Heme Oxygenase Protects against Placental Vascular Inflammation and Abortion by the Alarmin Heme in Mice

 ,
,
Abstract
:
1. Introduction
2. Results
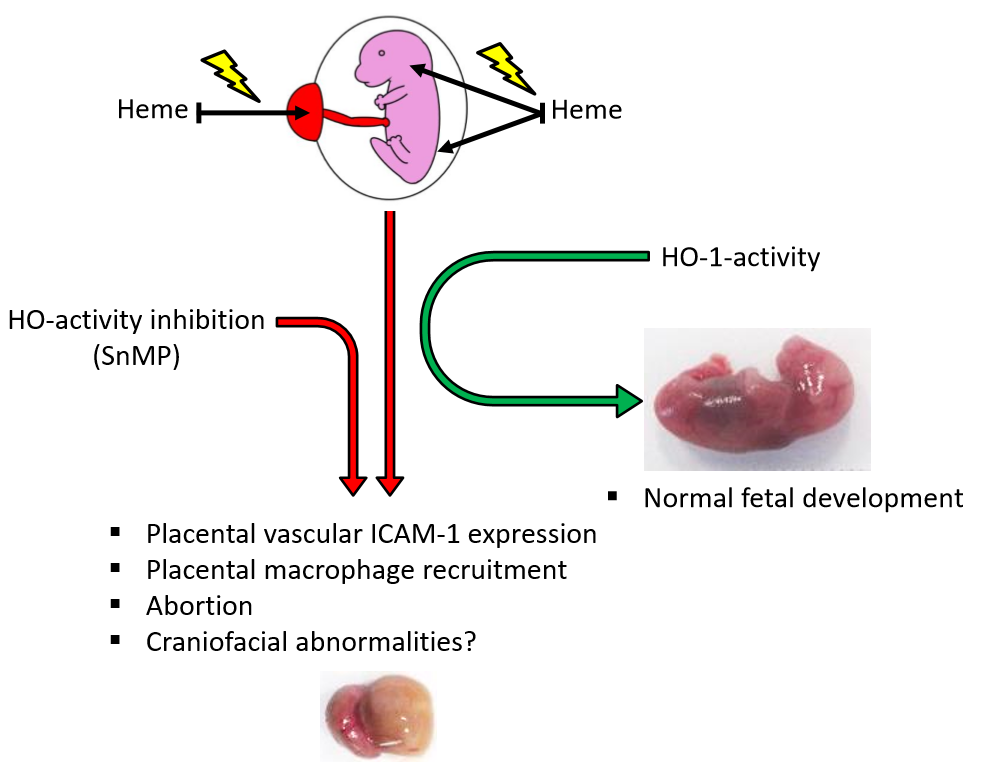
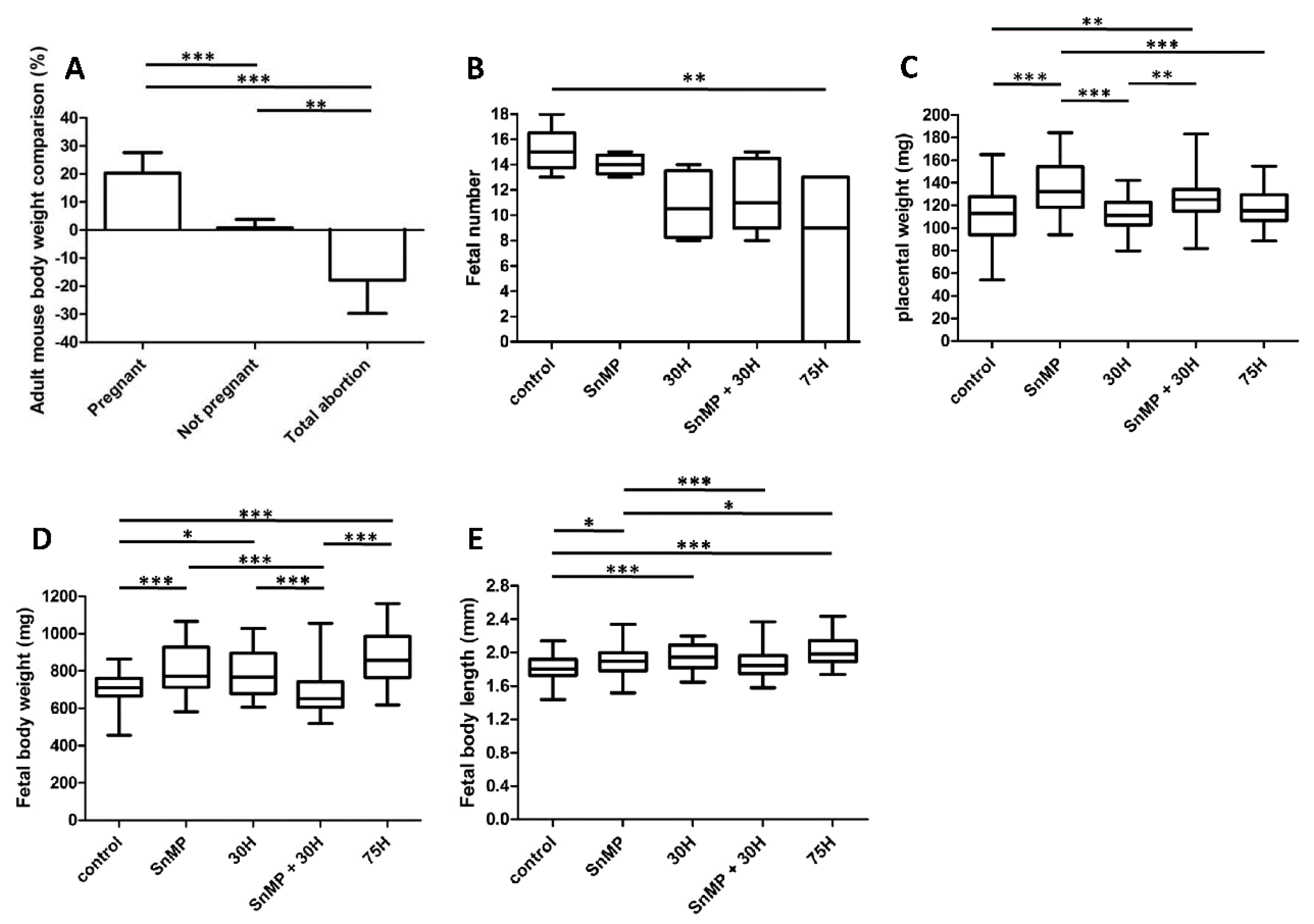
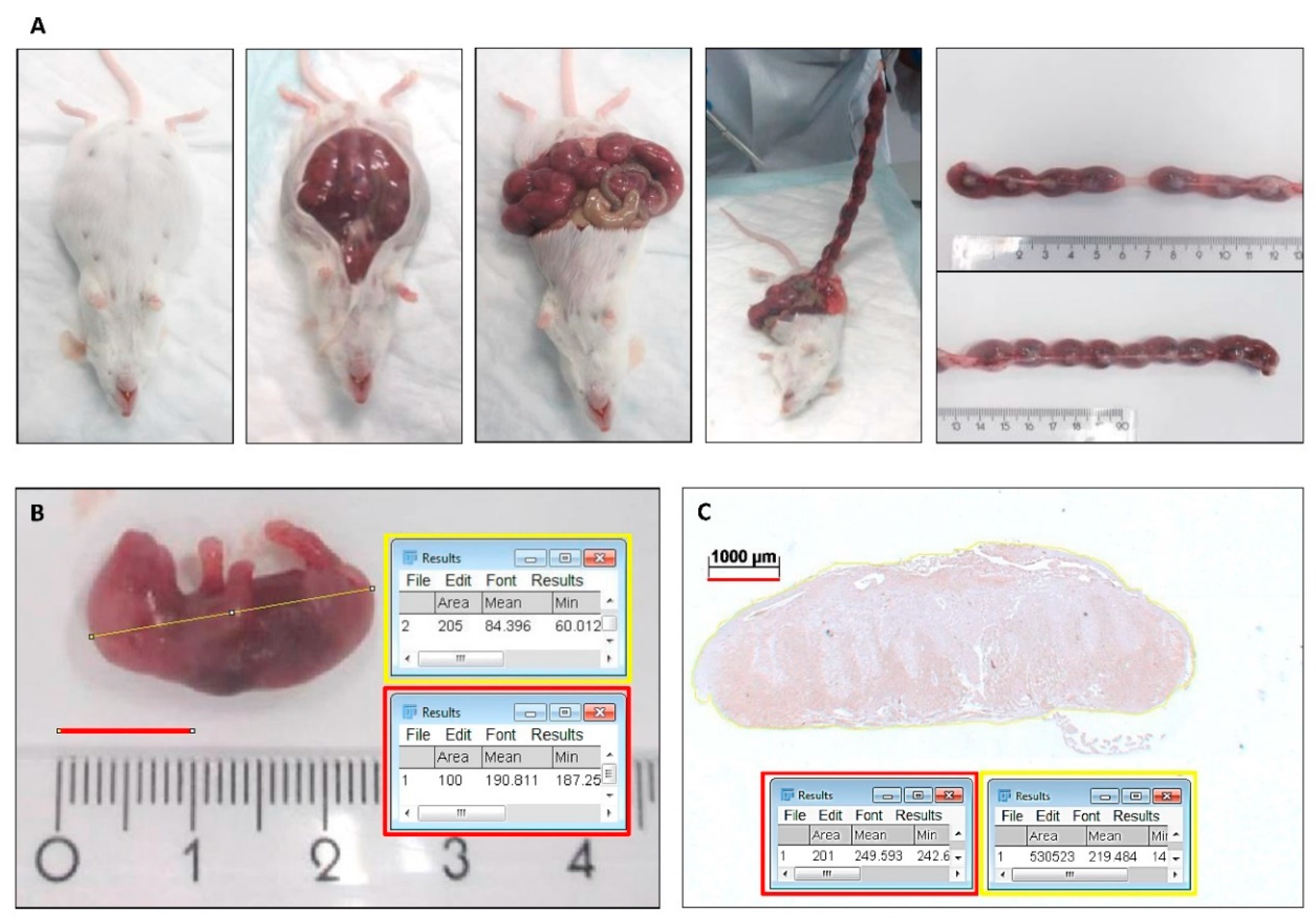
2.1. HO-Activity Protects against Heme-Induced Abortion
2.2. Placental Weight Increase after HO-Activity Inhibition
2.3. Fetal Body Size Increase after Inhibition of HO-Activity and Heme Administration
2.4. Palatal Fusion despite Inhibition of HO-Activity and Heme Administration
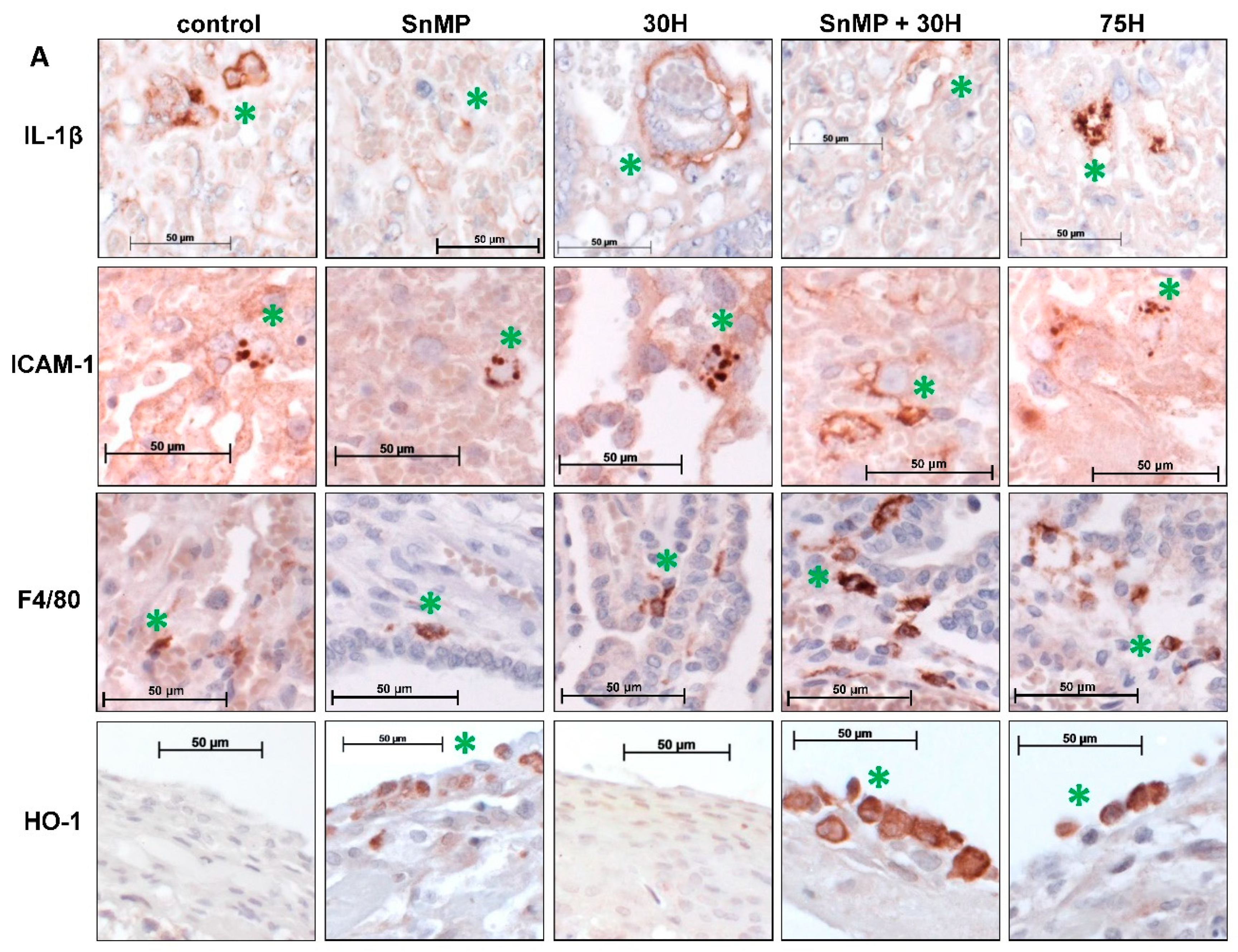
2.5. Inhibition of HO-Activity Reduces IL-1β Expression in Placental Blood Vessels
2.6. Heme-Induced ICAM-1 Expression in Placental Blood Vessels Is Counteracted by HO-Activity
2.7. Placental Macrophage Recruitment Increases after Heme Administration and Decreased HO-Activity
2.8. Placental HO-1 Expression Increases after Heme Administration
3. Discussion
4. Materials and Methods
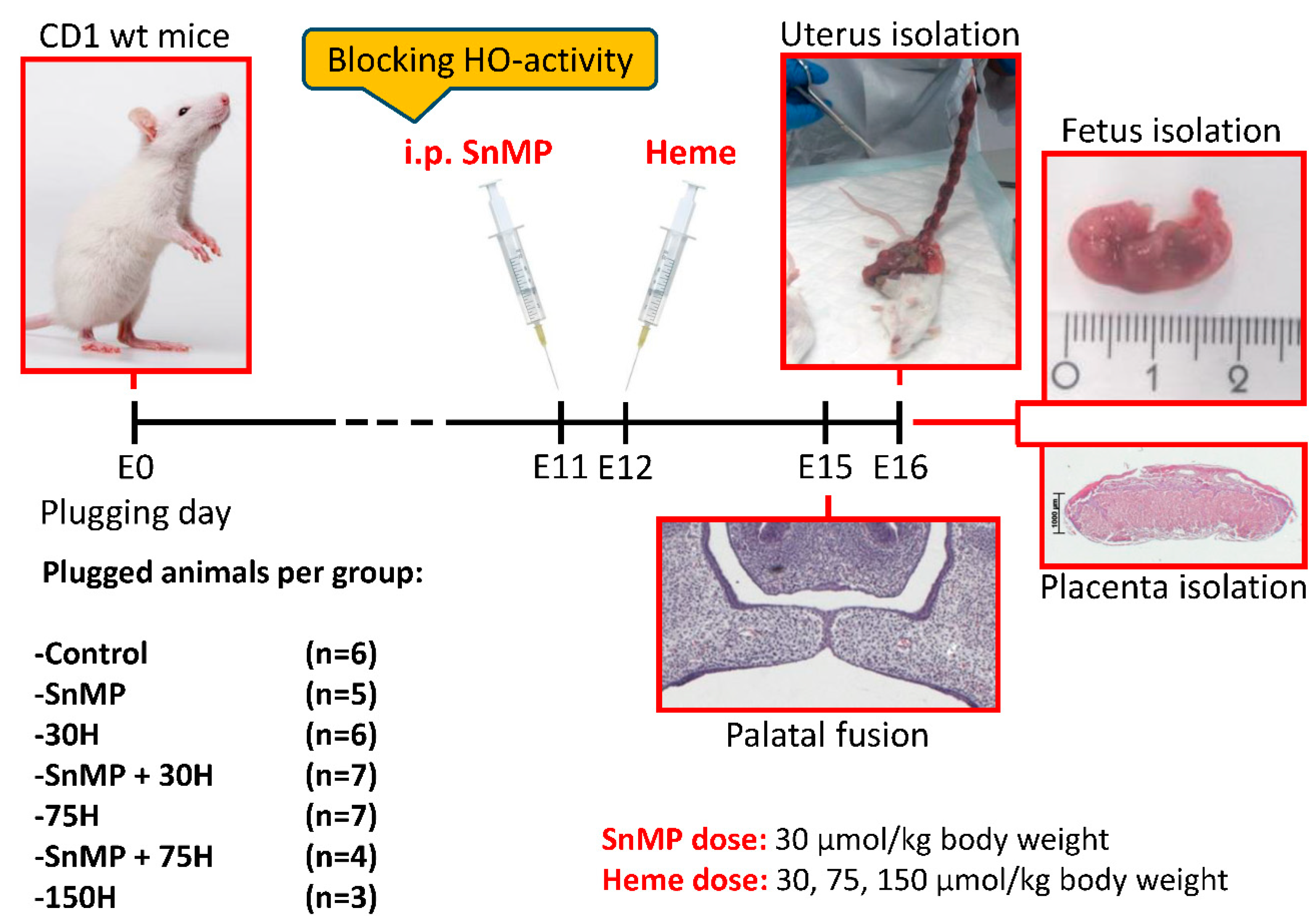
4.1. Mice Selection, Mating, Housing, Ethical Permission
4.2. Heme and/or SnMP Administration, Sample Size, Animal Welfare Monitoring
4.3. Adult Mouse Body Weight Comparison
4.4. Fetal Loss Rate and Fetal Number Calculation
4.5. Placental Weight and Fetal Body Weight and Length
4.6. Paraffin Embedding and Section Cutting of Head and Placenta Samples
4.7. Hematoxylin-Eosin Staining of Palatal Sections and Immunohistochemical Stainings of Placental Sections
4.8. Quantification of IL-1β, ICAM-1, F4/80 and HO-1 Immunoreactivity in Placental Sections
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mossey, P.A.; Modell, B. Epidemiology of oral clefts 2012: An international perspective. Front. Oral Biol. 2012, 16, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kuijpers-Jagtman, A.M. Cleft Lip and Palate: Role of the Orthodontist in the Inderdisciplinary Management Team. In Integrated Clinical Orthodontics; Krishnan, V., Davidovitch, Z., Eds.; Wiley-Blackwell: Chichester, UK, 2012; pp. 153–167. [Google Scholar]
- Nakajima, A.; Shuler, F.S.; Gulka, A.O.D.; Hanai, J.I. TGF-beta Signaling and the Epithelial-Mesenchymal Transition during Palatal Fusion. Int. J. Mol. Sci. 2018, 19, 3638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, Y.; Yeo, J.Y.; Chytil, A.; Han, J.; Bringas, P., Jr.; Nakajima, A.; Shuler, C.F.; Moses, H.L.; Chai, Y. Conditional inactivation of Tgfbr2 in cranial neural crest causes cleft palate and calvaria defects. Development 2003, 130, 5269–5280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, S.E.; Meyer, R.E.; Strauss, R.P.; Aylsworth, A.S. Classification, epidemiology, and genetics of orofacial clefts. Clin. Plast. Surg. 2014, 41, 149–163. [Google Scholar] [CrossRef]
- Brocardo, P.S.; Gil-Mohapel, J.; Christie, B.R. The role of oxidative stress in fetal alcohol spectrum disorders. Brain Res. Rev. 2011, 67, 209–225. [Google Scholar] [CrossRef]
- Spilson, S.V.; Kim, H.J.; Chung, K.C. Association between maternal diabetes mellitus and newborn oral cleft. Ann. Plast. Surg. 2001, 47, 477–481. [Google Scholar] [CrossRef]
- Brien, M.E.; Baker, B.; Duval, C.; Gaudreault, V.; Jones, R.L.; Girard, S. Alarmins at the maternal-fetal interface: Involvement of inflammation in placental dysfunction and pregnancy complications (1). Can. J. Physiol. Pharm. 2019, 97, 206–212. [Google Scholar] [CrossRef]
- Seki, H.; Kuromaki, K.; Takeda, S.; Kinoshita, K. Persistent subchorionic hematoma with clinical symptoms until delivery. Int. J. Gynecol. Obs. 1998, 63, 123–128. [Google Scholar] [CrossRef]
- Johns, J.; Hyett, J.; Jauniaux, E. Obstetric outcome after threatened miscarriage with and without a hematoma on ultrasound. Obs. Gynecol. 2003, 102, 483–487. [Google Scholar]
- Wagener, F.A.; Eggert, A.; Boerman, O.C.; Oyen, W.J.; Verhofstad, A.; Abraham, N.G.; Adema, G.; van Kooyk, Y.; de Witte, T.; Figdor, C.G. Heme is a potent inducer of inflammation in mice and is counteracted by heme oxygenase. Blood 2001, 98, 1802–1811. [Google Scholar] [CrossRef]
- Kassa, T.; Jana, S.; Meng, F.; Alayash, A.I. Differential heme release from various hemoglobin redox states and the upregulation of cellular heme oxygenase-1. FEBS. Open Bio 2016, 6, 876–884. [Google Scholar] [CrossRef]
- Vendrame, F.; Olops, L.; Saad, S.T.O.; Costa, F.F.; Fertrin, K.Y. Differences in heme and hemopexin content in lipoproteins from patients with sickle cell disease. J. Clin. Lipidol. 2018, 12, 1532–1538. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, V.A.; Padmanaban, G. Insights on Heme Synthesis in the Malaria Parasite. Trends Parasitol. 2017, 33, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Ahmad, Z.; Baid, N.; Kumar, A. Host heme oxygenase-1: Friend or foe in tackling pathogens? IUBMB Life 2018, 70, 869–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolisetty, S.; Zarjou, A.; Agarwal, A. Heme Oxygenase 1 as a Therapeutic Target in Acute Kidney Injury. Am. J. Kidney Dis. 2017, 69, 531–545. [Google Scholar] [CrossRef] [Green Version]
- Wagener, F.A.; Dankers, A.C.; van Summeren, F.; Scharstuhl, A.; van den Heuvel, J.J.; Koenderink, J.B.; Pennings, S.W.; Russel, F.G.; Masereeuw, R. Heme Oxygenase-1 and breast cancer resistance protein protect against heme-induced toxicity. Curr. Pharm. Des. 2013, 19, 2698–2707. [Google Scholar] [CrossRef]
- Balla, G.; Vercellotti, G.M.; Muller-Eberhard, U.; Eaton, J.; Jacob, H.S. Exposure of endothelial cells to free heme potentiates damage mediated by granulocytes and toxic oxygen species. Lab. Investig. 1991, 64, 648–655. [Google Scholar]
- Balla, J.; Jacob, H.S.; Balla, G.; Nath, K.; Eaton, J.W.; Vercellotti, G.M. Endothelial-cell heme uptake from heme proteins: Induction of sensitization and desensitization to oxidant damage. Proc. Natl. Acad. Sci. USA 1993, 90, 9285–9289. [Google Scholar] [CrossRef] [Green Version]
- Mueller, C.F.; Laude, K.; McNally, J.S.; Harrison, D.G. ATVB in focus: Redox mechanisms in blood vessels. Arter. Thromb. Vasc. Biol. 2005, 25, 274–278. [Google Scholar] [CrossRef] [Green Version]
- Nagy, E.; Eaton, J.W.; Jeney, V.; Soares, M.P.; Varga, Z.; Galajda, Z.; Szentmiklosi, J.; Mehes, G.; Csonka, T.; Smith, A.; et al. Red cells, hemoglobin, heme, iron, and atherogenesis. Arter. Thromb. Vasc. Biol. 2010, 30, 1347–1353. [Google Scholar] [CrossRef]
- Silva, G.; Jeney, V.; Chora, A.; Larsen, R.; Balla, J.; Soares, M.P. Oxidized hemoglobin is an endogenous proinflammatory agonist that targets vascular endothelial cells. J. Biol. Chem. 2009, 284, 29582–29595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posta, N.; Csosz, E.; Oros, M.; Petho, D.; Potor, L.; Kallo, G.; Hendrik, Z.; Sikura, K.E.; Mehes, G.; Toth, C.; et al. Hemoglobin oxidation generates globin-derived peptides in atherosclerotic lesions and intraventricular hemorrhage of the brain, provoking endothelial dysfunction. Lab. Investig. 2020, 100, 986–1002. [Google Scholar] [CrossRef] [PubMed]
- Wagener, F.A.; Scharstuhl, A.; Tyrrell, R.M.; Von den Hoff, J.W.; Jozkowicz, A.; Dulak, J.; Russel, F.G.; Kuijpers-Jagtman, A.M. The heme-heme oxygenase system in wound healing; implications for scar formation. Curr. Drug Targets 2010, 11, 1571–1585. [Google Scholar] [CrossRef] [PubMed]
- Zenclussen, M.L.; Casalis, P.A.; El-Mousleh, T.; Rebelo, S.; Langwisch, S.; Linzke, N.; Volk, H.D.; Fest, S.; Soares, M.P.; Zenclussen, A.C. Haem oxygenase-1 dictates intrauterine fetal survival in mice via carbon monoxide. J. Pathol. 2011, 225, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Wagener, F.A.; Feldman, E.; de Witte, T.; Abraham, N.G. Heme induces the expression of adhesion molecules ICAM-1, VCAM-1, and E selectin in vascular endothelial cells. Proc. Soc. Exp. Biol. Med. 1997, 216, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Wagener, F.A.; van Beurden, H.E.; von den Hoff, J.W.; Adema, G.J.; Figdor, C.G. The heme-heme oxygenase system: A molecular switch in wound healing. Blood 2003, 102, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Tuuli, M.G.; Norman, S.M.; Odibo, A.O.; Macones, G.A.; Cahill, A.G. Perinatal outcomes in women with subchorionic hematoma: A systematic review and meta-analysis. Obstet. Gynecol. 2011, 117, 1205–1212. [Google Scholar] [CrossRef]
- Xiang, L.; Wei, Z.; Cao, Y. Symptoms of an intrauterine hematoma associated with pregnancy complications: A systematic review. PLoS ONE 2014, 9, e111676. [Google Scholar] [CrossRef] [Green Version]
- Nyberg, D.A.; Mack, L.A.; Benedetti, T.J.; Cyr, D.R.; Schuman, W.P. Placental abruption and placental hemorrhage: Correlation of sonographic findings with fetal outcome. Radiology 1987, 164, 357–361. [Google Scholar] [CrossRef]
- Ascenzi, P.; Bocedi, A.; Visca, P.; Altruda, F.; Tolosano, E.; Beringhelli, T.; Fasano, M. Hemoglobin and heme scavenging. IUBMB Life 2005, 57, 749–759. [Google Scholar] [CrossRef]
- Wagener, F.A.; Volk, H.D.; Willis, D.; Abraham, N.G.; Soares, M.P.; Adema, G.J.; Figdor, C.G. Different faces of the heme-heme oxygenase system in inflammation. Pharm. Rev. 2003, 55, 551–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorlach, A.; Dimova, E.Y.; Petry, A.; Martinez-Ruiz, A.; Hernansanz-Agustin, P.; Rolo, A.P.; Palmeira, C.M.; Kietzmann, T. Reactive oxygen species, nutrition, hypoxia and diseases: Problems solved? Redox. Biol. 2015, 6, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Zenclussen, A.C.; Lim, E.; Knoeller, S.; Knackstedt, M.; Hertwig, K.; Hagen, E.; Klapp, B.F.; Arck, P.C. Heme oxygenases in pregnancy II: HO-2 is downregulated in human pathologic pregnancies. Am. J. Reprod. Immunol. 2003, 50, 66–76. [Google Scholar] [CrossRef]
- Zenclussen, M.L.; Linzke, N.; Schumacher, A.; Fest, S.; Meyer, N.; Casalis, P.A.; Zenclussen, A.C. Heme oxygenase-1 is critically involved in placentation, spiral artery remodeling, and blood pressure regulation during murine pregnancy. Front. Pharm. 2014, 5, 291. [Google Scholar] [CrossRef] [Green Version]
- Sollwedel, A.; Bertoja, A.Z.; Zenclussen, M.L.; Gerlof, K.; Lisewski, U.; Wafula, P.; Sawitzki, B.; Woiciechowsky, C.; Volk, H.D.; Zenclussen, A.C. Protection from abortion by heme oxygenase-1 up-regulation is associated with increased levels of Bag-1 and neuropilin-1 at the fetal-maternal interface. J. Immunol. 2005, 175, 4875–4885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miya, M.; Okamoto, A.; Nikaido, T.; Tachimoto-Kawaguchi, R.; Tanaka, T. Immunohistochemical localization of heme oxygenase-1 and bilirubin/biopyrrin of heme metabolites as antioxidants in human placenta with preeclampsia. Hypertens Pregnancy 2019, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Grochot-Przeczek, A.; Dulak, J.; Jozkowicz, A. Haem oxygenase-1: Non-canonical roles in physiology and pathology. Clin. Sci. (Lond) 2012, 122, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Ewing, J.F.; Maines, M.D. Regulation and expression of heme oxygenase enzymes in aged-rat brain: Age related depression in HO-1 and HO-2 expression and altered stress-response. J. Neural. Transm. (Vienna) 2006, 113, 439–454. [Google Scholar] [CrossRef]
- Suttorp, C.M.; Cremers, N.A.; van Rheden, R.; Regan, R.F.; Helmich, P.; van Kempen, S.; Kuijpers-Jagtman, A.M.; Wagener, F. Chemokine Signaling during Midline Epithelial Seam Disintegration Facilitates Palatal Fusion. Front. Cell Dev. Biol. 2017, 5, 94. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Fu, W.; Yao, J.; Ji, Z.; Wang, Y.; Zhou, Z.; Yan, J.; Li, W. Heme induces IL-1beta secretion through activating NLRP3 in kidney inflammation. Cell Biochem. Biophys. 2014, 69, 495–502. [Google Scholar] [CrossRef]
- Benallaoua, M.; Francois, M.; Batteux, F.; Thelier, N.; Shyy, J.Y.J.; Fitting, C.; Tsagris, L.; Boczkowski, J.; Savouret, J.F.; Corvol, M.T.; et al. Pharmacologic induction of heme oxygenase 1 reduces acute inflammatory arthritis in mice. Arthritis Rheum. 2007, 56, 2585–2594. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wong, R.J.; Kalish, F.S.; Nayak, N.R.; Stevenson, D.K. Effect of heme oxygenase-1 deficiency on placental development. Placenta 2009, 30, 861–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mossey, P.A.; Little, J.; Munger, R.G.; Dixon, M.J.; Shaw, W.C. Cleft lip and palate. Lancet 2009, 374, 1773–1785. [Google Scholar] [CrossRef]
- Belcher, J.D.; Mahaseth, H.; Welch, T.E.; Otterbein, L.E.; Hebbel, R.P.; Vercellotti, G.M. Heme oxygenase-1 is a modulator of inflammation and vaso-occlusion in transgenic sickle mice. J. Clin. Investig. 2006, 116, 808–816. [Google Scholar] [CrossRef] [Green Version]
- Belcher, J.D.; Chen, C.; Nguyen, J.; Milbauer, L.; Abdulla, F.; Alayash, A.I.; Smith, A.; Nath, K.A.; Hebbel, R.P.; Vercellotti, G.M. Heme triggers TLR4 signaling leading to endothelial cell activation and vaso-occlusion in murine sickle cell disease. Blood 2014, 123, 377–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, A.; Sarsam, S.D. The Impact of Incidental Ultrasound Finding of Subchorionic and Retroplacental Hematoma in Early Pregnancy. J. Obstet. Gynaecol. India 2019, 69, 43–49. [Google Scholar] [CrossRef]
- Ji, W.; Li, W.; Mei, S.; He, P. Intrauterine hematomas in the second and third trimesters associated with adverse pregnancy outcomes: A retrospective study. J. Matern. Fetal. Neonatal. Med. 2017, 30, 2151–2155. [Google Scholar] [CrossRef]
- Ott, J.; Pecnik, P.; Promberger, R.; Pils, S.; Binder, J.; Chalubinski, K.M. Intra- versus retroplacental hematomas: A retrospective case-control study on pregnancy outcomes. BMC Pregnancy Childbirth 2017, 17, 366. [Google Scholar] [CrossRef] [Green Version]
- MacMullen, N.J.; Dulski, L.A.; Meagher, B. Red alert: Perinatal hemorrhage. MCN Am. J. Matern. Child Nurs. 2005, 30, 46–51. [Google Scholar]
- Schumacher, A.; Wafula, P.O.; Teles, A.; El-Mousleh, T.; Linzke, N.; Zenclussen, M.L.; Langwisch, S.; Heinze, K.; Wollenberg, I.; Casalis, P.A.; et al. Blockage of heme oxygenase-1 abrogates the protective effect of regulatory T cells on murine pregnancy and promotes the maturation of dendritic cells. PLoS ONE 2012, 7, e42301. [Google Scholar] [CrossRef] [Green Version]
- Zenclussen, A.C.; Sollwedel, A.; Bertoja, A.Z.; Gerlof, K.; Zenclussen, M.L.; Woiciechowsky, C.; Volk, H.D. Heme oxygenase as a therapeutic target in immunological pregnancy complications. Int. Immunopharmacol. 2005, 5, 41–51. [Google Scholar] [CrossRef] [PubMed]
- El-Mousleh, T.; Casalis, P.A.; Wollenberg, I.; Zenclussen, M.L.; Volk, H.D.; Langwisch, S.; Jensen, F.; Zenclussen, A.C. Exploring the potential of low doses carbon monoxide as therapy in pregnancy complications. Med. Gas. Res. 2012, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Puri, N.; Sodhi, K.; Bellner, L.; Abraham, N.G.; Kappas, A. Apo A1 Mimetic Rescues the Diabetic Phenotype of HO-2 Knockout Mice via an Increase in HO-1 Adiponectin and LKBI Signaling Pathway. Int. J. Hypertens. 2012, 2012, 628147. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dore, S. Heme oxygenase 2 deficiency increases brain swelling and inflammation after intracerebral hemorrhage. Neuroscience 2008, 155, 1133–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Wei, X.; Cui, X.; Kobayashi, T.; Li, W. Effects of heme oxygenase 1 on brain edema and neurologic outcome after cardiopulmonary resuscitation in rats. Anesthesiology 2008, 109, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Wagner, K.R.; Hua, Y.; de Courten-Myers, G.M.; Broderick, J.P.; Nishimura, R.N.; Lu, S.Y.; Dwyer, B.E. Tin-mesoporphyrin, a potent heme oxygenase inhibitor, for treatment of intracerebral hemorrhage: In vivo and in vitro studies. Cell. Mol. Biol. (Noisy-le-grand) 2000, 46, 597–608. [Google Scholar]
- Daguindau, E.; Gautier, T.; Chague, C.; Pais de Barros, J.P.; Deckert, V.; Lagrost, L.; Saas, P. Is It Time to Reconsider the Lipopolysaccharide Paradigm in Acute Graft-Versus-Host Disease? Front. Immunol. 2017, 8, 952. [Google Scholar] [CrossRef] [Green Version]
- Toyama, R.P.; Xikota, J.C.; Schwarzbold, M.L.; Frode, T.S.; Buss Zda, S.; Nunes, J.C.; Funchal, G.D.; Nunes, F.C.; Walz, R.; Pires, M.M. Dose-dependent sickness behavior, abortion and inflammation induced by systemic LPS injection in pregnant mice. J. Matern. Fetal. Neonatal. Med. 2015, 28, 426–430. [Google Scholar] [CrossRef]
- Er, A. Azithromycin prevents pregnancy loss: Reducing the level of tumor necrosis factor-alpha and raising the level of interleukin-10 in rats. Mediat. Inflamm. 2013, 2013, 928137. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, Y.H.; Fu, L.; Yu, Z.; Xia, M.Z.; Hu, X.G.; Wang, H.; Xu, D.X. Vitamin D3 pretreatment protects against lipopolysaccharide-induced early embryo loss through its anti-inflammatory effects. Am. J. Reprod. Immunol. 2017, 77. [Google Scholar] [CrossRef]
- Mahdavi Siuki, M.; Fazel Nasab, N.; Barati, E.; Nezafat Firizi, M.; Jalilvand, T.; Namdar Ahmadabad, H. The protective effect of berberine against lipopolysaccharide-induced abortion by modulation of inflammatory/immune responses. Immunopharmacol. Immunotoxicol. 2018, 40, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Di Simone, N.; Di Nicuolo, F.; Marana, R.; Castellani, R.; Ria, F.; Veglia, M.; Scambia, G.; Surbek, D.; Barnea, E.; Mueller, M. Synthetic PreImplantation Factor (PIF) prevents fetal loss by modulating LPS induced inflammatory response. PLoS ONE 2017, 12, e0180642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Kong, N.; Zhou, C.; Mungun, D.; Iyan, Z.; Guo, Y.; Yang, Z. Effect of Remote Ischemic Preconditioning on Perioperative Cardiac Events in Patients Undergoing Elective Percutaneous Coronary Intervention: A Meta-Analysis of 16 Randomized Trials. Cardiol. Res. Pract. 2017, 2017, 6907167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luna, R.L.; Nunes, A.K.; Oliveira, A.G.; Araujo, S.M.; Lemos, A.J.; Rocha, S.W.; Croy, B.A.; Peixoto, C.A. Sildenafil (Viagra(R)) blocks inflammatory injury in LPS-induced mouse abortion: A potential prophylactic treatment against acute pregnancy loss? Placenta 2015, 36, 1122–1129. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.Y.; Zhao, L.; Wang, H.; Xu, D.X. Lipopolysaccharide (LPS) up-regulates the expression of haem oxygenase-1 in mouse placenta. Placenta 2007, 28, 951–957. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, H.; Sun, X.; Ren, L. The protective role of vitamin D3 in a murine model of asthma via the suppression of TGF-beta/Smad signaling and activation of the Nrf2/HO-1 pathway. Mol. Med. Rep. 2016, 14, 2389–2396. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Fan, C.; Yan, X.; Lu, X.; Jiang, H.; Di, S.; Ma, Z.; Feng, Y.; Zhang, Z.; Feng, P.; et al. Berberine ameliorates lipopolysaccharide-induced acute lung injury via the PERK-mediated Nrf2/HO-1 signaling axis. Phytother. Res. 2019, 33, 130–148. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Zhang, D.; Wang, L.; Sreeharsha, N.; Ning, Y. Curcumin synergistically potentiates the protective effect of sitagliptin against chronic deltamethrin nephrotoxicity in rats: Impact on pro-inflammatory cytokines and Nrf2/Ho-1 pathway. J. Biochem. Mol. Toxicol. 2019, e22386. [Google Scholar] [CrossRef]
- Jeong, J.H.; Kim, H.G.; Choi, O.H. Sildenafil Inhibits Advanced Glycation End Products-induced sFlt-1 Release Through Upregulation of Heme Oxygenase-1. J. Menopausal Med. 2014, 20, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Chen, S.; Hu, Z.; Xiao, F.; Lu, S.; Ma, X.; Luan, Z. Effect of unfractionated heparin on the expression of heme oxygenase-1 in intestinal mucosa of mice with sepsis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2016, 28, 423–426. [Google Scholar]
- Pontillo, A.; Girardelli, M.; Agostinis, C.; Masat, E.; Bulla, R.; Crovella, S. Bacterial LPS differently modulates inflammasome gene expression and IL-1beta secretion in trophoblast cells, decidual stromal cells, and decidual endothelial cells. Reprod. Sci. 2013, 20, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Duval, C.; Brien, M.E.; Gaudreault, V.; Boufaied, I.; Baker, B.; Jones, R.L.; Girard, S. Differential effect of LPS and IL-1beta in term placental explants. Placenta 2019, 75, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Wei, J.; Snowise, S.; DeSousa, J.; Stone, P.; Viall, C.; Chen, Q.; Chamley, L. Trophoblast debris extruded from preeclamptic placentae activates endothelial cells: A mechanism by which the placenta communicates with the maternal endothelium. Placenta 2014, 35, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Girard, S.; Heazell, A.E.; Derricott, H.; Allan, S.M.; Sibley, C.P.; Abrahams, V.M.; Jones, R.L. Circulating cytokines and alarmins associated with placental inflammation in high-risk pregnancies. Am. J. Reprod. Immunol. 2014, 72, 422–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdei, J.; Toth, A.; Balogh, E.; Nyakundi, B.B.; Banyai, E.; Ryffel, B.; Paragh, G.; Cordero, M.D.; Jeney, V. Induction of NLRP3 Inflammasome Activation by Heme in Human Endothelial Cells. Oxid. Med. Cell. Longev. 2018, 2018, 4310816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, J.K.; Mitchell, M.D. Intrauterine infection and the effects of inflammatory mediators on prostaglandin production by myometrial cells from pregnant women. Am. J. Obstet. Gynecol. 1996, 174, 682–686. [Google Scholar] [CrossRef]
- Oger, S.; Mehats, C.; Dallot, E.; Ferre, F.; Leroy, M.J. Interleukin-1beta induces phosphodiesterase 4B2 expression in human myometrial cells through a prostaglandin E2- and cyclic adenosine 3′,5′-monophosphate-dependent pathway. J. Clin. Endocrinol. Metab. 2002, 87, 5524–5531. [Google Scholar] [CrossRef] [Green Version]
- Blois, S.; Tometten, M.; Kandil, J.; Hagen, E.; Klapp, B.F.; Margni, R.A.; Arck, P.C. Intercellular adhesion molecule-1/LFA-1 cross talk is a proximate mediator capable of disrupting immune integration and tolerance mechanism at the feto-maternal interface in murine pregnancies. J. Immunol. 2005, 174, 1820–1829. [Google Scholar] [CrossRef] [Green Version]
- Takeshita, T.; Satomi, M.; Akira, S.; Nakagawa, Y.; Takahashi, H.; Araki, T. Preventive effect of monoclonal antibodies to intercellular adhesion molecule-1 and leukocyte function-associate antigen-1 on murine spontaneous fetal resorption. Am. J. Reprod. Immunol. 2000, 43, 180–185. [Google Scholar] [CrossRef]
- Korkmazer, E.; Nizam, R.; Arslan, E.; Akkurt, O. Relationship between intercellular adhesion molecule-1 and morbidly adherent placenta. J. Perinat. Med. 2018, 47, 45–49. [Google Scholar] [CrossRef]
- Szarka, A.; Rigo, J., Jr.; Lazar, L.; Beko, G.; Molvarec, A. Circulating cytokines, chemokines and adhesion molecules in normal pregnancy and preeclampsia determined by multiplex suspension array. BMC Immunol. 2010, 11, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Ryu, H.M.; Yang, J.H.; Kim, M.Y.; Ahn, H.K.; Lim, H.J.; Shin, J.S.; Woo, H.J.; Park, S.Y.; Kim, Y.M.; et al. Maternal serum levels of VCAM-1, ICAM-1 and E-selectin in preeclampsia. J. Korean. Med. Sci. 2004, 19, 688–692. [Google Scholar] [CrossRef] [Green Version]
- Docheva, N.; Romero, R.; Chaemsaithong, P.; Tarca, A.L.; Bhatti, G.; Pacora, P.; Panaitescu, B.; Chaiyasit, N.; Chaiworapongsa, T.; Maymon, E.; et al. The profiles of soluble adhesion molecules in the “great obstetrical syndromes”. J. Matern. Fetal. Neonatal. Med. 2019, 32, 2113–2136. [Google Scholar] [CrossRef] [PubMed]
- Abe, E.; Matsubara, K.; Oka, K.; Kusanagi, Y.; Ito, M. Cytokine regulation of intercellular adhesion molecule-1 expression on trophoblasts in preeclampsia. Gynecol. Obstet. Investig. 2008, 66, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Goksu, E.A.Y.; Nazli, M.; Elis, Y.S. Significance of platelet endothelial cell adhesion molecule-1 (PECAM-1) and intercellular adhesion molecule-1 (ICAM-1) expressions in preeclamptic placentae. Endocrine 2012, 42, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Salafia, C.M.; Weigl, C.; Silberman, L. The prevalence and distribution of acute placental inflammation in uncomplicated term pregnancies. Obstet. Gynecol. 1989, 73, 383–389. [Google Scholar] [CrossRef]
- Shynlova, O.; Nedd-Roderique, T.; Li, Y.; Dorogin, A.; Lye, S.J. Myometrial immune cells contribute to term parturition, preterm labour and post-partum involution in mice. J. Cell. Mol. Med. 2013, 17, 90–102. [Google Scholar] [CrossRef]
- Leong, A.S.; Norman, J.E.; Smith, R. Vascular and myometrial changes in the human uterus at term. Reprod. Sci. 2008, 15, 59–65. [Google Scholar] [CrossRef]
- Ledingham, M.A.; Thomson, A.J.; Jordan, F.; Young, A.; Crawford, M.; Norman, J.E. Cell adhesion molecule expression in the cervix and myometrium during pregnancy and parturition. Obstet. Gynecol. 2001, 97, 235–242. [Google Scholar]
- Hamilton, S.; Oomomian, Y.; Stephen, G.; Shynlova, O.; Tower, C.L.; Garrod, A.; Lye, S.J.; Jones, R.L. Macrophages infiltrate the human and rat decidua during term and preterm labor: Evidence that decidual inflammation precedes labor. Biol. Reprod. 2012, 86, 39. [Google Scholar] [CrossRef]
- Bhartiya, D.; James, K. Very small embryonic-like stem cells (VSELs) in adult mouse uterine perimetrium and myometrium. J. Ovarian. Res. 2017, 10, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsur, A.; Kalish, F.; Burgess, J.; Nayak, N.R.; Zhao, H.; Casey, K.M.; Druzin, M.L.; Wong, R.J.; Stevenson, D.K. Pravastatin improves fetal survival in mice with a partial deficiency of heme oxygenase-1. Placenta 2019, 75, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sussan, T.E.; Sudini, K.; Talbot, C.C.J.; Wang, X.; Wills-Karp, M.; Burd, I.; Biswal, S. Nrf2 regulates gene-environment interactions in an animal model of intrauterine inflammation: Implications for preterm birth and prematurity. Sci. Rep. 2017, 7, 40194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, M.J.; Moller, H.J.; Moestrup, S.K. Hemoglobin and heme scavenger receptors. Antioxid. Redox Signal. 2010, 12, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Frimat, M.; Boudhabhay, I.; Roumenina, L.T. Hemolysis Derived Products Toxicity and Endothelium: Model of the Second Hit. Toxins (Basel) 2019, 11, 660. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.; Knapp, S. Heme and hemolysis in innate immunity: Adding insult to injury. Curr. Opin. Immunol. 2017, 50, 14–20. [Google Scholar] [CrossRef]
- Wang, G.; Manaenko, A.; Shao, A.; Ou, Y.; Yang, P.; Budbazar, E.; Nowrangi, D.; Zhang, J.H.; Tang, J. Low-density lipoprotein receptor-related protein-1 facilitates heme scavenging after intracerebral hemorrhage in mice. J. Cereb. Blood Flow Metab. 2017, 37, 1299–1310. [Google Scholar] [CrossRef]
- Hvidberg, V.; Maniecki, M.B.; Jacobsen, C.; Hojrup, P.; Moller, H.J.; Moestrup, S.K. Identification of the receptor scavenging hemopexin-heme complexes. Blood 2005, 106, 2572–2579. [Google Scholar] [CrossRef]
- Lin, S.; Yin, Q.; Zhong, Q.; Lv, F.L.; Zhou, Y.; Li, J.Q.; Wang, J.Z.; Su, B.Y.; Yang, Q.W. Heme activates TLR4-mediated inflammatory injury via MyD88/TRIF signaling pathway in intracerebral hemorrhage. J. Neuroinflammation 2012, 9, 46. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Athayde, N.; Trudinger, B. Endothelial cell expression of adhesion molecules is induced by fetal plasma from pregnancies with umbilical placental vascular disease. Br. J. Obstet. Gynaecol. 2002, 109, 770–777. [Google Scholar] [CrossRef]
- Juliano, P.B.; Blotta, M.H.; Altemani, A.M. ICAM-1 is overexpressed by villous trophoblasts in placentitis. Placenta 2006, 27, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Belcher, J.D.; Beckman, J.D.; Balla, G.; Balla, J.; Vercellotti, G. Heme degradation and vascular injury. Antioxid. Redox Signal. 2010, 12, 233–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behringer, R.; Gertsenstein, M.; Nagy, K.V.; Nagy, A. Selecting Female Mice in Estrus and Checking Plugs. Cold Spring Harb. Protoc. 2016. [Google Scholar] [CrossRef] [PubMed]
- Dudas, M.; Li, W.Y.; Kim, J.; Yang, A.; Kaartinen, V. Palatal fusion—where do the midline cells go? A review on cleft palate, a major human birth defect. Acta Histochem. 2007, 109, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lutton, J.D.; Jiang, S.; Drummond, G.S.; Abraham, N.G.; Kappas, A. Comparative pharmacology of zinc mesoporphyrin and tin mesoporphyrin: Toxic actions of zinc mesoporphyrin on hematopoiesis and progenitor cell mobilization. Pharmacology 1999, 58, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, D.K.; Rodgers, P.A.; Vreman, H.J. The use of metalloporphyrins for the chemoprevention of neonatal jaundice. Am. J. Dis. Child 1989, 143, 353–356. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Lin, H.H.; Faunce, D.E.; Stacey, M.; Terajewicz, A.; Nakamura, T.; Zhang-Hoover, J.; Kerley, M.; Mucenski, M.L.; Gordon, S.; Stein-Streilein, J. The macrophage F4/80 receptor is required for the induction of antigen-specific efferent regulatory T cells in peripheral tolerance. J. Exp. Med. 2005, 201, 1615–1625. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.D.; Xie, R.; Klein-Nulend, J.; van Rheden, R.E.; Bronckers, A.L.; Kuijpers-Jagtman, A.M.; Von den Hoff, J.W.; Maltha, J.C. Orthodontic force stimulates eNOS and iNOS in rat osteocytes. J. Dent. Res. 2009, 88, 255–260. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| First Antibody | Specificity | Concentration (µg/mL) | Source |
|---|---|---|---|
| sc-1252 | IL-1B | 0.2 | Santa Cruz Biotechnology, Santa Cruz, CA, USA |
| CD54 | ICAM-1 | 0.34 | Proteintech via Sanbio, Uden, The Netherlands |
| A3-1 (ab6640) | F4/80 | 1.0 | Abcam Cambridge Biomedical Campus, Cambridge, UK |
| SPA 895 | HO-1 | 1.0 | Stressgen, Victoria, BC, USA |
| Secondary Antibody | Specificity | Concentration (µg/mL) | Source |
|---|---|---|---|
| 705-065-147 | Donkey anti-goat Biotin | 2.8 | Jackson Immunoresearch West Grove, PA, USA |
| 711-065-147 | Donkey anti-rabbit Biotin | 2.0 | Jackson Immunoresearch West Grove, PA, USA |
| 712-065-153 | Donkey anti-rat Biotin | 4.6 | Jackson Immunoresearch West Grove, PA, USA |
| 711-065-147 | Donkey anti-rabbit Biotin | 2.0 | Jackson Immunoresearch West Grove, PA, USA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suttorp, C.M.; van Rheden, R.E.M.; van Dijk, N.W.M.; Helmich, M.P.A.C.; Kuijpers-Jagtman, A.M.; Wagener, F.A.D.T.G. Heme Oxygenase Protects against Placental Vascular Inflammation and Abortion by the Alarmin Heme in Mice. Int. J. Mol. Sci. 2020, 21, 5385. https://doi.org/10.3390/ijms21155385
Suttorp CM, van Rheden REM, van Dijk NWM, Helmich MPAC, Kuijpers-Jagtman AM, Wagener FADTG. Heme Oxygenase Protects against Placental Vascular Inflammation and Abortion by the Alarmin Heme in Mice. International Journal of Molecular Sciences. 2020; 21(15):5385. https://doi.org/10.3390/ijms21155385
Chicago/Turabian StyleSuttorp, Christiaan M., René E. M. van Rheden, Natasja W. M. van Dijk, Maria P. A. C. Helmich, Anne Marie Kuijpers-Jagtman, and Frank A. D. T. G. Wagener. 2020. "Heme Oxygenase Protects against Placental Vascular Inflammation and Abortion by the Alarmin Heme in Mice" International Journal of Molecular Sciences 21, no. 15: 5385. https://doi.org/10.3390/ijms21155385
APA StyleSuttorp, C. M., van Rheden, R. E. M., van Dijk, N. W. M., Helmich, M. P. A. C., Kuijpers-Jagtman, A. M., & Wagener, F. A. D. T. G. (2020). Heme Oxygenase Protects against Placental Vascular Inflammation and Abortion by the Alarmin Heme in Mice. International Journal of Molecular Sciences, 21(15), 5385. https://doi.org/10.3390/ijms21155385