Shade Effects on Peanut Yield Associate with Physiological and Expressional Regulation on Photosynthesis and Sucrose Metabolism

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
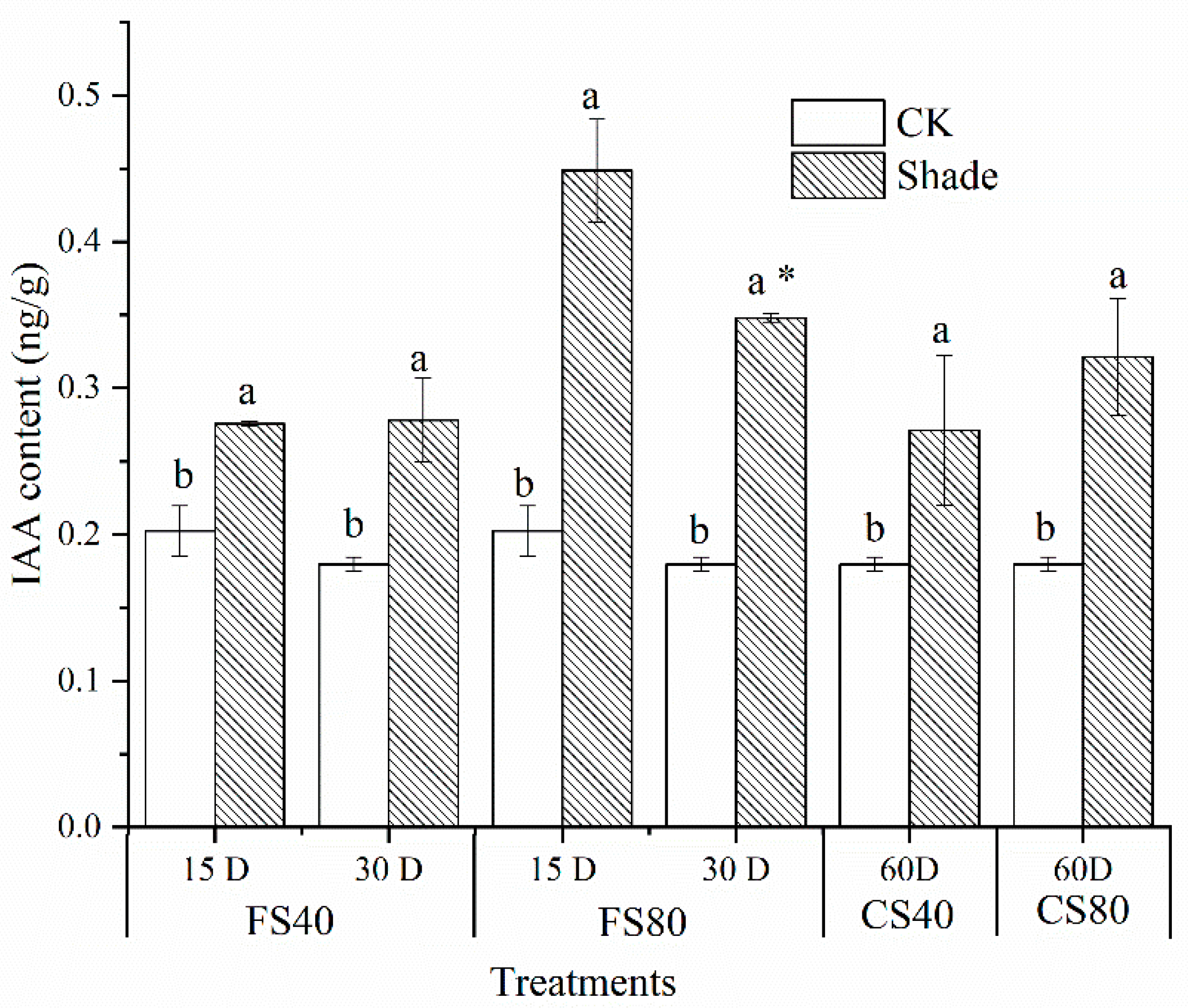
2.1. Effects of Shade Stress on Size of Peanut Plants and Association with Auxin
2.2. Effects of Shade Stress on Yield Components
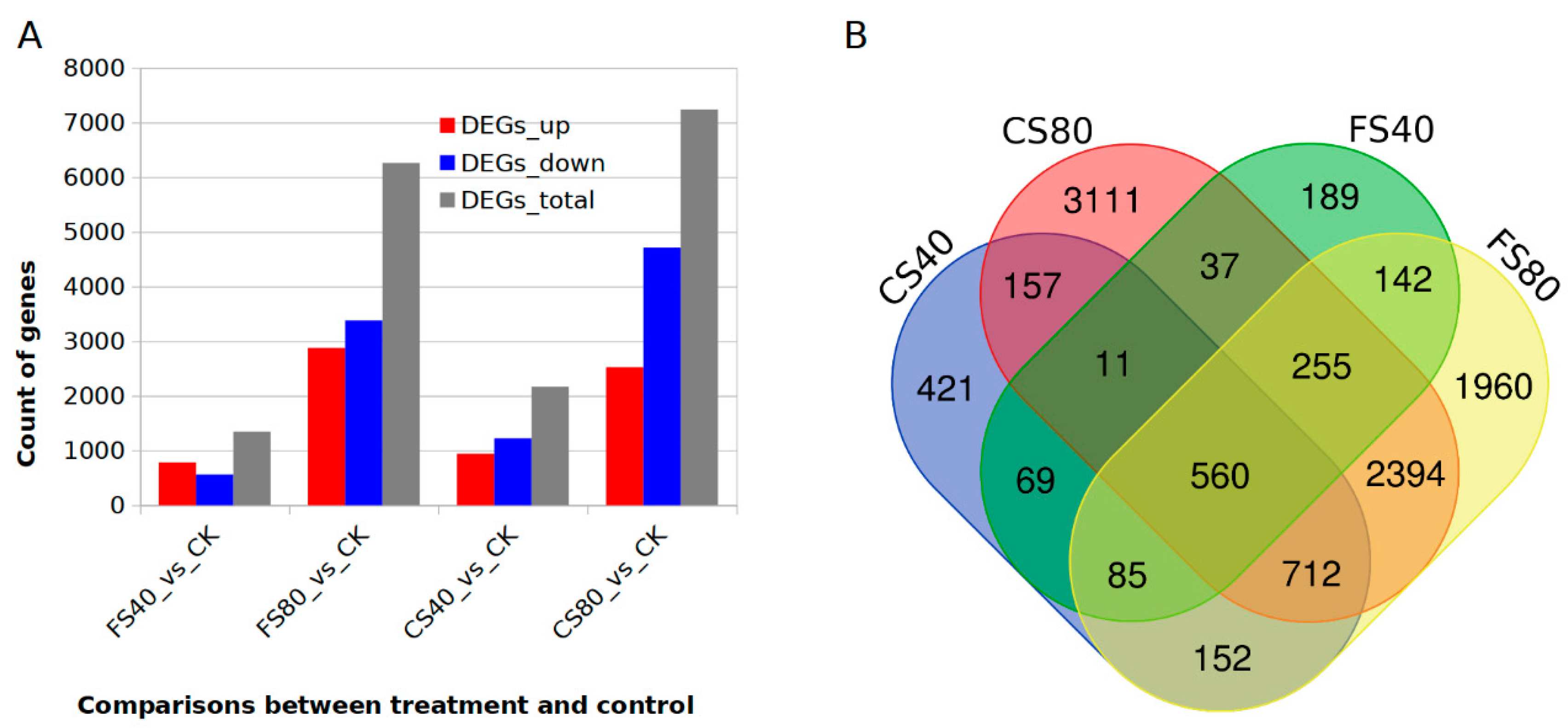
2.3. Transcriptome Profiles and Expressional Regulation Responsive to Shade
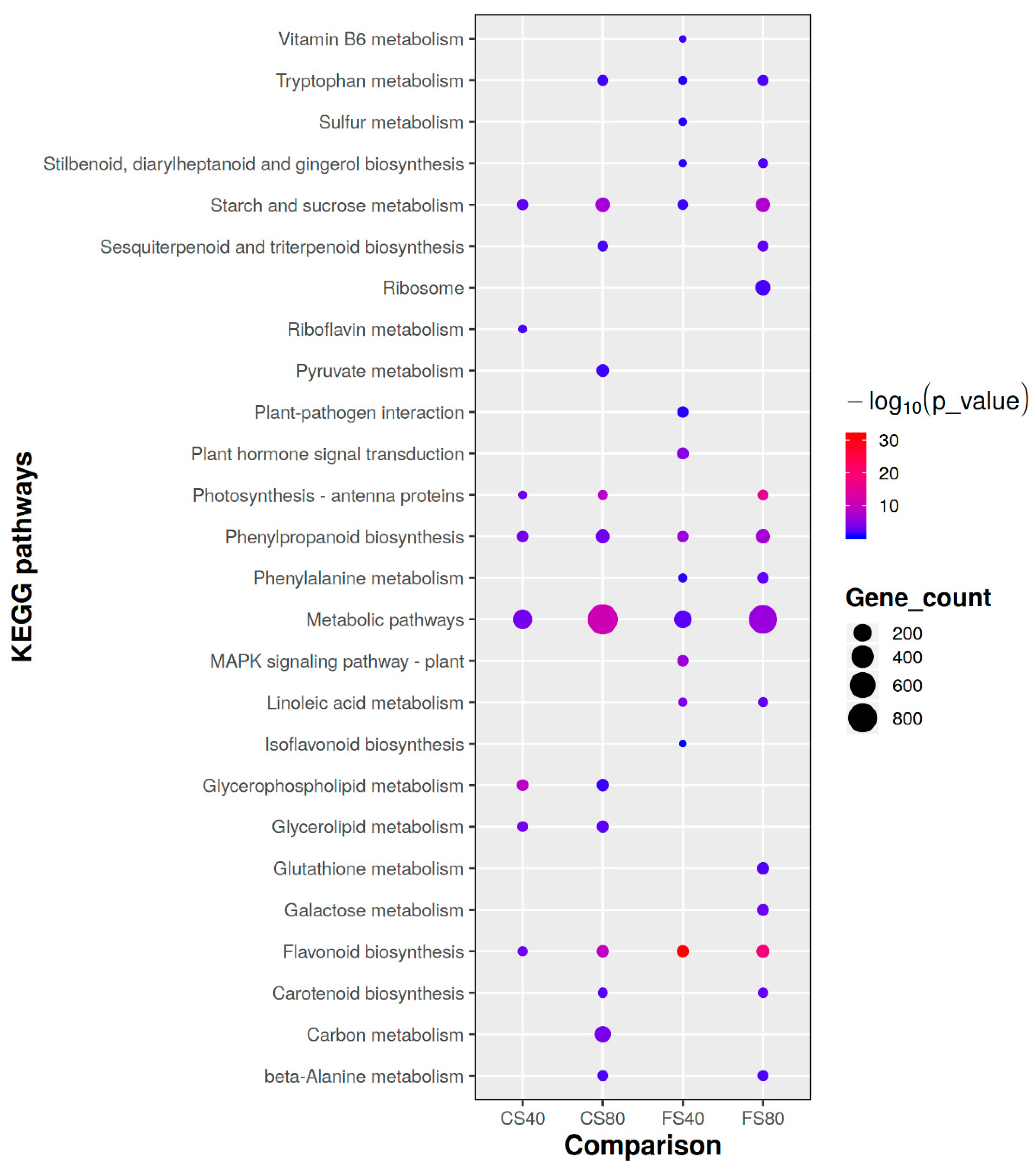
2.4. Function of Induced DEGs and Affected Pathways by Shade Stress
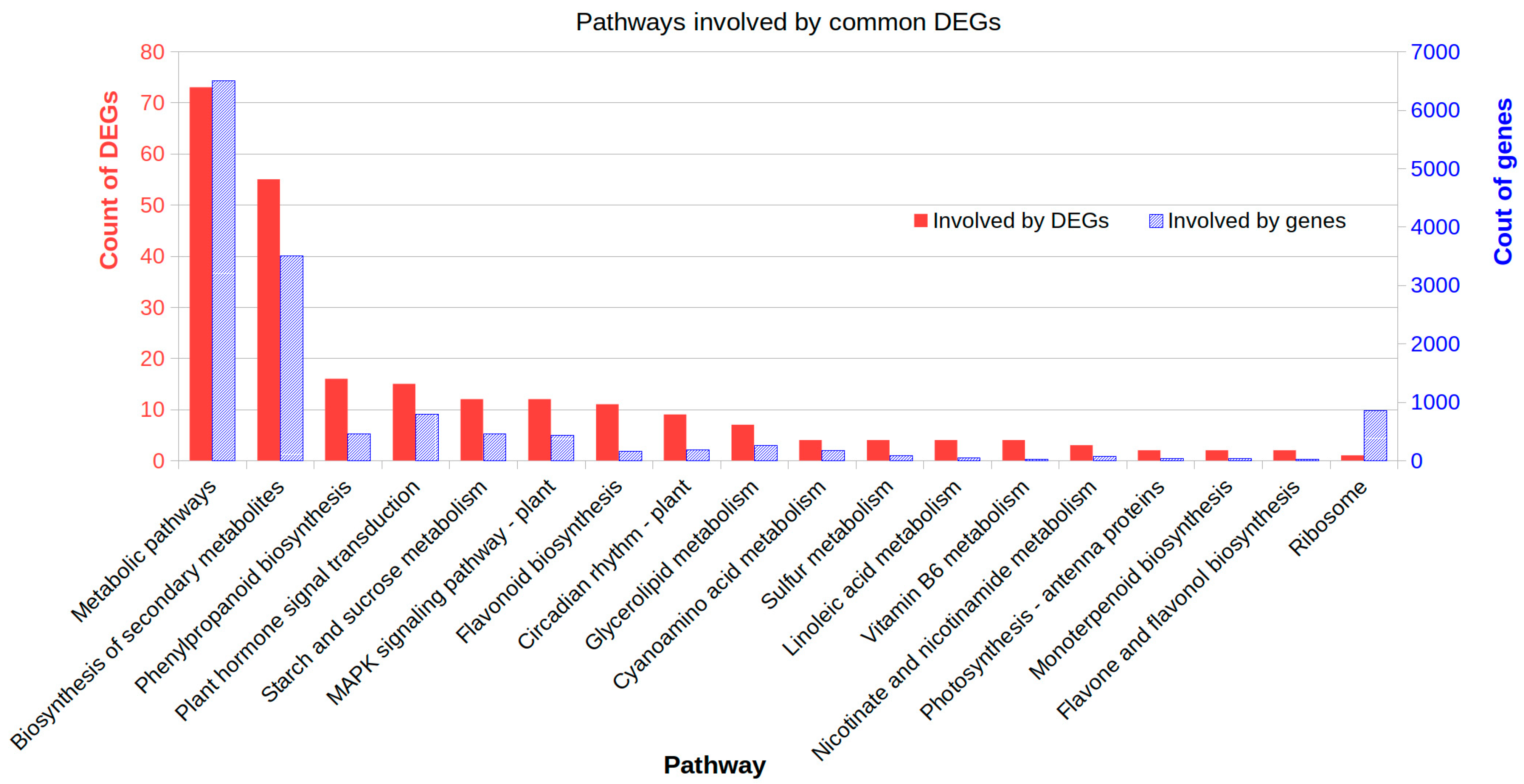
2.5. Affected Metabolism Pathways Involved by Common DEGs
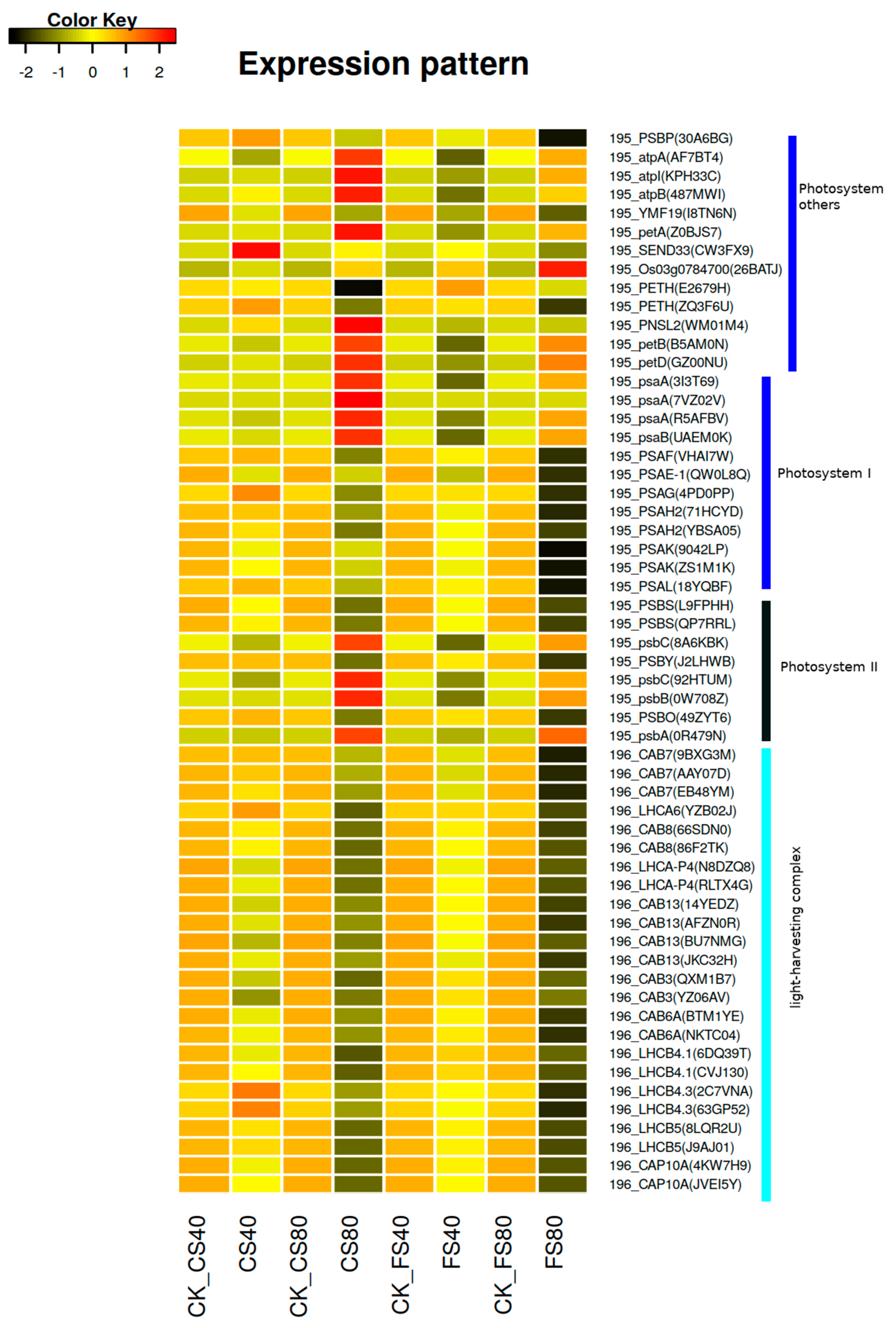
2.6. Regulation of DEGs and Association with Physiological Photosynthesis
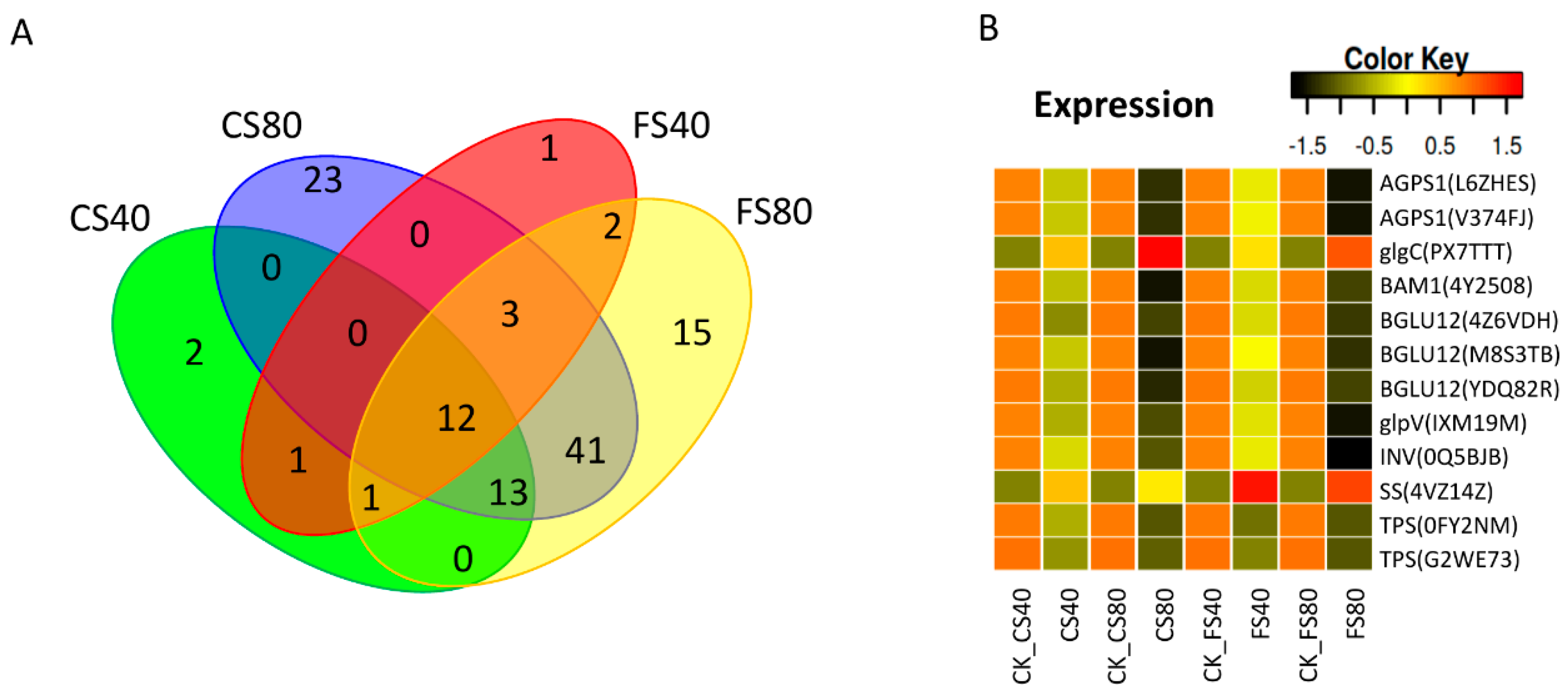
2.7. Regulation of DEGs in Starch and Sucrose Metabolism and Association with Biomass
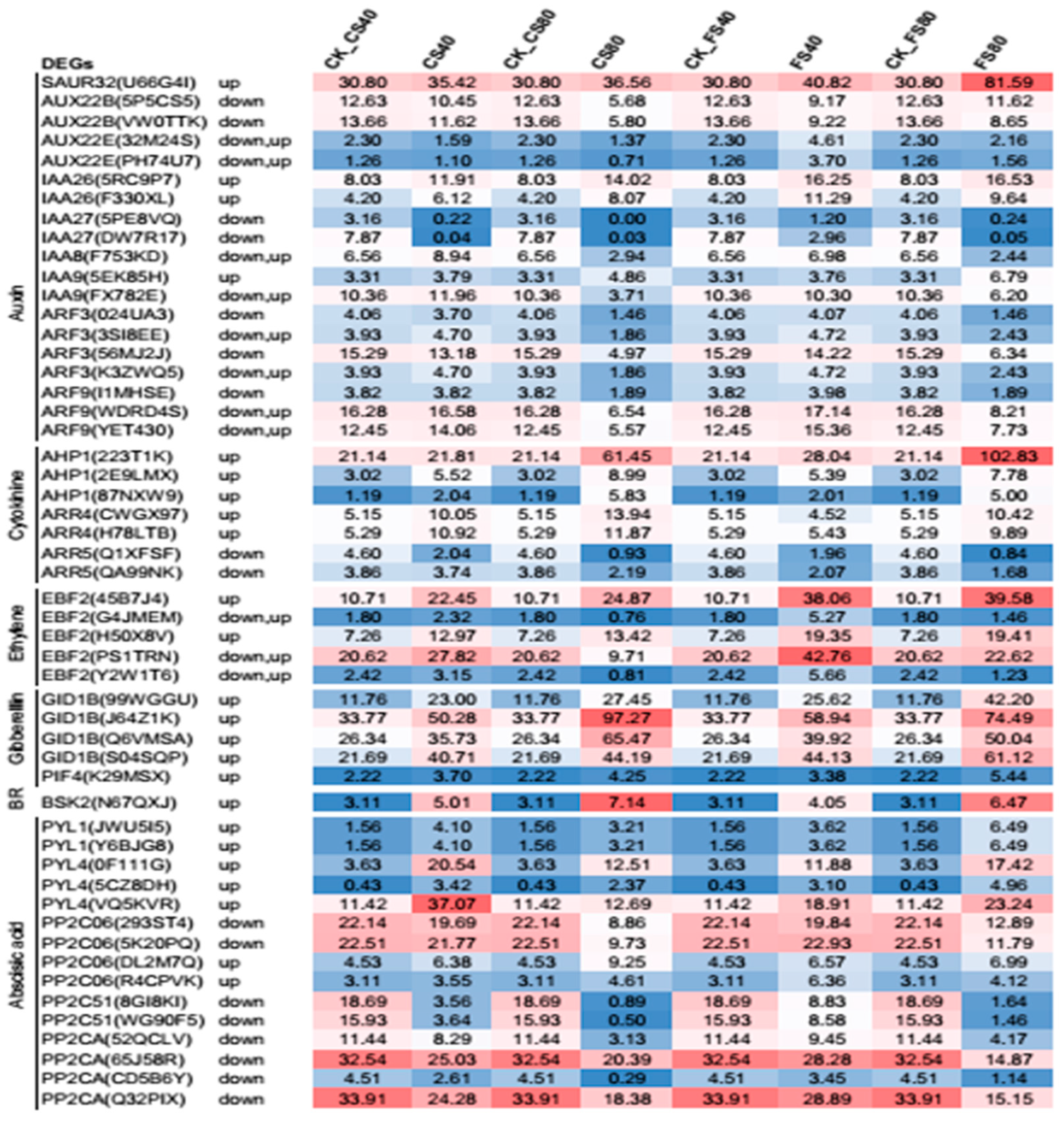
2.8. Regulation of DEGs in Hormone Signaling
3. Discussion
4. Materials and Methods
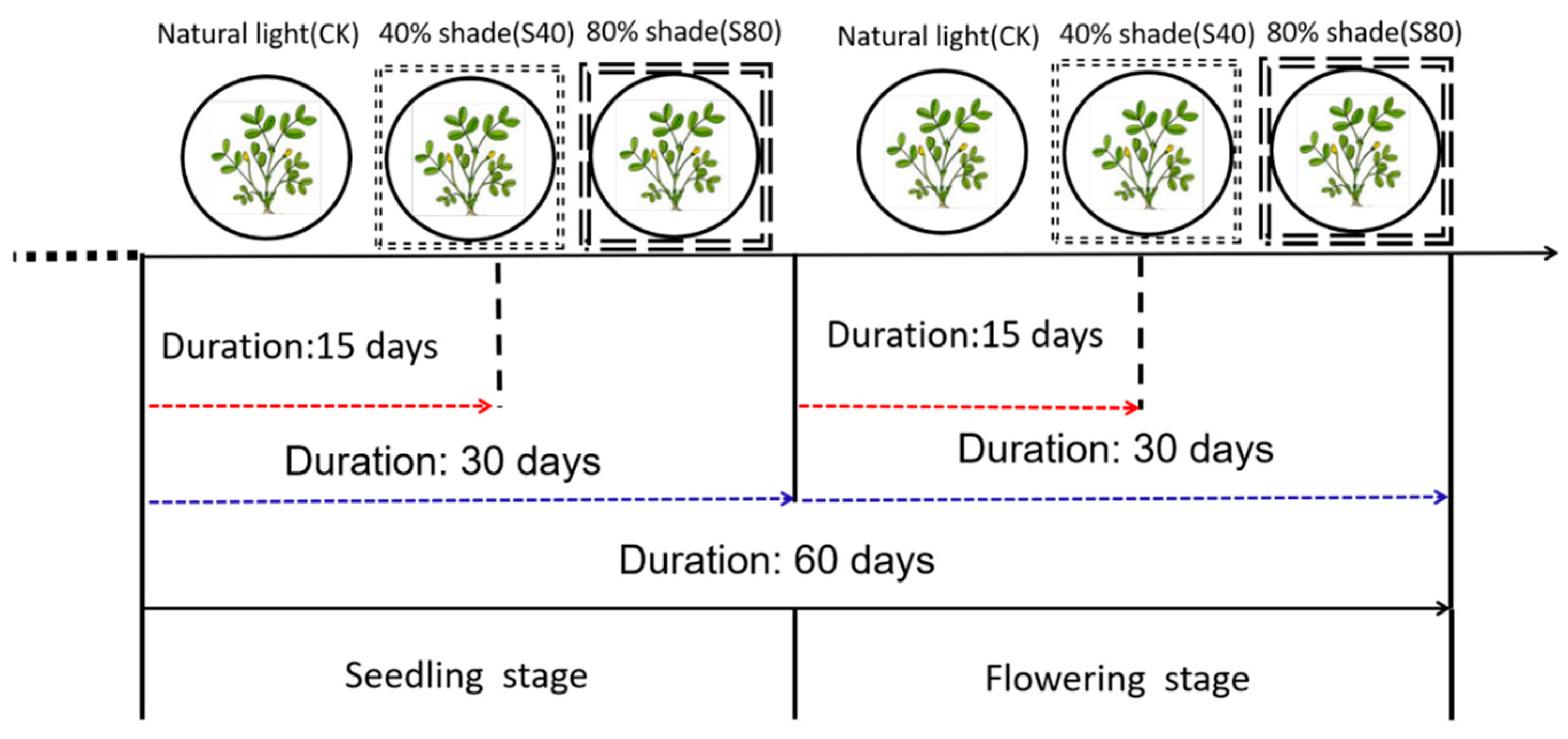
4.1. Plant and Growth Condition
4.2. Shade Stress Treatments
4.3. Length and Biomass Measurements
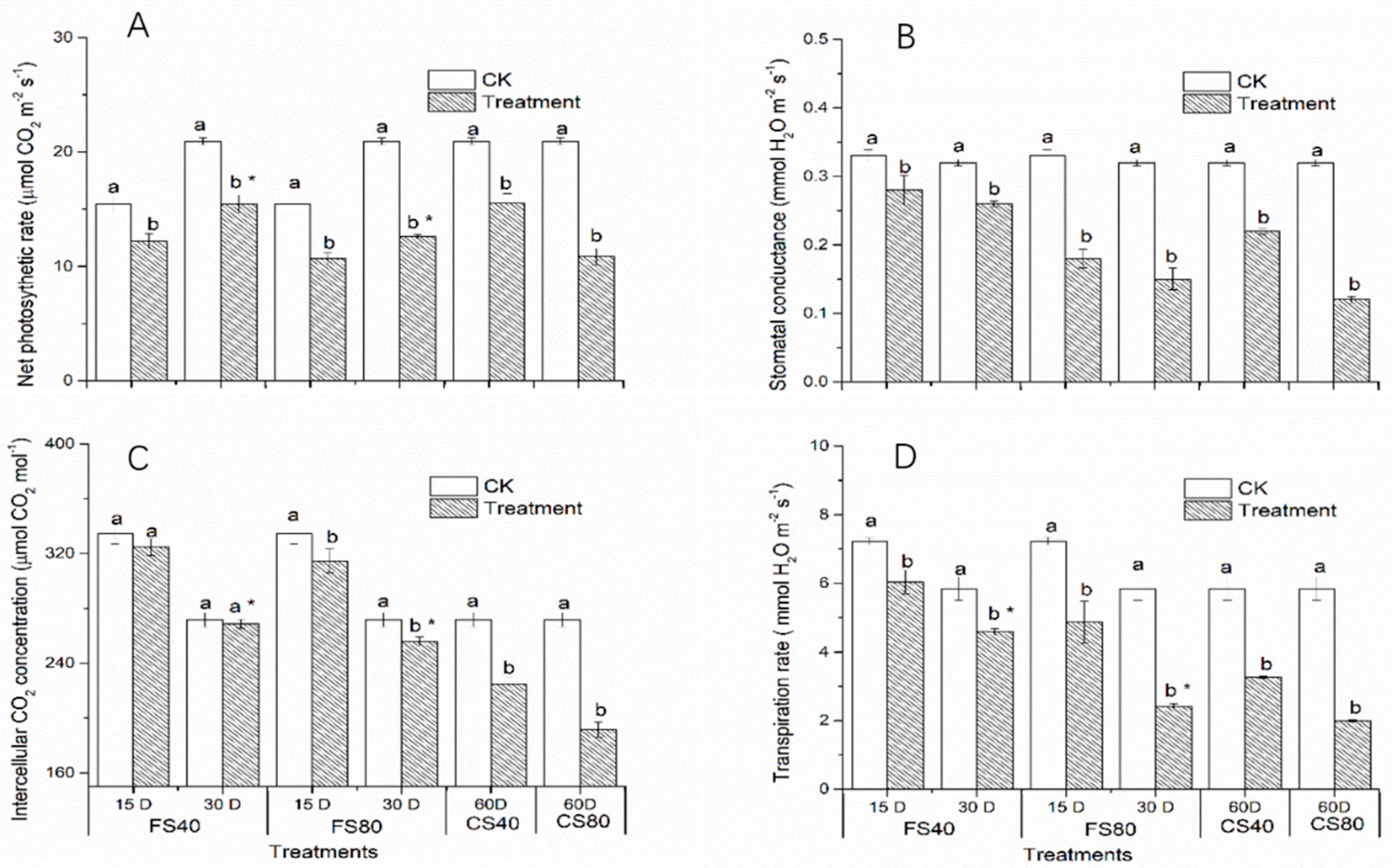
4.4. Analyses of Photosynthesis Parameters
4.5. Measurement of Peanut Yield
4.6. Analysis of Plant Hormone
4.7. RNA Extraction, mRNA Sequencing and Data Deposition
4.8. Analyses of Transcript Assembly, Abundance, Gene Ontology and Pathway
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kong, X. China must protect high-quality arable land. Nature 2014, 506, 7. [Google Scholar] [CrossRef] [PubMed]
- Bajželj, B.; Richards, K.S.; Allwood, J.M.; Smith, P.; Dennis, J.S.; Curmi, E.; Gilligan, C.A. Importance of food-demand management for climate mitigation. Nat. Clim. Chang. 2014, 4, 924–929. [Google Scholar] [CrossRef] [Green Version]
- Brooker, R.W.; Bennett, A.E.; Cong, W.; Daniell, T.; George, T.S.; Hallett, P.; Hawes, C.; Iannetta, P.; Jones, H.; Karley, A.J.; et al. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytol. 2014, 206, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Yong, T.; Chen, P.; Dong, Q.; Du, Q.; Yang, F.; Wang, X.-C.; Liu, W.-G.; Yang, W.-Y. Optimized nitrogen application methods to improve nitrogen use efficiency and nodule nitrogen fixation in a maize-soybean relay intercropping system. J. Integr. Agric. 2018, 17, 664–676. [Google Scholar] [CrossRef]
- Du, J.; Han, T.; Gai, J.-Y.; Yong, T.; Sun, X.; Wang, X.-C.; Yang, F.; Liu, J.; Shu, K.; Liu, W.-G.; et al. Maize-soybean strip intercropping: Achieved a balance between high productivity and sustainability. J. Integr. Agric. 2018, 17, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.-D.; Zhou, L.; Chen, P.; Du, Q.; Pang, T.; Song, C.; Wang, X.-C.; Liu, W.-G.; Yang, W.-Y.; Yong, T. Effects of maize-soybean relay intercropping on crop nutrient uptake and soil bacterial community. J. Integr. Agric. 2019, 18, 2006–2018. [Google Scholar] [CrossRef]
- Li, Y.-H.; Shi, D.-Y.; Li, G.-H.; Zhao, B.; Zhang, J.-W.; Liu, P.; Ren, B.; Dong, S. Maize/peanut intercropping increases photosynthetic characteristics, 13C-photosynthate distribution, and grain yield of summer maize. J. Integr. Agric. 2019, 18, 2219–2229. [Google Scholar] [CrossRef]
- Chi, B.; Zhang, Y.; Zhang, N.; Zhang, X.; Dai, J.; Dong, H. Wide-strip intercropping of cotton and peanut combined with strip rotation increases crop productivity and economic returns. Field Crop. Res. 2019, 243, 107617. [Google Scholar] [CrossRef]
- Wang, X.; Feng, Y.; Yu, L.; Shu, Y.; Tan, F.; Gou, Y.; Luo, S.; Yang, W.; Li, Z.; Wang, J. Sugarcane/soybean intercropping with reduced nitrogen input improves crop productivity and reduces carbon footprint in China. Sci. Total. Environ. 2020, 719, 137517. [Google Scholar] [CrossRef]
- Liu, W.-G.; Hussain, S.; Liu, T.; Zou, J.-L.; Ren, M.-L.; Zhou, T.; Liu, J.; Yang, F.; Yang, W. Shade stress decreases stem strength of soybean through restraining lignin biosynthesis. J. Integr. Agric. 2019, 18, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Roig-Villanova, I.; Khan, S.; Shanahan, H.; Quail, P.H.; Martínez-García, J.F.; Devlin, P.F. A novel high-throughput in vivo molecular screen for shade avoidance mutants identifies a novel phyA mutation. J. Exp. Bot. 2011, 62, 2973–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallemi, M.; Galstyan, A.; Paulišić, S.; Then, C.; Ferrández-Ayela, A.; Lorenzo-Orts, L.; Roig-Villanova, I.; Wang, X.; Micol, J.L.; Ponce, M.R.; et al. DRACULA2 is a dynamic nucleoporin with a role in regulating the shade avoidance syndrome in Arabidopsis. Development 2016, 143, 1623–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, K.A.; Whitelam, G.C. Phytochromes and shade-avoidance eesponses in plants. Ann. Bot. 2005, 96, 169–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef] [PubMed]
- Casal, J.J. Shade avoidance. Arab. Book 2012, 10, e0157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medzihradszky, M.; Bindics, J.; Ádám, É.; Viczián, A.; Klement, É.; Lorrain, S.; Gyula, P.; Mérai, Z.; Fankhauser, C.; Medzihradszky, K.F.; et al. Phosphorylation of phytochrome B inhibits light-induced signaling via accelerated dark reversion in Arabidopsis. Plant Cell 2013, 25, 535–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devlin, P.F.; Patel, S.R.; Whitelam, G.C. Phytochrome E influences internode elongation and flowering time in Arabidopsis. Plant Cell 1998, 10, 1479. [Google Scholar] [CrossRef] [Green Version]
- Salter, M.G.; Franklin, K.A.; Whitelam, G.C. Gating of the rapid shade-avoidance response by the circadian clock in plants. Nature 2003, 426, 680–683. [Google Scholar] [CrossRef]
- Gendron, J.M.; Pruneda-Paz, J.L.; Doherty, C.J.; Gross, A.M.; Kang, S.E.; Kay, S.A. Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor. Proc. Natl. Acad. Sci. USA 2012, 109, 3167–3172. [Google Scholar] [CrossRef] [Green Version]
- Devlin, P.F.; Yanovsky, M.J.; A Kay, S. A Genomic analysis of the shade avoidance response in arabidopsis. Plant Physiol. 2003, 133, 1617–1629. [Google Scholar] [CrossRef] [Green Version]
- Cagnola, J.I.; Ploschuk, E.; Benech-Arnold, T.; Finlayson, S.A.; Casal, J.J. Stem transcriptome reveals mechanisms to reduce the energetic cost of shade-avoidance responses in tomato. Plant Physiol. 2012, 160, 1110–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellaro, R.; Pacín, M.; Casal, J.J. Meta-analysis of the transcriptome reveals a core set of shade-avoidance genes in arabidopsis. Photochem. Photobiol. 2017, 93, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Kohnen, M.V.; Schmid-Siegert, E.; Trevisan, M.; Petrolati, L.A.; Sénéchal, F.; Müller-Moulé, P.; Maloof, J.N.; Xenarios, I.; Fankhauser, C. Neighbor detection induces organ-specific transcriptomes, revealing patterns underlying hypocotyl-specific growth. Plant Cell 2016, 28, 2889–2904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranade, S.; Delhomme, N.; García-Gil, M.R. Transcriptome analysis of shade avoidance and shade tolerance in conifers. Planta 2019, 250, 299–318. [Google Scholar] [CrossRef] [Green Version]
- Maron, L. Saving for the future: Alfalfa delays flowering in the shade. Plant J. 2019, 99, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Molina-Contreras, M.J.; Paulišić, S.; Then, C.; Moreno-Romero, J.; Pastor-Andreu, P.; Morelli, L.; Roig-Villanova, I.; Jenkins, H.; Hallab, A.; Gan, X.; et al. Photoreceptor activity contributes to contrasting responses to shade in cardamine and arabidopsis seedlings. Plant Cell 2019, 31, 2649–2663. [Google Scholar] [CrossRef]
- Shi, Q.; Kong, F.; Zhang, H.; Jiang, Y.; Heng, S.; Liang, R.; Ma, L.; Liu, J.; Lu, X.; Li, P.; et al. Molecular mechanisms governing shade responses in maize. Biochem. Biophys. Res. Commun. 2019, 516, 112–119. [Google Scholar] [CrossRef]
- Zhang, N.; Van Westreenen, A.; Anten, N.P.R.; Evers, J.B.; Marcelis, L.F.M. Disentangling the effects of photosynthetically active radiation and red to far-red ratio on plant photosynthesis under canopy shading. A simulation study using a functional-structural plant model. Ann. Bot. 2019. [Google Scholar] [CrossRef]
- Christophe, A.; Moulia, B.; Varlet-Grancher, C. Quantitative contributions of blue light and PAR to the photocontrol of plant morphogenesis in Trifolium repens (L.). J. Exp. Bot. 2006, 57, 2379–2390. [Google Scholar] [CrossRef] [Green Version]
- Hammons, R.O.; Herman, D.; Stalker, H.T. Chapter 1—Origin and early history of the peanut. In Peanuts; Stalker, H.T., Wilson, R.F., Eds.; AOCS Press: Urbana, IL, USA, 2016; pp. 1–26. [Google Scholar]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.S.; Liu, X.; Gao, N.; Clevenger, J.; Dash, S.; et al. The genome sequences of Arachis duranensis and Arachis ipaensis, the diploid ancestors of cultivated peanut. Nat. Genet. 2016, 48, 438–446. [Google Scholar] [CrossRef]
- Chen, X.; Li, H.; Pandey, M.K.; Yang, Q.; Wang, X.; Garg, V.; Li, H.; Chi, X.; Doddamani, D.; Hong, Y.; et al. Draft genome of the peanut A-genome progenitor (Arachis duranensis) provides insights into geocarpy, oil biosynthesis, and allergens. Proc. Natl. Acad. Sci. USA 2016, 113, 6785–6790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, W.; Chen, H.; Yang, M.; Wang, J.-P.; Pandey, M.K.; Zhang, C.; Chang, W.-C.; Zhang, L.; Zhang, X.; Tang, R.; et al. The genome of cultivated peanut provides insight into legume karyotypes, polyploid evolution and crop domestication. Nat. Genet. 2019, 51, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Dudchenko, O.; Gao, D.; Seijo, G.; Leal-Bertioli, S.C.M.; Ren, L.; Farmer, A.D.; Pandey, M.K.; et al. The genome sequence of segmental allotetraploid peanut Arachis hypogaea. Nat. Genet. 2019, 51, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Lu, Q.; Liu, H.; Zhang, J.; Hong, Y.; Lan, H.; Li, H.; Wang, J.; Liu, H.; Li, S.; et al. Sequencing of cultivated peanut, Arachis hypogaea, yields insights into genome evolution and oil improvement. Mol. Plant 2019, 12, 920–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirza, M.M.Q. Climate change and extreme weather events: Can developing countries adapt? Clim. Policy 2003, 3, 233–248. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 002832. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, B.-Y.; Zhao, Q.; Sun, X.; Li, Y.; Duan, Z.; Miao, X.; Luo, S.; Li, J. Genomic variance and transcriptional comparisons reveal the mechanisms of leaf color affecting palatability and stressed defense in tea plant. Genes 2019, 10, 929. [Google Scholar] [CrossRef] [Green Version]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [Green Version]
- Combes, D.; Monsan, P. Sucrose hydrolysis by invertase. Characterization of products and substrate inhibition. Carbohydr. Res. 1983, 117, 215–228. [Google Scholar] [CrossRef]
- Sakakibara, M.; Wang, D.; Takahashi, R.; Takahashi, K.; Mori, S. Influence of ultrasound irradiation on hydrolysis of sucrose catalyzed by invertase. Enzym. Microb. Technol. 1996, 18, 444–448. [Google Scholar] [CrossRef]
- Akgöl, S.; Kaçar, Y.; Denizli, A.; Arıca, M. Hydrolysis of sucrose by invertase immobilized onto novel magnetic polyvinylalcohol microspheres. Food Chem. 2001, 74, 281–288. [Google Scholar] [CrossRef]
- Yang, C.; Li, L. Hormonal regulation in shade avoidance. Front. Plant Sci. 2017, 8, 1527. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Gray, W.M. SAUR Proteins as effectors of hormonal and environmental signals in plant growth. Mol. Plant 2015, 8, 1153–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- To, J.; Deruère, J.; Maxwell, B.B.; Morris, V.F.; Hutchison, C.E.; Ferreira, F.J.; Schaller, G.E.; Kieber, J.J. Cytokinin regulates type-A arabidopsis response regulator activity and protein stability via two-component phosphorelay. Plant Cell 2007, 19, 3901–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, S.; Fu, Y.; He, T.; Wang, X. Analysis of dynamic global transcriptional atlas reveals common regulatory networks of hormones and photosynthesis across nicotiana varieties in response to long-term drought. Front. Plant Sci. 2020, 11, 672. [Google Scholar] [CrossRef] [PubMed]
- Courbier, S.; Pierik, R. Canopy light quality modulates stress responses in plants. iScience 2019, 22, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Li, G. Auxin-dependent cell elongation during the shade avoidance response. Front. Plant Sci. 2019, 10, 914. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, H.; Deng, T.; Liu, Z.; Wang, X. Time-coursed transcriptome analysis identifies key expressional regulation in growth cessation and dormancy induced by short days in Paulownia. Sci. Rep. 2019, 9, 16602. [Google Scholar] [CrossRef]
- Yang, D.; Seaton, D.D.; Krahmer, J.; Halliday, K.J. Photoreceptor effects on plant biomass, resource allocation, and metabolic state. Proc. Natl. Acad. Sci. USA 2016, 113, 7667–7672. [Google Scholar] [CrossRef] [Green Version]
- Devlin, P.F. Plants wait for the lights to change to red. Proc. Natl. Acad. Sci. USA 2016, 113, 7301–7303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortell, J.M.; Kennedy, J.A. Effect of shading on accumulation of flavonoid compounds in (Vitis vinifera L.) pinot noir fruit and extraction in a model system. J. Agric. Food Chem. 2006, 54, 8510–8520. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Welti, R.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Dong, J.; Bennetzen, J.L.; Zhong, M.; Yang, J.; Zhang, J.; Li, S.; Hao, X.; Zhang, Z.; Wang, X. Integrating transcriptome and microRNA analysis identifies genes and microRNAs for AHO-induced systemic acquired resistance in N. tabacum. Sci. Rep. 2017, 7, 12504. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Stem | Stem | Internode | Leaf Number | ||
|---|---|---|---|---|---|---|
| Stage | Shade | Duration | Length (cm) | Diameter (cm) | Length (cm) | (Count) |
| Seedling | CK | 15 | 13.8 ± 0.60c | NA | NA | 5.67 ± 0.58a |
| S40 | 15 | 16.4 ± 0.28b | NA | NA | 6.67 ± 0.58a | |
| S80 | 15 | 18.0 ± 0.28a | NA | NA | 7.00 ± 0.00a | |
| CK | 30 | 15.9 ± 0.40c | NA | NA | 7.33 ± 0.58a | |
| S40 | 30 | 18.7 ± 0.28b * | NA | NA | 7.67 ± 0.58a | |
| S80 | 30 | 20.2 ± 0.14a * | NA | NA | 8.00 ± 0.00a * | |
| Flowering | CK | 15 | 15.8 ± 0.23c | 0.45 ± 0.00a | 2.13 ± 0.25c | 9.00 ± 0.00a |
| S40 | 15 | 21.2 ± 0.75b | 0.40 ± 0.01b | 2.83 ± 0.21b | 11.33 ± 0.57a | |
| S80 | 15 | 28.3 ± 0.35a | 0.33 ± 0.01c | 3.70 ± 0.10a | 12.50 ± 2.12a | |
| CK | 30 | 17.9 ± 0.20c | 0.34 ± 0.01a | 2.38 ± 0.17c | 10.33 ± 0.57a | |
| S40 | 30 | 24.6 ± 0.47b* | 0.32 ± 0.01b * | 3.41 ± 0.17b | 11.85 ± 0.30a | |
| S80 | 30 | 30.8 ± 0.66a | 0.29 ± 0.01c | 4.07 ± 0.16a | 13.10 ± 1.21a | |
| Seedling | CK | 60 | 17.9 ± 0.20c | 0.34 ± 0.06a | 2.38 ± 0.17c | 10.33 ± 0.57b |
| and | S40 | 60 | 31.4 ± 0.38b | 0.30 ± 0.01b | 3.47 ± 0.22b | 15.00 ± 1.00a |
| flowering | S80 | 60 | 39.2 ± 0.54a | 0.28 ± 0.01c | 4.33 ± 0.22a | 14.50 ± 0.71a |
| Stage | Shade | Duration (Day) | Pods Per Plant | 100-Pod Weight (g) | 100-Kernel Weight (g) |
|---|---|---|---|---|---|
| Seedling | CK | 15 | 15.30 ± 0.90a | 114.80 ± 0.71a | 64.85 ± 1.07a |
| S40 | 15 | 10.00 ± 0.60b | 106.69 ± 1.41b | 55.81 ± 0.60b | |
| S80 | 15 | 8.70 ± 0.90b | 101.25 ± 2.15c | 44.93 ± 1.14c | |
| CK | 30 | 15.30 ± 0.90a | 114.80 ± 0.70a | 64.85 ± 1.07a | |
| S40 | 30 | 5.70 ± 0.70b ** | 100.35 ± 0.54b * | 44.87 ± 1.09b ** | |
| S80 | 30 | 5.30 ± 0.90b * | 96.58 ± 0.74c | 41.06 ± 1.04c | |
| Flowering | CK | 15 | 15.30 ± 0.90a | 114.80 ± 0.70a | 64.85 ± 1.07a |
| S40 | 15 | 6.00 ± 1.00b | 95.00 ± 2.22b | 49.58 ± 1.19b | |
| S80 | 15 | 5.3 ± 0.30b | 88.87 ± 1.74c | 39.67 ± 1.45c | |
| CK | 30 | 15.30 ± 0.90a | 114.80 ± 0.71a | 64.85 ± 1.07a | |
| S40 | 30 | 2.30 ± 0.90b * | 91.05 ± 1.17b | 40.30 ± 1.44b * | |
| S80 | 30 | 2.70 ± 0.70b * | 84.46 ± 1.17c | 36.14 ± 0.57c | |
| Seedling | CK | 60 | 15.30 ± 0.90a | 114.80 ± 0.71a | 64.85 ± 1.07a |
| and | S40 | 60 | 1.30 ± 0.90b | 81.24 ± 1.53b | 32.14 ± 1.68b |
| Flowering | S80 | 60 | 1.30 ± 0.70b | 67.67 ± 1.46c | 28.66 ± 0.62b |
| Gene_ID | Symbol $ | Description of Encoded Proteins | K_ID # | |
|---|---|---|---|---|
| 1 | 14YEDZ | CAB13 | light-harvesting complex II chlorophyll a/b binding protein 3 | K08914 |
| 2 | AFZN0R | CAB13 | light-harvesting complex II chlorophyll a/b binding protein 3 | K08914 |
| 3 | BU7NMG | CAB13 | light-harvesting complex II chlorophyll a/b binding protein 3 | K08914 |
| 4 | JKC32H | CAB13 | light-harvesting complex II chlorophyll a/b binding protein 3 | K08914 |
| 5 | QXM1B7 | CAB3 | light-harvesting complex II chlorophyll a/b binding protein 3 | K08912 |
| 6 | YZ06AV | CAB3 | light-harvesting complex II chlorophyll a/b binding protein 3 | K08912 |
| 7 | BTM1YE | CAB6A | light-harvesting complex II chlorophyll a/b binding protein 3 | K08907 |
| 8 | NKTC04 | CAB6A | light-harvesting complex II chlorophyll a/b binding protein 3 | K08907 |
| 9 | 9BXG3M | CAB7 | light-harvesting complex I chlorophyll a/b binding protein 2 | K08908 |
| 10 | AAY07D | CAB7 | light-harvesting complex I chlorophyll a/b binding protein 2 | K08908 |
| 11 | EB48YM | CAB7 | light-harvesting complex I chlorophyll a/b binding protein 2 | K08908 |
| 12 | 66SDN0 | CAB8 | light-harvesting complex I chlorophyll a/b binding protein 3 | K08909 |
| 13 | 86F2TK | CAB8 | light-harvesting complex I chlorophyll a/b binding protein 3 | K08909 |
| 14 | 4KW7H9 | CAP10A | light-harvesting complex II chlorophyll a/b binding protein 6 | K08917 |
| 15 | JVEI5Y | CAP10A | light-harvesting complex II chlorophyll a/b binding protein 6 | K08917 |
| 16 | N8DZQ8 | LHCA-P4 | light-harvesting complex I chlorophyll a/b binding protein 4 | K08910 |
| 17 | RLTX4G | LHCA-P4 | light-harvesting complex I chlorophyll a/b binding protein 4 | K08910 |
| 18 | YZB02J | LHCA6 | light-harvesting complex I chlorophyll a/b binding protein 2 | K08908 |
| 19 | 6DQ39T | LHCB4.1 | light-harvesting complex II chlorophyll a/b binding protein 4 | K08915 |
| 20 | CVJ130 | LHCB4.1 | light-harvesting complex II chlorophyll a/b binding protein 4 | K08915 |
| 21 | 2C7VNA | LHCB4.3 | light-harvesting complex II chlorophyll a/b binding protein 4 | K08915 |
| 22 | 63GP52 | LHCB4.3 | light-harvesting complex II chlorophyll a/b binding protein 4 | K08915 |
| 23 | 8LQR2U | LHCB5 | light-harvesting complex II chlorophyll a/b binding protein 5 | K08916 |
| 24 | J9AJ01 | LHCB5 | light-harvesting complex II chlorophyll a/b binding protein 5 | K08916 |
| Gene_ID | Symbol $ | Description of Encoded Proteins | K_ID # |
|---|---|---|---|
| AF7BT4 | atpA | ATP synthase F1, alpha subunit | K02111 |
| 487MWI | atpB | ATP synthase, F1 beta subunit | K02112 |
| KPH33C | atpI | ATP synthase subunit A | K02108 |
| 26BATJ | Os03g0784700 | Ferredoxin-NADP reductase family protein | K02641 |
| Z0BJS7 | petA | chloroplast envelope membrane protein-like isoform X2 [Glycine max] | K02634 |
| B5AM0N | petB | photosynthetic electron transfer B | K02704 |
| GZ00NU | petD | photosynthetic electron transfer D | K02637 |
| E2679H | PETH | ferredoxin-NADP(+)-oxidoreductase 1 | K02641 |
| ZQ3F6U | PETH | ferredoxin-NADP(+)-oxidoreductase 1 | K02641 |
| WM01M4 | PNSL2 | oxygen-evolving enhancer protein | K08901 |
| 3I3T69 | psaA | photosystem I P700 chlorophyll A apoprotein | K02689 |
| 7VZ02V | psaA | photosystem I P700 chlorophyll A apoprotein | K02689 |
| R5AFBV | psaA | photosystem I P700 chlorophyll A apoprotein | K02690 |
| UAEM0K | psaB | photosystem I P700 chlorophyll A apoprotein | K02690 |
| QW0L8Q | PSAE-1 | photosystem I reaction center subunit IV A | K02693 |
| VHAI7W | PSAF | photosystem I reaction center subunit III | K02694 |
| 4PD0PP | PSAG | photosystem I reaction center subunit V | K08905 |
| 71HCYD | PSAH2 | photosystem I reaction center subunit VI | K02695 |
| YBSA05 | PSAH2 | photosystem I reaction center subunit VI | K02695 |
| 9042LP | PSAK | photosystem I reaction center subunit X psaK | K02698 |
| ZS1M1K | PSAK | photosystem I reaction center subunit X psaK | K02698 |
| 18YQBF | PSAL | photosystem I reaction center subunit XI | K02699 |
| 0R479N | psbA | photosystem II protein D1 [Glycine max] | K02703 |
| 0W708Z | psbB | photosystem II CP47 chlorophyll A apoprotein | K02704 |
| 8A6KBK | psbC | Photosystem II chlorophyll-binding protein CP43 | K02705 |
| 92HTUM | psbC | photosystem II CP43 chlorophyll apoprotein | K02705 |
| 49ZYT6 | PSBO | photosystem II oxygen-evolving enhancer protein | K02716 |
| 30A6BG | PSBP | 23kDa polypeptide of the oxygen evolving complex of photosystem II | K02717 |
| L9FPHH | PSBS | photosystem II 22 kDa protein, chloroplastic-like [Glycine max] | K03542 |
| QP7RRL | PSBS | photosystem II 22 kDa protein, chloroplastic-like [Glycine max] | K03542 |
| J2LHWB | PSBY | photosystem II core complex family psbY protein | K02723 |
| CW3FX9 | SEND33 | ferredoxin 1 | K02639 |
| I8TN6N | YMF19 | ATPase subunit 8 (mitochondrion) [Glycine max] | K02109 |
| Gene ID | Encoded Enzyme | KEGG_ID | Abbreviation | Enzyme | Function |
|---|---|---|---|---|---|
| L6ZHES | glucose-1-phosphate adenylyltransferase 1 | K05349 | AGPS1 | [EC:3.2.1.21] | Glycosidases that hydrolyse O- and S-glycosyl compounds |
| V374FJ | glucose-1-phosphate adenylyltransferase 1 | K05349 | AGPS1 | [EC:3.2.1.21] | Glycosidases that hydrolyse O- and S-glycosyl compounds |
| PX7TTT | glucose-1-phosphate adenylyltransferase | K00975 | glgC | [EC:2.7.7.27] | Transferring phosphorus-containing groups for glycogen synthesis |
| 4Y2508 | beta-amylase 1 | K01177 | BAM1 | [EC:3.2.1.2] | hydrolyse O- and S-glycosyl compounds |
| 4Z6VDH | beta glucosidase 15 | K01188 | BGLU12 | [EC:2.4.1.1] | hydrolyse O- and S-glycosyl compounds |
| M8S3TB | beta glucosidase 15 | K01188 | BGLU12 | [EC:2.4.1.1] | hydrolyse O- and S-glycosyl compounds |
| YDQ82R | beta glucosidase 15 | K01188 | BGLU12 | [EC:2.4.1.1] | hydrolyse O- and S-glycosyl compounds |
| IXM19M | glycogen phosphorylase 1-like isoform X1 | K00688 | glpV | [EC:2.4.1.1] | Glycogen degradation, glycogen => glucose-6P |
| 0Q5BJB | beta-fructofuranosidase 5; or Invertase | K01193 | INV | [EC:3.2.1.26] | hydrolyse O- and S-glycosyl compounds, e.g. sucrose |
| 4VZ14Z | sucrose synthase 4 | K00695 | SS | [EC:2.4.1.13] | sucrose synthesis and hydrolysis |
| 0FY2NM | trehalose-6-phosphate phosphatase | K01087 | TPS | [EC:3.1.3.12] | catalyze trehalose-6p to trehalose |
| G2WE73 | trehalose-6-phosphate phosphatase | K01087 | TPS | [EC:3.1.3.12] | catalyze trehalose-6p to trehalose |
| Treatments | Root | Stem | Leaf | Pod | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Stage | Shade | Duration | Dry Weight (g) | Ratio (%) | Dry Weight (g) | Ratio (%) | Dry Weight (g) | Ratio (%) | Dry weight (g) | Ratio (%) |
| Seedling | CK | 15 | 0.19 ± 0.03a | 8.39 | 0.55 ± 0.07a | 24.10 | 1.55 ± 0.48a | 67.51 | NA | NA |
| S40 | 15 | 0.16 ± 0.05a | 12.89 | 0.42 ± 0.00a | 34.46 | 0.64 ± 0.08b | 52.64 | NA | NA | |
| S80 | 15 | 0.07 ± 0.01b | 13.64 | 0.15 ± 0.01b | 30.87 | 0.27 ± 0.01b | 55.49 | NA | NA | |
| CK | 30 | 0.63 ± 0.07a | 6.6.1 | 3.74 ± 0.29a | 39.30 | 5.15 ± 0.43a | 54.08 | NA | NA | |
| S40 | 30 | 0.31 ± 0.02b * | 5.06 | 2.67 ± 0.10b ** | 43.67 | 3.14 ± 0.45b ** | 51.26 | NA | NA | |
| S80 | 30 | 0.28 ± 0.03b ** | 9.53 | 1.11 ± 0.15c * | 37.26 | 1.58 ± 0.36c * | 53.21 | NA | NA | |
| Flowering | CK | 15 | 0.63 ± 0.07a | 6.62 | 3.74 ± 0.29a | 39.29 | 5.15 ± 0.43a | 54.10 | NA | NA |
| S40 | 15 | 0.44 ± 0.02b | 5.25 | 3.52 ± 0.17a | 42.00 | 4.42 ± 0.16a | 52.74 | NA | NA | |
| S80 | 15 | 0.26 ± 0.00c | 4.68 | 2.21 ± 0.13b | 39.82 | 3.08 ± 0.12b | 55.50 | NA | NA | |
| CK | 30 | 0.68 ± 0.05a | 6.07 | 3.78 ± 0.28a | 33.72 | 5.31 ± 0.56a | 47.37 | 1.44 ± 0.12a | 12.85 | |
| S40 | 30 | 0.46 ± 0.05b | 7.11 | 2.69 ± 0.24b * | 41.58 | 2.88 ± 0.34b * | 44.51 | 0.44 ± 0.07b | 6.80 | |
| S80 | 30 | 0.31 ± 0.06c | 6.95 | 1.71 ± 0.23c | 38.34 | 2.19 ± 0.19c * | 49.10 | 0.35 ± 0.10b | 5.61 | |
| Seedling | CK | 60 | 0.68 ± 0.05a | 6.07 | 3.78 ± 0.28a | 33.72 | 5.31 ± 0.56a | 47.37 | 1.44 ± 0.12a | 12.85 |
| and | S40 | 60 | 0.33 ± 0.13b | 6.52 | 2.36 ± 0.17b | 46.64 | 2.10 ± 0.09b | 41.50 | 1.07 ± 0.07b | 5.34 |
| flowering | S80 | 60 | 0.16 ± 0.02c | 8.12 | 0.86 ± 0.15c | 43.65 | 0.95 ± 0.04c | 48.22 | 0.00 ± 0.00c | 0.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Zhang, H.; Zeng, R.; Wang, X.; Huang, L.; Wang, L.; Wang, X.; Zhang, L. Shade Effects on Peanut Yield Associate with Physiological and Expressional Regulation on Photosynthesis and Sucrose Metabolism. Int. J. Mol. Sci. 2020, 21, 5284. https://doi.org/10.3390/ijms21155284
Chen T, Zhang H, Zeng R, Wang X, Huang L, Wang L, Wang X, Zhang L. Shade Effects on Peanut Yield Associate with Physiological and Expressional Regulation on Photosynthesis and Sucrose Metabolism. International Journal of Molecular Sciences. 2020; 21(15):5284. https://doi.org/10.3390/ijms21155284
Chicago/Turabian StyleChen, Tingting, Huajian Zhang, Ruier Zeng, Xinyue Wang, Luping Huang, Leidi Wang, Xuewen Wang, and Lei Zhang. 2020. "Shade Effects on Peanut Yield Associate with Physiological and Expressional Regulation on Photosynthesis and Sucrose Metabolism" International Journal of Molecular Sciences 21, no. 15: 5284. https://doi.org/10.3390/ijms21155284
APA StyleChen, T., Zhang, H., Zeng, R., Wang, X., Huang, L., Wang, L., Wang, X., & Zhang, L. (2020). Shade Effects on Peanut Yield Associate with Physiological and Expressional Regulation on Photosynthesis and Sucrose Metabolism. International Journal of Molecular Sciences, 21(15), 5284. https://doi.org/10.3390/ijms21155284