Peripheral Dopamine 2-Receptor Antagonist Reverses Hypertension in a Chronic Intermittent Hypoxia Rat Model

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
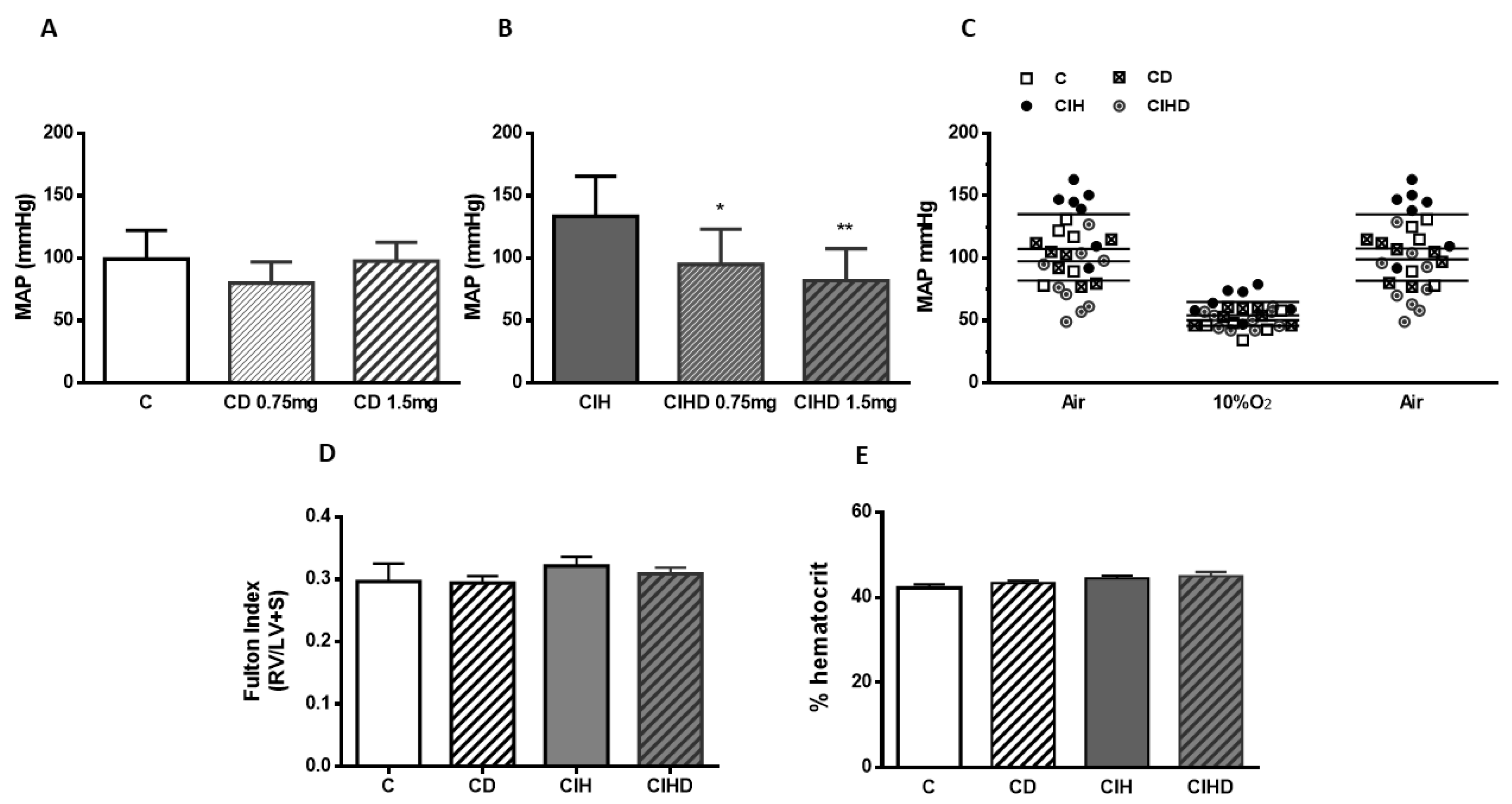
2.1. Effect of Domperidone on CIH Induced Hypertension
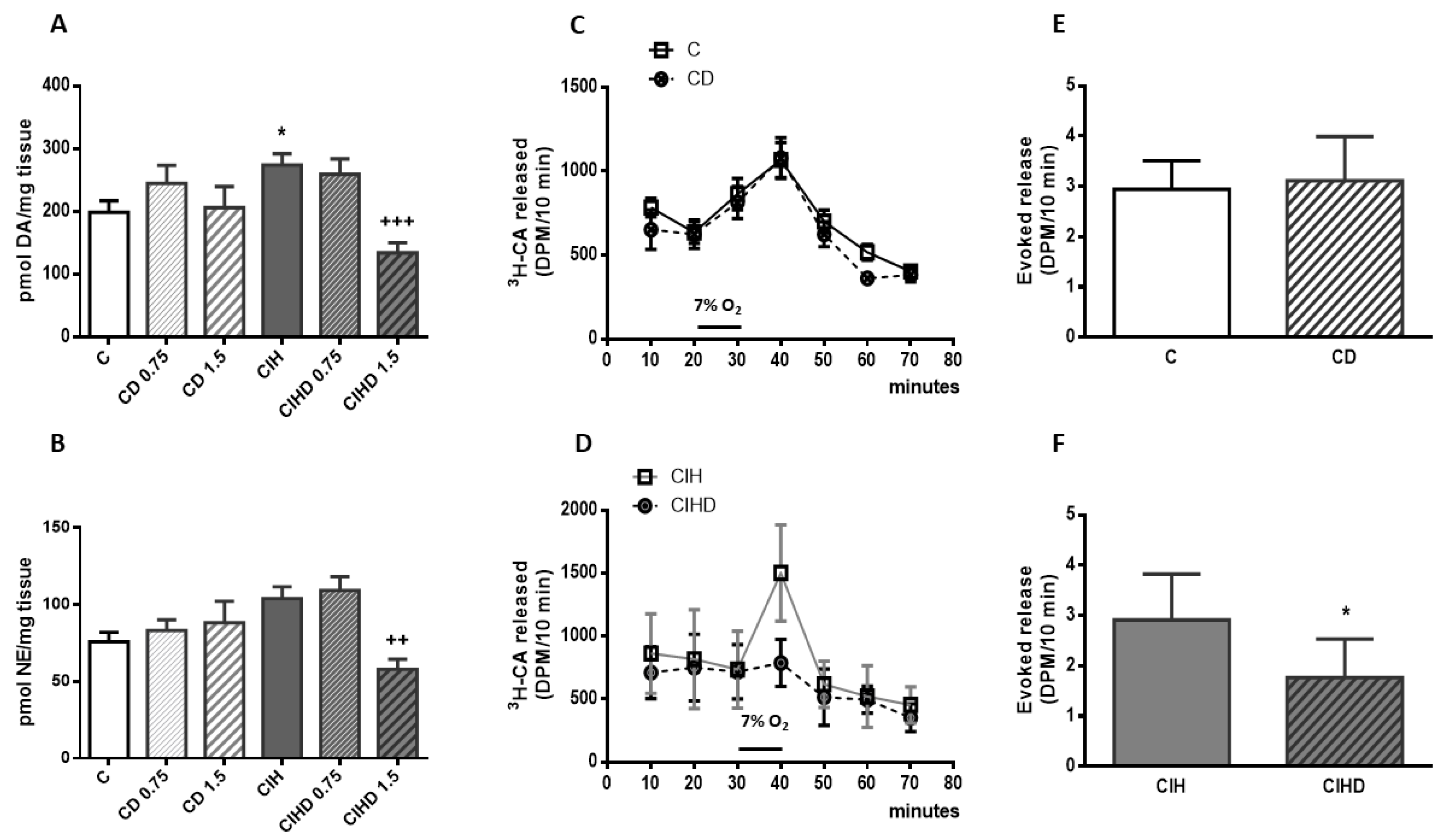
2.2. Effects of Domperidone on CB Catecholamine Content and CB Secretory Activity
2.3. Effects of Domperidone on Respiratory Parameters
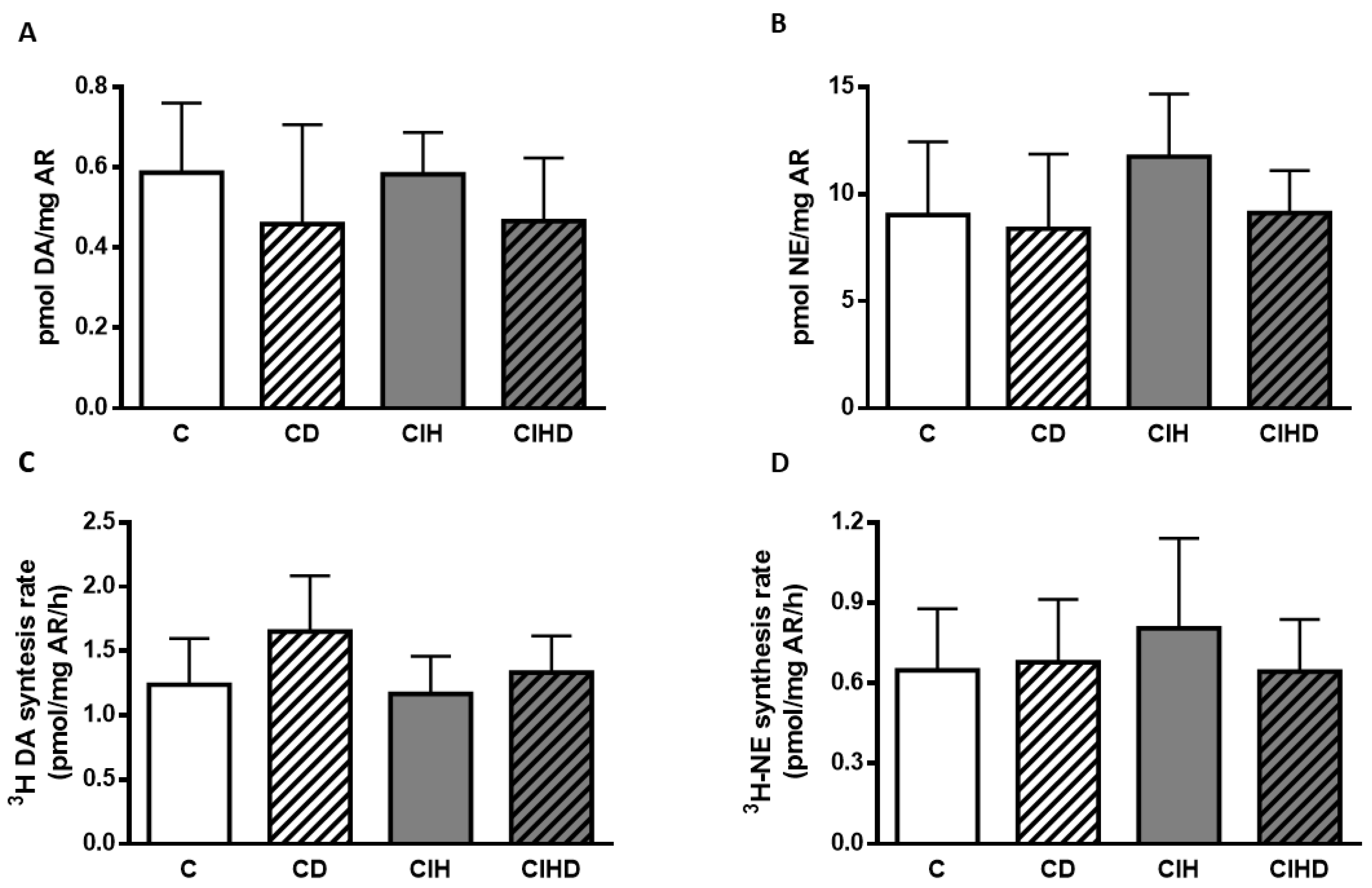
2.4. Effects of Domperidone on CA Content and Rate of CA Synthesis from Renal Artery (RA)
3. Discussion
4. Materials and Methods
4.1. Animals and Anesthesia
4.2. Arterial Blood Pressure Measurement
4.3. Plethysmography
4.4. Endogenous Catecholamine Content
4.5. Stimuli–Evoked Catecholamine Release from CB and Renal Artery Catecholamine Synthesis
4.6. Data Presentation and Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Konecny, T.; Kara, T.; Somers, V.K. Obstructive sleep apnea and hypertension: an update. Hypertension 2014, 63, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, E.C.; Lesske, J.; Qian, W.; Miller, C.C.; Unger, T. Repetitive, episodic hypoxia causes diurnal elevation of blood pressure in rats. Hypertension 1992, 19, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhang, J.; Gan, T.X.; Chen Izu, Y.; Hasday, J.D.; Karmazyn, M.; Balke, C.W.; Scharf, S.M. Left ventricular dysfunction and associated cellular injury in rats exposed to chronic intermittent hypoxia. J. Appl. Physiol. 2008, 104, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, R.; Andrade, D.C.; Lucero, C.; Arias, P.; Iturriaga, R. Carotid Body Ablation Abrogates Hypertension and Autonomic Alterations Induced by Intermittent Hypoxia in Rats. Hypertension 2016, 68, 436–445. [Google Scholar] [CrossRef] [Green Version]
- Mcnicholas, W.T.; Bonsigore, M.R. Management committee of EU cost action B26. Sleep apno ea as an independent risk factor for cardiovascular disease: current evidence, basic mechanisms and research priorities. Eur. Respir. J. 2007, 29, 156–178. [Google Scholar] [CrossRef]
- Fletcher, E.C.; Lesske, J.; Culman, J.; Miller, C.C.; Unger, T. Sympathetic denervation blocks blood pressure elevation in episodic hypoxia. Hypertension 1992, 20, 612–619. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.J.; Yuan, G.; Khan, S.; Nanduri, J.; Makarenko, V.V.; Reddy, V.D.; Vasavda, C.; Kumar, G.K.; Semenza, G.L.; Prabhakar, N.P. Regulation of hypoxia-inducible factor-α isoforms and redox state by carotid body neural activity in rats. J. Physiol. 2014, 592, 3841–3858. [Google Scholar] [CrossRef]
- Narkiewicz, K.; Ratcliffe, L.E.; Hart, E.C.; Briant, L.J.; Chrostowska, M.; Wolf, J.; Szyndler, A.; Hering, D.; Abdala, A.P.; Manghat, N.; et al. Unilateral carotid body resection in resistant hypertension: a safety and feasibility trial. JACC Basic. Transl. Sci. 2016, 1, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Fujii, K.; Saku, K.; Kishi, T.; Oga, Y.; Tohyama, T.; Nishikawa, T.; Sakamoto, T.; Ikeda, M.; Ide, T.; Tsutsui, H.; et al. Carotid Body Denervation Markedly Improves Survival in Rats with Hypertensive Heart Failure. Am. J. Hypertens. 2017, 1, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Iturriaga, R. Carotid Body Ablation: A New Target to Address Central Autonomic Dysfunction. Curr. Hypertens Rep. 2018, 22, 53. [Google Scholar] [CrossRef] [PubMed]
- Lesske, J.; Fletcher, E.C.; Bao, G.; Unger, T. Hypertension caused by chronic intermittent hypoxia--influence of chemoreceptors and sympathetic nervous system. J. Hypertens. 1997, 15, 1593–1603. [Google Scholar]
- Gonzalez, C.; Almaraz, L.; Obeso, A.; Rigual, R. Carotid body chemoreceptors: from natural stimuli to sensory discharges. Physiol. Rev. 1994, 74, 829–898. [Google Scholar] [CrossRef]
- Gauda, E.B. Gene expression in peripheral arterial chemoreceptors. Microsc. Res. Tech. 2002, 59, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Wakai, J.; Takayama, A.; Yokoyama, T.; Nakamuta, N.; Kusakabe, T.; Yamamoto, Y. Immunohistochemical localization of dopamine D2 receptor in the rat carotid body. Acta Histochem. 2015, 117, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Ford, C.P. The role of D2-autoreceptors in regulating dopamine neuron activity and transmission. Neuroscience 2014, 282, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, A.S.; Striet, J.B.; Gudelsky, G.; Soukhova, G.K.; Gozal, E.; Beitner-Johnson, D.; Guo, S.-Z.; Sachleben, L.R.; Haycock, J.W.; Gozal, D.; et al. Regulation of catecholamines by sustained and intermittent hypoxia in neuroendocrine cells and sympathetic neurons. Hypertension 2003, 42, 1130–1136. [Google Scholar] [CrossRef] [Green Version]
- Docio, I.; Olea, E.; Prieto-LLoret, J.; Gallego-Martin, T.; Obeso, A.; Gomez-Niño, A.; Rocher, A. Guinea Pig as a Model to Study the Carotid Body Mediated Chronic Intermittent Hypoxia Effects. Front. Physiol. 2018, 9, 6942018. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Obeso, E.; Docio, I.; Olea, E.; Cogolludo, A.; Obeso, A.; Rocher, A.; Gomez-Niño, A. Guinea Pig Oxygen-Sensing and Carotid Body Functional Properties. Front. Physiol. 2017, 8, 285. [Google Scholar] [CrossRef] [Green Version]
- Laduron, P.M.; Leysen, J.E. Domperidone, a specific in vitro dopamine antagonist, devoid of in vivo central dopaminergic activity. Biochem. Pharmacol. 1979, 28, 2161–2165. [Google Scholar] [CrossRef]
- Kohli, J.D.; Glock, D.; Goldberg, L.I. Selective DA2 versus DA1 antagonist activity of domperidone in the periphery. Eur. J. Pharm. 1983, 89, 137–141. [Google Scholar] [CrossRef]
- Schey, R.; Saadi, M.; Midani, D.; Roberts, A.C.; Parupalli, R.; Parkman, H.P. Domperidone to Treat Symptoms of Gastroparesis: Benefits and Side Effects from a Large Single-Center Cohort. Dig. Dis. Sci. 2016, 61, 3545–3551. [Google Scholar] [CrossRef] [PubMed]
- Quintero, M.; Olea, E.; Conde, S.V.; Obeso, A.; Gallego-Martin, T.; Gonzalez, C.; Montserrat, J.M.; Gomez-Niño, A.; Yubero, S.; Agapito, M.T. Age protects from harmful effects produced by chronic intermittent hypoxia. J. Physiol. 2016, 594, 1773–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrim, R.R.; Bonagamba, L.G.; Machado, B.H. Cardiovascular and respiratory outcome of preconditioned rats submitted to chronic intermittent hypoxia. Exp. Physiol. 2015, 100, 1008–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Gomez-Nino, A.; Gonzalez, C.; Dinger, B.; Fidone, S. Stimulus-specific mobilization of dopamine and norepinephrine stores in cat carotid body. J. Auton. Nerv. Syst. 1997, 67, 109–113. [Google Scholar] [CrossRef]
- Gonzalez-Martín, M.C.; Vega-Agapito, M.V.; Conde, S.V.; Castaneda, J.; Bustamante, R.; Olea, E.; Perez-Vizcaino, F.; Gonzalez, C.; Obeso, A. Carotid body function and ventilatory responses in intermittent hypoxia. Evidence for anomalous brainstem integration of arterial chemoreceptor input. J. Cell. Physiol. 2011, 226, 1961–1969. [Google Scholar] [CrossRef] [Green Version]
- Olea, E.; Agapito, M.T.; Gallego-Martin, T.; Rocher, A.; Gomez-Niño, A.; Obeso, A.; Gonzalez, C.; Yubero, S. Intermittent hypoxia and diet-induced obesity: effects on oxidative status, sympathetic tone, plasma glucose and insulin levels, and arterial pressure. J. Appl. Physiol. (1985) 2014, 117, 706–719. [Google Scholar] [CrossRef] [Green Version]
- Carroll, J.L.; Boyle, K.M.; Wasicko, M.J.; Sterni, L.M. Dopamine D2 receptor modulation of carotid body type 1 cell intracellular calcium in developing rats. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 288, L910–L916. [Google Scholar] [CrossRef]
- Iturriaga, R.; Larraín, C.; Zapata, P. Effects of dopaminergic blockade upon carotid chemosensory activity and its hypoxia-induced excitation. Brain Res. 1994, 663, 145–154. [Google Scholar] [CrossRef]
- Prabhakar, N.R.; Peng, Y.J.; Jacono, F.J.; Kumar, G.K.; Dick, T.E. Cardiovascular alterations by chronic intermittent hypoxia: importance of carotid body chemoreflexes. Clin. Exp. Pharmacol. Physiol. 2005, 32, 447–449. [Google Scholar] [CrossRef]
- Bucolo, C.; Leggio, G.M.; Drago, F.; Salomone, S. Dopamine outside the brain: The eye, cardiovascular system and endocrine pancreas. Pharmacol. Ther. 2019, 203, 107392. [Google Scholar] [CrossRef]
- Larrain, A.; Kapur, V.K.; Gooley, T.A.; Pope, C.E. Pharmacological treatment of obstructive sleep apnea with a combination of pseudoephedrine and domperidone. J. Clin. Sleep Med. 2010, 6, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucking, E.F.; O’Halloran, K.D.; Jones, J.F. Increased cardiac output contributes to the development of chronic intermittent hypoxia-induced hypertension. Exp. Physiol. 2014, 99, 1312–1324. [Google Scholar] [CrossRef]
- Tedjasaputra, V.; Bryan, T.L.; Van Diepen, S.; Moore, L.E.; Bouwsema, M.M.; Welsh, R.C.; Petersen, S.R.; Stickland, M.K. Dopamine receptor blockade improves pulmonary gas exchange but decreases exercise performance in healthy humans. J. Physiol. 2015, 593, 3147–3157. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Prabhakar, N.R. Peripheral chemoreceptors: function and plasticity of the carotid body. Compr. Physiol. 2012, 2, 141–219. [Google Scholar] [PubMed] [Green Version]
- Nurse, C.A. Synaptic and paracrine mechanisms at carotid body arterial chemoreceptors. J. Physiol. 2014, 592, 3419–3426. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, N.R.; Semenza, G.L. Adaptive and maladaptive cardiorespiratory responses to continuous and intermittent hypoxia mediated by hypoxia-inducible factors 1 and 2. Physiol. Rev. 2012, 92, 967–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamboa, J.L.; Macarlupú, J.L.; Rivera-Chira, M.; Monge, C.C.; León-Velarde, F. Effect of domperidone on ventilation and polycythemia after 5 weeks of chronic hypoxia in rats. Respir. Physiol. Neurobiol. 2003, 135, 1–8. [Google Scholar] [CrossRef]
- Julien, C.A.; Joseph, V.; Bairam, A. Alteration of carotid body chemoreflexes after neonatal intermittent hypoxia and caffeine treatment in rat pups. Respir. Physio. Neurobiol. 2011, 177, 301–312. [Google Scholar] [CrossRef]
- Walsh, T.S.; Foo, I.T.; Drummond, G.B.; Warren, P.M. Influence of dose of domperidone on the acute ventilatory response to hypoxia in humans. Br. J. Anaesth. 1998, 81, 322–326. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, S.; Dostal, J.; Erokwu, B.; Han, F.; Dick, T.E.; Strohl, K.P. Domperidone and ventilatory behavior: Sprague–Dawley versus Brown Norway rats. Respir. Physiol. Neurobiol. 2007, 155, 22–28. [Google Scholar] [CrossRef]
- Nakano, H.; Lee, S.D.; Farkas, G.A. Dopaminergic modulation of ventilation in obese Zucker rats. J. Appl. Physiol. (1985) 2002, 92, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Huey, K.A.; Brown, I.P.; Jordan, M.C.; Powell, F.L. Changes in dopamine D(2)-receptor modulation of the hypoxic ventilatory response with chronic hypoxia. Respir. Physiol. 2000, 123, 177–187. [Google Scholar] [CrossRef]
- Prabhakar, N.R.; Kumar, G.K. Mechanisms of sympathetic activation and blood pressure elevation by intermittent hypoxia. Respir. Physiol. Neurobiol. 2010, 174, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Xing, T.; Pilowsky, P.M.; Fong, A.Y. Mechanism of sympathetic activation and blood pressure elevation in humans and animals following acute intermittent hypoxia. Prog. Brain Res. 2014, 209, 131–146. [Google Scholar]
- Iturriaga, R.; Oyarce, M.P.; Dias, A.C.R. Role of Carotid Body in Intermittent Hypoxia-Related Hypertension. Curr. Hypertens Rep. 2017, 19, 38. [Google Scholar] [CrossRef] [PubMed]
- Narkiewicz, K.; Somers, V.K. Sympathetic nerve activity in obstructive sleep apnoea. Acta Physiol. Scand. 2003, 177, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Beecroft, J.; Duffin, J.; Pierratos, A.; Chan, C.T.; McFarlane, P.; Hanly, P.J. Enhanced chemo-responsiveness in patients with sleep apnoea and end-stage renal disease. Eur. Respir. J. 2006, 28, 151–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamisier, T.; Tan, C.O.; Pepin, J.L.; Levy, P.; Taylor, J.A. Blood Pressure Increases in OSA du5 to Maintained Neurovascular Sympathetic Transduction: Impact of CPAP. Sleep 2015, 38, 1973–1980. [Google Scholar] [CrossRef] [Green Version]
- Miglis, M.G. Autonomic dysfunction in primary sleep disorders. Sleep Med. 2016, 19, 40–49. [Google Scholar] [CrossRef]
- Goldstein, D.S.; Holmes, C. Neuronal source of plasma dopamine. Clin. Chem. 2008, 54, 1864–1871. [Google Scholar] [CrossRef] [Green Version]
- Carey, R.M. Theodore Cooper Lecture: Renal dopamine system: paracrine regulator of sodium homeostasis and blood pressure. Hypertension 2001, 38, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Gildea, J.J.; Shah, I.; Weiss, R.; Casscells, N.D.; McGrash, H.E.; Zhang, J.; Jones, J.E.; Felder, R.E. HK-2 human renal proximal tubule cells as a model for G protein-coupled receptor kinase type 4-mediated dopamine 1 receptor uncoupling. Hypertension 2010, 56, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Armando, I.; Villar, V.A.; Jose, P.A. Genomics and Pharmacogenomics of Salt-sensitive Hypertension. Curr. Hypertens Rev. 2015, 11, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorriento, D.; Santulli, G.; Del Giudice, C.; Anastasio, A.; Trimarco, B.; Iaccarino, G. Endothelial Cells Are Able to Synthesize and Release Catecholamines both In Vitro and In Vivo. Hypertension 2012, 60, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jose, P.A.; Asico, L.D.; Eisner, G.M.; Pocchiari, F.; Semeraro, C.; Felder, R.A. Effects of costimulation of dopamine D1- and D2-like receptors on renal function. Am. J. Physiol. 1998, 275, R986–R994. [Google Scholar] [CrossRef] [PubMed]
- Bao, G.; Metreveli, N.; Li, R.; Taylor, A.; Fletcher, E.C. Blood pressure response to chronic episodic hypoxia: Role of the sympathetic nervous system. J. Appl. Physiol. 1997, 83, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L.; Prabhakar, N.R. The role of hypoxia-inducible factors in carotid body (patho) physiology. J. Physiol. 2018, 596, 2977–2983. [Google Scholar] [CrossRef] [Green Version]
- Rocher, A.; Caceres, A.I.; Almaraz, L.; Gonzalez, C. EPAC signalling pathways are involved in low P-O2 chemoreception in carotid body chemoreceptor cells. J. Physiol. 2009, 587, 4015–4027. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animals | C | CD | ||||
| Respiratory Parameter | Breathing Frequency (Breaths min−1) | Tidal Volume (mL kg−1) | Minute Ventilation (mL min−1 kg−1) | Breathing Frequency (Breaths min−1) | Tidal Volume (mL kg−1) | Minute Ventilation (mL min−1 kg−1) |
| 21% O2 (air) | 129 (93) | 4.6 (0.5) | 532 (288) | 114 (47) | 5.4 (0.5) | 564 (178) |
| 12% O2 | 131 (23) | 5.4 (0.7) | 664 (45) | 160 (17) | 5.7 (0.7) | 809 (178) |
| 10% O2 | 139 (30) | 5.1 (0,7) | 921 (100) | 134 (26) | 6.2 (0.6) | 951 (89) |
| 7% O2 | 139 (30) | 8.0 (0.8) | 1066 (201) | 134 (26) | 8.6 (0.8) | 1110 (138) |
| 5% CO2 in air | 150 (24) | 7.9 (0.8) | 1151 (136) | 141 (30) | 7.6 (0.8) | 1041 (200) |
| Animals | CIH | CIHD | ||||
| Respiratory Parameter | Breathing Frequency (Breaths min−1) | Tidal Volume (mL kg−1) | Minute Ventilation (mL min−1 kg−1) | Breathing Frequency (Breaths min−1) | Tidal Volume (mL kg−1) | Minute Ventilation (mL min−1 kg−1) |
| 21% O2 (air) | 92 (31) | 4.9 (0.5) | 409 (82) | 93 (28) | 5.0 (0.9) | 418 (68) |
| 12% O2 | 124 (17) | 5.3 (0.7) | 622 (74) | 119 (17) | 5.4 (0.7) | 624 (106) |
| 10% O2 | 152 (28) | 6.1 (0.8) | 851 (156) | 147 (16) | 6.4 (1.0) | 892 (81) |
| 7% O2 | 121 (12) | 7,6 (1.1) | 886 (126) | 123 (18) | 7.9 (1.1) | 953 (182) |
| 5% CO2 in air | 130 (66) | 5.2 (0.6) | 969 (97) | 132 (17) | 7.6 (1.0) | 984 (152) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olea, E.; Docio, I.; Quintero, M.; Rocher, A.; Obeso, A.; Rigual, R.; Gomez-Niño, A. Peripheral Dopamine 2-Receptor Antagonist Reverses Hypertension in a Chronic Intermittent Hypoxia Rat Model. Int. J. Mol. Sci. 2020, 21, 4893. https://doi.org/10.3390/ijms21144893
Olea E, Docio I, Quintero M, Rocher A, Obeso A, Rigual R, Gomez-Niño A. Peripheral Dopamine 2-Receptor Antagonist Reverses Hypertension in a Chronic Intermittent Hypoxia Rat Model. International Journal of Molecular Sciences. 2020; 21(14):4893. https://doi.org/10.3390/ijms21144893
Chicago/Turabian StyleOlea, Elena, Inmaculada Docio, Miguel Quintero, Asunción Rocher, Ana Obeso, Ricardo Rigual, and Angela Gomez-Niño. 2020. "Peripheral Dopamine 2-Receptor Antagonist Reverses Hypertension in a Chronic Intermittent Hypoxia Rat Model" International Journal of Molecular Sciences 21, no. 14: 4893. https://doi.org/10.3390/ijms21144893
APA StyleOlea, E., Docio, I., Quintero, M., Rocher, A., Obeso, A., Rigual, R., & Gomez-Niño, A. (2020). Peripheral Dopamine 2-Receptor Antagonist Reverses Hypertension in a Chronic Intermittent Hypoxia Rat Model. International Journal of Molecular Sciences, 21(14), 4893. https://doi.org/10.3390/ijms21144893