The Impact of Small Extracellular Vesicles on Lymphoblast Trafficking across the Blood-Cerebrospinal Fluid Barrier In Vitro

 and
and 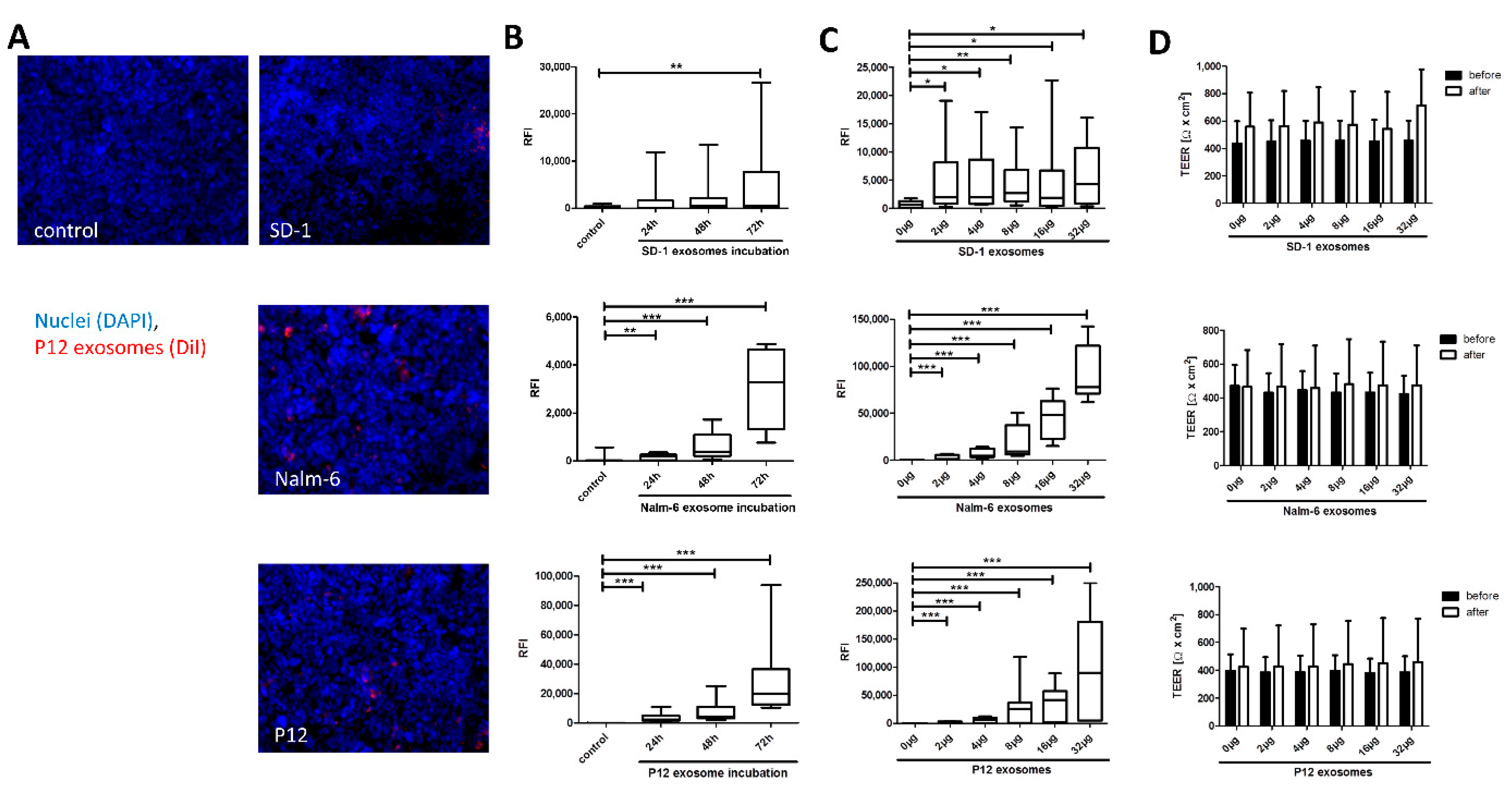{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Exosome Isolation
2.2. Time- and Dose-Dependent Uptake to HiBCPP Cells
2.3. Inhibition of Exosome Association and Uptake
2.4. Leukemia Cell Transmigration across the BCSFB
3. Discussion
3.1. Exosome Conditioning Does Not Alter BCSFB Integrity
3.2. Mechanisms of Exosome Uptake to HiBCPP Cells
3.3. Differential Relevance in BCP-ALL and T-ALL Cell Lines
3.4. Translational Implications
3.5. Summary
4. Materials and Methods
4.1. Cell Culture
4.2. Exosome Isolation
4.3. BCSFB In Vitro Model
4.4. Uptake of Exosomes by BCSFB In Vitro Model
4.5. Transmigration Assay
4.6. Western Blotting
4.7. Viability Testing
4.8. Immunofluorescence Staining
4.9. Nanoparticle Tracking Analysis (NTA)
4.10. Electon Microscopy
4.11. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALL | acute lymphoblastic leukemia |
| BBB | blood-brain barrier |
| BCP-ALL | B-cell precursor acute lymphoblastic leukemia |
| BCSFB | blood-cerebrospinal fluid barrier |
| CCR7 | C-C chemokine receptor type 7 |
| CNS | central nervous system |
| CP | choroid plexus |
| ECM | extracellular matrix |
| ITG | integrin |
| NTA | nanoparticle tracking analysis |
| PSGL-1 | P-selectin glycoprotein ligand 1 |
| RT | room temperature |
| RFI | relative fluorescence intensity |
| T-ALL | T-linage acute lymphoblastic leukemia |
| TEER | transepithelial electrical resistance |
References
- Hunger, S.P.; Mullighan, C.G. Acute lymphoblastic leukemia in children. N. Engl. J. Med. 2015, 373, 1541–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeha, S.; Pei, D.; Choi, J.; Cheng, C.; Sandlund, J.T.; Coustan-Smith, E.; Campana, D.; Inaba, H.; Rubnitz, J.E.; Ribeiro, R.C.; et al. Improved cns control of childhood acute lymphoblastic leukemia without cranial irradiation: St jude total therapy study 16. J. Clin. Oncol. 2019, 37, 3377–3391. [Google Scholar] [CrossRef] [PubMed]
- Lenk, L.; Alsadeq, A.; Schewe, D.M. Involvement of the central nervous system in acute lymphoblastic leukemia: Opinions on molecular mechanisms and clinical implications based on recent data. Cancer Metastasis Rev. 2020, 39, 173–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frishman-Levy, L.; Izraeli, S. Advances in understanding the pathogenesis of cns acute lymphoblastic leukaemia and potential for therapy. Br. J. Haematol. 2017, 176, 157–167. [Google Scholar] [CrossRef]
- Kinjyo, I.; Bragin, D.; Grattan, R.; Winter, S.S.; Wilson, B.S. Leukemia-derived exosomes and cytokines pave the way for entry into the brain. J. Leukoc. Biol. 2019, 105, 741–753. [Google Scholar] [CrossRef]
- Zhou, F.; Wen, Y.; Jin, R.; Chen, H. New attempts for central nervous infiltration of pediatric acute lymphoblastic leukemia. Cancer Metastasis Rev. 2019, 38, 657–671. [Google Scholar] [CrossRef]
- Williams, M.T.; Yousafzai, Y.M.; Elder, A.; Rehe, K.; Bomken, S.; Frishman-Levy, L.; Tavor, S.; Sinclair, P.; Dormon, K.; Masic, D.; et al. The ability to cross the blood-cerebrospinal fluid barrier is a generic property of acute lymphoblastic leukemia blasts. Blood 2016, 127, 1998–2006. [Google Scholar] [CrossRef] [Green Version]
- Gaudichon, J.; Jakobczyk, H.; Debaize, L.; Cousin, E.; Galibert, M.D.; Troadec, M.B.; Gandemer, V. Mechanisms of extramedullary relapse in acute lymphoblastic leukemia: Reconciling biological concepts and clinical issues. Blood Rev. 2019, 36, 40–56. [Google Scholar] [CrossRef]
- Pui, C.H.; Howard, S.C. Current management and challenges of malignant disease in the cns in paediatric leukaemia. Lancet Oncol. 2008, 9, 257–268. [Google Scholar] [CrossRef]
- Si, M.; Jiao, X.; Li, Y.; Chen, H.; He, P.; Jiang, F. The role of cytokines and chemokines in the microenvironment of the blood-brain barrier in leukemia central nervous system metastasis. Cancer Manag. Res. 2018, 10, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Yang, H.; Liu, J.; Zhu, L.; Yu, S.; Zhang, X.; Gao, L. Focus on exosomes: Novel pathogenic components of leukemia. Am. J. Cancer Res. 2019, 9, 1815–1829. [Google Scholar] [PubMed]
- Gener Lahav, T.; Adler, O.; Zait, Y.; Shani, O.; Amer, M.; Doron, H.; Abramovitz, L.; Yofe, I.; Cohen, N.; Erez, N. Melanoma-derived extracellular vesicles instigate proinflammatory signaling in the metastatic microenvironment. Int. J. Cancer 2019, 145, 2521–2534. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Costa-Silva, B.; Shen, T.L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyiadzis, M.; Whiteside, T.L. The emerging roles of tumor-derived exosomes in hematological malignancies. Leukemia 2017, 31, 1259–1268. [Google Scholar] [CrossRef]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.M.; Molaei, F.; Alahari, S.K. Exosomes: Composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol. Cancer 2019, 18, 75. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Garcia, M.; Murakami, J.L.; Chen, C.C. Exosome-mediated microenvironment dysregulation in leukemia. Biochim. Biophys. Acta 2016, 1863, 464–470. [Google Scholar] [CrossRef]
- Almeida, R.S.; Costa, E.S.M.; Coutinho, L.L.; Garcia Gomes, R.; Pedrosa, F.; Massaro, J.D.; Donadi, E.A.; Lucena-Silva, N. Microrna expression profiles discriminate childhood t- from b-acute lymphoblastic leukemia. Hematol. Oncol. 2019, 37, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Price, T.T.; Cantelli, G.; Ngo, B.; Warner, M.J.; Olivere, L.; Ridge, S.M.; Jablonski, E.M.; Therrien, J.; Tannheimer, S.; et al. Leukaemia hijacks a neural mechanism to invade the central nervous system. Nature 2018, 560, 55–60. [Google Scholar] [CrossRef]
- Ghersi-Egea, J.F.; Strazielle, N.; Catala, M.; Silva-Vargas, V.; Doetsch, F.; Engelhardt, B. Molecular anatomy and functions of the choroidal blood-cerebrospinal fluid barrier in health and disease. Acta Neuropathol. 2018, 135, 337–361. [Google Scholar] [CrossRef] [Green Version]
- Naumann, J.A.; Gordon, P.M. In vitro model of leukemia cell migration across the blood-cerebrospinal fluid barrier. Leuk. Lymphoma 2017, 58, 1747–1749. [Google Scholar] [CrossRef]
- Marz, M.; Meyer, S.; Erb, U.; Georgikou, C.; Horstmann, M.A.; Hetjens, S.; Weiss, C.; Fallier-Becker, P.; Vandenhaute, E.; Ishikawa, H.; et al. Pediatric acute lymphoblastic leukemia-conquering the cns across the choroid plexus. Leuk. Res. 2018, 71, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Gunther, R.; Chelstrom, L.M.; Tuel-Ahlgren, L.; Simon, J.; Myers, D.E.; Uckun, F.M. Biotherapy for xenografted human central nervous system leukemia in mice with severe combined immunodeficiency using b43 (anti-cd19)-pokeweed antiviral protein immunotoxin. Blood 1995, 85, 2537–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, M.; Castro, F.V.; Alexander, S.; Smith, D.; Liu, J.; Walker, M.; Bitton, D.; Mulryan, K.; Ashton, G.; Blaylock, M.; et al. Rac2, aep, and icam1 expression are associated with cns disease in a mouse model of pre-b childhood acute lymphoblastic leukemia. Blood 2011, 118, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Shimosato, Y.; Kameya, T.; Kuroki, M.; Kitahara, T.; Minato, K.; Shimoyama, M. Leukemic distribution of a human acute lymphocytic leukemia cell line (ichikawa strain) in nude mice conditioned with whole-body irradiation. Cancer Res. 1978, 38, 3494–3498. [Google Scholar] [PubMed]
- Gossai, N.P.; Gordon, P.M. The role of the central nervous system microenvironment in pediatric acute lymphoblastic leukemia. Front. Pediatr. 2017, 5, 90. [Google Scholar] [CrossRef] [Green Version]
- Thastrup, M.; Marquart, H.V.; Levinsen, M.; Grell, K.; Abrahamsson, J.; Albertsen, B.K.; Frandsen, T.L.; Harila-Saari, A.; Lahteenmaki, P.M.; Niinimaki, R.; et al. Flow cytometric detection of leukemic blasts in cerebrospinal fluid predicts risk of relapse in childhood acute lymphoblastic leukemia: A nordic society of pediatric hematology and oncology study. Leukemia 2020, 34, 336–346. [Google Scholar] [CrossRef]
- Yousafzai, Y.M.; Smith, L.; Smith, A.; Bhatti, S.; Gardiner, M.; Cousins, A.; Fee, F.; Chudleigh, S.; Spence, A.; Taylor, W.; et al. Use of quantitative polymerase chain reaction (qpcr) for the diagnosis and monitoring of cns leukaemia. Leuk. Res. 2019, 87, 106232. [Google Scholar] [CrossRef]
- Macia, E.; Ehrlich, M.; Massol, R.; Boucrot, E.; Brunner, C.; Kirchhausen, T. Dynasore, a cell-permeable inhibitor of dynamin. Dev. Cell 2006, 10, 839–850. [Google Scholar] [CrossRef] [Green Version]
- Nakase, I.; Noguchi, K.; Aoki, A.; Takatani-Nakase, T.; Fujii, I.; Futaki, S. Arginine-rich cell-penetrating peptide-modified extracellular vesicles for active macropinocytosis induction and efficient intracellular delivery. Sci. Rep. 2017, 7, 1991. [Google Scholar] [CrossRef]
- Tian, T.; Zhu, Y.L.; Zhou, Y.Y.; Liang, G.F.; Wang, Y.Y.; Hu, F.H.; Xiao, Z.D. Exosome uptake through clathrin-mediated endocytosis and macropinocytosis and mediating mir-21 delivery. J. Biol. Chem. 2014, 289, 22258–22267. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christianson, H.C.; Svensson, K.J.; van Kuppevelt, T.H.; Li, J.P.; Belting, M. Cancer cell exosomes depend on cell-surface heparan sulfate proteoglycans for their internalization and functional activity. Proc. Natl. Acad. Sci. USA 2013, 110, 17380–17385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, J.; Gomes-Alves, P.; Carvalho, S.B.; Peixoto, C.; Alves, P.M.; Altevogt, P.; Costa, J. Extracellular vesicles from ovarian carcinoma cells display specific glycosignatures. Biomolecules 2015, 5, 1741–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, H.; Tachikawa, M.; Yagi, Y.; Umetsu, M.; Nurdin, A.; Miyauchi, E.; Watanabe, M.; Uchida, Y.; Terasaki, T. Cluster of differentiation 46 is the major receptor in human blood-brain barrier endothelial cells for uptake of exosomes derived from brain-metastatic melanoma cells (sk-mel-28). Mol. Pharm. 2019, 16, 292–304. [Google Scholar] [CrossRef]
- Scharff, B.; Modvig, S.; Marquart, H.V.; Christensen, C. Integrin-mediated adhesion and chemoresistance of acute lymphoblastic leukemia cells residing in the bone marrow or the central nervous system. Front. Oncol. 2020, 10, 775. [Google Scholar] [CrossRef]
- Harima, A.; Nakaseko, C.; Yokota, A.; Kitagawa, M.; Morimoto, C.; Harigaya, K.; Saito, Y. Fibronectin promotes cell proliferation of human pre-b cell line via its interactions with vla-4 and vla-5. Hematology 2008, 13, 236–243. [Google Scholar] [CrossRef]
- Lysaght, J.; Verma, N.K.; Maginn, E.N.; Ryan, J.M.; Campiani, G.; Zisterer, D.M.; Williams, D.C.; Browne, P.V.; Lawler, M.P.; McElligott, A.M. The microtubule targeting agent pbox-15 inhibits integrin-mediated cell adhesion and induces apoptosis in acute lymphoblastic leukaemia cells. Int. J. Oncol. 2013, 42, 239–246. [Google Scholar] [CrossRef]
- Shah Scharff, B.F.S.; Modvig, S.; Thastrup, M.; Levinsen, M.; Degn, M.; Ryder, L.P.; Schmiegelow, K.; Christensen, C.; Marquart, H.V. A comprehensive clinical study of integrins in acute lymphoblastic leukemia indicates a role of alpha6/cd49f in persistent minimal residual disease and alpha5 in the colonization of cerebrospinal fluid. Leuk. Lymphoma 2020, 61, 1714–1718. [Google Scholar] [CrossRef]
- Wortzel, I.; Dror, S.; Kenific, C.M.; Lyden, D. Exosome-mediated metastasis: Communication from a distance. Dev. Cell 2019, 49, 347–360. [Google Scholar] [CrossRef]
- Jost, T.R.; Borga, C.; Radaelli, E.; Romagnani, A.; Perruzza, L.; Omodho, L.; Cazzaniga, G.; Biondi, A.; Indraccolo, S.; Thelen, M.; et al. Role of cxcr4-mediated bone marrow colonization in cns infiltration by t cell acute lymphoblastic leukemia. J. Leukoc. Biol. 2016, 99, 1077–1087. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.T.; Yousafzai, Y.; Cox, C.; Blair, A.; Carmody, R.; Sai, S.; Chapman, K.E.; McAndrew, R.; Thomas, A.; Spence, A.; et al. Interleukin-15 enhances cellular proliferation and upregulates cns homing molecules in pre-b acute lymphoblastic leukemia. Blood 2014, 123, 3116–3127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsadeq, A.; Fedders, H.; Vokuhl, C.; Belau, N.M.; Zimmermann, M.; Wirbelauer, T.; Spielberg, S.; Vossen-Gajcy, M.; Cario, G.; Schrappe, M.; et al. The role of zap70 kinase in acute lymphoblastic leukemia infiltration into the central nervous system. Haematologica 2017, 102, 346–355. [Google Scholar] [CrossRef] [Green Version]
- Roth, P.; Keller, A.; Hoheisel, J.D.; Codo, P.; Bauer, A.S.; Backes, C.; Leidinger, P.; Meese, E.; Thiel, E.; Korfel, A.; et al. Differentially regulated mirnas as prognostic biomarkers in the blood of primary cns lymphoma patients. Eur. J. Cancer 2015, 51, 382–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pando, A.; Reagan, J.L.; Quesenberry, P.; Fast, L.D. Extracellular vesicles in leukemia. Leuk. Res. 2018, 64, 52–60. [Google Scholar] [CrossRef]
- Engelhardt, B.; Ransohoff, R.M. Capture, crawl, cross: The t cell code to breach the blood-brain barriers. Trends Immunol. 2012, 33, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Van der Velden, V.H.; de Launaij, D.; de Vries, J.F.; de Haas, V.; Sonneveld, E.; Voerman, J.S.; de Bie, M.; Revesz, T.; Avigad, S.; Yeoh, A.E.; et al. New cellular markers at diagnosis are associated with isolated central nervous system relapse in paediatric b-cell precursor acute lymphoblastic leukaemia. Br. J. Haematol. 2016, 172, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Buonamici, S.; Trimarchi, T.; Ruocco, M.G.; Reavie, L.; Cathelin, S.; Mar, B.G.; Klinakis, A.; Lukyanov, Y.; Tseng, J.C.; Sen, F.; et al. Ccr7 signalling as an essential regulator of cns infiltration in t-cell leukaemia. Nature 2009, 459, 1000–1004. [Google Scholar] [CrossRef] [Green Version]
- Shechter, R.; London, A.; Schwartz, M. Orchestrated leukocyte recruitment to immune-privileged sites: Absolute barriers versus educational gates. Nat. Rev. Immunol. 2013, 13, 206–218. [Google Scholar] [CrossRef]
- Fernández-Sevilla, L.M.; Valencia, J.; Flores-Villalobos, M.A.; Gonzalez-Murillo, A.; Sacedon, R.; Jimenez, E.; Ramirez, M.; Varas, A.; Vicente, A. The choroid plexus stroma constitutes a sanctuary for paediatric b-cell precursor acute lymphoblastic leukaemia in the central nervous system. J. Pathol. 2020. [Google Scholar] [CrossRef]
- Ishiwata, I.; Ishiwata, C.; Ishiwata, E.; Sato, Y.; Kiguchi, K.; Tachibana, T.; Hashimoto, H.; Ishikawa, H. Establishment and characterization of a human malignant choroids plexus papilloma cell line (hibcpp). Hum. Cell 2005, 18, 67–72. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erb, U.; Hikel, J.; Meyer, S.; Ishikawa, H.; Worst, T.S.; Nitschke, K.; Nuhn, P.; Porubsky, S.; Weiss, C.; Schroten, H.; et al. The Impact of Small Extracellular Vesicles on Lymphoblast Trafficking across the Blood-Cerebrospinal Fluid Barrier In Vitro. Int. J. Mol. Sci. 2020, 21, 5491. https://doi.org/10.3390/ijms21155491
Erb U, Hikel J, Meyer S, Ishikawa H, Worst TS, Nitschke K, Nuhn P, Porubsky S, Weiss C, Schroten H, et al. The Impact of Small Extracellular Vesicles on Lymphoblast Trafficking across the Blood-Cerebrospinal Fluid Barrier In Vitro. International Journal of Molecular Sciences. 2020; 21(15):5491. https://doi.org/10.3390/ijms21155491
Chicago/Turabian StyleErb, Ulrike, Julia Hikel, Svenja Meyer, Hiroshi Ishikawa, Thomas S. Worst, Katja Nitschke, Philipp Nuhn, Stefan Porubsky, Christel Weiss, Horst Schroten, and et al. 2020. "The Impact of Small Extracellular Vesicles on Lymphoblast Trafficking across the Blood-Cerebrospinal Fluid Barrier In Vitro" International Journal of Molecular Sciences 21, no. 15: 5491. https://doi.org/10.3390/ijms21155491
APA StyleErb, U., Hikel, J., Meyer, S., Ishikawa, H., Worst, T. S., Nitschke, K., Nuhn, P., Porubsky, S., Weiss, C., Schroten, H., Adam, R., & Karremann, M. (2020). The Impact of Small Extracellular Vesicles on Lymphoblast Trafficking across the Blood-Cerebrospinal Fluid Barrier In Vitro. International Journal of Molecular Sciences, 21(15), 5491. https://doi.org/10.3390/ijms21155491