Mesenchymal Stem Cells in Aplastic Anemia and Myelodysplastic Syndromes: The “Seed and Soil” Crosstalk

Abstract
:1. Introduction
2. The Role of Mesenchymal Stem Cells and Microenvironment in Aplastic Anemia
2.1. Pathogenic Mechanisms of Mesenchymal Stem Cells in AA
2.2. Therapeutic Employment of MSCs in AA
2.2.1. MSC Infusions in AA Patients
2.2.2. MSC and HSC Cotransplantation in AA Patients
2.2.3. Other Studies on Therapeutic Use of MSCs in AA
3. The Role of Mesenchymal Stem Cells in Myelodysplastic Syndromes
3.1. Preclinical Evidence of MSCs’ Role in MDSs
3.1.1. MSCs’ Role in Inducing Clonal Hematopoiesis in MDSs
3.1.2. MSCs’ Role in Facilitating Clonal Hematopoiesis in MDSs
3.2. Therapeutic Strategies in MDSs Involving MSCs
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fracchiolla, N.S.; Fattizzo, B.; Cortelezzi, A. Mesenchymal Stem Cells in Myeloid Malignancies: A Focus on Immune Escaping and Therapeutic Implications. Stem Cells Int. 2017, 2017, 6720594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, J.S.; Choi, Y.; Kim, H.S.; Kim, H.O. Comparison of molecular profiles of human mesenchymal stem cells derived from bone marrow, umbilical cord blood, placenta and adipose tissue. Int. J. Mol. Med. 2016, 37, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef]
- Sacchetti, B.; Funari, A.; Michienzi, S.; Di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Gehron Robey, P.; Riminucci, M.; et al. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Salter, A.B.; Meadows, S.K.; Muramoto, G.G.; Himburg, H.; Doan, P.; Daher, P.; Russell, L.; Chen, B.; Chao, N.J.; Chute, J.P. Endothelial progenitor cell infusion induces hematopoietic stem cell reconstitution in vivo. Blood 2009, 113, 2104–2107. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Saunders, T.L.; Enikolopov, G.; Morrison, S.J. Endothelial and perivascular cells maintain haematopoietic stem cells. Nature 2012, 481, 457–462. [Google Scholar] [CrossRef] [Green Version]
- Winkler, I.G.; Barbier, V.; Nowlan, B.; Jacobsen, R.N.; Forristal, C.E.; Patton, J.T.; Magnani, J.L.; Lévesque, J.-P. Vascular niche E-selectin regulates hematopoietic stem cell dormancy, self renewal and chemoresistance. Nat. Med. 2012, 18, 1651–1657. [Google Scholar] [CrossRef]
- Lévesque, J.P.; Helwani, F.M.; Winkler, I.G. The endosteal ‘osteoblastic’ niche and its role in hematopoietic stem cell homing and mobilization. Leukemia 2010, 24, 1979–1992. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Chatterjee, S.; Basak, P.; Das, P.; Pereira, J.A.; Dutta, R.K.; Chaklader, M.; Chaudhuri, S.; Law, S. The bone marrow stem stromal imbalance—A key feature of disease progression in case of myelodysplastic mouse model. J. Stem Cells 2010, 5, 49–64. [Google Scholar]
- Mishima, S.; Nagai, A.; Abdullah, S.; Matsuda, C.; Taketani, T.; Kumakura, S.; Shibata, H.; Ishikura, H.; Kim, S.U.; Masuda, J. Effective ex vivo expansion of hematopoietic stem cells using osteoblast-differentiated mesenchymal stem cells is CXCL12 dependent. Eur. J. Haematol. 2010, 84, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Kohara, H.; Noda, M.; Nagasawa, T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006, 25, 977–988. [Google Scholar] [CrossRef] [Green Version]
- Arranz, L.; Sánchez-Aguilera, A.; Martín-Pérez, D.; Isern, J.; Langa, X.; Tzankov, A.; Lundberg, P.; Muntión, S.; Tzeng, I.-S.; Lai, D.-M.; et al. Neuropathy of haematopoietic stem cell niche is essential for myeloproliferative neoplasms. Nature 2014, 512, 78–81. [Google Scholar] [CrossRef]
- Pleyer, L.; Valent, P.; Greil, R. Mesenchymal stem and progenitor cells in normal and dysplastic hematopoiesis-masters of survival and clonality? Int. J. Mol. Sci. 2016, 17, 1009. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, A.; Sturgeon, C.; Siatskas, M.; Ferrer, K.; McIntosh, K.; Patil, S.; Hardy, W.; Devine, S.; Ucker, D.; Deans, R.; et al. Mesenchymal stem cells suppress lymphocyte proliferation in vitro and prolong skin graft survival in vivo. Exp Hematol. 2002, 30, 42–48. [Google Scholar] [CrossRef]
- Nemeth, K.; Leelahavanichkul, A.; Yuen, P.S.; Mayer, B.; Parmelee, A.; Doi, K.; Robey, P.G.; Leelahavanichkul, K.; Koller, B.H.; Brown, J.M.; et al. Bone marrow stromal cells attenuate sepsis via prostaglandin E(2)-dependent reprogramming of host macrophages to increase their interleukin-10 production. Nat. Med. 2009, 15, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Zhang, Y.; Ye, L.; Zhang, T.; Cheng, J.; Chen, G.; Zhang, K.; Yang, Y. Umbilical cord-derived mesenchymal stem cells instruct monocytes towards an IL10-producing phenotype by secreting IL6 and HGF. Sci. Rep. 2016, 6, 37566. [Google Scholar] [CrossRef]
- Luk, F.; Carreras-Planella, L.; Korevaar, S.S.; de Witte, S.F.H.; Borras, F.E.; Betjes, M.G.H.; Baan, C.C.; Hoogduijn, M.J.; Franquesa, M. Inflammatory conditions dictate the effect of mesenchymal stem or stromal cells on B cell function. Front Immunol. 2017, 8, 1042. [Google Scholar] [CrossRef] [Green Version]
- Spaggiari, G.M.; Capobianco, A.; Abdelrazik, H.; Becchetti, F.; Mingari, M.C.; Moretta, L. Mesenchymal stem cells inhibit natural killer-cell proliferation, cytotoxicity, and cytokine production: Role of indoleamine 2,3-dioxygenase and prostaglandin E2. Blood 2008, 111, 1327–1333. [Google Scholar] [CrossRef]
- Najar, M.; Raicevic, G.; Fayyad-Kazan, H.; Bron, D.; Toungouz, M.; Lagneaux, L. Mesenchymal stromal cells and immunomodulation: A gathering of regulatory immune cells. Cytotherapy 2016, 18, 160–171. [Google Scholar] [CrossRef]
- Augello, A.; Tasso, R.; Negrini, S.M.; Amateis, A.; Indiveri, F.; Cancedda, R.; Pennesi, G. Bone marrow mesenchymal progenitor cells inhibit lymphocyte proliferation by activation of the programmed death 1 pathway. Eur. J. Immunol. 2005, 35, 1482–1490. [Google Scholar] [CrossRef]
- Starczynowski, D.T.; Kuchenbauer, F.; Argiropoulos, B.; Sung, S.; Morin, R.; Muranyi, A.; Hirst, M.; Hogge, D.; Marra, M.; Wells, R.A.; et al. Identification of miR-145 and miR-146a as mediators of the 5q-syndrome phenotype. Nat. Med. 2010, 16, 49–58. [Google Scholar] [CrossRef]
- Hu, Y.Z.; Xue, Q.; Chen, Y.J.; Yu, G.H.; Qing, M.-D.; Shen, Y.; Wang, M.Y.; Shi, Q.; Zhang, X.-G. Different roles of PD-L1 and FasL in immunomodulation mediated by human placenta-derived mesenchymal stem cells. Hum. Immunol. 2013, 74, 267–276. [Google Scholar]
- Ghannam, S.; Pene, J.; Moquet-Torcy, G.; Jorgensen, C.; Yssel, H. Mesenchymal stem cells inhibit human Th17 cell differentiation and function and induce a T regulatory cell phenotype. J. Immunol. 2010, 185, 302–312. [Google Scholar] [CrossRef] [Green Version]
- Luz-Crawford, P.; Noel, D.; Fernandez, X.; Khoury, M.; Figueroa, F.; Carrión, F.; Jorgensen, C.; Djouad, F. Mesenchymal stem cells repress Th17 molecular program through the PD-1 pathway. PLoS ONE 2012, 7, e45272. [Google Scholar] [CrossRef]
- Medinger, M.; Drexler, B.; Lengerke, C.; Passweg, J. Pathogenesis of Acquired Aplastic Anemia and the Role of the Bone Marrow Microenvironment. Front Oncol. 2018, 5, 587. [Google Scholar] [CrossRef] [Green Version]
- Scheinberg, P. Activity of eltrombopag in severe aplastic anemia. Hematol. Am. Soc. Hematol. Educ. Program. 2018, 2018, 450–456. [Google Scholar] [CrossRef] [Green Version]
- Young, N.S.; Maciejewski, J. Mechanisms of disease—The pathophysiology of acquired aplastic anemia. N. Engl. J. Med. 1997, 336, 1365–1372. [Google Scholar] [CrossRef]
- Hoffman, R.; Zanjani, E.D.; Lutton, J.D.; Zalusky, R.; Wasserman, L.R. Suppression of erythroid-colony formation by lymphocytes from patients with aplastic anemia. N. Engl. J. Med. 1977, 296, 10–13. [Google Scholar] [CrossRef]
- Risitano, A.M.; Maciejewski, J.P.; Green, S.; Plasilova, M.; Zeng, W.; Young, N.S. In-vivo dominant immune responses in aplastic anaemia: Molecular tracking of putatively pathogenetic T-cell clones by TCR beta-CDR3 sequencing. Lancet 2004, 364, 355–364. [Google Scholar] [CrossRef]
- Shallis, R.M.; Ahmad, R.; Zeidan, A.M. Aplastic anemia: Etiology, molecular pathogenesis and emerging concepts. Eur. J. Haematol. 2018, 101, 711–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.H.; Miyazato, A.; Chen, G.B.; Kajigaya, S.; Young, N.S.; Maciejewski, J.P. Interferon-γ-induced gene expression in CD34 cells: Identification of pathologic cytokine-specific signature profiles. Blood 2006, 107, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Wu, Z.; Lin, Z.; Hollinger, M.; Chen, J.; Feng, X.; Young, N.S. Macrophage TNF-α licenses donor T cells in murine bone marrow failure and can be implicated in human aplastic anemia. Blood J. Am. Soc. Hematol. 2018, 132, 2730–2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacigalupo, A. How I Treat Acquired Aplastic Anemia. Blood 2017, 129, 1428–1436. [Google Scholar] [CrossRef]
- Lu, S.H.; Ge, M.L.; Zheng, Y.Z.; Yang, S.G.; Chen, F.; You, Y.H.; Han, Z.C. Effect of CD106 + Mesenchymal Stem Cell on Bone Marrow Vascular Failure in Patients With Aplastic Anemia. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2018, 40, 178–186. [Google Scholar]
- Deng, S.; Xiang, J.J.; Shen, Y.Y.; Lin, S.Y.; Zeng, Y.Q.; Shen, J.P. Effects of VEGF-Notch Signaling Pathway on Proliferation and Apoptosis of Bone Marrow MSC in Patients with Aplastic Anemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2019, 27, 1925–1932. [Google Scholar]
- Li, N.; Liu, L.; Liu, Y.; Luo, S.; Song, Y.; Fang, B. miR-144-3p Suppresses Osteogenic Differentiation of BMSCs from Patients with Aplastic Anemia through Repression of TET2. Mol. Ther. Nucleic Acids. 2020, 19, 619–626. [Google Scholar] [CrossRef]
- Li, H.; Wang, L.; Pang, Y.; Jiang, Z.; Liu, Z.; Xiao, H.; Chen, H.; Ge, X.; Lan, H.; Xiao, Y. In patients with chronic aplastic anemia, bone marrow-derived MSCs regulate the Treg/Th17 balance by influencing the Notch/RBP-J/FOXP3/RORγt pathway. Sci. Rep. 2017, 7, 42488. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Chen, J.; Huang, F.; Wang, J.; Su, W.; Zhou, J.; Qi, Q.; Cao, F.; Sun, B.; Liu, Z.; et al. Human Gingiva Tissue-Derived MSC Ameliorates Immune-Mediated Bone Marrow Failure of Aplastic Anemia via Suppression of Th1 and Th17 Cells and Enhancement of CD4 + Foxp3+ Regulatory T Cells Differentiation. Am. J. Transl. Res. 2019, 11, 7627–7643. [Google Scholar]
- Li, J.; Yang, S.; Lu, S.; Zhao, H.; Feng, J.; Li, W.; Ma, F.; Ren, Q.; Liu, B.; Zhang, L.; et al. Differential Gene Expression Profile Associated With the Abnormality of Bone Marrow Mesenchymal Stem Cells in Aplastic Anemia. PLoS ONE 2012, 7, e47764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, J.; Zhang, L.; Ren, X.; Li, C.; Li, X.; Dong, P.; Zheng, X.; Huang, J.; Shao, Y.; Ge, M.; et al. Multifaceted Characterization of the Signatures and Efficacy of Mesenchymal stem/stromal Cells in Acquired Aplastic Anemia. Stem Cell Res. Ther. 2020, 11, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bueno, C.; Roldan, M.; Anguita, E.; Romero-Moya, D.; Martín-Antonio, B.; Rosu-Myles, M.; del Cañizo, C.; Campos, F.; García, R.; Gómez-Casares, M.; et al. Bone Marrow Mesenchymal Stem Cells From Patients With Aplastic Anemia Maintain Functional and Immune Properties and Do Not Contribute to the Pathogenesis of the Disease. Haematologica 2014, 99, 1168–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantelli, M.; Avanzini, M.A.; Rosti, V.; Ingo, D.M.; Conforti, A.; Novara, F.; Arrigo, G.; Boni, M.; Zappatore, R.; Lenta, E.; et al. Comprehensive Characterization of Mesenchymal Stromal Cells From Patients With Fanconi Anaemia. Br. J. Haematol. 2015, 170, 826–836. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; He, Y.; Xing, W.; Zhang, P.; Shi, H.; Chen, S.; Shi, J.; Bai, J.; Rhodes, S.D.; Zhang, F.; et al. An abnormal bone marrow microenvironment contributes to hematopoietic dysfunction in Fanconi anemia. Haematologica 2017, 102, 1017–1027. [Google Scholar] [CrossRef] [Green Version]
- Bardelli, D.; Dander, E.; Bugarin, C.; Cappuzzello, C.; Pievani, A.; Fazio, G.; Pierani, P.; Corti, P.; Farruggia, P.; Dufour, C.; et al. Mesenchymal stromal cells from Shwachman-Diamond syndrome patients fail to recreate a bone marrow niche in vivo and exhibit impaired angiogenesis. Br. J. Haematol. 2018, 182, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Zambetti, N.A.; Ping, Z.; Chen, S.; Kenswil, K.J.G.; Mylona, M.A.; Sanders, M.A.; Hoogenboezem, R.M.; Bindels, E.M.J.; Adisty, M.N.; Van Strien, P.M.H.; et al. Mesenchymal Inflammation Drives Genotoxic Stress in Hematopoietic Stem Cells and Predicts Disease Evolution in Human Pre-leukemia. Cell Stem Cell 2016, 19, 613–627. [Google Scholar] [CrossRef] [Green Version]
- Gonzaga, V.F.; Wenceslau, C.V.; Lisboa, G.S.; Frare, E.O.; Kerkis, I. Mesenchymal stem cell benefits observed in bone marrow failure and acquired aplastic anemia. Stem Cells Int. 2017, 2017, 8076529. [Google Scholar] [CrossRef] [Green Version]
- Kfoury, Y.; Scadden, D.T. Mesenchymal cell contributions to the stem cell niche. Cell Stem Cell 2015, 16, 239–253. [Google Scholar] [CrossRef] [Green Version]
- Scheinberg, P. Mesenchymal stromal cells: Filling the void of immunosuppressive therapy in aplastic anemia? Cytotherapy 2013, 15, 751–752. [Google Scholar] [CrossRef]
- de Lima, M.; McNiece, I.; Robinson, S.N.; Munsell, M.; Eapen, M.; Horowitz, M.; Alousi, A.; Saliba, R.; McMannis, J.D.; Kaur, I.; et al. Cord-blood engraftment with ex vivo mesenchymal-cell coculture. N. Engl. J. Med. 2012, 367, 2305–2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Mo, W.; Zhang, Y.; Zhou, M.; Li, Y.; Zhou, R.; Xu, S.; Pan, S.; Deng, H.; Mao, P.; et al. Vascular and Perivascular Niches, but Not the Osteoblastic Niche, Are Numerically Restored Following Allogeneic Hematopoietic Stem Cell Transplantation in Patients With Aplastic Anemia. Int. J. Hematol. 2017, 106, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Jiang, Z.-J.; Pang, Y.; Li, L.; Gao, Y.; Xiao, H.W.; Li, Y.-H.; Zhang, H.; Liu, Q. Efficacy and safety of mesenchymal stromal cell treatment from related donors for patients with refractory aplastic anemia. Cytotherapy 2013, 15, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Clé, D.V.; Santana-Lemos, B.; Tellechea, M.F.; Prata, K.L.; Orellana, M.D.; Covas, D.T.; Calado, R.T. Intravenous Infusion of Allogeneic Mesenchymal Stromal Cells in Refractory or Relapsed Aplastic Anemia. Cytotherapy 2015, 17, 1696–1705. [Google Scholar] [CrossRef]
- Pang, Y.; Xiao, H.W.; Zhang, H.; Liu, Z.H.; Li, L.; Gao, Y.; Li, H.B.; Jiang, Z.J.; Tan, H.; Lin, J.R.; et al. Allogeneic Bone-Marrow-Derived Mesenchymal stromal Cells Expanded in vitro for Treatment of Aplastic Anemia: A Multicenter Phase II Trial. Stem Cells Transl. Med. 2017, 6, 1949. [Google Scholar] [CrossRef]
- Xu, L.X.; Cao, Y.B.; Wang, Z.H.; Liu, Z.Y.; Liu, B.; Zhao, D.D.; DA, W.M.; Gao, C.J.; Wu, X.X. Efficacy of haploidentical allogeneic bone marrow hematopoietic stem cell transplantation combined with umbilical cord blood derived mesenchymal stem cells for severe aplastic anemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2011, 19, 1241–1245. [Google Scholar]
- Xu, L.X.; Liu, Z.Y.; Wu, Y.M.; Cao, Y.B.; Li, X.H.; Yan, B.; Yang, X.L.; DA, W.M.; Wu, X.X. Clinical observation of haploidentical-hematopoietic stem cell transplantation combined with human umbilical cord-derived mesenchymal stem cells for severe aplastic anemia-II. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2014, 22, 774–778. [Google Scholar]
- Li, X.H.; Gao, C.J.; Da, W.M.; Cao, Y.B.; Wang, Z.H.; Xu, L.X.; Wu, Y.M.; Liu, B.; Liu, Z.Y.; Yan, B.; et al. Reduced intensity conditioning, combined transplantation of haploidentical hematopoietic stem cells and mesenchymal stem cells in patients with severe aplastic anemia. PLoS ONE 2014, 9, e89666. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhang, Y.; Xiao, H.; Yao, Z.; Zhang, H.; Liu, Q.; Wu, B.; Nie, D.; Li, Y.; Pang, Y.; et al. Cotransplantation of bone marrow-derived mesenchymal stem cells in haploidentical hematopoietic stem cell transplantation in patients with severe aplastic anemia: An interim summary for a multicenter phase II trial results. Bone Marrow Transpl. 2017, 52, 704–710. [Google Scholar] [CrossRef]
- Xu, L.; Liu, Z.; Wu, Y.; Yang, X.; Cao, Y.; Li, X.; Yan, B.; Li, S.; Da, W.; Wu, X. Clinical Evaluation of Haploidentical Hematopoietic Combined With Human Umbilical Cord-Derived Mesenchymal Stem Cells in Severe Aplastic Anemia. Eur. J. Med. Res. 2018, 23, 12. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.Y.; Fu, Y.W.; Wang, Q.; Ai, H.; Wang, Y.Q.; Li, T.T.; Zhou, J.; Fang, B.J.; Wei, X.D.; Song, Y.P. The efficacy and safety of co-transplantation of unrelated donor peripheral blood stem cells combined with umbilical mesenchymal stem cells in patients with refractory severe aplastic anemia-Ⅱ. Zhonghua Nei Ke Za Zhi 2019, 58, 819–822. [Google Scholar]
- Wang, H.; Yan, H.; Wang, Z.; Zhu, L.; Liu, J.; Guo, Z. Cotransplantation of allogeneic mesenchymal and hematopoietic stem cells in children with aplastic anemia. Pediatrics 2012, 29, e1612–e1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Wang, Z.-D.; Zheng, X.-L.; Ding, L.; Han, D.-M.; Yan, H.-M.; Wang, H.-X. Clinical Observation of Haploidentical Hematopoietic Stem Cell Transplantation Combined With Bone-marrow Derived Mesenchymal Stem Cells Transfusion for Treatment of Children With Severe Aplastic Anemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2017, 25, 1158–1164. [Google Scholar] [PubMed]
- Wang, Z.; Yu, H.; Cao, F.; Liu, Z.; Liu, Z.; Feng, W.; Liu, X.; Yu, Y.; Xiao, Y.; Li, L.; et al. Donor-derived marrow mesenchymal stromal cell co-transplantation following a haploidentical hematopoietic stem cell transplantation trail to treat severe aplastic anemia in children. Ann. Hematol. 2019, 98, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Ding, Y.; Gao, Y.; Li, L.; Pang, Y.; Liu, Z.; Zhang, H.; Xiao, Y.; Jiang, Z.; Xiao, H. Cotransplantation of haploidentical hematopoietic stem cells and allogeneic bone marrow-derived mesenchymal stromal cells as a first-line treatment in very severe aplastic anemia patients with refractory infections. Eur. J. Haematol. 2018, 100, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhu, C.; Ma, D.; Gu, Z.; Xu, C.; Wang, F.; Chen, J.-G.; Liu, C.-J.; Guan, L.-X.; Gao, R.; et al. Efficacy and safety of mesenchymal stromal cells for the prophylaxis of chronic graft-versus-host disease after allogeneic hematopoietic stem cell transplantation: A meta-analysis of randomized controlled trials. Ann. Hematol. 2018, 97, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Hinden, L.; Avner, M.; Stepensky, P.; Or, R.; Almogi-Hazan, O. Lymphocyte counts may predict a good response to mesenchymal stromal cells therapy in graft versus host disease patients. PLoS ONE 2019, 14, e0217572. [Google Scholar] [CrossRef]
- Liu, L.L.; Liu, L.; Liu, H.H.; Ren, S.-S.; Dou, C.-Y.; Cheng, P.-P.; Wang, C.-L.; Wang, L.-N.; Chen, X.-L.; Zhanget, H.; et al. Levamisole suppresses adipogenesis of aplastic anaemia-derived bone marrow mesenchymal stem cells through ZFP36L1-PPARGC1B axis. J. Cell Mol. Med. 2018, 22, 4496–4506. [Google Scholar] [CrossRef]
- Qu, Y.; Lin, Q.; Yuan, Y.; Sun, Z.; Li, P.; Wang, F.; Jiang, H.; Chen, T. Cyclosporin A inhibits adipogenic differentiation and regulates immunomodulatory functions of murine mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2018, 498, 516–522. [Google Scholar] [CrossRef]
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 Revision to the World Health Organization Classification of Myeloid Neoplasms and Acute Leukemia. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef]
- Stahl, M.; DeVeaux, M.; de Witte, T.; Neukirchen, J.; Sekeres, M.A.; Brunner, A.M.; Roboz, G.J.; Steensma, D.P.; Bhatt, V.R.; Platzbecker, U.; et al. The use of immunosuppressive therapy in MDS: Clinical outcomes and their predictors in a large international patient cohort. Blood Adv. 2018, 2, 1765–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaninoni, A.; Imperiali, F.G.; Cattaneo, A.; Soverini, G.; Binda, F.; Porretti, L.; Cortelezzi, A.; Barcellini, W. Detection of erythroblast antibodies in mitogen-stimulated bone marrow cultures from patients with myelodysplastic syndromes. Transfusion 2016, 56, 2037–2041. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, P.L.; Tuechler, H.; Schanz, J.; Sanz, G.; Garcia-Manero, G.; Solé, F.; Bennett, J.M.; Bowen, D.; Fenaux, P.; Dreyfus, F.; et al. Revised international prognostic scoring system for myelodysplastic syndromes. Blood 2012, 120, 2454–2465. [Google Scholar] [CrossRef] [PubMed]
- Malcovati, L.; Papaemmanuil, E.; Ambaglio, I.; Elena, C.; Gallì, A.; Della Porta, M.G.; Travaglino, E.; Pietra, D.; Pascutto, C.; Ubezio, M.; et al. Driver somatic mutations identify distinct disease entities within myeloid neoplasms with myelodysplasia. Blood 2014, 124, 1513–1521. [Google Scholar] [CrossRef]
- Sekeres, M.A. The Epidemiology of Myelodysplastic Syndromes. Hematol. Oncol. Clin. North Am. 2010, 24, 287–294. [Google Scholar] [CrossRef]
- Montalban-Bravo, G.; Garcia-Manero, G. Myelodysplastic syndromes: 2018 update on diagnosis, risk-stratification and management. Am. J. Hematol. 2018, 93, 129–147. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.; Fenaux, P.; List, A.F.; Raza, A.; Platzbecker, U.; Santini, V. Recent advances in the treatment of lower-risk non-del(5q) myelodysplastic syndromes (MDS). Leuk. Res. 2017, 52, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Santini, V. How I Treat MDS After Hypomethylating Agent Failure. Blood 2019, 133, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Scalzulli, E.; Pepe, S.; Colafigli, G.; Breccia, B. Therapeutic Strategies in Low and High-Risk MDS: What Does the Future Have to Offer? Blood Rev. 2020, 100689. [Google Scholar] [CrossRef]
- Zhao, Z.G.; Xu, W.; Yu, H.P.; Fang, B.L.; Wu, S.H.; Li, F.; Li, W.M.; Li, Q.B.; Chen, Z.C.; Zou, P. Functional Characteristics of Mesenchymal Stem Cells Derived From Bone Marrow of Patients With Myelodysplastic Syndromes. Cancer Lett. 2012, 317, 136–143. [Google Scholar] [CrossRef]
- Ferrer, R.A.; Wobus, M.; List, C.; Wehner, R.; Schönefeldt, C.; Brocard, B.; Mohr, B.; Rauner, M.; Schmitz, M.; Stiehler, M.; et al. Mesenchymal Stromal Cells From Patients With Myelodyplastic Syndrome Display Distinct Functional Alterations That Are Modulated by Lenalidomide. Haematologica 2013, 98, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Geyh, S.; Oz, S.; Cadeddu, R.P.; Fröbel, J.; Brückner, B.; Kündgen, A.; Fenk, R.; Bruns, I.; Zilkens, C.; Hermsen, D.; et al. Insufficient Stromal Support in MDS Results From Molecular and Functional Deficits of Mesenchymal Stromal Cells. Leukemia 2013, 27, 1841–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, S.; Kumar, S.; Srivastava, V.M.; Therese, M.M.; Nair, S.C.; Abraham, A.; Mathews, V.; George, B.; Srivastava, A. Heterogeneity of Mesenchymal Stromal Cells in Myelodysplastic Syndrome-with Multilineage Dysplasia (MDS-MLD). Indian J. Hematol. Blood Transfus. 2019, 35, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Umezu, T.; Imanishi, S.; Asano, M.; Yoshizawa, S.; Katagiri, S.; Ohyashiki, K.; Ohyashiki, J.H. Genetic variations of bone marrow mesenchymal stromal cells derived from acute leukemia and myelodysplastic syndrome by targeted deep sequencing. Leuk. Res. 2017, 62, 23–28. [Google Scholar] [CrossRef]
- Blau, O.; Baldus, C.D.; Hofmann, W.K.; Thiel, G.; Nolte, F.; Burmeister, T.; Türkmen, S.; Benlasfer, O.; Schümann, E.; Sindram, A.; et al. Mesenchymal stromal cells of myelodysplastic syndrome and acute myeloid leukemia patients have distinct genetic abnormalities compared with leukemic blasts. Blood 2011, 118, 5583–5592. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Villar, O.; Garcia, J.L.; Sanchez-Guijo, F.M.; Robledo, C.; Villaron, E.M.; Hernández-Campo, P.; Lopez-Holgado, N.; Diez-Campelo, M.; Barbado, M.V.; Perez-Simon, J.A.; et al. Both expanded and uncultured mesenchymal stem cells from MDS patients are genomically abnormal, showing a specific genetic profile for the 5q- syndrome. Leukemia 2009, 23, 664–672. [Google Scholar] [CrossRef] [Green Version]
- Flynn, C.M.; Kaufman, D.S. Donor Cell Leukemia: Insight Into Cancer Stem Cells and the Stem Cell Niche. Blood 2007, 109, 2688–2692. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, M.H.G.P.; Mukherjee, S.; Guo, S.; Zhang, S.; Kobayashi, T.; Schoonmaker, J.A.; Ebert, B.L.; Al-Shahrour, F.; Hasserjian, R.P.; Scadden, E.O.; et al. Bone Progenitor Dysfunction Induces Myelodysplasia and Secondary Leukaemia. Nature 2010, 464, 852–857. [Google Scholar] [CrossRef] [Green Version]
- Ozdogan, H.; Gur Dedeoglu, B.; Oztemur Islakoglu, Y.; Aydos, A.; Kose, S.; Atalay, A.; Yegin, Z.A.; Avcu, F.; Uckan Cetinkaya, D.; Ilhan, O. DICER1 Gene and miRNA Dysregulation in Mesenchymal Stem Cells of Patients With Myelodysplastic Syndrome and Acute Myeloblastic Leukemia. Leuk. Res. 2017, 63, 62–71. [Google Scholar] [CrossRef]
- Kode, A.; Manavalan, J.S.; Mosialou, I.; Bhagat, G.; Rathinam, C.V.; Luo, N.; Khiabanian, H.; Lee, A.; Murty, V.V.; Friedman, R.; et al. Leukaemogenesis Induced by an Activating β-catenin Mutation in Osteoblasts. Nature 2014, 506, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Fernald, A.A.; Anastasi, J.; Le Beau, M.M.; Qian, Z. Haploinsufficiency of Apc Leads to Ineffective Hematopoiesis. Blood 2010, 115, 3481–3488. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, A.; Wang, J.; Hu, C.; Fernald, A.A.; Davis, E.M.; Cheng, J.X.; Le Beau, M.M. Inhibition of WNT Signaling in the Bone Marrow Niche Prevents the Development of MDS in the Apcdel/+ MDS Mouse Model. Blood 2017, 129, 2959–2970. [Google Scholar] [CrossRef] [PubMed]
- Ping, Z.; Chen, S.; Hermans, S.J.F.; Kenswil, K.J.G.; Feyen, J.; van Dijk, C.; Bindels, E.M.J.; Mylona, A.M.; Adisty, N.M.; Hoogenboezem, R.M.; et al. Activation of NF-kB driven inflammatory programs in mesenchymal elements attenuates hematopoiesis in low-risk myelodysplastic syndromes. Leukemia 2019, 33, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zambetti, N.A.; Bindels, E.M.J.; Kenswill, K.; Mylona, A.M.; Adisty, N.M.; Hoogenboezem, R.M.; Sanders, M.A.; Cremers, E.M.P.; Westers, T.M.; et al. Massive Parallel RNA Sequencing of Highly Purified Mesenchymal Elements in Low-Risk MDS Reveals Tissue-Context-Dependent Activation of Inflammatory Programs. Leukemia 2016, 30, 1938–1942. [Google Scholar] [CrossRef] [Green Version]
- Abegunde, S.O.; Buckstein, R.; Wells, R.A.; Rauh, M.J. An inflammatory environment containing TNF-alpha favors Tet2-mutant clonal hematopoiesis. Exp. Hematol. 2018, 59, 60–65. [Google Scholar] [CrossRef]
- Fozza, C.; Crobu, V.; Isoni, M.A.; Dore, F. The immune landscape of myelodysplastic syndromes. Crit. Rev. Oncol. Hematol. 2016, 107, 90–99. [Google Scholar] [CrossRef]
- Kotsianidis, I.; Bouchliou, I.; Nakou, E.; Spanoudakis, E.; Margaritis, D.; Christophoridou, A.V.; Anastasiades, A.; Tsigalou, C.; Bourikas, G.; Karadimitris, A.; et al. Kinetics, Function and Bone Marrow Trafficking of CD4 + CD25 + FOXP3+ Regulatory T Cells in Myelodysplastic Syndromes (MDS). Leukemia 2009, 23, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Kordasti, S.Y.; Ingram, W.; Hayden, J.; Darling, D.; Barber, L.; Afzali, B.; Lombardi, G.; Wlodarski, M.W.; Maciejewski, J.P.; Farzaneh, F.; et al. CD4+ CD25high Foxp3+ Regulatory T Cells in Myelodysplastic Syndrome (MDS). Blood 2007, 110, 847–850. [Google Scholar] [CrossRef] [Green Version]
- Munn, D.H.; Mellor, A.L. IDO in the Tumor Microenvironment: Inflammation, Counter-Regulation, and Tolerance. Trends Immunol. 2016, 37, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Zhi-Gang, Z.; Wei-Ming, L.; Zhi-Chao, C.; Yong, Y.; Ping, Z. Immunosuppressive Properties of Mesenchymal Stem Cells Derived From Bone Marrow of Patient With Hematological Malignant Diseases. Leuk. Lymphoma 2008, 49, 2187–2195. [Google Scholar] [CrossRef]
- Wu, L.; Amarachintha, S.; Xu, J.; Oley, F.; Du, W. Mesenchymal COX2-PG Secretome Engages NR4A-WNT Signalling Axis in Haematopoietic Progenitors to Suppress Anti-Leukaemia Immunity. Br. J. Haematol. 2018, 183, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Sarhan, D.; Wang, J.; Sunil Arvindam, U.; Hallstrom, C.; Verneris, M.R.; Grzywacz, B.; Warlick, E.; Blazar, B.R.; Miller, J.S. Mesenchymal Stromal Cells Shape the MDS Microenvironment by Inducing Suppressive Monocytes That Dampen NK Cell Function. JCI Insight 2020, 5, e130155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterman, R.S.; Tomchuck, S.L.; Henkle, S.L.; Betancourt, A.M. A New Mesenchymal Stem Cell (MSC) Paradigm: Polarization Into a Pro-Inflammatory MSC1 or an Immunosuppressive MSC2 Phenotype. PLoS ONE 2010, 5, e10088. [Google Scholar] [CrossRef] [PubMed]
- Medyouf, H.; Mossner, M.; Jann, J.C.; Nolte, F.; Raffel, S.; Herrmann, C.; Lier, A.; Eisen, C.; Nowak, V.; Zens, B.; et al. Myelodysplastic cells in patients reprogram mesenchymal stromal cells to establish a transplantable stem cell niche disease unit. Cell Stem Cell 2014, 14, 824–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cull, A.H.; Snetsinger, B.; Buckstein, R.; Wells, R.A.; Rauh, M.J. Tet2 restrains inflammatory gene expression in macrophages. Exp. Hematol. 2017, 55, 56–70.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vas, V.; Senger, K.; Dörr, K.; Niebel, A.; Geiger, H. Aging of the microenvironment influences clonality in hematopoiesis. PLoS ONE 2012, 7, e42080. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Lee, W.Y.; Huang, B.; Zhang, J.F.; Wu, T.; Jiang, X.; Wang, C.C.; Li, G. Secretome of Human Fetal Mesenchymal Stem Cell Ameliorates Replicative Senescen. Stem Cells Dev. 2016, 25, 1755–1766. [Google Scholar] [CrossRef]
- Zheng, Q.; Zhao, Y.; Guo, J.; Zhao, S.; Fei, C.; Xiao, C.; Wu, D.; Wu, L.; Li, X.; Chang, C. Iron Overload Promotes Mitochondrial Fragmentation in Mesenchymal Stromal Cells From Myelodysplastic Syndrome Patients Through Activation of the AMPK/MFF/Drp1 Pathway. Cell Death Dis. 2018, 9, 515. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Hope, K.J.; Zhai, Q.; Smadja-Joffe, F.; Dick, J.E. Targeting of CD44 eradicates human acute myeloid leukemic stem cells. Nat. Med. 2006, 12, 1167–1174. [Google Scholar] [CrossRef]
- Matsunaga, T.; Takemoto, N.; Sato, T.; Takimoto, R.; Tanaka, I.; Fujimi, A.; Akiyama, T.; Kuroda, H.; Kawano, Y.; Kobune, M.; et al. Interaction between leukemic-cell VLA-4 and stromal fibronectin is a decisive factor for minimal residual disease of acute myelogenous leukemia. Nat. Med. 2003, 9, 1158–1165. [Google Scholar] [CrossRef]
- Zeng, Z.; Shi, Y.X.; Samudio, I.J.; Wang, R.Y.; Ling, X.; Frolova, O.; Levis, M.; Rubin, J.B.; Negrin, R.R.; Estey, E.H.; et al. Targeting the leukemia microenvironment by CXCR4 inhibition overcomes resistance to kinase inhibitors and chemotherapy in AML. Blood 2009, 113, 6215–6224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uy, G.L.; Rettig, M.P.; Motabi, I.H.; McFarland, K.; Trinkaus, K.M.; Hladnik, L.M.; Kulkarni, S.; Abboud, C.N.; Cashen, A.F.; Stockerl-Goldstein, K.E.; et al. A phase 1/2 study of chemosensitization with the CXCR4 antagonist plerixafor in relapsed or refractory acute myeloid leukemia. Blood 2012, 119, 3917–3924. [Google Scholar] [CrossRef] [PubMed]
- Carter, B.Z.; Mak, P.Y.; Wang, X.; Yang, H.; Garcia-Manero, G.; Mak, D.H.; Mu, H.; Ruvolo, V.R.; Qiu, Y.; Coombes, K.; et al. Focal Adhesion Kinase as a Potential Target in AML and MDS. Mol. Cancer Ther. 2017, 16, 1133–1144. [Google Scholar] [CrossRef] [Green Version]
- Kastrinaki, M.C.; Pavlaki, K.; Batsali, A.K.; Kouvidi, E.; Mavroudi, I.; Pontikoglou, C.; Papadaki, H.A. Mesenchymal Stem Cells in Immune-Mediated Bone Marrow Failure Syndromes. Clin. Dev. Immunol. 2013, 2013, 265608. [Google Scholar] [CrossRef] [Green Version]
- Chao, M.P.; Takimoto, C.H.; Feng, D.D.; McKenna, K.; Gip, P.; Liu, J.; Volkmer, J.P.; Weissman, I.L.; Majeti, R. Therapeutic Targeting of the Macrophage Immune Checkpoint CD47 in Myeloid Malignancies. Front. Oncol. 2020, 9, 1380. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Fu, R.; Wang, H.; Li, L.; Liu, H.; Shao, Z. CD47 Is Expressed Abnormally on Hematopoietic Cells in Myelodysplastic Syndrome. Leuk. Res. 2013, 37, 907–910. [Google Scholar] [CrossRef] [PubMed]
- Wobus, M.; Benath, G.; Ferrer, R.A.; Wehner, R.; Schmitz, M.; Hofbauer, L.C.; Rauner, M.; Ehninger, G.; Bornhäuser, M.; Platzbecker, U. Impact of Lenalidomide on the Functional Properties of Human Mesenchymal Stromal Cells. Exp. Hematol. 2012, 40, 867–876. [Google Scholar] [CrossRef]
- Iancu-Rubin, C.; Mosoyan, G.; Wang, J.; Kraus, T.; Sung, V.; Hoffman, R. Stromal cell-mediated inhibition of erythropoiesis can be attenuated by Sotatercept (ACE-011), an activin receptor type II ligand trap. Exp. Hematol. 2013, 41, 155–166. [Google Scholar] [CrossRef]
- Boada, M.; Echarte, L.; Guillermo, C.; Diaz, L.; Touriño, C.; Grille, S. 5-Azacytidine Restores Interleukin 6-increased Production in Mesenchymal Stromal Cells From Myelodysplastic Patients. Hematol. Transfus. Cell Ther. 2020. [Google Scholar] [CrossRef]
- Wenk, C.; Garz, A.K.; Grath, S.; Huberle, C.; Witham, D.; Weickert, M.; Malinverni, R.; Niggemeyer, J.; Kyncl, M.; Hecker, J.; et al. Direct Modulation of the Bone Marrow Mesenchymal Stromal Cell Compartment by Azacitidine Enhances Healthy Hematopoiesis. Blood Adv. 2018, 2, 3447–3461. [Google Scholar] [CrossRef] [Green Version]
- Roversi, F.M.; Cury, N.M.; Lopes, M.R.; Ferro, K.P.; Machado-Neto, J.A.; Alvarez, M.C.; Dos Santos, G.P.; Giardini Rosa, R.; Longhini, A.L.; Duarte, A.D.S.S.; et al. Up-regulation of SPINT2/HAI-2 by Azacytidine in bone marrow mesenchymal stromal cells affects leukemic stem cell survival and adhesion. J. Cell. Mol. Med. 2019, 23, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Geng, S.; Zhang, H.; Lai, P.; Liao, P.; Zeng, L.; Lu, Z.; Weng, J.; Du, X. Phenotype of mesenchymal stem cells from patients with myelodyplastic syndrome maybe partly modulated by decitabine. Oncol. Lett. 2019, 18, 4457–4466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaian, E.; Weidner, H.; Wobus, M.; Baschant, U.; Jacobi, A.; Mies, A.; Bornhäuser, M.; Guck, J.; Hofbauer, L.C.; Rauner, M.; et al. Effects of rigosertib on the osteo-hematopoietic niche in myelodysplastic syndromes. Ann. Hematol. 2019, 98, 2063–2072. [Google Scholar] [CrossRef] [PubMed]


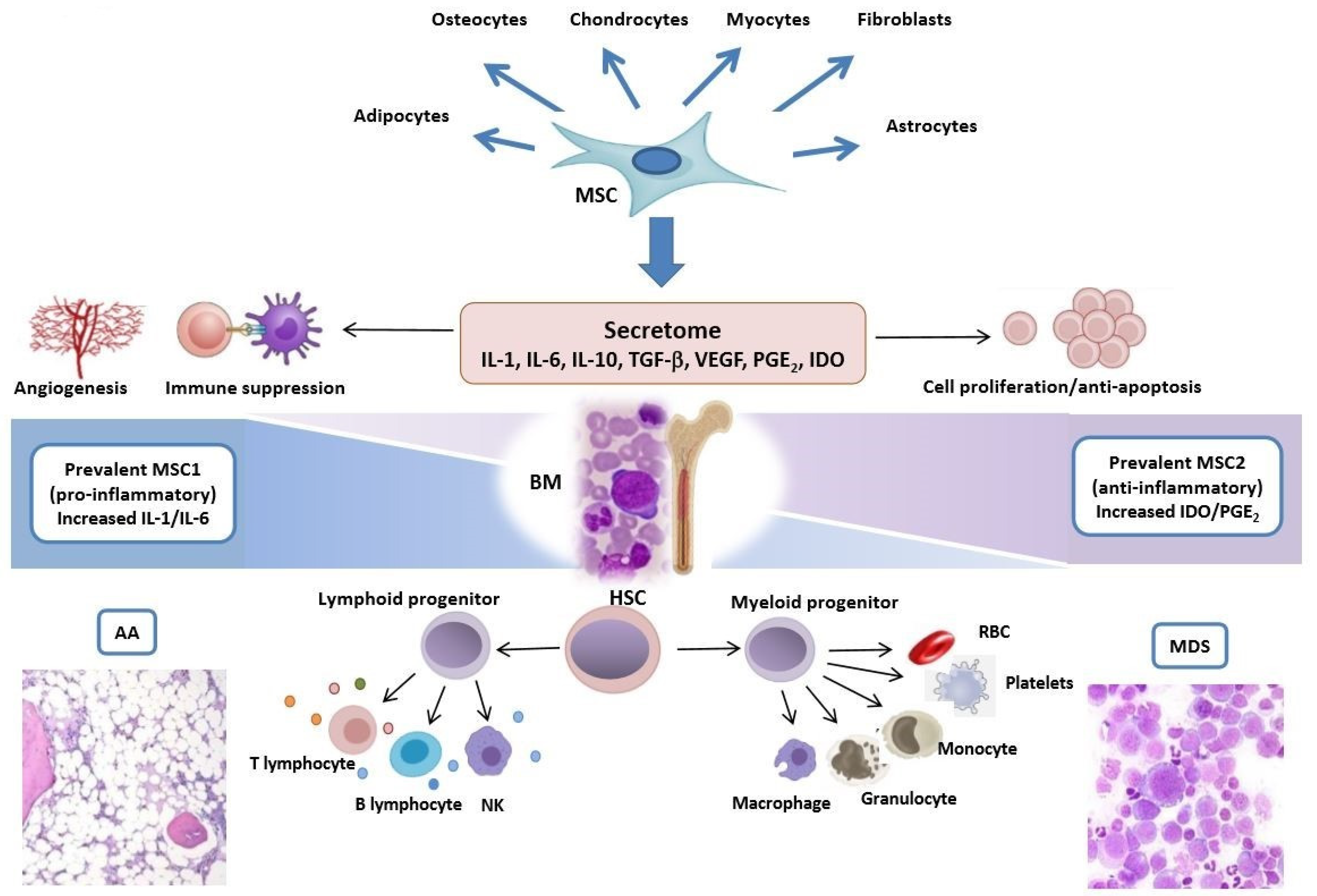{kind=link}
| Reference | N. of Patients | Main Findings |
|---|---|---|
| Lu S.H., et al. 2018 | - | Deficient or decreased expression of CD106 + MSCs accelerates bone marrow vascularization failure in AA patients |
| Deng S., et al. 2019 | - | The activation of VEGF–Notch pathway can restore the proliferation function of MSCs in AA patients |
| Li N., et al. 2020 | 3 | The microRNA miR-144-3p is involved in AA pathogenesis. Its targeting may be a therapeutic strategy |
| Li H., et al. 2017 | 15 | Bone marrow-derived MSCs modulate the levels of T-helper (Th)1, Th2, Th17, and T-regulatory cells, as well as their related cytokines in AA patients |
| Zhao J., et al. 2019 | - | Gingival-derived MSCs markedly improved survival and attenuated histological bone marrow damage in AA murine models |
| Li J., et al. 2012 | - | MSCs from AA patients show differential gene expression profiles associated with bone marrow failure |
| Huo J., et al. 2020 | 49 | MSCs from AA patients exhibited multifaceted defects in biological characteristics and alterative molecular genetics in the whole genome |
| Bueno C., et al. 2014 | - | MSC cultures from the bone marrow of AA patients display the same phenotype and differentiation potential as their counterparts from normal controls |
| Reference | N. of Patients | Main Findings |
|---|---|---|
| Infusion of MSCs in AA patients | ||
| Xiao Y., et al. 2012 | 18 | Sequential infusions of MSCs may improve hematopoiesis in AA. Six patients responded, 2 reached complete response at 3 months. |
| Clé D.V., et al. 2015 | 9 | Infusion of allogeneic MSCs in AA is safe but does not improve clinical hematologic response or engraft in recipient bone marrow. Two patients responded at 6 months. |
| Pang Y., et al. 2017 | 74 | In phase II prospective trial, MSC infusions evoked a response in 28.4% of cases (6.8% complete) in less than one month, with 88% OS at 17 months |
| Cotransplantation of MSCs and allogeneic hematopoietic stem cells | ||
| Xu L-X., et al. 2011 and 2014 | 5 | Haploidentical HSC transplant combined with umbilical cord MSC infusion allows engraftment with 12.5% incidence of severe GVHD and 25% mortality |
| Li X-H., et al. 2014 | 17 | Combined transplantation of haploidentical HSCs and MSCs on AA without an HLA-identical sibling was safe, reduced the incidence of severe GVHD (23.5%), and lead to good survival (88.2% and 76.5% at 3 and 6 months) |
| Liu Z., et al. 2017 | 44 | Cotransplantation of MSCs could reduce the risk of graft failure and severe GvHD in haploidentical setting. Acute GVHD occurred in 1/3 of cases and chronic one in 14.6%. The 12-month overall survival was 77.3% |
| Xu L., et al. 2018 | 24 | MSC and haploidentical HSC cotransplantation is effective and safe, with 50% acute GVHD, not related to gender, age, donor–recipient relations, and patient/donor pair. Patient/donor pair was significantly correlated with chronic GVHD. One year mortality was 20.8% |
| Zhao M., et al. 2019 | 25 | Peripheral blood HSCs from unrelated and related donor transplants and MSC infusions are effective and safe in AA. No severe acute GVHD and 21.7% chronic GVHD were registered. Overall survival was 84% at 22.9 months (median follow up). The 5-year overall survival rate after transplantation was 83.6% ± 7.5% |
| Wang H., et al. 2012 | 6 | Cotransplant of haploidentical HSCs and MSCs in children was effective and safe with no severe acute GVHD nor chronic GvHD, and 100% survival at 15 months |
| Wei W., et al. 2017 | 25 | Cotransplant of haploidentical HSCs and bone marrow-derived MSCs led to engraftment in 100% of cases (median time 12 days, range 11–22 days). Acute GvHD occurred in 64% cases (5 cases grades II–IV), and one patient died of grade IV skin, gut, and liver GvHD. Five cases developed chronic GVHD. |
| Wang Z., et al. 2019 | 35 | Cotransplant of haploidentical HSCs and MSCs led to hematopoietic reconstitution in 100% of cases (median time 14 days, range 10–22 days). Grade II/IV acute GvHD occurred in 26% cases and 23% developed chronic GVHD. The overall survival rate was 85.71%, with a median of 22 months (range 3.5–37 months). |
| Yue C., et al. 2018 | 6 | HLA-related donor HSCT and MSC infusion led to sustained, full donor chimerism, with a median time of myeloid/platelet engraftment of 13–15.5 days. One patient died of acute GVHD, and 5 patients were alive after a median follow-up of 21 months (range 17–40.5). |
| Hinden L., et al. 2019 | 26 | T-cell and NK-cell increased levels may predict a good response to HSCT and MSC cotransplantation. A better response was observed among patients who expressed low levels of IL-6 and IL-22, Th17-related cytokines, prior to therapy. |
| Other therapeutic mechanisms for MSCs | ||
| Liu L.L., et al. 2018 | - | Levamisole displayed a significant suppressive effect on the in-vitro adipogenic differentiation of bone marrow-derived MSCs from AA patients |
| Qu Y., et al. 2018 | - | Cyclosporin A suppressed adipogenic differentiation of MSCs by inhibiting interleukin-6 expression in AA |
| Reference | N. of Patients | Main Findings |
|---|---|---|
| MSC alterations in MDSs | ||
| Zhao Z.G., et al. 2012 | 14 | BM-derived MDS–MSCs showed reduced hematopoiesis support function compared to their normal counterparts and impaired capacity to inhibit T-lymphocyte activation and proliferation in vitro. |
| Ferrer R.A., et al. 2013 | - | Cocultures of MDS–MSCs with CD34+ cells from healthy donors resulted in reduced numbers of colony-forming units. Lenalidomide exposure of low-risk MDS–MSCs was able to rescue erythroid and myeloid colony formation. |
| Geyh S., et al. 2013 | 106 | MDS–MSCs exhibit reduced proliferative capacities and altered expression of key molecules involved in HSC proliferation. |
| Abbas S., et al. 2019 | 6 | Compared with healthy controls’ MSCs, MDS–MSCs displayed a shift towards increased apoptosis, lower expression of VEGF, SCF, and ANGPT, aberrant expression pattern in the Notch signaling pathway, and increase in Wnt signaling inhibitors. |
| Azuma K., et al. 2017 | 5 | MDS–MSCs showed some genetic variants with very low allelic frequency (7–8%), such as NF1–G2114D and NF1–G140, not shared by dysplastic HSCs. |
| Blau O., et al. 2011 | 43 | Cytogenetic aberrations in MSCs were detected in 16% of MDS/AML patients and were different from those observed in the neoplastic HSCs. No chromosomal abnormalities were identified in MSCs of healthy subjects. |
| Lopez-Villar O., et al. 2009 | 36 | MDS–MSCs display genomic alterations, some of them associated with the 5q- syndrome. |
| MSCs induce clonal hematopoiesis in MDSs | ||
| Raaijmakers M., et al. 2010 | - | Deletion of Dicer1, specifically in mouse osteoprogenitors, disrupts hematopoiesis, resulting in MDSs and AML, with neoplastic cells having Dicer1 intact. |
| Ozdogan H., et al. 2017 | 10 | DICER1 gene expression was lower in MDS–MSCs than healthy controls’ MSCs, and resulted in suppression of the physiologic osteogenic differentiation. |
| Kode A., et al. 2014 | 45 | An activating mutation of β-catenin in mouse osteoblasts alters the differentiation potential of HSCs via the activation of Notch signaling, leading to the development of AML. |
| Stoddart A., et al. 2017 | - | Loss of 1 copy of Ctnnb1 is sufficient to prevent the development of MDSs in Apcdel/+ mice; the alteration of WNT signaling in the BM niche is responsible for the disease. |
| MSCs facilitate clonal hematopoiesis in MDSs and immune escape | ||
| Ping Z., et al. 2019 | 45 | Activation of NF-κB in MDS–MSCs leads to transcriptional overexpression of inflammatory factors, including negative regulators of hematopoiesis. |
| Chen S., et al. 2016 | 12 | MSCs from low-risk MDS patients display global activation of inflammatory patterns, with increased NF-kB, EGF, TGF-β, and TNF signaling. |
| Medyouf H., et al. 2014 | 31 | Healthy MSCs acquire MDS–MSC molecular features when exposed to MDS–HSCs and contribute to the propagation of dysplastic HSCs in orthotopic xenografts through the overproduction of N-cadherin, IGFBP2, VEGFA, and LIF. |
| Zheng Q., et al. 2018 | 81 | Iron overload damages MDS–MSCs via the enhancement of the AMPK/MFF/Drp1 pathway, resulting in increased apoptosis, higher ROS levels, and increased mitochondrial fragmentation compared with MSCs from noniron-overloaded patients. |
| Reference | N. of Patients | Main Findings |
|---|---|---|
| Carter B.Z., et al. 2017 | 133 | Inhibition of focal adhesion kinase decreases MSC-mediated adhesion/migration and viability of MDS/AML cells and prolongs survival in a xenograft murine model. |
| Wobus M., et al. 2012 | - | Lenalidomide modulates expression of cell surface molecules and chemokine secretion of MSCs in vitro, reducing the migration of HSCs. |
| Iancu-Rubin C., et al. 2013 | - | MSCs treated with sotatercept changed their molecular and secretory profile, increasing the expression and secretion of erythropoiesis-stimulating factors in vitro. |
| Boada M., et al. 2020 | 35 | In vitro treatment with AZA leads to a significant reduction in IL-6 production by the MDS–MSCs. |
| Wenk C., et al. 2018 | - | AZA regulates the expression of extracellular matrix molecules and interferon pathway components, exerting a direct effect on MDS–MSCs and favoring healthy over malignant HSC expansion in vitro. |
| Roversi F.M., et al. 2019 | 10 | AZA upregulates SPINT2/HAI-2 expression, which is downregulated in MDSs due to methylation in AML/MDS–MSCs in an in-vitro study. |
| Pang Y., et al. 2019 | 28 | Treatment with decitabine increases the number of MSCs in G2/M phase and powers the ability of MDS–MSCs to induce the differentiation of T-cells into regulatory T-cells in vitro. |
| Balaian E., et al. 2019 | - | Rigosertib exerts inhibitory effects on the stromal components of the osteohematopoietic niche, including MSCs, in a murine model. This may explain the dissociation between antileukemic activity and the absence of hematological improvement. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fattizzo, B.; Giannotta, J.A.; Barcellini, W. Mesenchymal Stem Cells in Aplastic Anemia and Myelodysplastic Syndromes: The “Seed and Soil” Crosstalk. Int. J. Mol. Sci. 2020, 21, 5438. https://doi.org/10.3390/ijms21155438
Fattizzo B, Giannotta JA, Barcellini W. Mesenchymal Stem Cells in Aplastic Anemia and Myelodysplastic Syndromes: The “Seed and Soil” Crosstalk. International Journal of Molecular Sciences. 2020; 21(15):5438. https://doi.org/10.3390/ijms21155438
Chicago/Turabian StyleFattizzo, Bruno, Juri A. Giannotta, and Wilma Barcellini. 2020. "Mesenchymal Stem Cells in Aplastic Anemia and Myelodysplastic Syndromes: The “Seed and Soil” Crosstalk" International Journal of Molecular Sciences 21, no. 15: 5438. https://doi.org/10.3390/ijms21155438
APA StyleFattizzo, B., Giannotta, J. A., & Barcellini, W. (2020). Mesenchymal Stem Cells in Aplastic Anemia and Myelodysplastic Syndromes: The “Seed and Soil” Crosstalk. International Journal of Molecular Sciences, 21(15), 5438. https://doi.org/10.3390/ijms21155438