Google Scholar] [CrossRef]
- Beaumont, M.A.; Balding, D.J. Identifying adaptive genetic divergence among populations from genome scans. Mol. Ecol. 2004, 13, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.; Lammer, D.; Lyon, S.; Carter, B.; Jones, S.S. Breeding for organic and low-input farming systems: An evolutionary-participatory breeding method for inbred cereal grains. Renew. Agric. Food Syst. 2005, 20, 48–55. [Google Scholar] [CrossRef]
- Henry, R.J. (Ed.) Plant Genotyping—The DNA Fingerprinting of Plants; CABI Publishing: Wallingford, UK, 2001. [Google Scholar]
- Abdel-Mawgood, A.L. DNA Based techniques for studying genetic diversity. In Genetic Diversity in Microorganisms; IntechOpen Limited: London, UK, 2012; pp. 95–122. [Google Scholar]
- Loera-Sanchez, M.; Studer, B.; Kolliker, R. DNA-based assessment of genetic diversity in grassland plant species: Challenges, approaches, and applications. Agronomy 2019, 9, 881. [Google Scholar] [CrossRef] [Green Version]
- Tabor, H.K.; Risch, N.J.; Myers, R.M. Candidate-gene approaches for studying complex genetic traits: Practical considerations. Nat. Rev. Genet. 2002, 3, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, A.; Srivastava, S. Plant Breeding. In Participatory Plant Breeding: Concept and Applications; Springer: Singapore, 2019; pp. 29–68. [Google Scholar]
- Lubberstedt, T.; Zein, I.; Andersen, J.; Wenzel, G.; Krutzfeldt, B.; Eder, J.; Ouzunova, M.; Chun, S. Development and application of functional markers in maize. Euphytica 2005, 146, 101–108. [Google Scholar] [CrossRef]
- Lema, M. Marker assisted selection in comparison to conventional plant breeding. Agric. Res. Technol. 2018, 14, 555914. [Google Scholar] [CrossRef] [Green Version]
- Semagn, K.; Bjornstad, A.; Ndjiondjop, M.N. An overview of molecular marker methods for plants. Afr. J. Biotechnol. 2006, 2540, 25–68. [Google Scholar]
- Archak, S.; Lakshminarayanareddy, V.; Nagaraju, J. High-throughput multiplex microsatellite marker assay for detection and quantification of adulteration in basmati rice (Oryza sativa). Electrophoresis 2007, 28, 2396–2405. [Google Scholar] [CrossRef] [PubMed]
- Salgotra, R.K.; Millwood, R.J.; Agarwal, S.; Stewart, N.C. High-throughput functional marker assay for detection of Xa/xa and fgr genes in rice (Oryza sativa L.). Electrophoresis 2011, 32, 2216–2222. [Google Scholar] [CrossRef] [PubMed]
- Abdulmalik, R.O.; Menkir, A.; Meseka, S.; Unachukwu, N.; Ado, S.G.; Olarewaju, J.D.; Aba, D.A.; Hearne, S.; Crossa, J.; Gedil, M. Genetic gains in grain yield of a maize population improved through marker assisted recurrent selection under stress and non-stress conditions in West Africa. Front. Plant Sci. 2017, 8, 841. [Google Scholar] [CrossRef] [Green Version]
- Mullis, K.; Faloona, F.; Scharf, S.; Saiki, R.; Horn, G.; Erlich, H. Specific enzymatic amplification of DNA in vitro: The polymerase chain reaction. Cold Spring Harb. Symp. Quant. Biol. 1986, 51, 263–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, A.S.; McCouch, S.R. Functional markers for xa5 mediated resistance in rice (Oryza sativa L.). Mol. Breed. 2007, 19, 291–296. [Google Scholar] [CrossRef]
- Chukwu, S.C.; Rafii, M.Y.; Ramlee, S.I.; Ismail, S.I.; Oladosu, Y.; Okporie, E.E.; Onyashi, G.; Utobo, E.; Ekwu, L.; Swaray, S.; et al. Marker-assisted selection and gene pyramiding for resistance to bacterial leaf blight disease of rice (Oryza sativa L.). Biotechnol. Biotechnol. Equip. 2019, 33, 440–455. [Google Scholar] [CrossRef] [Green Version]
- Azhaguvel, P.; Saraswathi, D.; Sharma, A.; Varshney, R.K. Methodological advancement in molecular markers to delimit the gene(s) for crop improvement. In Advances in Molecular Markers for Crop Improvement; Azhaguvel, P., Ed.; Global Science Books: Isleworth, UK, 2006; pp. 460–469. [Google Scholar]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [PubMed]
- Saint Pierre, C.; Burgueno, J.; Crossa, J.; Davila, G.F.; Lopez, P.F.; Moya, E.S.; Moreno, I.I.; Muela, V.M.H.; Villa, V.M.H.; Vikram, P.; et al. Genomic prediction models for grain yield of spring bread wheat in diverse agro-ecological zones. Sci. Rep. 2016, 6, 27312. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.A.; Ali, S.; Salgotra, R.K.; Mir, Z.A.; Dutta, S.; Jadon, V.; Tyagi, A.; Mushtaq, M.; Jain, N.; Singh, P.K.; et al. Genomic selection in the era of next generation sequencing for complex traits in plant breeding. Front. Genet. 2016, 7, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crossa, J.; Beyene, Y.; Kassa, S.; Perez, P.; Hickey, J.M.; Chen, C.; de los Campos, G.; Burgueño, J.; Windhausen, V.S.; Buckler, E.; et al. Genomic prediction in maize breeding populations with genotyping-bysequencing. G3 (Bethesda) 2013, 3, 1903–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newell, M.A.; Jannink, J.L. Genomic selection in plant breeding. In Crop Breeding: Methods and Protocols; Fleury, D., Whitford, R., Eds.; Humana Press: New York, NY, USA, 2014; pp. 117–130. [Google Scholar]
- Lorenz, A.J.; Chao, S.; Asoro, F.G.; Heffner, E.L.; Hayashi, T.; Iwata, H.; Smith, K.P.; Sorrells, M.E.; LucJannink, J. Genomic selection in plant breeding: Knowledge and prospects. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: San Diego, CA, USA, 2011; Volume 110, pp. 77–123. [Google Scholar]
- Poland, J.A.; Brown, P.J.; Sorrells, M.E.; Jannink, J.L. Development of high-density genetic maps for barley and wheat using a novel two-enzyme genotyping-by-sequencing approach. PLoS ONE 2012, 7, e32253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorjanc, G.; Jenko, J.; Hearne, S.J.; Hickey, J.M. Initiating maize pre-breeding programs using genomic selection to harness polygenic variation from landrace populations. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, C.R.; Voss-Fels, K.P.; Miller, C.N.; Quan, W.; Hua, W.; Guan, C.Y.; Snowdon, R.J.; Qian, L. Effective genomic selection in a narrow-genepool crop with low-density markers: Asian rapeseed as an example. Plant Genome 2018, 11, 170084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.B.; Yang, M.H.; Zeng, F.; Biligetu, B. Searching for an accurate marker-based prediction of an individual quantitative trait in molecular plant breeding. Front. Plant Sci. 2017, 8, 1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, T.G.; Andrew, T.; Cooper, G.M.; Margulies, E.H.; Mullikin, J.C.; Balding, D.J. Functional constraint and small insertions and deletions in the ENCODE regions of the human genome. Genome Biol. 2007, 8, R180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, E.; Chu, W.C.; Ayele, M.; Ho, J.; Bruggeman, E.; Yourstone, K.; Rafalski, A.; Smith, O.S.; McMullen, M.D.; Bezawada, C.; et al. Development of single nucleotide polymorphism (SNP) markers for use in commercial maize (Zea mays L.) germplasm. Mol. Breed. 2009, 24, 165–176. [Google Scholar] [CrossRef]
- Liu, Y.; He, Z.H.; Appels, R.; Xia, X.C. Functional markers in wheat: Current status and future prospects. Theor. Appl. Genet. 2012, 125, 1–10. [Google Scholar] [CrossRef]
- Jiang, Q.; Hou, J.; Hao, C.; Wang, L.; Ge, H.; Dong, Y.; Zhang, X. The wheat (T. aestivum) sucrose synthase 2 gene (TaSus2) active in endosperm development is associated with yield traits. Funct. Integr. Genom. 2011, 11, 49–61. [Google Scholar] [CrossRef]
- Yang, Z.; Bai, Z.; Li, X.; Wang, P.; Wu, Q.; Yang, L.; Li, L.; Li, X. SNP identification and allelic-specific PCR markers development for TaGW2, a gene linked to wheat kernel weight. Theor. Appl. Genet. 2012, 125, 1057–1068. [Google Scholar] [CrossRef]
- Zhang, Y.; Miao, X.; Xia, X.; He, Z. Cloning of seed dormancy gene (TaSdr) associated with tolerance to pre-harvest sprouting in common wheat and development of a functional marker. Theor. Appl. Genet. 2014, 127, 855–866. [Google Scholar] [CrossRef]
- Ellis, M.; Spielmeyer, W.; Gale, K.; Rebetzke, J.; Richards, A. “Perfect” markers for the Rht-B1b and Rht-D1b dwarfing genes in wheat. Theor. Appl. Genet. 2002, 105, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Nakazaki, T.; Chen, S.; Chen, W.; Saito, H.; Tsukiyama, T.; Okumoto, Y.; Xu, Z.; Tanisaka, T. Identification and characterization of the erect-pose panicle gene EP conferring high grain yield in rice (Oryza sativa L.). Theor. Appl. Genet. 2009, 119, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Spielmeyer, W.; Ellis, M.; Robertson, M.; Ali, S.; Lenton, J.R.; Chandler, P.M. Isolation of gibberellin metabolic pathway genes from barley and comparative mapping in barley, wheat and rice. Theor. Appl. Genet. 2004, 109, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Raina, M.; Salgotra, R.K.; Pandotra, P.; Rathour, R.; Singh, K. Genetic enhancement for semi-dwarf and bacterial blight resistance with enhanced grain quality characteristics in traditional Basmati rice through marker-assisted selection. Comptes Rendus Biol. 2019, 342, 142–153. [Google Scholar] [CrossRef]
- Qi, Y.; Wang, L.; Gui, J.; Zhang, L.; Liu, Q.; Wang, J. Development and validation of a functional co-dominant SNP marker for the photoperiod thermo-sensitive genic male sterility pms3 (p/tms12-1) gene in rice. Breed. Sci. 2017, 67, 535–539. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Szucs, P.; Yan, L.; Helguera, M.; Skinner, J.S.; von Zitzewitz, J.; Hayes, P.M.; Dubcovsky, J. Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol. Genet. Genom. 2005, 273, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Mirlohi, S.; Li, X.; Hea, Y. Identification of functional single-nucleotide polymorphisms affecting leaf hair number in Brassica rapa. Plant Physiol. 2018, 177, 490–503. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yang, S.; Zhou, Y.; He, Z.; Xia, X. Distribution of the Rht-B1b, Rht-D1b and Rht8 reduced height genes in autumn-sown Chinese wheats detected by molecular markers. Euphytica 2006, 152, 109–116. [Google Scholar] [CrossRef]
- Su, Z.; Hao, C.; Wang, L.; Dong, Y.; Zhang, X. Identification and development of a functional marker of TaGW2 associated with grain weight in bread wheat (T. aestivum L.). Theor. Appl. Genet. 2011, 122, 211–223. [Google Scholar] [CrossRef]
- Andeden, E.; Yediay, F.; Baloch, F.; Shaaf, S.; Kilian, B.; Nachit, M.; Ozkan, H. Distribution of vernalization and photoperiod genes (Vrn-A1, Vrn-B1, Vrn-D1, Vrn-B3, Ppd-D1) in Turkish bread wheat cultivars and landraces. Cereal Res. Commun. 2011, 39, 352–364. [Google Scholar] [CrossRef]
- Feuillet, C.; Stein, N.; Rossini, L.; Praud, S.; Mayer, K.; Schulman, A.; Eversole, K.; Appels, R. Integrating cereal genomics to support innovation in the Triticeae. Funct. Integr. Genom. 2012, 12, 573–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, Y.Y.; Hong, L.Y.; Fei, T.J.; Qasim, S.M.; Zhi-Xiong, C.; Lan, W.; Jin-Quan, L.; Xiang-Dong, L.; Yong-Gen, L. Wide-compatibility gene S5n exploited by functional molecular markers and its effect on fertility of intersubspecific rice hybrids. Crop Sci. 2012. [Google Scholar] [CrossRef]
- Doebley, J.; Stec, A.; Gustus, C. Teosinte branched1 and the origin of maize: Evidence for epistasis and the evolution of dominance. Genetics 1995, 141, 333–346. [Google Scholar] [PubMed]
- Dunford, R.P.; Yano, M.; Kurata, N.; Sasaki, T.; Huestis, G.; Rocheford, T.; Laurie, D.A. Comparative mapping of the barley Phd-H1 photoperiod response gene region, which lies close to a junction between two rice linkage segments. Genetics 2002, 161, 825–834. [Google Scholar] [PubMed]
- Kole, C.; Quijada, P.; Michaels, S.D.; Amasino, R.M.; Osborn, T.C. Evidence for homology of flowering-time genes VFR2 from Brassica rapa and FLC from Arabidopsis thaliana. Theor. Appl. Genet. 2001, 102, 425–430. [Google Scholar] [CrossRef]
- Fileppi, M.; Gallsso, I.; Tagliabue, G.; Campion, B.; Doria, E.; Sparvoli, F. Characterisation of structural genes involved in phytic acid biosynthesis in common bean (Phaseolus vulgaris L.). Mol. Breed. 2010, 25, 453–470. [Google Scholar] [CrossRef]
- Hecht, V.; Foucher, F.; Ferrandiz, C.; Macknight, R.; Navarro, C.; Morin, J.; Vardy, M.E.; Ellis, N.; Beltran, J.P.; Rameau, C.; et al. Conservation of Arabidopsis flowering genes in model legumes. Plant Physiol. 2005, 137, 1420–1434. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Tang, F.; Gao, M.; Krishnan, H.B.; Zhu, H. R gene-controlled host specificity in the legume–rhizobia symbiosis. Proc. Natl. Acad. Sci. USA 2010, 107, 18735–18740. [Google Scholar] [CrossRef] [Green Version]
- Nesbitt, T.C.; Tanksley, S.D. Comparative sequencing in the genus Lycopersicon. Implications for the evolution of fruit size in the domestication of cultivated tomatoes. Genetics 2002, 162, 365–379. [Google Scholar]
- Fukino, N.; Ohara, T.; Sugiyama, M.; Kubo, N.; Hirai, M.; Sakata, Y.; Matsumoto, S. Mapping of a gene that confers short lateral branching (slb) in melon (Cucumis melo L.). Euphytica 2012, 187, 133–143. [Google Scholar] [CrossRef]
- Diaz, A.; Fergany, M.; Formisano, G.; Ziarsolo, P.; Blanca, J.; Fei, Z.; Staub, J.E.; Zalapa, J.E.; Cuevas, H.E.; Dace, G.; et al. A consensus linkage map for molecular markers and quantitative trait loci associated with economically important traits in melon (Cucumis melo L.). BMC Plant Biol. 2011, 11, 111. [Google Scholar] [CrossRef] [Green Version]
- Naresh, P.; Lin, S.; Lin, C.; Wang, Y.; Schafleitner, R.; Kilian, A.; Kumar, S. Molecular markers associated to two non-allelic genic male sterility genes in peppers (Capsicum annuum L.). Front. Plant Sci. 2018, 9, 1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogales, A.; Nobre, T.; Gardoso, H.G.; Munoz-Sanhueza, L.; Valadas, V.; Campos, M.D.; Arnholdt-Schmitt, B. Allelic variation on DcAOX1 gene in carrot (Daucus carota L.): An interesting simple sequence repeat in a highly variable intron. Plant Gene 2016, 5, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Sheng, X.; Yu, H.; Wang, J.S.; Shen, Y.S.; Gu, H.H. Identification of QTLs associated with curd architecture in cauliflower. BMC Plant Biol. 2020, 20, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abuyusuf, M.; Nath, U.K.; Kim, H.; Islam, R.M.; Park, J.I.; Nou, I.S. Molecular markers based on sequence variation in BoFLC1.C9 for characterizing early- and late-flowering cabbage genotypes. BMC Genet. 2019, 20, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Zhuang, M.; Fang, Z.; Wang, Q.; Zhang, Y.; Liu, Y.; Yang, L.; Cheng, F. A co-dominant marker BoE332 applied to marker-assisted selection of homozygous male-sterile plants in cabbage (Brassica oleracea var. capitata L.). J. Integr. Agric. 2013, 12, 596–602. [Google Scholar] [CrossRef]
- Kim, S.; Jung, Y.; Seong, K.; Kim, S.C.; Jung, Y.H.; Kim, M.; Jang, K.C.; Song, E.Y.; Seong, K.C.; Um, Y.C. Development of a SCAR marker for sex identification in asparagus. Korean J. Plant Res. 2014, 27, 236–241. [Google Scholar] [CrossRef]
- Zhao, X.L.; Ma, E.W.; Gale, E.K.R.; Lei, Z.S.; He, Z.H.; Sun, Q.X.; Xia, X.C. Identification of SNPs and development of functional markers for LMW-GS genes at Glu-D3 and Glu-B3 loci in bread wheat (Triticum aestivum L.). Mol. Breed. 2007, 20, 223–231. [Google Scholar] [CrossRef]
- He, X.; Zhang, Y.; He, Z.; Wu, Y.P.; Xiao, Y.G.; Ma, C.X.; Xia, X.C. Characterization of phytoene synthase 1 gene (Psy1) located on common wheat chromosome 7A and development of a functional marker. Theor. Appl. Genet. 2008, 116, 213–221. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, X.; He, Z.; Ma, W.; Appels, R.; Pena, R.J.; Xia, X.C. Characterization of low molecular-weight glutenin subunit Glu-B3 genes and development of STS markers in common wheat (T. aestivum L.). Theor. Appl. Genet. 2009, 118, 525–539. [Google Scholar] [CrossRef]
- Zhang, C.; Dong, C.; He, X.; Zhang, L.; Xia, X.; Zhonghu, H. Allelic variants at the TaZds-D1 locus on wheat chromosome 2DL and their association with yellow pigment content. Crop Sci. 2011, 51, 1580–1590. [Google Scholar] [CrossRef]
- Geng, H.; Xia, X.; Zhang, L.; Qu, Y.; He, Z. Development of functional markers for Lipoxygenase gene Talox-B1 on chromosome 4 BS in common wheat. Crop Sci. 2012, 52, 568–576. [Google Scholar] [CrossRef]
- Shi, W.; Yang, Y.; Chen, S.; Xu, M. Discovery of a new fragrance allele and the development of functional markers for the breeding of fragrant rice varieties. Mol. Breed. 2008, 22, 185–192. [Google Scholar] [CrossRef]
- Amarawathi, Y.; Singh, R.; Singh, A.K.; Singh, V.P.; Mohapatra, T.; Sharma, T.R. Mapping of quantitative trait loci for basmati quality traits in rice (Oryza sativa L.). Mol. Breed. 2008, 21, 49–65. [Google Scholar] [CrossRef]
- Sakthivel, K.; Sundaram, R.M.; Rani, N.S.; Balachandran, S.M.; Neereja, C.N. Genetic and molecular basis of fragrance in rice. Biotechnol. Adv. 2009, 27, 468–473. [Google Scholar] [CrossRef]
- Chen, T.; Meng-xiang, T.; Zhang, Y.; Zhu, Z.; Zhao, L.; Zhao, Q.; Lin, J.; Zhou, L.; Wang, C. Development of simple functional markers for low glutelin content gene 1 (Lgc1) in rice (Oryza sativa). Rice Sci. 2010, 17, 173–178. [Google Scholar] [CrossRef]
- Anuradha, K.; Agarwal, S.; Rao, Y.V.; Rao, K.V.; Viraktamath, B.C.; Sarla, N. Mapping QTLs and candidate genes for iron and zinc concentrations in unpolished rice of Madhukar × Swarna RILs. Gene 2012, 508, 233–240. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, S.; Yang, G.; Zha, W.; Cai, H.; Li, S.; Chen, Z.; Liu, K.; Xu, H.; You, A. A perfect functional marker for the gene of intermediate amylose content Wx-in in rice (Oryza sativa L.). Crop Breed. Appl. Biotechnol. 2018, 18, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Chai, Y.; Hao, X.; Yang, X.; Allen, W.B.; Li, J.; Yan, J.; Shen, B.; Li, J. Validation of DGAT1-2 polymorphisms associated with oil content and development of functional markers for molecular breeding of high-oil maize. Mol. Breed. 2012, 29, 939–949. [Google Scholar] [CrossRef]
- Zhou, Y.; Han, Y.; Li, Z.; Fu, Y.; Fu, Z.; Xu, S.; Li, J.; Yan, J.; Jang, X. ZmcrtRB3 encodes a carotenoid hydroxylase that affects the accumulation of a-carotene in maize kernel. J. Integr. Plant Biol. 2012, 54, 260–269. [Google Scholar] [CrossRef]
- Chhabra, R.; Hossain, F.; Muthusamy, V.; Baveja, A.; Mehta, B.K.; Zunjare, R.U. Development and validation of breeder-friendly functional markers of sugary1 gene encoding starch-debranching enzyme affecting kernel sweetness in maize (Zea mays). Crop Pasture Sci. 2019, 70, 868–875. [Google Scholar] [CrossRef]
- Obeng-Bio, E.; Badu-Apraku, B.; Elorhor Ifie, B.; Danquah, A.; Blay, E.T.; Dadzie, M.A. Phenotypic characterization and validation of provitamin A functional genes in early maturing provitamin A-quality protein maize (Zea mays) inbred lines. Plant Breed. 2019. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Nie, Y.D.; Han, F.X.; Zhao, X.N.; Dun, B.Q.; Lu, M.; Li, G. Allelic variation of a soluble acid invertase gene (SAI-1) and development of a functional marker in sweet sorghum (Sorghum bicolor (L.). Mol. Breed. 2014, 33, 721–730. [Google Scholar] [CrossRef]
- Zanan, R.; Khandagale, K.; Hinge, V.; Elangovan, M.; Henry, R.J.; Nadaf, A. Characterization of fragrance in sorghum (Sorghum bicolor L.) Moench) grain and development of a gene-based marker for selection in breeding. Mol. Breed. 2016, 36. [Google Scholar] [CrossRef]
- Juwattanasomran, R.; Somta, P.; Kaga, A.; Chankaew, S.; Shimizu, T.; Sorajjapinun, W.; Srinives, P. Identification of a new fragrance allele in soybean and development of its functional marker. Mol. Breed. 2010. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Staub, J.E.; Simon, P.W.; Zalapa, J.E. A consensus linkage map identifies genomic regions controlling fruit maturity and beta-carotene-associated flesh color in melon (Cucumis melo L.). Theor. Appl. Genet. 2009, 119, 741–756. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; He, L.; Xu, S.; Wan, Y.; Wang, H.; Wang, Y.; Yu, L.; Zhu, W. Expression analysis, functional marker development and verification of AgFNSI in celery. Sci. Rep. 2020, 10, 531. [Google Scholar] [CrossRef]
- Cheng, T.; Zheng, X.; Xie, K.; Liu, J.; Zheng, X.; Jin, S.; Diao, Y.; Hu, Z.; Wang, J. Development and identification of three functional markers associated with starch content in lotus (Nelumbo nucifera). Sci. Rep. 2020, 10, 4242. [Google Scholar] [CrossRef]
- Saritnum, O.; Minami, M.; Matsushima, K.; Minamiyama, Y.; Hirai, M.; Baba, T.; Bansho, H.; Nemoto, K. Inheritance of few-pungent trait in chilli pepper ‘S3212’ (Capsicum frutescens). J. Jpn. Soc. Hort. Sci. 2008, 77, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, N.; Torres, A.M. Characterization and diagnostic marker for TTG1 regulating tannin and anthocyanin biosynthesis in faba bean. Sci. Rep. 2019, 9, 16174. [Google Scholar] [CrossRef] [Green Version]
- Gemenet, D.C.; Kitavi, M.N.; David, M. Development of diagnostic SNP markers for quality assurance and control in sweetpotato [Ipomoea batatas (L.) Lam.] breeding programs. PLoS ONE 2020, 15, e0232173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arizio, C.M.; Costa Tártara, S.M.; Manifesto, M.M. Carotenoids gene markers for sweetpotato (Ipomoea batatas L. Lam): Applications in genetic mapping, diversity evaluation and cross-species transference. Mol. Genet. Genom. 2014. [Google Scholar] [CrossRef]
- Tommasini, L.; Yahiaoui, N.; Srichumpa, P.; Keller, B. Development of functional markers specific for seven Pm3 resistance alleles and their validation in the bread wheat gene pool. Theor. Appl. Genet. 2006, 114, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Periyannan, S.; Bansal, U.; Bariana, H.; Deal, K.; Luo, M.; Dvorak, J.; Lagudah, E. Identification of a robust molecular marker for the detection of the stem rust resistance gene Sr45 in common wheat. Theor. Appl. Genet. 2014, 127, 947–955. [Google Scholar] [CrossRef]
- Hur, Y.J.; Jeung, J.; Kim, S.Y.; Park, H.; Cho, J.; Lee, J.Y.; Sohan, Y.; Song, Y.C.; Park, D.; Lee, C.; et al. Functional markers for bacterial blight resistance gene Xa3 in rice. Mol. Breed. 2013, 31, 981–985. [Google Scholar] [CrossRef]
- Hajira, S.K.; Sundaram, R.M.; Laha, G.S.; Yugander, A.; Balachandran, S.M.; Viraktamath, B.C.; Sujatha, K.; Balachiranjeevi, C.H.; Pranathi, K.; Anila, M.; et al. A single-tube, functional marker-based multiplex PCR assay for simultaneous detection of major bacterial blight resistance genes Xa21, xa13 and xa5 in rice. Rice Sci. 2016, 23, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, R.M.; Vishnupriya, M.R.; Biradar, S.K.; Laha, G.S.; Reddy, G.A.; Shobha, R.N.; Sarma, N.P.; Sonti, R.V. Marker-assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 2008, 80, 411–422. [Google Scholar] [CrossRef]
- Song, W.Y.; Pi, L.Y.; Wang, G.L.; Gardner, J.; Holsten, T.; Ronald, P.C. Evolution of the rice Xa21 disease resistance gene family. Plant Cell 1997, 9, 1279–1287. [Google Scholar]
- Ellur, R.K.; Khanna, A.; Gopala Krishnan, S.; Bhowmick, P.K.; Vinod, K.K.; Nagarajan, M.; Mondal, K.K.; Singh, N.K.; Singh, K.; Prabhu, K.B.; et al. Marker-aided incorporation of Xa38, a novel bacterial blight resistance gene, in PB1121 and comparison of its resistance spectrum with xa13 + Xa21. Sci. Rep. 2016, 6, 29188. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, Z.; Lang, X.; Du, B.; Liu, K.; Yang, G.; Hu, G.; Li, S.; He, G.; You, A. Development and validation of a PCR-based functional marker system for the brown planthopper resistance gene Bph14 in rice. Breed. Sci. 2013, 63, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Yasuda, N.; Fujita, Y.; Koizumi, S.; Yoshida, H. Identification of the blast resistance gene Pit in rice cultivars using functional markers. Theor. Appl. Genet. 2010, 121, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, G.; Srinivasarao, K.; Mohan, K.M.; Sudarshan, I.; Sivaranjani, A.K.P.; Gopalkrishna, K.; Neeraja, C.N.; Balachandran, S.M.; Sundaram, S.M.; Prasad, M.S.; et al. Development and validation of functional marker targeting an InDel in the major rice blast disease resistance gene Pi54 (Pikh). Mol. Breed. 2011, 27, 129–135. [Google Scholar]
- Ma, J.; Ma, X.D.; Zhao, Z.C.; Wang, S.; Wang, J.L.; Wang, J.; Cheng, Z.J.; Lei, C.L. Development and application of a functional marker of the blast resistance gene Pi35 in rice. Acta Agron. Sin. 2015, 41, 1779–1790. [Google Scholar]
- Quan, W.F.; Hui, C.Z.; Yang, X.U.; Wang, J.; Li, W.; Fan, F.; Chen, L.; Tao, Y.; Zhong, W.; Yang, J. Development and application of the functional marker for the broad-spectrum blast resistance gene PigmR in rice. Sci. Agric. Sin. 2019, 52, 955–967. [Google Scholar]
- Wang, Y.; Jiang, W.; Liu, H.; Zeng, Y.; Du, B.; Zhu, L.; He, G.; Chen, R. Marker assisted pyramiding of Bph6 and Bph9 into elite restorer line 93–11 and development of functional marker for Bph9. Rice 2017, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Madsen, L.H.; Collins, N.C.; Rakwalska, M.; Backes, G.; Sandal, N.; Krusell, L.; Jensen, J.; Waterman, E.H.; Jahoor, A.; Ayliffe, M. Barley disease resistance gene analogs of the NBSLRR class: Identification and mapping. Mol. Genet. Genom. 2003, 269, 150–161. [Google Scholar] [CrossRef]
- Singh, K.B.; Foley, R.C.; Sanchez, L.O. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Brunner, S.; Keller, B.; Feuillet, C. A large rearrangement involving genes and low copy DNA interrupts the micro-collinearity between rice and barley at the Rph7 locus. Genetics 2003, 164, 673–683. [Google Scholar] [PubMed]
- Nevame, A.Y.M.; Xia, L.; Nchongboh, C.G.; Hasan, M.M.; Alam, M.A.; Yongbo, L.; Wenting, Z.; Yafei, H.; Emon, R.M.; Ismail, M.R.; et al. Development of a new molecular marker for the resistance to tomato yellow leaf curl virus. BioMed Res. Int. 2018. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.; Hwang, I.S.; Lee, H.J.; Lee, J.M.; Seo, E.; Choi, D.; Oh, C. Identification of a molecular marker tightly linked to bacterial wilt resistance in tomato by genome-wide SNP analysis. Theor. Appl. Genet. 2018, 131, 1017–1030. [Google Scholar] [CrossRef]
- Devran, Z.; Kahveci, E.; Hong, Y.; Studholme, D.J.; Tör, M. Identifying molecular markers suitable for Frl selection in tomato breeding. Theor. Appl. Genet. 2018, 131, 2099–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oumouloud, A.; Otmani, M.E.; Alvarez, J. Molecular characterization of Fom-1 gene and development of functional markers for molecular breeding of resistance to Fusarium race 2 in melon. Euphytica 2015. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.F.; Hoy, J.W.; Kinbeng, C.A.; Baisakh, N. Identification of genomic regions controlling leaf scald resistance in sugarcane using a bi-parental mapping population and selective genotyping by sequencing. Front. Plant Sci. 2018, 9, 877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Ren, Y.; Guo, S.; Zhang, H.; Gong, G.; Du, Y.; Xu, Y. Application of comparative genomics in developing markers tightly linked to the Pm-2F gene for powdery mildew resistance in melon (Cucumis melo L.). Euphytica 2012, 190, 157–168. [Google Scholar] [CrossRef]
- Nieto, C.; Morales, M.; Orjeda, G.; Clepet, C.; Monfort, A.; Sturbois, B.; Puigdomenech, P.; Pitrat, M.; Caboche, M.; Dogimont, C.; et al. An eIF4E allele confers resistance to an uncapped and non-polyadenylated RNA virus in melon. Plant J. 2006, 48, 452–462. [Google Scholar] [CrossRef]
- Ling, K.S.; Harris, K.R.; Meyer, J.D.F.; Levi, A.; Guner, N.; Wehner, T.C.; Bendahmane, A.; Havey, M.J. Non-synonymous single nucleotide polymorphisms in the watermelon eIF4E gene are closely associated with resistance to Zucchini yellow mosaic virus. Theor. Appl. Genet. 2009, 120, 191–200. [Google Scholar] [CrossRef]
- Gama, R.N.C.; Santos, C.A.F.; Dias, R.C.S.; Souza, R.R.C.; Queiróz, M.A. Microsatellite markers linked to powdery mildew resistance locus in watermelon. Aust. J. Crop Sci. 2015, 9, 92–97. [Google Scholar]
- Liu, P.N.; Miao, H.; Lu, H.W.; Cui, J.Y.; Tian, G.L.; Wehner, T.C.; Gu, X.F.; Zhang, S.P. Molecular mapping and candidate gene analysis for resistance to powdery mildew in Cucumis sativus stem. Genet. Mol. Res. 2017, 16, 16039680. [Google Scholar] [CrossRef]
- Shi, L.; Yang, Y.; Xie, Q.; Miao, H.; Bo, K.; Song, Z.; Wang, Y.; Xie, B.; Zhang, S.; Gu, X. Inheritance and QTL mapping of cucumber mosaic virus resistance in cucumber. PLoS ONE 2018, 13, e0200571. [Google Scholar] [CrossRef]
- Cao, B.H.; Lei, J.J.; Wang, Y.; Chen, G.J. Inheritance and identification of SCAR marker linked to bacterial wilt-resistance in eggplant. Afr. J. Biotechnol. 2009, 8, 5201–5207. [Google Scholar]
- Karna, S.; Ahn, Y. K Development of InDel markers to identify Capsicum disease resistance using whole genome resequencing. J. Plant Biotechnol. 2018, 45, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lu, H.; Hu, J. Molecular mapping to bacterial leaf spot in lettuce PI358001-1. Genet. Resist. 2016. [Google Scholar] [CrossRef]
- Li, G.; Qian, W.; Zhang, S.; Zhang, S.; Li, F.; Zhang, H.; Wu, J.; Wang, X.; Sun, R. Development of gene-based markers for the Turnip mosaic virus resistance gene retr02 in Brassica rapa. Plant Breed. 2016, 135. [Google Scholar] [CrossRef]
- Sun, S.; Deng, D.; Wang, Z.; Duan, C.; Wu, X.; Wang, X.; Zong, X.; Zhu, Z. A novel er1 allele and the development and validation of its functional marker for breeding pea (Pisum sativum L.) resistance to powdery mildew. Theor. Appl. Genet. 2016, 129, 909–919. [Google Scholar] [CrossRef]
- Maiti, S.; Basak, J.; Kundagrami, S.; Kundu, A.; Pal, A. Molecular marker-assisted genotyping of mungbean yellow mosaic India virus resistant germplasms of mungbean and urdbean. Mol. Biotechnol. 2011, 47, 95–104. [Google Scholar] [CrossRef]
- Lan, N.T.N.; Thao, H.T.; Son, L.V.; Mau, C.H. Overexpression of VrPDF1 gene confers resistance to weevils in transgenic mung bean plants. PeerJ Prepr. 2017. [Google Scholar] [CrossRef]
- Garg, B.; Lata, C.; Prasad, M. A study of the role of gene TaMYB2 and an associated SNP in dehydration tolerance in common wheat. Mol. Biol. Rep. 2012, 39, 10865–10871. [Google Scholar] [CrossRef]
- Pandey, B.; Sharma, P.; Pandey, D.; Sharma, I.; Chatrath, R. Identification of new aquaporin genes and single nucleotide polymorphism in bread wheat. Evol. Bioinform. 2013, 9, 437–452. [Google Scholar] [CrossRef]
- Huseynova, I.M. Application of PCR-based functional markers for identification of DREB1 genes in Triticum aestivum L. SF Biotechnol. Bioeng. J. 2018, 1, 1. [Google Scholar]
- Hamdi, K.; Brini, F.; Kharrat, N.; Masmoudi, K.; Yakoubi, I. Abscisic acid, stress, and ripening (TtASR1) gene as a functional marker for salt tolerance in durum wheat. BioMed J. Int. 2020. [Google Scholar] [CrossRef] [Green Version]
- Neeraja, C.; Maghirang-Rodriguez, R.; Pamplona, A.; Heuer, S.; Collard, B.C.Y.; Septiningsih, E.M.; Vergara, G.; Sanchez, D.; Xu, K.; Ismail, A.M.; et al. A marker-assisted backcross approach for developing submergence tolerant rice cultivars. Theor. Appl. Genet. 2007, 115, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Chin, J.H.; Gamuyao, R.; Dalid, C.; Bustamam, M.; Prasetiyono, J.; Moeljopawiro, S.; Wissuwa, M.; Heuer, S. Developing rice with high yield under phosphorus deficiency: Pup1 sequence to application. Plant Physiol. 2011, 156, 1202–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, D.; Wang, H.; Liang, G.; Yu, D. OsSAPK2 Confers abscisic acid sensitivity and tolerance to drought stress in rice. Front. Plant Sci. 2017, 8, 993. [Google Scholar] [CrossRef] [Green Version]
- Assenov, B.; Andjelkovic, V.; Ignjatovic-Micic, D.; Pagnotta, M.A. Identification of SNP mutations in In MYBF-1 gene involved in drought stress tolerance in maize. Bulg. J. Agric. Sci. 2013, 19, 181–185. [Google Scholar]
- Too, E.J.; Onkware, A.O.; Were, B.A.; Gudu, S.; Carlsson, A.; Geleta, M. Molecular markers associated with aluminium tolerance in Sorghum bicolor. Hereditas 2018, 155, 20. [Google Scholar] [CrossRef]
- Contour-Ansel, D.; Torres-Franklin, M.L.; Cruz, D.E.; Lameta, A.; Fodil, F.Z. Glutathione reductase in leaves of cowpea: Cloning of two cDNAs, expression and enzymatic activity under progressive drought stress desiccation and abscisic acid treatment. Ann. Bot. 2006, 98, 1279–1287. [Google Scholar] [CrossRef] [Green Version]
- Lata, C.; Prasad, M. Validation of an allele-specific marker associated with dehydration stress tolerance in a core set of foxtail millet accessions. Plant Breed. 2013, 132, 496–499. [Google Scholar] [CrossRef]
- Leonforte, A.; Sudheesh, S.; Cogan, N.O.; Salisbury, P.A.; Nicolas, M.E.; Materne, M.; Forster, J.W.; Kaur, S. SNP marker discovery, linkage map construction and identification of QTLs for enhanced salinity tolerance in field pea (Pisum sativum L.). BMC Plant Biol. 2013, 13, 161. [Google Scholar] [CrossRef] [Green Version]
- Ramkumar, G.; Sivaranjani, A.; Pandey, M.K.; Sakthivel, K.; Shobha, N.R.; Sudarshan, I.; Prasad, G.S.V.; Neeraja, C.N.; Sundaram, R.M.; Viraktamath, B.C.; et al. Development of a PCR-based SNP marker system for effective selection of kernel length and kernel elongation in rice. Mol. Breed. 2010, 26, 735–740. [Google Scholar] [CrossRef]
- Randhawa, M.S.; Bains, N.S.; Sohu, V.S.; Chhuneja, P.; Trethwon, R.M.; Bariana, H.S.; Banasl, U. Marker assisted transfer of stripe rust and stem rust resistance genes into four wheat cultivars. Agronomy 2019, 9, 497. [Google Scholar] [CrossRef] [Green Version]
- Lagudah, E.S.; Krattinger, S.G.; Herrera-Foessel, S.; Singh, R.P.; Huerta-Espino, J.; Spielmeyer, W.; Brown-Guedira, G.; Selter, L.L.; Keller, B. Gene specific markers for the wheat gene Lr34/Yr18/Pm38 which confers resistance to multiple fungal pathogens. Theor. Appl. Genet. 2009, 119, 889–898. [Google Scholar] [CrossRef] [Green Version]
- Yugander, A.; Sundaram, R.M.; Singh, K.; Ladhalakshmi, D.; Subba Rao, L.V.; Madhav, M.S.; Badri, J.; Prasad, M.S.; Laha, G.S. Incorporation of the novel bacterial blight resistance gene Xa38 into the genetic background of elite rice variety ‘Improved Samba Mahsuri’. PLoS ONE 2018, 13, e0198260. [Google Scholar] [CrossRef]
- Joseph, M.; Gopalakrishnan, S.; Sharma, R.K.; Singh, V.P.; Singh, A.K.; Singh, N.K.; Mohapatra, T. Combining bacterial blight resistance and Basmati quality characteristics by phenotypic and molecular marker-assisted selection in rice. Mol. Breed. 2004, 13, 377–387. [Google Scholar] [CrossRef]
- Das, G.; Rao, G.J.N.; Varier, M.; Prakash, A.; Prasad, D. Improved Tapaswini having four BB resistance genes pyramided with six genes/QTLs, resistance/ tolerance to biotic and abiotic stresses in rice. Sci. Rep. 2018, 8, 2413. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Gene (s) | Chromosomal Location | Sequence | Crop | References |
|---|---|---|---|---|---|
| Agronomic traits | |||||
| Semi-dwarf stature | Rht-B1 and Rht-D1 | 4B, 4D | F-TCTCCTCCCTCCCCACCCCAAC R-CCATGGCCATCTCGAGCTGC & F-CGCGCAATTATTGGCCAGAGATAG R-CCCCATGGCCATCTCGAGCTGCTA | Wheat | [129] |
| Grain weight | TaSus2-2B | 2 | F-CGCCCTGAGCCG CATCCACA R-CGCTCGCCCGC CATTTATTTCTCT | Wheat | [118] |
| Grain weight | TaGW2 | 6 | F-ATGGGGAACAGAATAGGAGGGAGGA R-CGAGTATGCCTAGAATGGAAAGAC | Wheat | [130] |
| Photoperiod response | Phd-H1 | 2 | F-ACGCCTCCCACTACACTG R-CACTGGTGGTAGCTGAGATT | Wheat | [131] |
| Vernalization | Vrn-D4 | 5 | F-CATAATGCCAAGCCGGTGAGTAC R-ATGTCTGCCAATTAGCTAGC | Wheat | [132] |
| Semi-dwarf | sd1 | 1 | F-CACGCACGGGTTCTTCCAGGTG R-AGGAGAATAGGAGATGGTTTACC | Rice | [123] |
| Wide-compatibility gene | S5n | 6 | F-CGTCTTGCTTCTTCATTCCC R-GTAGGTAAACACAGGCAGAG | Rice | [133] |
| Photoperiod-thermo-sensitive genic male (PGMS and TGMS) sterility | pms3 (p/tms12-1) | 12 | F-GAATGCCATCTAAACACT R-ATTTTACTCTTGATGGATGGTC | Rice | [126] |
| Plant stature | tb1 | 1 | F-CACATGAGCCCATGCCTCTC R-AAAGCGGTAAGTCCATGGGG | Maize | [134] |
| Plant height | Dwarf8 | 1 | F-ACACTATCACCGCTCTATTG R-ACTCTTTCCCTGACTTCATT | Maize | [31] |
| Photoperiod response | Phd-H1 | 7 | F-CCTCTTCGCTATTAC GCCAG R –GCCCTTCCCAACAGTTGCG | Barley | [135] |
| Vernalization requirements | VRN-H 1 | 5 | F-TTCATCATGGATCGCCAGTA R-AAAGCTCCTGCCAACTACGA | Barley | [127] |
| Vernalization requirements | VFR2 | A8 | F-CTCGTAGCCCCGAGAACATC R-ACTCAAGCAACTTACCAAGTGGA | Brassica | [136] |
| Leaf hair number | BrpHL1 | A9 | F-TACTCCTCGTTCCCTCTGGG R-GGGGGAAATGCAGATTCCGA | Brassica | [128] |
| Seed development | PvMIPSs and PvMIPSv | 1 | F-TTGCCACGCACCTGCTAATA R-CCTGCAGCTGCGATTTTCAA | Common bean | [137] |
| Controlling flowering time | MADS-box, Constants and Flowering locus T/Terminal Flower1 | 16 | F-ATGCACCTAGCCCAAGTGAC R-TGTTTGCATTCATGGCGTGT & R-ATCTGTTGTGCCGGGAATGT F-AACCGAAATGCAAAACAGGTGA | Pea, soybean, and burclover | [138] |
| Nodulation formation | Rj2 and Rfg1 | 16, 3 | F-AAGTCTTAAATTGTGTTTGGATGGA R-TGAGAATTGTCACCACCGGG & F-AAGTCTTAAATTGTGTTTGGATGGA R-TGAGAATTGTCACCACCGGG | Soybean | [139] |
| Fruit size | w2.2 | 2 | F-TCTGCTCAGAAGCATGCACA R-TTGTGACCTGTACCCCAGGA | Tomato | [140] |
| Short lateral branching | slb | 11 | F-CTTGCGCTCCTTGGTATTCC R-CAAGATCGGCAAGAGACAGC | Melon | [141] |
| Sutures on the rind | s-2 | 9 | F-GCATCGGAATCTTGTTCGGC R-TCCGGTGGGAGATACCCAAT | Melon | [142] |
| Male sterility | ms3 | 5 | F-GGTACTTTGA CCCTCATAATTGG R-TTGTTTGT GGTGTACG TGCT | Capsicum | [143] |
| Alternative respiration | DcAOX1 | 1 | F-AAAATAACAATGATGATGACACG R-CTCCACTTCAGTGATATCCAA | Carrot | [144] |
| Curd architecture | qCS.C6–1 and qCS.C6–2 | 6C | F-CGGTACTGGAATGTGGACGT R-TGAATTGGTATGAACACGCCTC | Cauliflower | [145] |
| Early and late flowering | BoFLC1.C9 | Unmapped | F-GGAAAGCAACATGGTGATGA R-CATGGTGTGAACCAGAGTCC | Cabbage | [146] |
| Male sterility | CDMs399-3 | 7C | F-TCCCTTTCACATCGTCCACA R-TGCAGCCCAGAACAGTGATA | Cabbage | [147] |
| Sex identification | MYB35 | 5 | F-TTGCTTGGCGGATCATATTATG R-TTGCTTGGCGATGTCCCTTTTG | Asparagus | [148] |
| Quality traits | |||||
| Low molecular weight glutenin | Glu-D3 and Glu-B3 | 1D | F-CAGCTAAACCCATGCAAGC R-CAATGGAAGTCATCACCTCAA | Wheat | [149] |
| Yellow pigment content | Psy1 | 7A | F-ACATGCCGCTACTCCTATCC R-GTAGAGTGGCCAGACAAGGT | Wheat | [150] |
| Low molecular weight glutenin | Glu-B3 | 1B | F-ACAACAGGTTCAGGGTTCCA R-GCTATTTGGTGTGGCTGCAA | Wheat | [151] |
| Yellow pigment content | TaZds-D1 | 2D | F-ACATAGTCCTGACCGCCAAA R-AGAGTTGCTCCTTCCATGCT | Wheat | [152] |
| Lipoxygenase gene | Talox-B1 | 4B | F-ATGATACTGGGCGGGCTCGT R-TCAGATGGAGATGCTGTTGGG | Wheat | [153] |
| Fragrance | badh2 | 8 | F-AGTTATGGTCTGGCTGGTGC R-TTGTGTGCTACCCACCCTTC | Rice | [154] |
| Fragrance | nksbad2 | 4 | F-ATGGCAACATGGAAGGTAGC R-CATCAGCAAGCTCCAAACAA | Rice | [155] |
| Fragrance | BADEX7-5 | 8 | F-TTAGGTTCTGAAGCCGGTGC R-TCCCAGTAAATGCAACCTAACAGA | Rice | [156] |
| Low glutenin content | Lgc1 | 10 | F-TTCTACAATGAAGGCGATGC R-CTGGGCTTTAACGGGACT & F-ACCGTGTTATGGCAGTTT R-ATTCAAGGGCTATCGTCT | Rice | [157] |
| Fe and Zn | OsNAS3, OsNRAMP1 | 7 | F-TCCATCGCTTGCTACCTCAC R-CCCGGAGATCGATCGAGACA & F-AGCACTCCCCCATCAATCAA R-ACTACACGGGTGGCTCTTTG | Rice | [158] |
| Intermediate amylose content | Wx-in | 6 | F-CAGCGTCGACGTAAGCCTAT R-CAGGCCCCTGAAATCCATGT | Rice | [159] |
| Oil content | DGAT1-2 | 6 | F-TGGCTCTGCAATCAGGAGAA R-TGAAGCAGCAAACAACGAGC | Maize | [160] |
| Forage quality for digestibility | Bm3 | 4 | F-TTCAACAAGGCGTACGGGAT R-AGTGGTTCTTCATGCCCTCG | Maize | [96] |
| Provitamin A | ZmcrtRB3 | 2 | F-GTCGGTACTGGCAAGTGGAA R-TAGTACGTGGCCATGACGTG | Maize | [161] |
| Sweetness | sugary1 | 4 | F-TCCCGACTTCAGAACGGTTG R-ACAACAGAGCAACCCCAACA | Maize | [162] |
| Provitamin A | crtRB1 and LcyE | 10,8 | F-CACAGGTCGCTGCGTACTTA R-GGGAGACAGCTCACAGGAAC & F-CAGTGCGCTGAAGGCTACTA R-GGATGAAAGGGTCGAGCCAA | Maize | [163] |
| Soluble acid invertase | SAI-1 | 4 | F-GGATTCCACTTCCAGCCACA R-CGACGGGGTAGAAGTCGATG | Sorghum | [164] |
| Fragrance | SbBADH2 | 4 | F-CGCAGTAGTGGAGTGGTTGT R-ACTGTGGCGGTTCTTGCATA | Sorghum | [165] |
| Fragrance allele | Gmbadh2-1 and Gmbadh2-2 | 5 | F-GTGATCTGCGAGGGAGGGAG R-TGAGTTGCAGGCAGTGTCAT | Soybean | [166] |
| White flesh | wf | 9 | F-TTGGAGGTTCAATGCTTGCC R-CAAAGACCAGAGCACCATCG | Melon | [167] |
| Green flesh color | gf | 8 | F-TCTGCAAAATGGTTGCTTTGAA R-AGGTGGATGTGGCACACAAA | Melon | [141] |
| Flavonoids | AgFNSI | 4 | F-ATGGCTCCATCAAC TATAAC R-CTGCCCTGGCAATCTCCG | Celery | [168] |
| Starch content | NnHXK and NnGBSS | Unmapped | F-TCTAAATCCCAATCCGTCC R-GCACGAACTCTTGGCAATC | Lotus | [169] |
| Pungency | Pup1 | 2 | F-CCATGGATTGTTGCTCGGGCCTCC R-CCGTACCGCCCCATTGCGATTCC | Chilli | [170] |
| Anthocyanin biosynthesis | VfTTG1 | Unmapped | F-TATGAATTCATTTTTAGTTCCCACCTAAC R-GTATCCGGTTGAGGACTCTCATAGATA | Faba bean | [171] |
| β-Carotene & Flesh | QA/QC | 3 | F-AGTGCGGGACAAGATGATCA R-TCCCGAACATCTGAGCAAGT | Sweet potato | [172] |
| Carotenoids | b_CHYβ-1 | Unmapped | F-TCCAGCTTGGGAATTACGTC R-ACAACGAAGCGTGCCATAG | Sweet potato | [173] |
| Biotic stresses | |||||
| Powdery mildew | Pm3 | 1A | F-CAAGTACCAACCACAGCCAC R-CCATTGCAACCACAGGAACA | Wheat | [174] |
| Stem rust resistance | Sr45 | 1D | F-GTCCATTTTACGACGGTCCG R-CTGGTCGGTAGGGAAGCTAG | Wheat | [175] |
| Bacterial blight resistance | Xa3 | 11 | F-GAATGGGTGGGGTTGGGAAG R-CCATGCACGCTTGTCGAATC | Rice | [176] |
| Bacterial blight resistance | xa5 | 5 | F-ACGGAGTTGCAATGTTGCTG R-GGCCAGGAGTAAAGCGGATT | Rice | [177] |
| Bacterial blight resistance | xa13 | 8 | F-GGCCATGGCTCAGTGTTTAT R-GAGCTCCAGATCTCCAAATG | Rice | [178] |
| Bacterial blight resistance | Xa21 | 11 | F-AGACGCGGAAGGGTGGTTCCCGG R-AGACCGGTAATCGAAAGATGAAA | Rice | [179] |
| Bacterial blight resistance | Xa38 | 4 | F-TCTTCTATTGCTAACATTGGTG R-AGCGTAAGTAAAAGTCTC | Rice | [180] |
| Brown plant hopper resistance | Bph14 | 3 | F-CAATCCGAGCTTACGTGGTG R-GGTGGAGAAGGCAAGAGTCT | Rice | [181] |
| Blast resistance | Pit | 1 | F-GTGACGGAAGTGCATGGGTA R-ACCAGGGAACCCGACAAGAA | Rice | [182] |
| Blast resistance | Pi54 (Pikh) | 11 | F-CCTCTTGAGTTGAATTGGCACG R-CCTCGTGCAGCTGTTTTCAC | Rice | [183] |
| Blast resistance | Pi35 | 1 | F-TCCATGGCGGAGGTGGTGTTGGCTG R-AGAGCAAATCTTGGGGTGTCTGCAA | Rice | [184] |
| Blast Resistance | PigmR | 6 | F-ATGTCGGAGGAAGCAGGTC R-ATGTCACGCAGCAAAACCAT | Rice | [185] |
| Brown plant hopper resistance | Bph9 | 12 | F-CACTCGCACGGATACAATGG R-GATCGTGACACATGCATGCT | Rice | [186] |
| Powdery mildew | NBS–LRR class of resistance genes | 2 | F-CGTTTTGTATGGCGTCCGAT R-TTGTCGCTGAGGTCCATCTT | Barley | [187] |
| General stress response | ERF transcription factors | 1 | F-ACAGTGGTGGCAAGTGTGAA R-ACGGCCTCCTTCTTACTCCT | Several crops | [188] |
| Leaf rust resistance | Rph7 | 3H | F-TGGAAACCACTGTACAGCCT R-CAGGCATGGGAGTGAACCTA | Barley | [189] |
| Tomato yellow leaf curl virus | ACY | 6 | F-CCTTATGATGTCTCGTGAAAGG R-GAAGCACAGATTGAAGAAAACC | Tomato | [190] |
| Bacterial wilt | Bwr-6, Bwr-12 | 6,12 | F-TCAAGGTCCACTACCTTCATCC R-TCGGTATAGAGGGTACGTTG | Tomato | [191] |
| Fusarium wilt | frl | 9 | F-TACGATGACGTCGGT R-ATGCTACTGCGATGAAAC | Tomato | [192] |
| Fusarium wilt | Fom 1 | 7 | F-AACGAGAAGGCGGTGGAAAT R-CGATCTCCTCAAGGGAAGGTG | Melon | [193] |
| Leaf scald resistance | Rpf | Unmapped | F-TTGTTGGAACCTTTCGCTGG R-TAGACCTGTGCTGCCGTAAA | Sugarcane | [194] |
| Powdery mildew resistance | Pm-2 F | 1 | F-GCCCAACCTTCAACTCGATA R-TTGAATCTCATTTTTCTGTTGCAT | Melon | [195] |
| Melon necrotic spot virus | nsv | 4 | F-GTTTCTGATACGATGTTGTTTCCCTG R-GCCGAGATGCAGCAGGATGCTTTGCAC | Melon | [196] |
| Zucchini yellow mosaic virus (ZYMV) | eIF4E | 3 | F-TGGACITTYTGGTTYGAYAA R-GGRTCYTCCCAYTTIGGYTC | Watermelon | [197] |
| Powdery mildew | Pm | 5 | F-ATTTTCTTGCTTCAAATGGA R-ATAAGCAAAAGCATCGAAAG | Watermelon | [198] |
| Powdery mildew | Pm-s | 5 | F-CCCTATGCGTGAAAGCCACT R-CGCCTCAAACCCATACCCAA | Cucumber | [199] |
| Cauliflower mosaic virus (CMV) | cmv6.1 | 6 | F-ACAAAGCTTCTCCGCAAATG R-GGAGGGAAAGGAAGGAGAGA | Cucumber | [200] |
| Bacterial wilt-resistance | S401 | 6 | F--G ACTGCGTACCAATTCAGTT R-GATGAGTCCTGAGTAACACGATG | Eggplant | [201] |
| Powdery mildew resistance | InDel 1 | 1 | F-AACTTGGTAGCAATTTTATTGGGT R-TGGAGACAATGTGCATAAGTCTCT | Capsicum | [202] |
| Bacterial leaf spot resistance | Xcvr | 2 | F-TATCAAACGTAAAGTTGGAGCTTGT R-CCAAACACCTTGTGCATTGCT | Lettuce | [203] |
| Turnip mosaic virus resistance | retr02 | 4A | F-GGAGAAGACAAACAAACCCCC R-A TACCTTCGACACCGTCCAAGACTT | Turnip | [204] |
| Powdery mildew resistance | er1-7 | 6 | F-CGACACCGTATTCAAGCAGG R-TGTTGCCCTGTTTGATCGTT | Pea | [205] |
| Mungbean Yellow Mosaic Virus | YR4 | Unmapped | F-ACAAACATGGGCTGGAACAC R-GTGCCTGTAACTGCTCACAC | Mung bean | [206] |
| Resistance to weevils | VrPDF1 | Unmapped | F-CCAAGCTTGGTTAACAGTTTCTAGTGCACC R-GCGTCGACGATGGAGAAGAAATCACTGGCC | Mung bean | [207] |
| Abiotic stresses | |||||
| Dehydration tolerance | TaMYB2 | Unmapped | F-GAGGCCAGCTAGCAGCTGCC R-ATTGCCGGACGCGCAAGAGG | Wheat | [208] |
| Drought stress tolerance | TaAQP | Unmapped | F-ACATCAATTTTACCGTGCTTTG R-CAATCAATCTGCCGACTGTG | Wheat | [209] |
| Drought stress tolerance | DREB1 | 3D | F-GAATGGATCCCGGAAAGCAC R-GGGAATGAACCAAGCCACAG | Wheat | [210] |
| Salt tolerance | TtASR1 | Unmapped | F-ACCCCTACTTCTACATGCCG R-ATGATGGAGCTGTGGGACG | Wheat | [211] |
| Submergence tolerance | SubA1 | 9 | F-CTAGTTGGGCATACGATGGC R-ACGCTTATATGTTACGTCAAC | Rice | [212] |
| Tolerance to phosphorus (P) deficiency | Pup 1 | 12 | F-CTGGACTTGACCCCAATGTA R-TCTGATGGAGTGTTCGGAGT | Rice | [213] |
| Drought stress tolerance | OsSAPK2 | 5 | F-AAGGACATAGGGTCGGGGAA R-TGGCCAAATGTGTGGGAGTT | Rice | [214] |
| Drought tolerance | MYBE1 | 5 | F-GGTACCCTGTCAAGGTTCGG R-AATTACTGGCCCCAGGTTCG | Maize | [215] |
| Photoperiod response | Phd-H1 | 2H | F-GTTGAGATCGACAGTCCCCA R-GGGCTCCTATCTCCAACTCC | Barley | [135] |
| Aluminum stress tolerance | SbMATE | 4 | F-TAAGGCGCAATCATCATGGC R-CAACAAGATTCTGGAGCCGG | Sorghum | [216] |
| Drought and salt stress tolerance | CPRD12 | 11 | F-AAAGCATGCCCTAGTGGGAC R-ATGTCGGAAGCTACGGTTTCT | Cowpea | [217] |
| Dehydration response | SiDREB2 | Unmapped | F-CAACGGACTTGGGGCAAATG R-ATCGTTCGCTTCTGCCTTCA | Foxtail | [218] |
| Salinity tolerance | Salt index_QTL 1 | Unmapped | F-TGTACACTGTGTTTCTGTTGGT R-GTATTCGATCGTCCCTCCCG | Field pea | [219] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salgotra, R.K.; Stewart, C.N., Jr. Functional Markers for Precision Plant Breeding. Int. J. Mol. Sci. 2020, 21, 4792. https://doi.org/10.3390/ijms21134792
Salgotra RK, Stewart CN Jr. Functional Markers for Precision Plant Breeding. International Journal of Molecular Sciences. 2020; 21(13):4792. https://doi.org/10.3390/ijms21134792
Chicago/Turabian StyleSalgotra, Romesh K., and C. Neal Stewart, Jr. 2020. "Functional Markers for Precision Plant Breeding" International Journal of Molecular Sciences 21, no. 13: 4792. https://doi.org/10.3390/ijms21134792
APA StyleSalgotra, R. K., & Stewart, C. N., Jr. (2020). Functional Markers for Precision Plant Breeding. International Journal of Molecular Sciences, 21(13), 4792. https://doi.org/10.3390/ijms21134792