Lipid Peroxidation and Chlorophyll Fluorescence of Photosystem II Performance during Drought and Heat Stress is Associated with the Antioxidant Capacities of C3 Sunflower and C4 Maize Varieties



Abstract
:1. Introduction
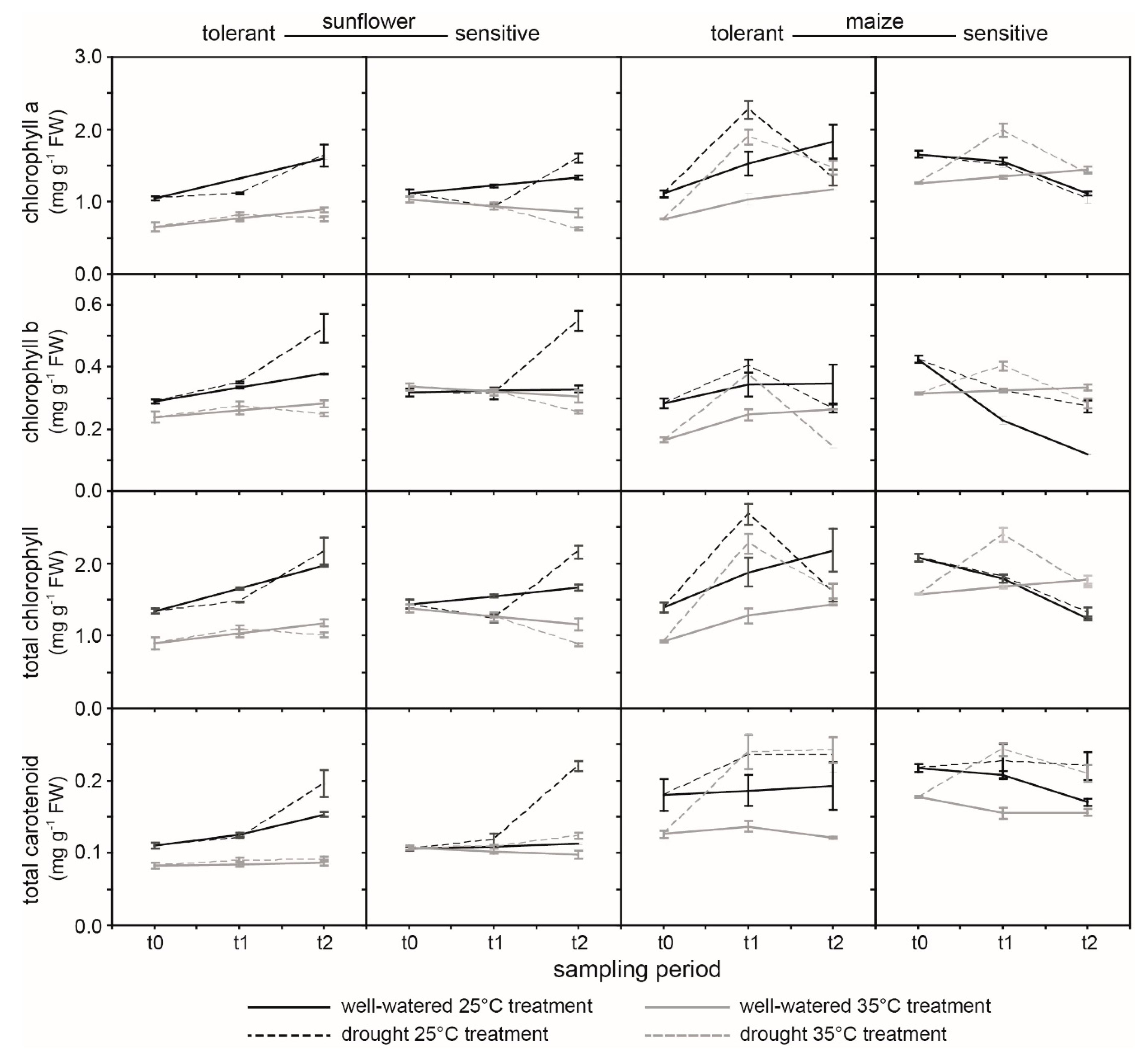
2. Results
2.1. Leaf Gas Exchange
2.2. Chlorophyll Fluorescence Parameters
2.3. Antioxidant Enzyme Activities
3. Discussion
3.1. Chlorophyll Fluorescence Analysis of the Effects of Drought and Heat Stress
3.2. Antioxidant Enzyme Responses to Drought and Heat Stress
4. Materials and Methods
4.1. Plant Growth Conditions and Drought Application
4.2. Chlorophyll a Fluorescence Transient Analysis and Parameters
4.3. Leaf Sampling and Assays of Antioxidant Enzyme Activities
4.4. Determination of Lipid Peroxidation
4.5. Determination of Pigment Contents
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FTSW | Fraction of transpirable soil water |
| SWC | Soil water content |
| PN | Net photosynthesis |
| Gs | Stomatal conductance |
| PSI | Photosystem I |
| PSII | Primary photochemistry/photosystem II (PSII) |
| Fv/Fm(φP0) | The maximum quantum yield of primary photochemistry (PSII), ((Fm-Fo)/Fm) |
| PAR | Photosynthetically active radiation |
| Chl | Chlorophyll |
| ChlF | Chlorophyll fluorescence |
| Fo | Minimal fluorescence |
| FM | Maximal fluorescence |
| FV | Maximal variable fluorescence |
| ψE0 | The efficiency of trapped energy to move an electron further than QA− |
| TBARS | Thiobarbituric acid reactive substances |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| APX | Ascorbate peroxidases |
| POX | Peroxidases |
| GR | Glutathione reductase |
| TS | Drought-tolerant sunflower variety (var. Bosfora) |
| SS | Drought-sensitive sunflower variety (var. 08 TR 003) |
| TM | Drought-tolerant maize variety (var. ADA-9516) |
| SM | Drought-sensitive maize variety (var. ADA-523). |
References
- Schär, C.; Vidale, P.L.; Lüthi, D.; Frei, C.; Häberli, C.; Liniger, M.A.; Appenzeller, C. The role of increasing temperature variability in European summer heatwaves. Nature 2004, 427, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Vautard, R.; Yiou, P.; D’Andrea, F.; De Noblet, N.; Viovy, N.; Cassou, C.; Polcher, J.; Ciais, P.; Kageyama, M.; Fan, Y. Summertime European heat and drought waves induced by wintertime Mediterranean rainfall deficit. Geophys. Res. Lett. 2007, 34, 7. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2012, 3, 30–36. [Google Scholar] [CrossRef]
- Semenov, M.; Stratonovitch, P.; Alghabari, F.; Gooding, M. Adapting wheat in Europe for climate change. J. Cereal Sci. 2014, 59, 245–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratonovitch, P.; Semenov, M. Heat tolerance around flowering in wheat identified as a key trait for increased yield potential in Europe under climate change. J. Exp. Bot. 2015, 66, 3599–3609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobell, D.B.; Hammer, G.; McLean, G.; Messina, C.; Roberts, M.J.; Schlenker, W. The critical role of extreme heat for maize production in the United States. Nat. Clim. Chang. 2013, 3, 497–501. [Google Scholar] [CrossRef]
- Marino, G.; Haworth, M.; Scartazza, A.; Tognetti, R.; Centritto, M. A comparison of the Variable J and carbon-isotopic composition of sugars methods to assess mesophyll conductance from the leaf to the canopy scale in drought-stressed cherry. Int. J. Mol. Sci. 2020, 21, 1222. [Google Scholar] [CrossRef] [Green Version]
- Haworth, M.; Marino, G.; Riggi, E.; Avola, G.; Brunetti, C.; Scordia, D.; Testa, G.; Gomes, M.T.G.; Loreto, F.; Cosentino, S.L.; et al. The effect of summer drought on the yield of Arundo donax is reduced by the retention of photosynthetic capacity and leaf growth later in the growing season. Ann. Bot. 2019, 124, 567–579. [Google Scholar] [CrossRef]
- Crafts-Brandner, S.J.; Salvucci, M.E. Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2. Proc. Natl. Acad. Sci. USA 2000, 97, 13430–13435. [Google Scholar] [CrossRef] [Green Version]
- Killi, D.; Bussotti, F.; Raschi, A.; Haworth, M. Adaptation to high temperature mitigates the impact of water deficit during combined heat and drought stress in C3 sunflower and C4 maize varieties with contrasting drought tolerance. Physiol. Plant. 2016, 159, 130–147. [Google Scholar] [CrossRef]
- Nguyen, H.; Babu, R.C.; Blum, A. Breeding for drought resistance in rice: Physiology and molecular genetics considerations. Crop. Sci. 1997, 37, 1426–1434. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2006, 58, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [Green Version]
- Kalaji, H.M.; Kalaji, H.M.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Haworth, M.; Marino, G.; Brunetti, C.; Killi, D.; De Carlo, A.; Centritto, M. The impact of heat stress and water deficit on the photosynthetic and stomatal physiology of olive (Olea europaea L.)—A case study of the 2017 heat wave. Plants 2018, 7, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crafts-Brandner, S.J.; Salvucci, M.E. Sensitivity of photosynthesis in a C4 plant, maize, to heat stress. Plant Physiol. 2002, 129, 1773–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haworth, M.; Belcher, C.M.; Killi, D.; Dewhirst, R.A.; Materassi, A.; Raschi, A.; Centritto, M. Impaired photosynthesis and increased leaf construction costs may induce floral stress during episodes of global warming over macroevolutionary timescales. Sci. Rep-UK 2018, 8, 6206. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2010, 62, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop. Sci. 1981, 21, 43. [Google Scholar] [CrossRef]
- Lauteri, M.; Haworth, M.; Serraj, R.; Monteverdi, M.C.; Centritto, M. Photosynthetic diffusional constraints affect yield in drought stressed rice cultivars during flowering. PLoS ONE 2014, 9, e109054. [Google Scholar] [CrossRef] [PubMed]
- Centritto, M.; Loreto, F.; Chartzoulakis, K. The use of low [CO2] to estimate diffusional and non-diffusional limitations of photosynthetic capacity of salt-stressed olive saplings. Plant Cell Environ. 2003, 26, 585–594. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Energy dissipation in C3 plants under drought. Funct. Plant Boil. 2002, 29, 1209. [Google Scholar] [CrossRef]
- Smirnoff, N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef]
- Feller, U.; Berenbaum, M. Moderately high temperatures inhibit Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase (Rubisco) Activase-mediated activation of Rubisco. Plant Physiol. 1998, 116, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Yordanov, I.; Velikova, V.; Tsonev, T. Influence of drought, high temperature, and carbamide cytokinin 4-PU-30 on photosynthetic activity of bean plants. 1. Changes in chlorophyll fluorescence quenching. Photosynthetica 1999, 37, 447–457. [Google Scholar] [CrossRef]
- Havaux, M. Characterization of thermal damage to the photosynthetic electron transport system in potato leaves. Plant Sci. 1993, 94, 19–33. [Google Scholar] [CrossRef]
- Gounaris, K.; Brain, A.P.; Quinn, P.J.; Williams, W. Structural and functional changes associated with heat-induced phase-separations of non-bilayer lipids in chloroplast thylakoid membranes. FEBS Lett. 1983, 153, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Berry, J.; Bjorkman, O. Photosynthetic response and adaptation to temperature in higher plants. Annu. Rev. Plant Physiol. 1980, 31, 491–543. [Google Scholar] [CrossRef]
- Sinsawat, V.; Leipner, J.; Stamp, P.; Fracheboud, Y. Effect of heat stress on the photosynthetic apparatus in maize (Zea mays L.) grown at control or high temperature. Environ. Exp. Bot. 2004, 52, 123–129. [Google Scholar] [CrossRef]
- Haworth, M.; Cosentino, S.L.; Marino, G.; Brunetti, C.; Scordia, D.; Testa, G.; Riggi, E.; Avola, G.; Loreto, F.; Centritto, M. Physiological responses of Arundo donax ecotypes to drought: A common garden study. GCB Bioenergy 2016, 9, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Oukarroum, A.; El Madidi, S.; Schansker, G.; Strasser, R.J. Probing the responses of barley cultivars (Hordeum vulgare L.) by chlorophyll a fluorescence OLKJIP under drought stress and re-watering. Environ. Exp. Bot. 2007, 60, 438–446. [Google Scholar] [CrossRef]
- Costa, J.M.; Da Silva, J.M.; Pinheiro, C.; Barón, M.; Mylona, P.; Centritto, M.; Haworth, M.; Loreto, F.; Uzilday, B.; Turkan, I.; et al. Opportunities and limitations of crop phenotyping in southern European countries. Front. Plant Sci. 2019, 10, 1125. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Huang, B. Drought and heat stress injury to two cool-season turfgrasses in relation to antioxidant metabolism and lipid peroxidation. Crop Sci. 2001, 41, 436–442. [Google Scholar] [CrossRef]
- Turkan, I.; Turkan, I.; Ozdemir, F.; Koca, H. Differential responses of lipid peroxidation and antioxidants in the leaves of drought-tolerant P. acutifolius Gray and drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress. Plant Sci. 2005, 168, 223–231. [Google Scholar] [CrossRef]
- Xu, S.; Li, J.; Zhang, X.; Wei, H.; Cui, L. Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation, antioxidant metabolites, and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress. Environ. Exp. Bot. 2006, 56, 274–285. [Google Scholar] [CrossRef]
- Kumar, S.; Gupta, D.; Nayyar, H. Comparative response of maize and rice genotypes to heat stress: Status of oxidative stress and antioxidants. Acta Physiol. Plant. 2011, 34, 75–86. [Google Scholar] [CrossRef]
- Liu, X.; Huang, B. Heat stress injury in relation to membrane lipid peroxidation in creeping bentgrass. Crop Sci. 2000, 40, 503–510. [Google Scholar] [CrossRef]
- Sairam, R.; Srivastava, G.; Saxena, D. Increased antioxidant activity under elevated temperatures: A mechanism of heat stress tolerance in wheat genotypes. Boil. Plant. 2000, 43, 245–251. [Google Scholar] [CrossRef]
- Sekmen, A.H.; Ozgur, R.; Uzilday, B.; Turkan, I. Reactive oxygen species scavenging capacities of cotton (Gossypium hirsutum) cultivars under combined drought and heat induced oxidative stress. Environ. Exp. Bot. 2014, 99, 141–149. [Google Scholar] [CrossRef]
- Panković, D.; Sakač, Z.; Kevrešan, S.; Plesničar, M. Acclimation to long-term water deficit in the leaves of two sunflower hybrids: Photosynthesis, electron transport and carbon metabolism. J. Exp. Bot. 1999, 50, 128–138. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.R. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukhov, N.G.; Carpentier, R. Effects of Water Stress on the Photosynthetic Efficiency of Plants. In Plant Cell Monographs; Springer Science and Business Media LLC: Dordrecht, Dutch, 2007; Volume 19, pp. 623–635. [Google Scholar]
- Haworth, M.; Centritto, M.; Giovannelli, A.; Marino, G.; Proietti, N.; Capitani, D.; De Carlo, A.; Loreto, F. Xylem morphology determines the drought response of two Arundo donax ecotypes from contrasting habitats. GCB Bioenergy 2016, 9, 119–131. [Google Scholar] [CrossRef]
- Haworth, M.; Catola, S.; Marino, G.; Brunetti, C.; Michelozzi, M.; Riggi, E.; Avola, G.; Cosentino, S.L.; Loreto, F.; Centritto, M. Moderate drought stress induces increased foliar dimethylsulphoniopropionate (DMSP) concentration and isoprene emission in two contrasting ecotypes of Arundo donax. Front. Plant Sci. 2017, 8, 1016. [Google Scholar] [CrossRef] [PubMed]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Martinazzo, E.G.; Ramm, A.; Bacarin, M.A. The chlorophyll a fluorescence as an indicator of the temperature stress in the leaves of Prunus persica. Braz. J. Plant Physiol. 2013, 24, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Haworth, M.; Gallagher, A.; Sum, E.; Hill-Donnelly, M.; Steinthorsdottir, M.; McElwain, J.C. On the reconstruction of plant photosynthetic and stress physiology across the Triassic–Jurassic boundary. Turk. J. Earth Sci. 2014, 23, 321–329. [Google Scholar] [CrossRef]
- Schansker, G.; Toth, S.Z.; Strasser, R.J. Methylviologen and dibromothymoquinone treatments of pea leaves reveal the role of photosystem I in the Chl a fluorescence rise OJIP. Biochim. et Biophys. Acta (BBA)-Gen. Subj. 2005, 1706, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Ceppi, M.G.; Oukarroum, A.; Çiçek, N.; Strasser, R.J.; Schansker, G. The IP amplitude of the fluorescence rise OJIP is sensitive to changes in the photosystem I content of leaves: A study on plants exposed to magnesium and sulfate deficiencies, drought stress and salt stress. Physiol. Plant. 2012, 144, 277–288. [Google Scholar] [CrossRef]
- Pospíšil, P. Production of reactive oxygen species by photosystem II. Biochim. et Biophys. Acta (BBA)-Bioenerg. 2009, 1787, 1151–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pospíšil, P. Production of reactive oxygen species by Photosystem II as a response to light and temperature stress. Front. Plant Sci. 2016, 7, 1950. [Google Scholar] [CrossRef] [PubMed]
- Jung, S. Variation in antioxidant metabolism of young and mature leaves of Arabidopsis thaliana subjected to drought. Plant Sci. 2004, 166, 459–466. [Google Scholar] [CrossRef]
- Huang, C.; Zhao, S.; Wang, L.; Anjum, S.A.; Chen, M.; Zhou, H.; Zou, C. Alteration in chlorophyll fluorescence, lipid peroxidation and antioxidant enzymes activities in hybrid ramie (Boehmeria nivea L.) under drought stress. Aust. J. Crop Sci. 2013, 7, 594–599. [Google Scholar]
- Srivastava, A.; Guissé, B.; Greppin, H.; Strasser, R.J. Regulation of antenna structure and electron transport in Photosystem II of Pisum sativum under elevated temperature probed by the fast polyphasic chlorophyll a fluorescence transient: OKJIP. Biochim. et Biophys. Acta (BBA)-Bioenerg. 1997, 1320, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence; Springer Science and Business Media LLC: Dordrecht, Dutch, 2004; Volume 19, pp. 321–362. [Google Scholar]
- Müller, P.; Li, X.-P.; Niyogi, K.K. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [Green Version]
- Agati, G.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Pollastri, S.; Tattini, M. Functional roles of flavonoids in photoprotection: New evidence, lessons from the past. Plant Physiol. Biochem. 2013, 72, 35–45. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Rampino, P.; Mita, G.; Fasano, P.; Borrelli, G.M.; Aprile, A.; Dalessandro, G.; De Bellis, L.; Perrotta, C. Novel durum wheat genes up-regulated in response to a combination of heat and drought stress. Plant Physiol. Biochem. 2012, 56, 72–78. [Google Scholar] [CrossRef]
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef]
- Haworth, M.; Elliott-Kingston, C.; McElwain, J.C. Stomatal control as a driver of plant evolution. J. Exp. Bot. 2011, 62, 2419–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demmig-Adams, B.; Adams, W.W. Photosynthesis: Harvesting sunlight safely. Nature 2000, 303, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Peltzer, D.; Dreyer, E.; Polle, A. Differential temperature dependencies of antioxidative enzymes in two contrasting species: Fagus sylvatica and Coleus blumei. Plant Physiol. Biochem. 2002, 40, 141–150. [Google Scholar] [CrossRef]
- Elstner, E.F. Oxygen activation and oxygen toxicity. Annu. Rev. Plant Physiol. 1982, 33, 73–96. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C.; Tadolini, B.; Puntarulo, S.; I Cederbaum, A.; I Aruoma, O.; Laughton, M.J.; Quinlan, G.J.; O’Connell, M.J.; Ward, R.J.; et al. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Gambarova, N.G.; Gins, M.S. Characteristics of oxidative stress of plants with C3 and C4 photosynthesis during salinization. Russ. Agric. Sci. 2008, 34, 77–80. [Google Scholar] [CrossRef]
- Materassi, A.; Fasano, G.; Arca, A. Climatic chamber for plant physiology: A new project concept. Rivista di Ingegneria Agraria (Italy) 2005, 4, 79–87. [Google Scholar]
- Sinclair, T.; Ludlow, M. Influence of soil water supply on the plant water balance of four tropical grain legumes. Funct. Plant Boil. 1986, 13, 329–341. [Google Scholar] [CrossRef]
- Killi, D.; Anlauf, R.; Kavdır, Y.; Haworth, M.; Kavdir, Y. Assessing the impact of agro-industrial olive wastes in soil water retention: Implications for remediation of degraded soils and water availability for plant growth. Int. Biodeterior. Biodegradation 2014, 94, 48–56. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, Á.; Dabrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Bergmeyer, H.U.; Gawehn, K. Methoden der Enzymatischen Analyse; Akademie Verlag: Berlin, Germany, 1970; Volume 1, pp. 636–647. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Kanner, J.; Kinsella, J.E. Lipid deterioration initiated by phagocytic cells in muscle foods: .beta.-carotene destruction of a myeloperoxidase-hydrogen peroxide-halide system. J. Agric. Food Chem. 1983, 31, 370–376. [Google Scholar] [CrossRef]
- Rao, K.M.; Sresty, T. Antioxidative parameters in the seedlings of pigeonpea (Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Fo | Fm | Fv/Fm | ΨEo | RC/ABS | PIabs | PItot | I-Pphase | KBand | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp. | Var. | Water | |||||||||
| 25 °C | TM | WW | 263.4 ± 13.1 | 1366.1 ± 59.7○ | 0.775 ± 0.01 | 0.610 ± 0.03 | 4.404 ± 0.38 | 24.437 ± 5.71 | 16.001 ± 7.16 | 0.234 ± 0.06 | 0.443 ± 0.03 |
| TM | D | 287.6 ± 24.7 | 1323.0 ± 75.6▲ | 0.746 ± 0.02○ | 0.600 ± 0.02 | 4.025 ± 0.42 | 18.951 ± 7.30 | 13.086 ± 4.04 | 0.248 ± 0.03○ | 0.467 ± 0.03 | |
| SM | WW | 280.1 ± 10.1 | 1455.4 ± 63.8○ | 0.773 ± 0.01 | 0.602 ± 0.01 | 4.221 ± 0.20 | 21.844 ± 2.37 | 11.887 ± 1.37 | 0.210 ± 0.00 | 0.458 ± 0.01 | |
| SM | D | 302.8 ± 13.7▲○ | 1322.9 ± 59.0○ | 0.728 ± 0.02▲○ | 0.561 ± 0.04▲○ | 3.569 ± 0.30▲○ | 12.909 ± 4.88▲○ | 9.171 ± 2.49○ | 0.234 ± 0.02 | 0.512 ± 0.02▲○ | |
| 35 °C | TM | WW | 258.3 ± 13.5○ | 1219.3 ± 59.7▲○ | 0.755 ± 0.00▲○ | 0.577 ± 0.02 | 4.267 ± 0.06 | 19.581 ± 2.28 | 14.274 ± 3.56 | 0.229 ± 0.02 | 0.445 ± 0.00 |
| TM | D | 276.0 ± 15.0 | 1261.2 ± 54.9▲○ | 0.749 ± 0.02▲○ | 0.640 ± 0.01○ | 3.943 ± 0.31▲ | 21.678 ± 5.22 | 13.402 ± 3.54 | 0.240 ± 0.02 | 0.477 ± 0.02 | |
| SM | WW | 276.4 ± 7.3 | 1224.3 ± 42.3▲○ | 0.735 ± 0.00▲○ | 0.537 ± 0.02 | 3.961 ± 0.13 | 14.958 ± 2.02▲ | 12.216 ± 4.40 | 0.233 ± 0.04 | 0.469 ± 0.01 | |
| SM | D | 292.9 ± 25.7▲ | 1250.3 ± 70.8▲○ | 0.726 ± 0.01▲○ | 0.577 ± 0.03 | 3.646 ± 0.24▲○ | 14.338 ± 1.21▲○ | 10.465 ± 2.58 | 0.244 ± 0.05 | 0.501 ± 0.02▲○ | |
| 25 °C | TS | WW | 226.5 ± 9.0 | 1650.4 ± 40.3● | 0.847 ± 0.00 | 0.749 ± 0.01● | 6.477 ± 0.25 | 108.032 ± 9.44● | 98.229 ± 12.29 | 0.355 ± 0.02 | 0.328 ± 0.01 |
| TS | D | 236.4 ± 6.9 | 1716.0 ± 58.2□ | 0.845 ± 0.00 | 0.724 ± 0.02 | 6.185 ± 0.32 | 93.674 ± 13.73 | 74.735 ± 18.56 | 0.308 ± 0.03 | 0.343 ± 0.01 | |
| SS | WW | 232.5 ± 13.5 | 1707.3 ± 61.7□ | 0.846 ± 0.00 | 0.718 ± 0.01□ | 6.335 ± 0.25 | 89.967 ± 14.86□ | 82.747 ± 9.75 | 0.345 ± 0.01 | 0.334 ± 0.01 | |
| SS | D | 231.6 ± 10.9 | 1649.6 ± 61.3● | 0.842 ± 0.00 | 0.723 ± 0.03 | 6.117 ± 0.29 | 89.016 ± 22.01 | 73.243 ± 30.65 | 0.316 ± 0.05 | 0.345 ± 0.01 | |
| 35 °C | TS | WW | 236.6 ± 18.4 | 1534.6 ± 100.3□● | 0.828 ± 0.00□● | 0.717 ± 0.01□ | 6.151 ± 0.46 | 75.968 ± 12.30□ | 73.436 ± 16.29□ | 0.350 ± 0.02 | 0.338 ± 0.02 |
| TS | D | 261.4 ± 17.9□● | 1537.1 ± 102.5□● | 0.811 ± 0.00□● | 0.708 ± 0.01□ | 5.669 ± 0.47□● | 59.649 ± 11.68□● | 65.109 ± 19.35□● | 0.366 ± 0.02 | 0.360 ± 0.02□ | |
| SS | WW | 292.2 ± 38.9□● | 1529.2 ± 61.2□● | 0.775 ± 0.02□● | 0.593 ± 0.03□● | 4.548 ± 0.58□● | 26.128 ± 10.52□● | 33.084 ± 8.93□● | 0.340 ± 0.02 | 0.432 ± 0.04□● | |
| SS | D | 296.6 ± 35.0□● | 1511.5 ±80.9□● | 0.773 ± 0.01□● | 0.555 ± 0.02□● | 4.791 ±0.41□● | 20.994 ±3.58□● | 28.027 ±4.22□● | 0.317 ± 0.02□● | 0.405 ± 0.02□● | |
| Species | Factor | CAT | APX | POX | GR | SOD | TBARS | Chl a | Chl b | Tot. Chl | Tot. Carot. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Maize | Variety (V) | n.s. | n.s. | n.s. | ** | n.s. | *** | ** | n.s. | * | n.s. |
| Temperature (T) | *** | *** | *** | n.s. | ** | *** | n.s. | n.s. | n.s. | n.s. | |
| Water (W) | n.s. | *** | n.s. | *** | ** | *** | n.s. | n.s. | n.s. | *** | |
| V * T | *** | * | n.s. | n.s. | n.s. | n.s. | *** | *** | *** | n.s. | |
| V * W | *** | n.s. | ** | n.s. | n.s. | *** | n.s. | *** | n.s. | n.s. | |
| T * W | *** | *** | * | n.s. | *** | * | ** | *** | n.s. | n.s. | |
| V * T * W | *** | ** | *** | n.s. | n.s. | n.s. | ** | * | ** | n.s. | |
| Sunflower | Variety (V) | *** | n.s. | *** | *** | *** | *** | *** | *** | ** | *** |
| Temperature (T) | n.s. | *** | ** | n.s. | *** | *** | *** | *** | *** | *** | |
| Water (W) | n.s. | *** | ** | *** | * | *** | n.s. | * | n.s. | *** | |
| V * T | *** | *** | ** | n.s. | *** | *** | *** | *** | *** | ** | |
| V * W | *** | ** | *** | * | *** | *** | n.s. | *** | n.s. | * | |
| T * W | *** | *** | *** | n.s. | ** | *** | n.s. | *** | n.s. | n.s. | |
| V * T * W | n.s. | *** | *** | * | *** | n.s. | *** | ** | *** | ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Killi, D.; Raschi, A.; Bussotti, F. Lipid Peroxidation and Chlorophyll Fluorescence of Photosystem II Performance during Drought and Heat Stress is Associated with the Antioxidant Capacities of C3 Sunflower and C4 Maize Varieties. Int. J. Mol. Sci. 2020, 21, 4846. https://doi.org/10.3390/ijms21144846
Killi D, Raschi A, Bussotti F. Lipid Peroxidation and Chlorophyll Fluorescence of Photosystem II Performance during Drought and Heat Stress is Associated with the Antioxidant Capacities of C3 Sunflower and C4 Maize Varieties. International Journal of Molecular Sciences. 2020; 21(14):4846. https://doi.org/10.3390/ijms21144846
Chicago/Turabian StyleKilli, Dilek, Antonio Raschi, and Filippo Bussotti. 2020. "Lipid Peroxidation and Chlorophyll Fluorescence of Photosystem II Performance during Drought and Heat Stress is Associated with the Antioxidant Capacities of C3 Sunflower and C4 Maize Varieties" International Journal of Molecular Sciences 21, no. 14: 4846. https://doi.org/10.3390/ijms21144846
APA StyleKilli, D., Raschi, A., & Bussotti, F. (2020). Lipid Peroxidation and Chlorophyll Fluorescence of Photosystem II Performance during Drought and Heat Stress is Associated with the Antioxidant Capacities of C3 Sunflower and C4 Maize Varieties. International Journal of Molecular Sciences, 21(14), 4846. https://doi.org/10.3390/ijms21144846