Pharmacogenomics of Antibiotics


Abstract
:1. Introduction
2. Antibiotics
2.1. The Beta-Lactams
2.1.1. Penicillins
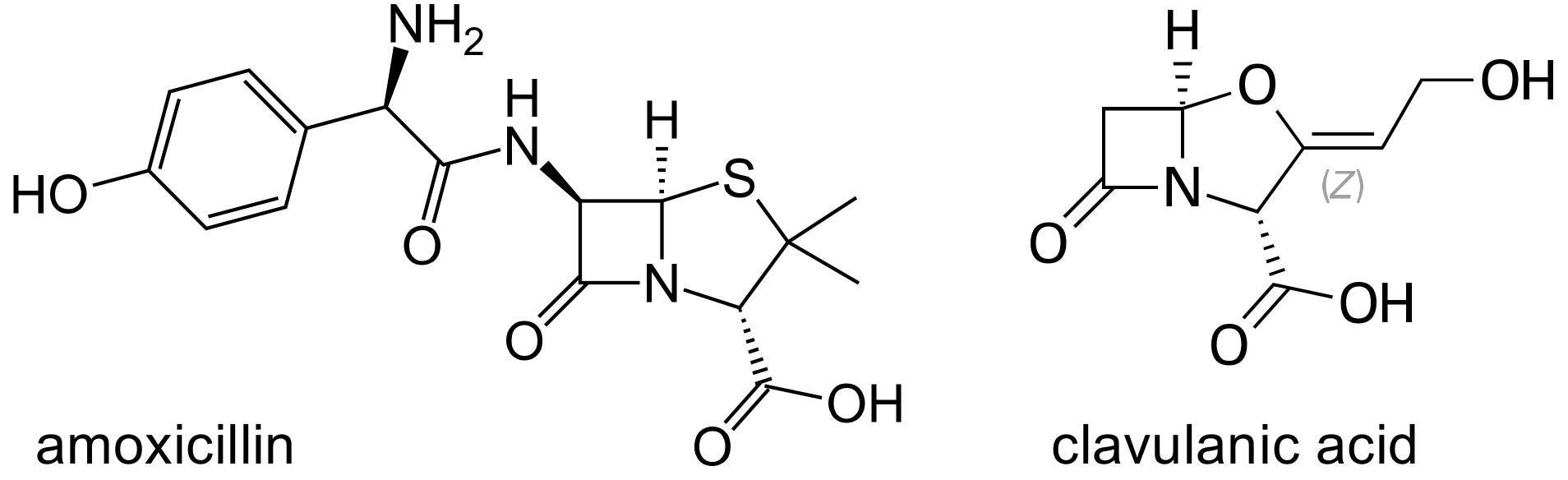
The Aminopenicillins, Ampicillin and Amoxicillin
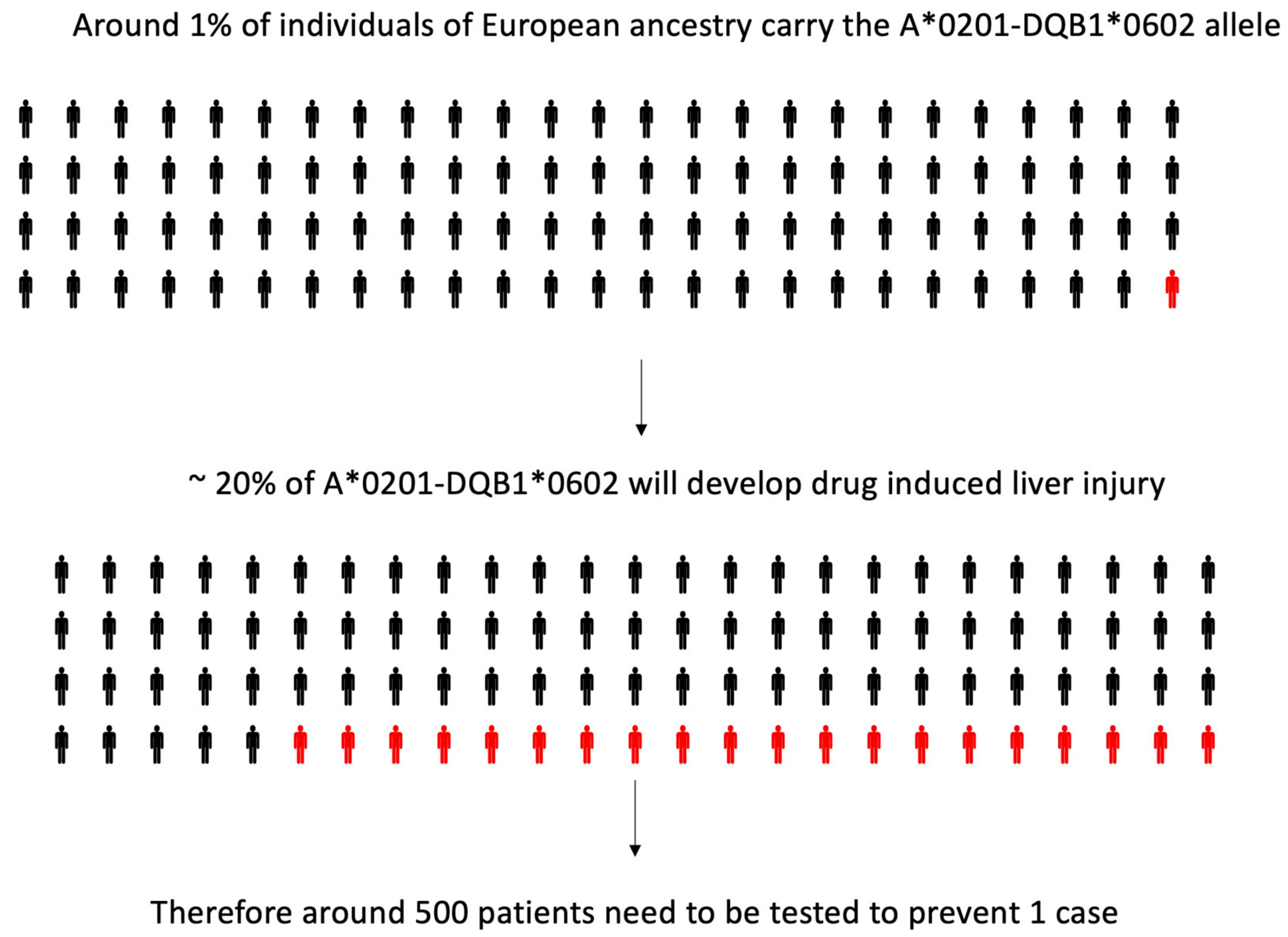
Amoxicillin Clavulanate
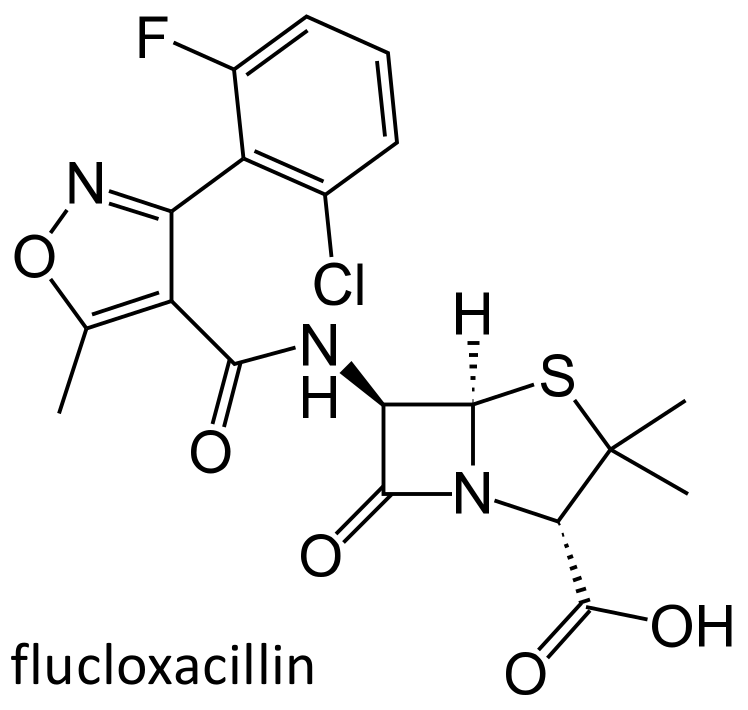
Flucloxacillin
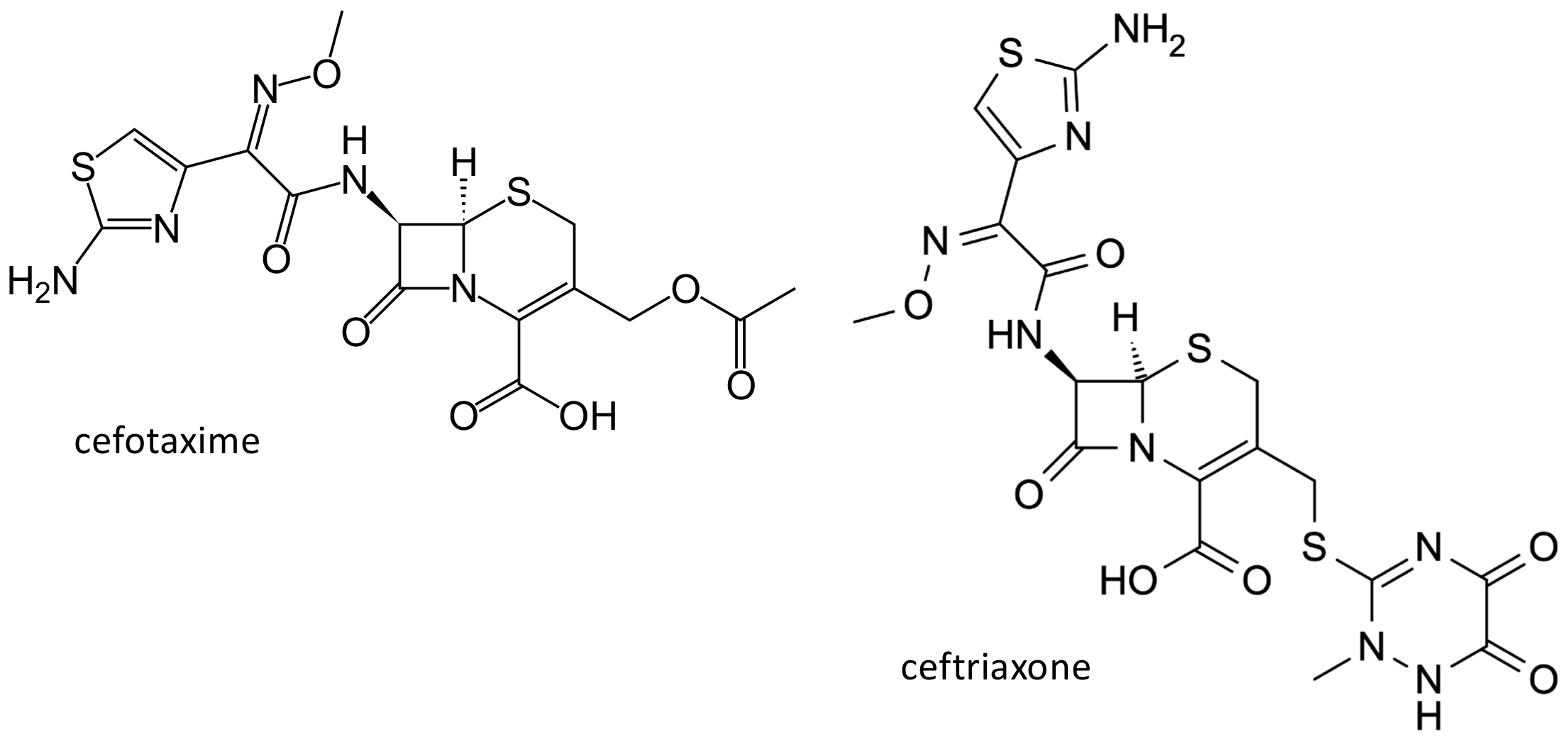
2.1.2. Cephalosporins
2.1.3. Beta-Lactam Induced Neutropenia
2.2. The Macrolides
2.2.1. Erythromycin
2.2.2. Azithromycin
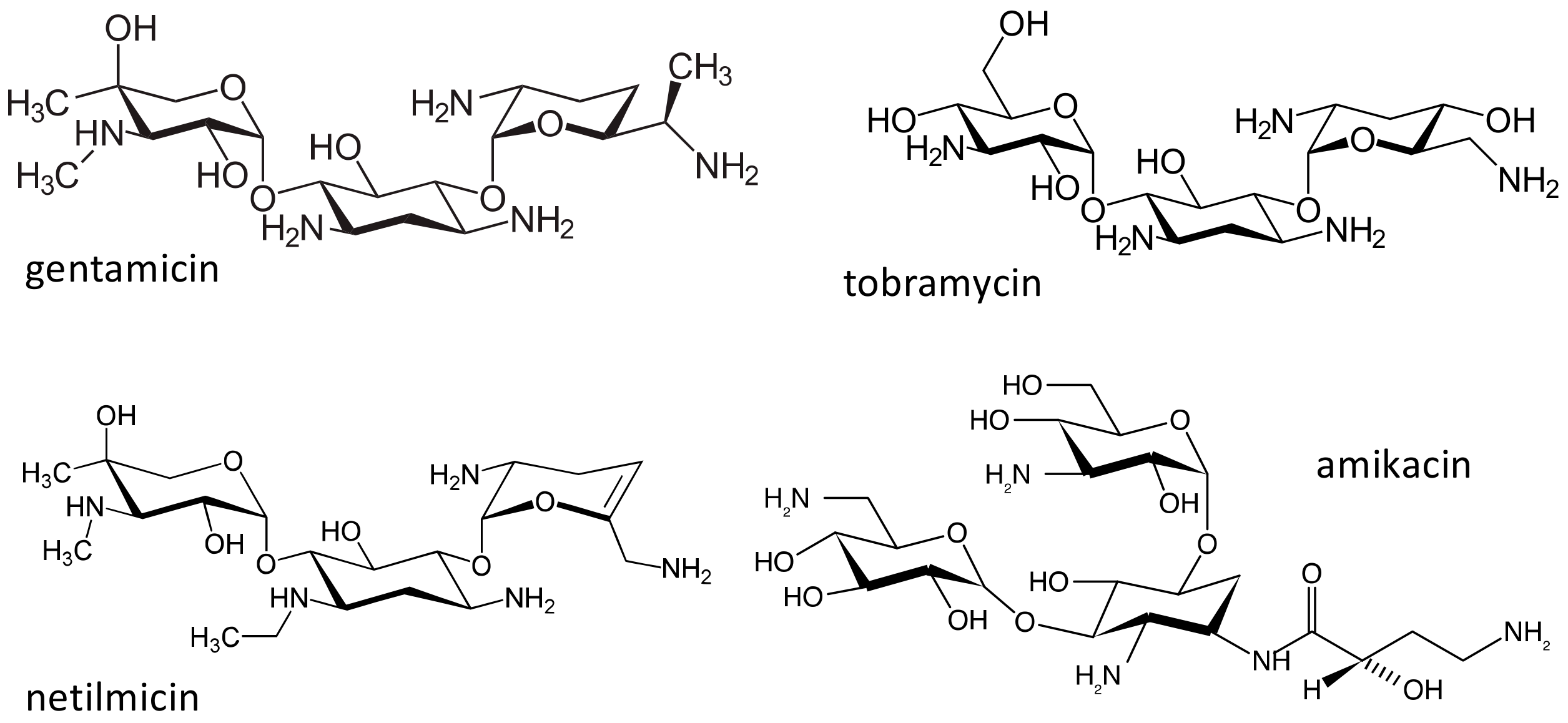
2.3. The Aminoglycosides
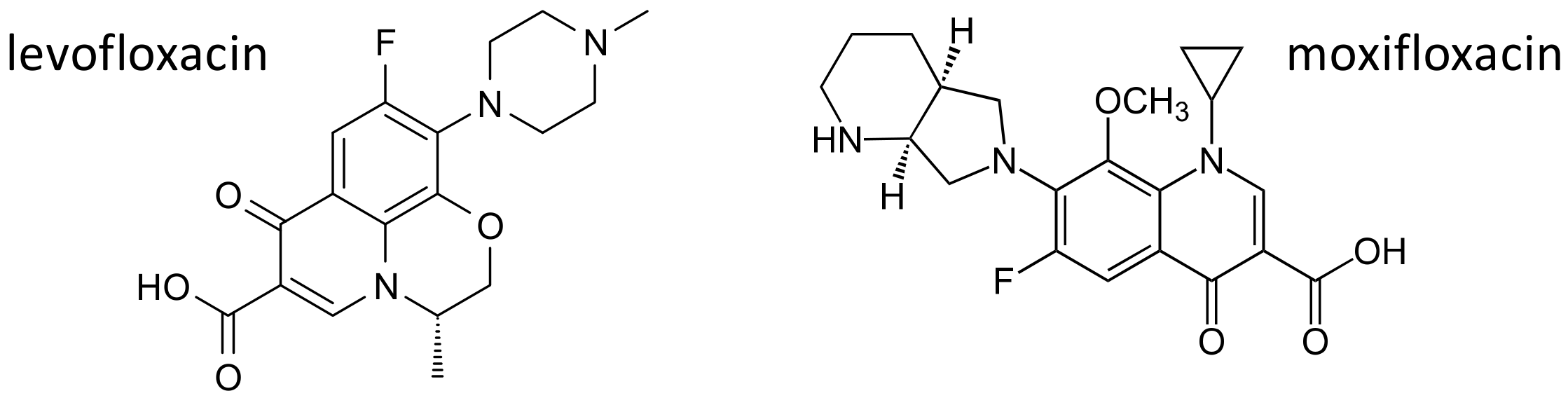
2.4. The Fluoroquinolones
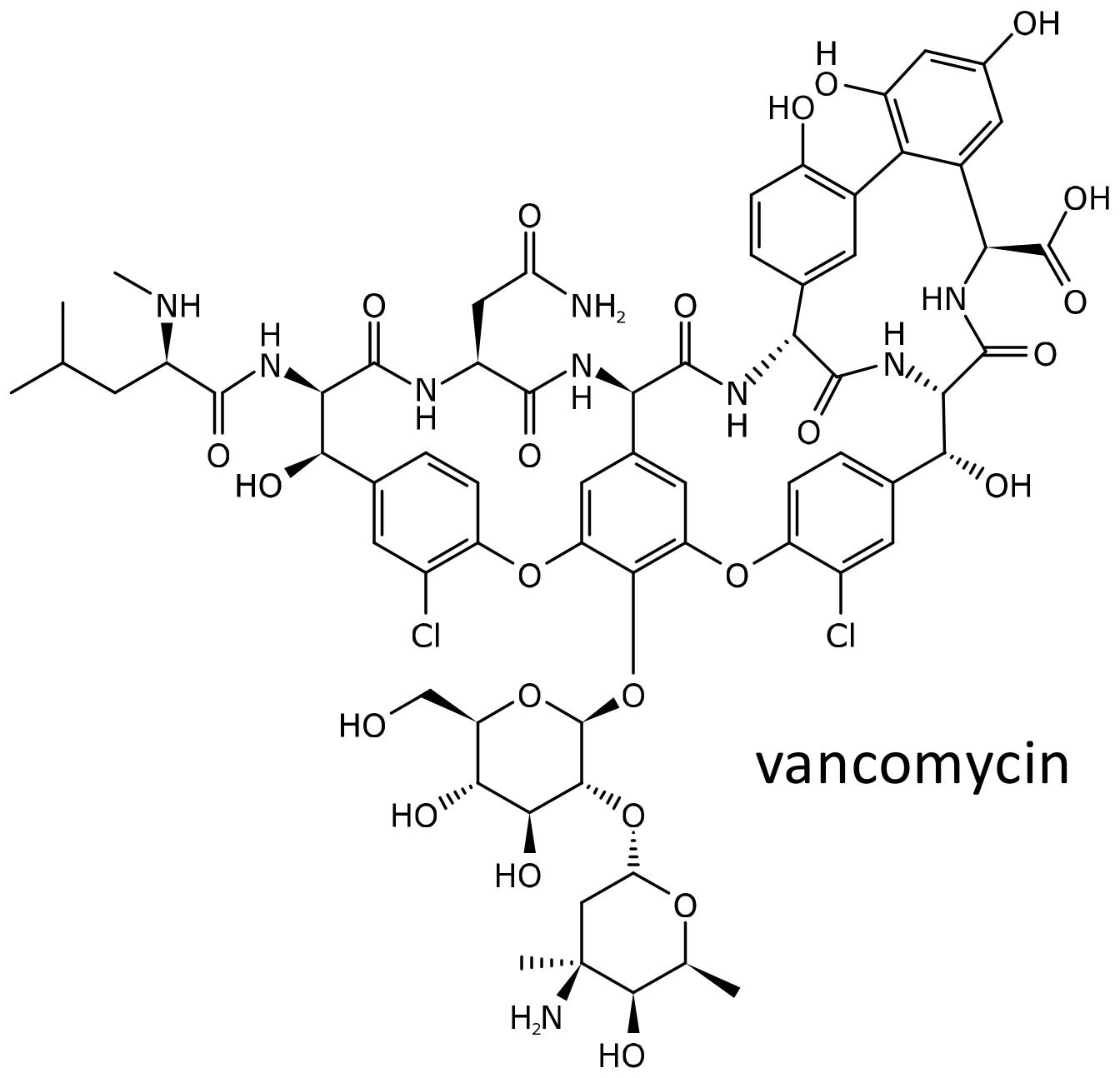
2.5. Vancomycin
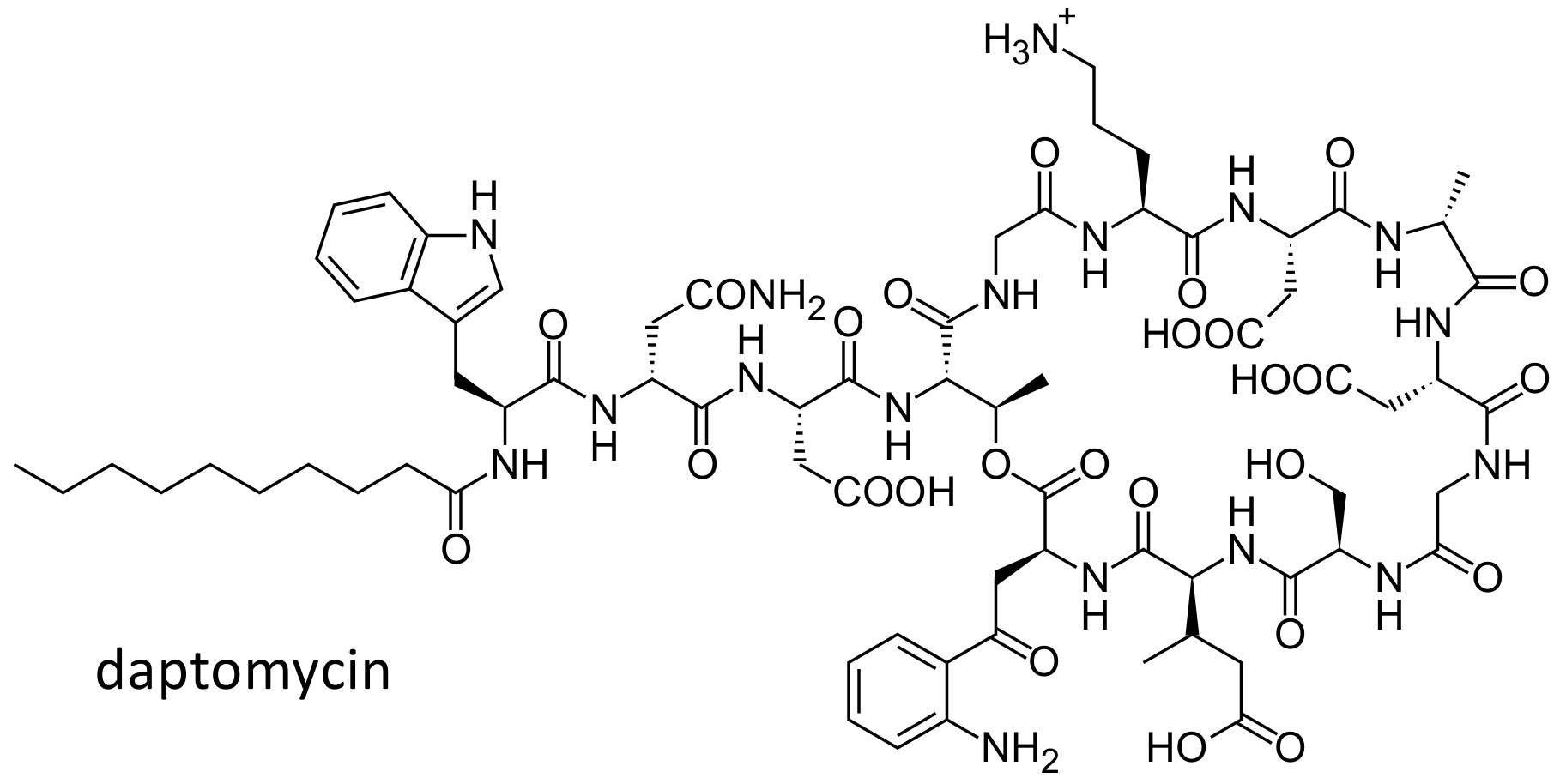
2.6. Daptomycin
2.7. Linezolid
2.8. Minocycline
2.9. Clindamycin
3. Conclusions
Funding
Conflicts of Interest
Abbreviations
| MHC | Major histocompatibility complex |
| HLA | Human leukocyte antigen |
| DILI | Drug induced liver injury |
| OR | Odds ratios |
| CI | Confidence index |
| GWA | Genome wide association |
| SNP | Single nucleotide polymorphism |
| MRSA | Methicillin resistant Staphylococcus aureus |
| MRSE | Methicillin resistant Staphylococcus epidermidis |
| PXR | Pregnane X receptor |
| OAT | Organic anion transporter |
| MRP | Multidrug resistant-associated protein |
| ABC | ATP binding cassette |
| OATP | Organic anion transporter polypeptide |
| SLCO1A2 | Solute carrier organic anion transporter family member 1A2 |
| CYP | Cytochrome P450 |
| UGT | Uridine 5′ diphosphate glucuronosyltransferase |
| AUC0-24 | Area under the concentration-time curve from 0 to 24 h |
| DRESS | Drug reaction with eosinophilia and systemic symptoms |
References
- Lesch, J. The First Miracle Drugs: How the sulfa Drugs Transformed Medicine; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Lax, E. The Mold in Dr. Florey’s Coat: The Story of Penicillin Miracle; Henry Holt: New York, NY, USA, 2004. [Google Scholar]
- Attar, M.; Lee, V.H. Pharmacogenomic considerations in drug delivery. Pharmacogenomics 2003, 4, 443–461. [Google Scholar] [CrossRef] [PubMed]
- Ho, R.H.; Kim, R.B. Transporters and drug therapy: Implications for drug disposition and disease. Clin. Pharmacol. Ther. 2005, 78, 260–277. [Google Scholar] [CrossRef] [PubMed]
- Pavlos, R.; Mallal, S.; Phillips, E. HLA and pharmacogenetics of drug hypersensitivity. Pharmacogenomics 2012, 13, 1285–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, A.; Mondino, C.; Viola, M.; Montuschi, P. Immediate allergic reactions to beta-lactams: diagnosis and therapy. Int. J. Immunopathol Pharm. 2003, 16, 19–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerns, D.; Shira, J.E.; Go, S.; Summers, R.J.; Schwab, J.A.; Plunket, D.C. Ampicillin rash in children. Relationship to penicillin allergy and infectious mononucleosis. Am. J. Dis. Child. 1973, 125, 187–190. [Google Scholar] [CrossRef]
- Andrade, R.J.; Lucena, M.I.; Fernandez, M.C.; Pelaez, G.; Pachkoria, K.; Garcia-Ruiz, E.; Garcia-Munoz, B.; Gonzalez-Grande, R.; Pizarro, A.; Duran, J.A.; et al. Drug-induced liver injury: an analysis of 461 incidences submitted to the Spanish registry over a 10-year period. Gastroenterology 2005, 129, 512–521. [Google Scholar] [CrossRef]
- De Abajo, F.J.; Montero, D.; Madurga, M.; Rodriguez, L.A.G. Acute and clinically relevant drug-induced liver injury: A population based case-control study. Br. J. Clin. Pharmacol. 2004, 58, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Chalasani, N.; Fontana, R.J.; Bonkovsky, H.L.; Watkins, P.B.; Davern, T.; Serrano, J.; Yang, H.; Rochon, J.; Drug Induced Liver Injury Network (DILIN). Causes, clinical features, and outcomes from a prospective study of drug-induced liver injury in the United States. Gastroenterology 2008, 135, 1924–1934. [Google Scholar] [CrossRef] [Green Version]
- Bjornsson, E.S.; Bergmann, O.M.; Bjornsson, H.K.; Kvaran, R.B.; Olafsson, S. Incidence, presentation, and outcomes in patients with drug-induced liver injury in the general population of Iceland. Gastroenterology 2013, 144, 1419–1425. [Google Scholar] [CrossRef]
- Andrade, R.J.; Lucena, M.I.; Kaplowitz, N.; Garcia-Munoz, B.; Borraz, Y.; Pachkoria, K.; Garcia-Cortes, M.; Fernandez, M.C.; Pelaez, G.; Rodrigo, L.; et al. Outcome of acute idiosyncratic drug-induced liver injury: Long-term follow-up in a hepatotoxicity registry. Hepatology 2006, 44, 1581–1588. [Google Scholar] [CrossRef]
- Lucena, M.I.; Andrade, R.J.; Fernandez, M.C.; Pachkoria, K.; Pelaez, G.; Duran, J.A.; Villar, M.; Rodrigo, L.; Romero-Gomez, M.; Planas, R.; et al. Determinants of the clinical expression of amoxicillin-clavulanate hepatotoxicity: a prospective series from Spain. Hepatology 2006, 44, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, L.A.G.; Stricker, B.H.; Zimmerman, H.J. Risk of acute liver injury associated with the combination of amoxicillin and clavulanic acid. Arch. Intern. Med. 1996, 156, 1327–1332. [Google Scholar] [CrossRef] [PubMed]
- Fontana, R.J.; Shakil, A.O.; Greenson, J.K.; Boyd, I.; Lee, W.M. Acute liver failure due to amoxicillin and amoxicillin/clavulanate. Dig. Dis. Sci. 2005, 50, 1785–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hautekeete, M.L.; Horsmans, Y.; van Waeyenberge, C.; Demanet, C.; Henrion, J.; Verbist, L.; Brenard, R.; Sempoux, C.; Michielsen, P.P.; Yap, P.S.; et al. HLA association of amoxicillin-clavulanate--induced hepatitis. Gastroenterology 1999, 117, 1181–1186. [Google Scholar] [CrossRef]
- O’Donohue, J.; Oien, K.A.; Donaldson, P.; Underhill, J.; Clare, M.; MacSween, R.N.; Mills, P.R. Co-amoxiclav jaundice: Clinical and histological features and HLA class II association. Gut 2000, 47, 717–720. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, P.T.; Daly, A.K.; Henderson, J.; Graham, J.; Pirmohamed, M.; Bernal, W.; Day, C.P.; Aithal, G.P. Human leucocyte antigen class II genotype in susceptibility and resistance to co-amoxiclav-induced liver injury. J. Hepatol. 2010, 53, 1049–1053. [Google Scholar] [CrossRef]
- Andrade, R.J.; Lucena, M.I.; Alonso, A.; Garcia-Cortes, M.; Garcia-Ruiz, E.; Benitez, R.; Fernandez, M.C.; Pelaez, G.; Romero, M.; Corpas, R.; et al. HLA class II genotype influences the type of liver injury in drug-induced idiosyncratic liver disease. Hepatology 2004, 39, 1603–1612. [Google Scholar] [CrossRef]
- Lucena, M.I.; Molokhia, M.; Shen, Y.; Urban, T.J.; Aithal, G.P.; Andrade, R.J.; Day, C.P.; Ruiz-Cabello, F.; Donaldson, P.T.; Stephens, C.; et al. Susceptibility to amoxicillin-clavulanate-induced liver injury is influenced by multiple HLA class I and II alleles. Gastroenterology 2011, 141, 338–347. [Google Scholar] [CrossRef]
- Stephens, C.; Lopez-Nevot, M.A.; Ruiz-Cabello, F.; Ulzurrun, E.; Soriano, G.; Romero-Gomez, M.; Moreno-Casares, A.; Lucena, M.I.; Andrade, R.J. HLA alleles influence the clinical signature of amoxicillin-clavulanate hepatotoxicity. PLoS ONE 2013, 8, e68111. [Google Scholar] [CrossRef]
- Cirulli, E.T.; Nicoletti, P.; Abramson, K.; Andrade, R.J.; Bjornsson, E.S.; Chalasani, N.; Fontana, R.J.; Hallberg, P.; Li, Y.J.; Lucena, M.I.; et al. A missense variant in PTPN22 is a risk factor for drug-induced liver injury. Gastroenterology 2019, 156, 1707–1716. [Google Scholar] [CrossRef] [Green Version]
- Peacock, S.J.; Paterson, G.K. Mechanisms of methicillin resistance in staphylococcus aureus. Annu. Rev. Biochem. 2015, 84, 577–601. [Google Scholar] [CrossRef] [PubMed]
- Daly, A.K.; Donaldson, P.T.; Bhatnagar, P.; Shen, Y.; Pe’er, I.; Floratos, A.; Daly, M.J.; Goldstein, D.B.; John, S.; Nelson, M.R.; et al. HLA-B*5701 genotype is a major determinant of drug-induced liver injury due to flucloxacillin. Nat. Genet. 2009, 41, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, H.A.; Woloski, B.M.; Hellman, M.; Jamieson, J.C. Studies on the effect of inflammation on rat liver and serum sialyltransferase. Evidence that inflammation causes release of Gal beta 1 leads to 4GlcNAc alpha 2 leads to 6 sialyltransferase from liver. J. Biol. Chem. 1983, 258, 11505–11509. [Google Scholar] [PubMed]
- Nicoletti, P.; Aithal, G.P.; Chamberlain, T.C.; Coulthard, S.; Alshabeeb, M.; Grove, J.I.; Andrade, R.J.; Bjornsson, E.; Dillon, J.F.; Hallberg, P.; et al. Drug-induced liver injury due to flucloxacillin: Relevance of multiple human leukocyte antigen alleles. Clin. Pharmacol. Ther. 2019, 106, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Andrews, E.; Armstrong, M.; Tugwood, J.; Swan, D.; Glaves, P.; Pirmohamed, M.; Aithal, G.P.; Wright, M.C.; Day, C.P.; Daly, A.K. A role for the pregnane X receptor in flucloxacillin-induced liver injury. Hepatology 2010, 51, 1656–1664. [Google Scholar] [CrossRef]
- Ings, R.M.; Reeves, D.S.; White, L.O.; Bax, R.P.; Bywater, M.J.; Holt, H.A. The human pharmacokinetics of cefotaxime and its metabolites and the role of renal tubular secretion on their elimination. J. Pharm. Biopharm. 1985, 13, 121–142. [Google Scholar] [CrossRef]
- Ueo, H.; Motohashi, H.; Katsura, T.; Inui, K. Human organic anion transporter hOAT3 is a potent transporter of cephalosporin antibiotics, in comparison with hOAT1. Biochem. Pharmacol. 2005, 70, 1104–1113. [Google Scholar] [CrossRef]
- Uwai, Y.; Saito, H.; Inui, K. Rat renal organic anion transporter rOAT1 mediates transport of urinary-excreted cephalosporins, but not of biliary-excreted cefoperazone. Drug Metab. Pharmacokinet. 2002, 17, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Jariyawat, S.; Sekine, T.; Takeda, M.; Apiwattanakul, N.; Kanai, Y.; Sophasan, S.; Endou, H. The interaction and transport of beta-lactam antibiotics with the cloned rat renal organic anion transporter 1. J. Pharmacol. Exp. Ther. 1999, 290, 672–677. [Google Scholar]
- Fujita, T.; Brown, C.; Carlson, E.J.; Taylor, T.; de la Cruz, M.; Johns, S.J.; Stryke, D.; Kawamoto, M.; Fujita, K.; Castro, R.; et al. Functional analysis of polymorphisms in the organic anion transporter, SLC22A6 (OAT1). Pharm. Genom. 2005, 15, 201–209. [Google Scholar] [CrossRef]
- 1000 Genome Project Consortium; Abecasis, G.R.; Auton, A.; Brooks, L.D.; DePristo, M.A.; Durbin, R.M.; Handsaker, R.E.; Kang, H.M.; Marth, G.T.; McVean, G.A. An integrated map of genetic variation from 1092 human genomes. Nature 2012, 491, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdman, A.R.; Mangravite, L.M.; Urban, T.J.; Lagpacan, L.L.; Castro, R.A.; de la Cruz, M.; Chan, W.; Huang, C.C.; Johns, S.J.; Kawamoto, M.; et al. The human organic anion transporter 3 (OAT3; SLC22A8): Genetic variation and functional genomics. Am. J. Physiol. Renal. Physiol. 2006, 290, F905–F912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yee, S.W.; Nguyen, A.N.; Brown, C.; Savic, R.M.; Zhang, Y.; Castro, R.A.; Cropp, C.D.; Choi, J.H.; Singh, D.; Tahara, H.; et al. Reduced renal clearance of cefotaxime in asians with a low-frequency polymorphism of OAT3 (SLC22A8). J. Pharm. Sci. 2013, 102, 3451–3457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Takahara, S.; Kato, S.; Kubo, Y.; Sai, Y.; Tamai, I.; Yabuuchi, H.; Tsuji, A. Involvement of multidrug resistance-associated protein 2 (Abcc2) in molecular weight-dependent biliary excretion of beta-lactam antibiotics. Drug Metab. Dispos. 2008, 36, 1088–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehn, L.M. ABC efflux transporters at blood-central nervous system barriers and their implications for treating spinal cord disorders. Neural Regen. Res. 2020, 15, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Hartz, A.M.; Bauer, B. ABC transporters in the CNS—An inventory. Curr. Pharm. Biotechnol. 2011, 12, 656–673. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Glaeser, H.; Smith, L.H.; Roberts, R.L.; Moeckel, G.W.; Gervasini, G.; Leake, B.F.; Kim, R.B. Polymorphisms in human organic anion-transporting polypeptide 1A2 (OATP1A2): implications for altered drug disposition and central nervous system drug entry. J. Biol. Chem. 2005, 280, 9610–9617. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.; Hagenbuch, B.; Kullak-Ublick, G.A.; Benke, D.; Aguzzi, A.; Meier, P.J. Organic anion-transporting polypeptides mediate transport of opioid peptides across blood-brain barrier. J. Pharmacol. Exp. Ther. 2000, 294, 73–79. [Google Scholar]
- Allegra, S.; Cardellino, C.S.; Fatiguso, G.; Cusato, J.; de Nicolo, A.; Avataneo, V.; Bonora, S.; D’Avolio, A.; di Perri, G.; Calcagno, A. Effect of ABCC2 and ABCG2 gene polymorphisms and CSF-to-serum albumin ratio on ceftriaxone plasma and cerebrospinal fluid concentrations. J. Clin. Pharmacol. 2018, 58, 1550–1556. [Google Scholar] [CrossRef]
- Neftel, K.A.; Hauser, S.P.; Muller, M.R. Inhibition of granulopoiesis in vivo and in vitro by beta-lactam antibiotics. J. Infect. Dis. 1985, 152, 90–98. [Google Scholar] [CrossRef]
- Andres, E.; Maloisel, F. Idiosyncratic drug-induced agranulocytosis or acute neutropenia. Curr. Opin. Hematol. 2008, 15, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.; Fukuda, T.; Hahn, D.; Mizuno, T.; Frenck, R.W., Jr.; Vinks, A.A. Pharmacokinetics and pharmacogenomics of beta-lactam-induced neutropenia. Pharmacogenomics 2016, 17, 547–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russel, F.G.; Koenderink, J.B.; Masereeuw, R. Multidrug resistance protein 4 (MRP4/ABCC4): a versatile efflux transporter for drugs and signalling molecules. Trends Pharmacol. Sci. 2008, 29, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Ci, L.; Kusuhara, H.; Adachi, M.; Schuetz, J.D.; Takeuchi, K.; Sugiyama, Y. Involvement of MRP4 (ABCC4) in the luminal efflux of ceftizoxime and cefazolin in the kidney. Mol. Pharmacol. 2007, 71, 1591–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanwert, A.L.; Bailey, R.M.; Sweet, D.H. Organic anion transporter 3 (Oat3/Slc22a8) knockout mice exhibit altered clearance and distribution of penicillin G. Am. J. Physiol. Renal. Physiol. 2007, 293, F1332–F1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mc, G.J.; Bunch, R.L.; Anderson, R.C.; Boaz, H.E.; Flynn, E.H.; Powell, H.M.; Smith, J.W. Ilotycin, a new antibiotic. Antibiot. Chemother. (Northfield) 1952, 2, 281–283. [Google Scholar]
- Washington, J.A., 2nd; Wilson, W.R. Erythromycin: A microbial and clinical perspective after 30 years of clinical use (2). Mayo Clin. Proc. 1985, 60, 271–278. [Google Scholar] [CrossRef] [Green Version]
- Washington, J.A., 2nd; Wilson, W.R. Erythromycin: A microbial and clinical perspective after 30 years of clinical use (1). Mayo Clin. Proc. 1985, 60, 189–203. [Google Scholar] [CrossRef] [Green Version]
- Zuckerman, J.M.; Qamar, F.; Bono, B.R. Review of macrolides (azithromycin, clarithromycin), ketolids (telithromycin) and glycylcyclines (tigecycline). Med. Clin. North Am. 2011, 95, 761–791. [Google Scholar] [CrossRef]
- Franke, R.M.; Lancaster, C.S.; Peer, C.J.; Gibson, A.A.; Kosloske, A.M.; Orwick, S.J.; Mathijssen, R.H.; Figg, W.D.; Baker, S.D.; Sparreboom, A. Effect of ABCC2 (MRP2) transport function on erythromycin metabolism. Clin. Pharmacol. Ther. 2011, 89, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, C.S.; Bruun, G.H.; Peer, C.J.; Mikkelsen, T.S.; Corydon, T.J.; Gibson, A.A.; Hu, S.; Orwick, S.J.; Mathijssen, R.H.; Figg, W.D.; et al. OATP1B1 polymorphism as a determinant of erythromycin disposition. Clin. Pharmacol. Ther. 2012, 92, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Sugie, M.; Asakura, E.; Zhao, Y.L.; Torita, S.; Nadai, M.; Baba, K.; Kitaichi, K.; Takagi, K.; Takagi, K.; Hasegawa, T. Possible involvement of the drug transporters P glycoprotein and multidrug resistance-associated protein Mrp2 in disposition of azithromycin. Antimicrob. Agents Chemother. 2004, 48, 809–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.J.; Zhao, L.M.; Qiu, F.; Sun, Y.X.; Li-Ling, J. Influence of ABCB1 gene polymorphisms on the pharmacokinetics of azithromycin among healthy Chinese Han ethnic subjects. Pharmacol. Rep. 2009, 61, 843–850. [Google Scholar] [CrossRef]
- Becker, B.; Cooper, M.A. Aminoglycoside antibiotics in the 21st century. ACS Chem. Biol. 2013, 8, 105–115. [Google Scholar] [CrossRef]
- Guthrie, O.W. Aminoglycoside induced ototoxicity. Toxicology 2008, 249, 91–96. [Google Scholar] [CrossRef]
- Selimoglu, E. Aminoglycoside-induced ototoxicity. Curr. Pharm. Des. 2007, 13, 119–126. [Google Scholar] [CrossRef]
- Guan, M.X. Mitochondrial 12S rRNA mutations associated with aminoglycoside ototoxicity. Mitochondrion 2011, 11, 237–245. [Google Scholar] [CrossRef]
- Prezant, T.R.; Agapian, J.V.; Bohlman, M.C.; Bu, X.; Oztas, S.; Qiu, W.Q.; Arnos, K.S.; Cortopassi, G.A.; Jaber, L.; Rotter, J.I.; et al. Mitochondrial ribosomal RNA mutation associated with both antibiotic-induced and non-syndromic deafness. Nat. Genet. 1993, 4, 289–294. [Google Scholar] [CrossRef]
- Estivill, X.; Govea, N.; Barcelo, E.; Badenas, C.; Romero, E.; Moral, L.; Scozzri, R.; D’Urbano, L.; Zeviani, M.; Torroni, A. Familial progressive sensorineural deafness is mainly due to the mtDNA A1555G mutation and is enhanced by treatment of aminoglycosides. Am. J. Hum. Genet. 1998, 62, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Bitner-Glindzicz, M.; Pembrey, M.; Duncan, A.; Heron, J.; Ring, S.M.; Hall, A.; Rahman, S. Prevalence of mitochondrial 1555A-->G mutation in European children. N. Engl. J. Med. 2009, 360, 640–642. [Google Scholar] [CrossRef] [Green Version]
- Vandebona, H.; Mitchell, P.; Manwaring, N.; Griffiths, K.; Gopinath, B.; Wang, J.J.; Sue, C.M. Prevalence of mitochondrial 1555A-->G mutation in adults of European descent. N. Engl. J. Med. 2009, 360, 642–644. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, R.; Wang, Q.; Yan, Q.; Deng, J.H.; Han, D.; Bai, Y.; Young, W.Y.; Guan, M.X. Maternally inherited aminoglycoside-induced and nonsyndromic deafness is associated with the novel C1494T mutation in the mitochondrial 12S rRNA gene in a large Chinese family. Am. J. Hum. Genet. 2004, 74, 139–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA Reinforces Safety Information about Serious Low Blood Sugar Levels and Mental Health Side Effects With Fluoroquinolone Antibiotics. Requires Label Changes. Available online: https://www.fda.gov/drugs/drug-safety-and-availability/fda-reinforces-safety-information-about-serious-low-blood-sugar-levels-and-mental-health-side (accessed on 27 July 2020).
- Disabling and Potentially Permanent Side Effects Lead to Suspension or Restrictions of Quinolone and Fluoroquinolone Antibiotics. Available online: https://www.ema.europa.eu/en/documents/referral/quinolone-fluoroquinolone-article-31-referral-disabling-potentially-permanent-side-effects-lead_en.pdf (accessed on 27 July 2020).
- Mehlhorn, A.J.; Brown, D.A. Safety concerns with fluoroquinolones. Ann. Pharmacother. 2007, 41, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Leslie, E.M.; Deeley, R.G.; Cole, S.P. Multidrug resistance proteins: role of P-glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in tissue defense. Toxicol. Appl. Pharmacol. 2005, 204, 216–237. [Google Scholar] [CrossRef]
- Alvarez, A.I.; Perez, M.; Prieto, J.G.; Molina, A.J.; Real, R.; Merino, G. Fluoroquinolone efflux mediated by ABC transporters. J. Pharm. Sci. 2008, 97, 3483–3493. [Google Scholar] [CrossRef]
- Ooie, T.; Terasaki, T.; Suzuki, H.; Sugiyama, Y. Quantitative brain microdialysis study on the mechanism of quinolones distribution in the central nervous system. Drug Metab. Dispos. 1997, 25, 784–789. [Google Scholar]
- Ooie, T.; Terasaki, T.; Suzuki, H.; Sugiyama, Y. Kinetic evidence for active efflux transport across the blood-brain barrier of quinolone antibiotics. J. Pharmacol. Exp. Ther. 1997, 283, 293–304. [Google Scholar]
- De Lange, E.C.; Marchand, S.; van den Berg, D.; van der Sandt, I.C.; de Boer, A.G.; Delon, A.; Bouquet, S.; Couet, W. In vitro and in vivo investigations on fluoroquinolones; effects of the P-glycoprotein efflux transporter on brain distribution of sparfloxacin. Eur. J. Pharm. Sci. 2000, 12, 85–93. [Google Scholar] [CrossRef]
- Gervasoni, C.; Cattaneo, D.; Falvella, F.S.; Vitiello, P.; Cheli, S.; Milazzo, L.; Clementi, E.; Riva, A. Levofloxacin-induced seizures in a patient without predisposing risk factors: the impact of pharmacogenetics. Eur. J. Clin. Pharmacol. 2013, 69, 1611–1613. [Google Scholar] [CrossRef]
- Kroetz, D.L.; Pauli-Magnus, C.; Hodges, L.M.; Huang, C.C.; Kawamoto, M.; Johns, S.J.; Stryke, D.; Ferrin, T.E.; DeYoung, J.; Taylor, T.; et al. Sequence diversity and haplotype structure in the human ABCB1 (MDR1, multidrug resistance transporter) gene. Pharmacogenetics 2003, 13, 481–494. [Google Scholar] [CrossRef]
- Franke, R.M.; Scherkenbach, L.A.; Sparreboom, A. Pharmacogenetics of the organic anion transporting polypeptide 1A2. Pharmacogenomics 2009, 10, 339–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, T.; Takahashi, K.; Ohtsu, N.; Oguma, T.; Ohnishi, T.; Atsumi, R.; Tamai, I. Identification of influx transporter for the quinolone antibacterial agent levofloxacin. Mol. Pharm. 2007, 4, 85–94. [Google Scholar] [CrossRef] [PubMed]
- MacGowan, A.P. Moxifloxacin (Bay 12-8039): a new methoxy quinolone antibacterial. Expert Opin. Investig. Drugs 1999, 8, 181–199. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Tuberculosis Report. 2019. Available online: https://www.who.int/tb/publications/global_report/en/ (accessed on 27 July 2020).
- Naidoo, A.; Naidoo, K.; McIlleron, H.; Essack, S.; Padayatchi, N. A review of moxifloxacin for the treatment of drug-susceptible tuberculosis. J. Clin. Pharmacol. 2017, 57, 1369–1386. [Google Scholar] [CrossRef] [PubMed]
- Pranger, A.D.; Kosterink, J.G.; van Altena, R.; Aarnoutse, R.E.; van der Werf, T.S.; Uges, D.R.; Alffenaar, J.W. Limited-sampling strategies for therapeutic drug monitoring of moxifloxacin in patients with tuberculosis. Ther. Drug Monit. 2011, 33, 350–354. [Google Scholar] [CrossRef] [Green Version]
- Barbarino, J.M.; Haidar, C.E.; Klein, T.E.; Altman, R.B. PharmGKB summary: Very important pharmacogene information for UGT1A1. Pharmacogenet. Genomics 2014, 24, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Stingl, J.C.; Bartels, H.; Viviani, R.; Lehmann, M.L.; Brockmoller, J. Relevance of UDP-glucuronosyltransferase polymorphisms for drug dosing: A quantitative systematic review. Pharmacol. Ther. 2014, 141, 92–116. [Google Scholar] [CrossRef]
- Naidoo, A.; Ramsuran, V.; Chirehwa, M.; Denti, P.; McIlleron, H.; Naidoo, K.; Yende-Zuma, N.; Singh, R.; Ngcapu, S.; Chaudhry, M.; et al. Effect of genetic variation in UGT1A and ABCB1 on moxifloxacin pharmacokinetics in South African patients with tuberculosis. Pharmacogenomics 2018, 19, 17–29. [Google Scholar] [CrossRef]
- Weiner, M.; Burman, W.; Luo, C.C.; Peloquin, C.A.; Engle, M.; Goldberg, S.; Agarwal, V.; Vernon, A. Effects of rifampin and multidrug resistance gene polymorphism on concentrations of moxifloxacin. Antimicrob. Agents Chemother. 2007, 51, 2861–2866. [Google Scholar] [CrossRef] [Green Version]
- Weiner, M.; Gelfond, J.; Johnson-Pais, T.L.; Engle, M.; Peloquin, C.A.; Johnson, J.L.; Sizemore, E.E.; Mac Kenzie, W.R. Elevated plasma moxifloxacin concentrations and SLCO1B1 g-11187G>A polymorphism in adults with pulmonary tuberculosis. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.; Hnatkova, K.; Schmidt, A.; Smetana, P. Electrocardiographic QTc changes due to moxifloxacin infusion. J. Clin. Pharmacol. 2009, 49, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Levine, D.P. Vancomycin: A history. Clin. Infect. Dis. 2006, 42 (Suppl 1), S5–S12. [Google Scholar] [CrossRef] [PubMed]
- Konvinse, K.C.; Trubiano, J.A.; Pavlos, R.; James, I.; Shaffer, C.M.; Bejan, C.A.; Schutte, R.J.; Ostrov, D.A.; Pilkinton, M.A.; Rosenbach, M.; et al. HLA-A*32:01 is strongly associated with vancomycin-induced drug reaction with eosinophilia and systemic symptoms. J. Allergy Clin. Immunol. 2019, 144, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Driest, S.L.; McGregor, T.L.; Edwards, D.R.V.; Saville, B.R.; Kitchner, T.E.; Hebbring, S.J.; Brilliant, M.; Jouni, H.; Kullo, I.J.; Creech, C.B.; et al. Genome-wide association study of serum creatinine levels during vancomycin therapy. PLoS ONE 2015, 10, e0127791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoemaker, D.M.; Simou, J.; Roland, W.E. A review of daptomycin for injection (Cubicin) in the treatment of complicated skin and skin structure infections. Ther. Clin. Risk Manag. 2006, 2, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Lemaire, S.; van Bambeke, F.; Mingeot-Leclercq, M.P.; Tulkens, P.M. Modulation of the cellular accumulation and intracellular activity of daptomycin towards phagocytized Staphylococcus aureus by the P-glycoprotein (MDR1) efflux transporter in human THP-1 macrophages and madin-darby canine kidney cells. Antimicrob. Agents Chemother. 2007, 51, 2748–2757. [Google Scholar] [CrossRef] [Green Version]
- Baietto, L.; D’Avolio, A.; Cusato, J.; Pace, S.; Calcagno, A.; Motta, I.; Corcione, S.; di Perri, G.; de Rosa, F.G. Effect of SNPs in human ABCB1 on daptomycin pharmacokinetics in Caucasian patients. J. Antimicrob. Chemother. 2015, 70, 307–308. [Google Scholar] [CrossRef] [Green Version]
- Rivera, A.M.; Boucher, H.W. Current concepts in antimicrobial therapy against select gram-positive organisms: methicillin-resistant Staphylococcus aureus, penicillin-resistant pneumococci, and vancomycin-resistant enterococci. Mayo Clin. Proc. 2011, 86, 1230–1243. [Google Scholar] [CrossRef] [Green Version]
- Bolhuis, M.S.; Tiberi, S.; Sotgiu, G.; de Lorenzo, S.; Kosterink, J.G.; van der Werf, T.S.; Migliori, G.B.; Alffenaar, J.W. Linezolid tolerability in multidrug-resistant tuberculosis: A retrospective study. Eur. Respir. J. 2015, 46, 1205–1207. [Google Scholar] [CrossRef] [Green Version]
- Zoller, M.; Maier, B.; Hornuss, C.; Neugebauer, C.; Dobbeler, G.; Nagel, D.; Holdt, L.M.; Bruegel, M.; Weig, T.; Grabein, B.; et al. Variability of linezolid concentrations after standard dosing in critically ill patients: a prospective observational study. Crit. Care 2014, 18, R148. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Xie, J.; Wang, T.; Chen, L.; Zeng, X.; Sun, J.; Wang, X.; Dong, Y. Pharmacokinetic/pharmacodynamic evaluation of linezolid for the treatment of staphylococcal infections in critically ill patients. Int. J. Antimicrob. Agents 2016, 48, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Allegra, S.; di Paolo, A.; Cusato, J.; Fatiguso, G.; Arrigoni, E.; Danesi, R.; Corcione, S.; D’Avolio, A. A common mdr1 gene polymorphism is associated with changes in linezolid clearance. Ther. Drug Monit. 2018, 40, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Bjornsson, E.; Talwalkar, J.; Treeprasertsuk, S.; Kamath, P.S.; Takahashi, N.; Sanderson, S.; Neuhauser, M.; Lindor, K. Drug-induced autoimmune hepatitis: clinical characteristics and prognosis. Hepatology 2010, 51, 2040–2048. [Google Scholar] [CrossRef] [PubMed]
- Urban, T.J.; Nicoletti, P.; Chalasani, N.; Serrano, J.; Stolz, A.; Daly, A.K.; Aithal, G.P.; Dillon, J.; Navarro, V.; Odin, J.; et al. Minocycline hepatotoxicity: Clinical characterization and identification of HLA-B *35:02 as a risk factor. J. Hepatol. 2017, 67, 137–144. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, S.; Yang, F.; Zhang, L.; Alterovitz, G.; Zhu, H.; Xuan, J.; Yang, X.; Luo, H.; Mu, J.; et al. HLA-B*51:01 is strongly associated with clindamycin-related cutaneous adverse drug reactions. Pharm. J. 2017, 17, 501–505. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Polymorphism | Antibiotic | Effect | Reference |
|---|---|---|---|---|
| SLC22A8 | rs11568482 | Cefotaxime (Cephalosporins) | Lower clearance | [33] |
| ABCC2 | rs2273697 | Ceftriaxone (Cephalosporins) | Higher drug concentration in CSF | [41] |
| ABCG2 | rs13120400 | Reduction on drug CSF/plasma ratio | ||
| ABCC2 | rs717620 | Erythromycin (Macrolides) | Increase in drug metabolism | [52] |
| SLCO1B1 | rs4149056 | Reduction in drug metabolism | [53] | |
| ABCB1 | 2677TT/3435TT | Azithromycin (Macrolides) | Lower Cmax and higher Tmax | [55] |
| UGT1A | rs8175347 | Moxifloxacin (Fluoroquinolones) | Lower clearance | [83] |
| rs3755319 | Higher clearance | |||
| ABCB1 | rs2032582 | Higher clearance | [83] | |
| ABCB1 | rs1045642 | Higher Tmax | [84] | |
| SLCO1B1 | rs4149015 | Higher AUC0-24 and Cmax | [85] | |
| ABCB1 | 1236C > T rs1128503 2677G > T/A rs2032582 3435C > T rs1045642 | Daptomycin | Higher AUC0-24 | [92] |
| ABCB1 | rs1045642 | Linezolid | Lower clearance | [97] |
| Gene | HLA Association | Antibiotics | Effect | Reference |
|---|---|---|---|---|
| HLA-DRB1 | DRB1*15:01 | Amoxicillin clavulanate (Penicillins) | DILI | [16,17,18] |
| HLA-DQB1 | DQB1*06:14 | [19] | ||
| HLA-DQB1 | rs9274407 | [20] | ||
| HLA-DRA | rs3135388 | [20] | ||
| - | rs2523822 | [20] | ||
| HLA-DRB1-HLA-DQB1 | DRB1*15:01-DQB1*06:02 | [8,16,17,18] | ||
| HLA-A | A*30:02 | [21] | ||
| HLA-B | B*18:01 | [21] | ||
| HLA-B | B*57:01 | Flucloxacillin (Penicillins) | [24,26] | |
| HLA-B | B*57:03 | [26] | ||
| HLA-A | A*32:01 | Vancomycin | DRESS | [88] |
| HLA-B | B*35:02 | Minocycline | DILI | [99] |
| HLA-B | B*51:01 | Clindamycin | Cutaneous reaction | [100] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stocco, G.; Lucafò, M.; Decorti, G. Pharmacogenomics of Antibiotics. Int. J. Mol. Sci. 2020, 21, 5975. https://doi.org/10.3390/ijms21175975
Stocco G, Lucafò M, Decorti G. Pharmacogenomics of Antibiotics. International Journal of Molecular Sciences. 2020; 21(17):5975. https://doi.org/10.3390/ijms21175975
Chicago/Turabian StyleStocco, Gabriele, Marianna Lucafò, and Giuliana Decorti. 2020. "Pharmacogenomics of Antibiotics" International Journal of Molecular Sciences 21, no. 17: 5975. https://doi.org/10.3390/ijms21175975
APA StyleStocco, G., Lucafò, M., & Decorti, G. (2020). Pharmacogenomics of Antibiotics. International Journal of Molecular Sciences, 21(17), 5975. https://doi.org/10.3390/ijms21175975