A Translational Paradigm to Study the Effects of Uncontrollable Stress in Humans


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Study 1: Results of the Initial Paradigm
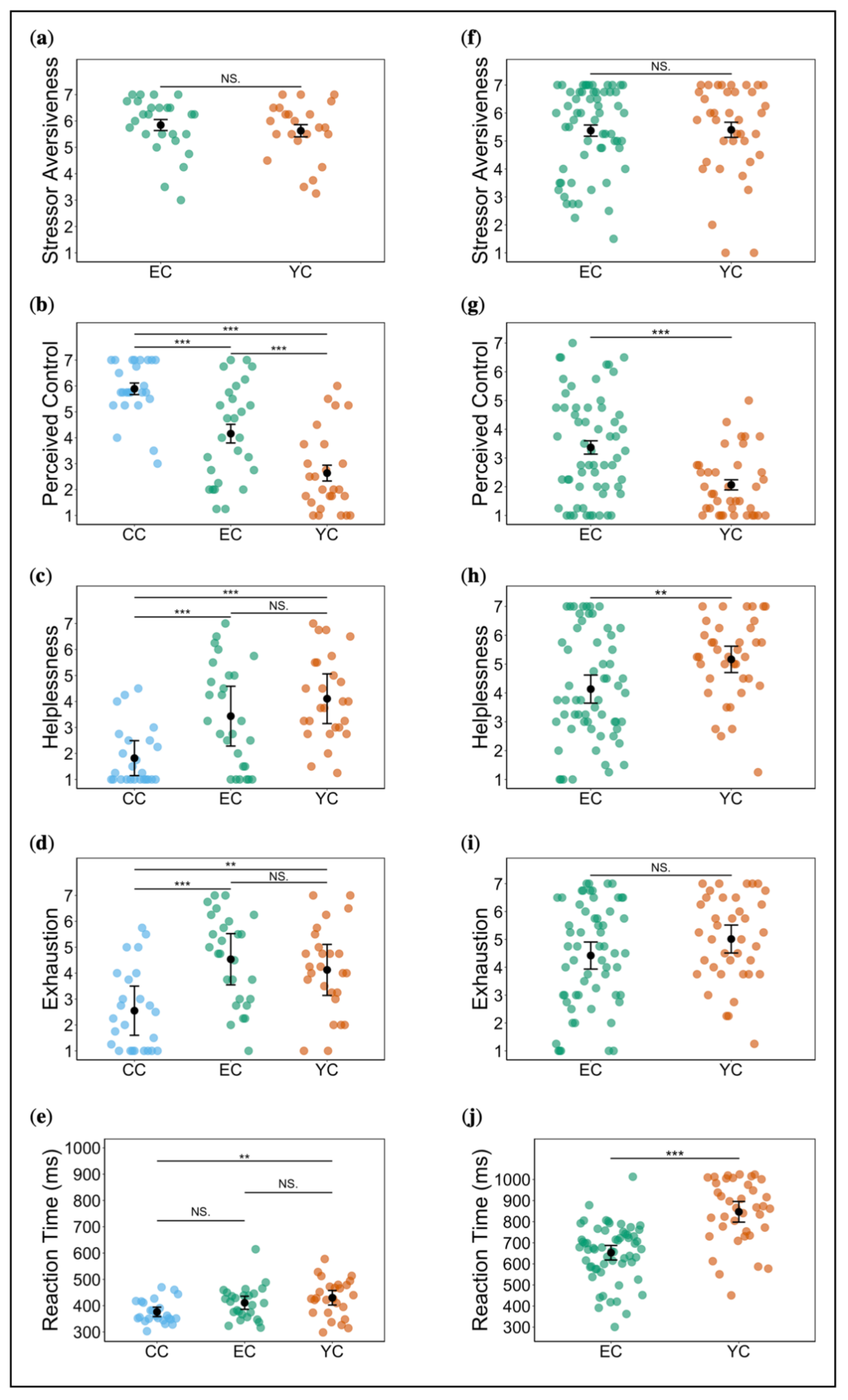
2.1.1. A Priori Group Comparisons and Manipulation Check
2.1.2. Ratings and RT during Acute Stress Exposure
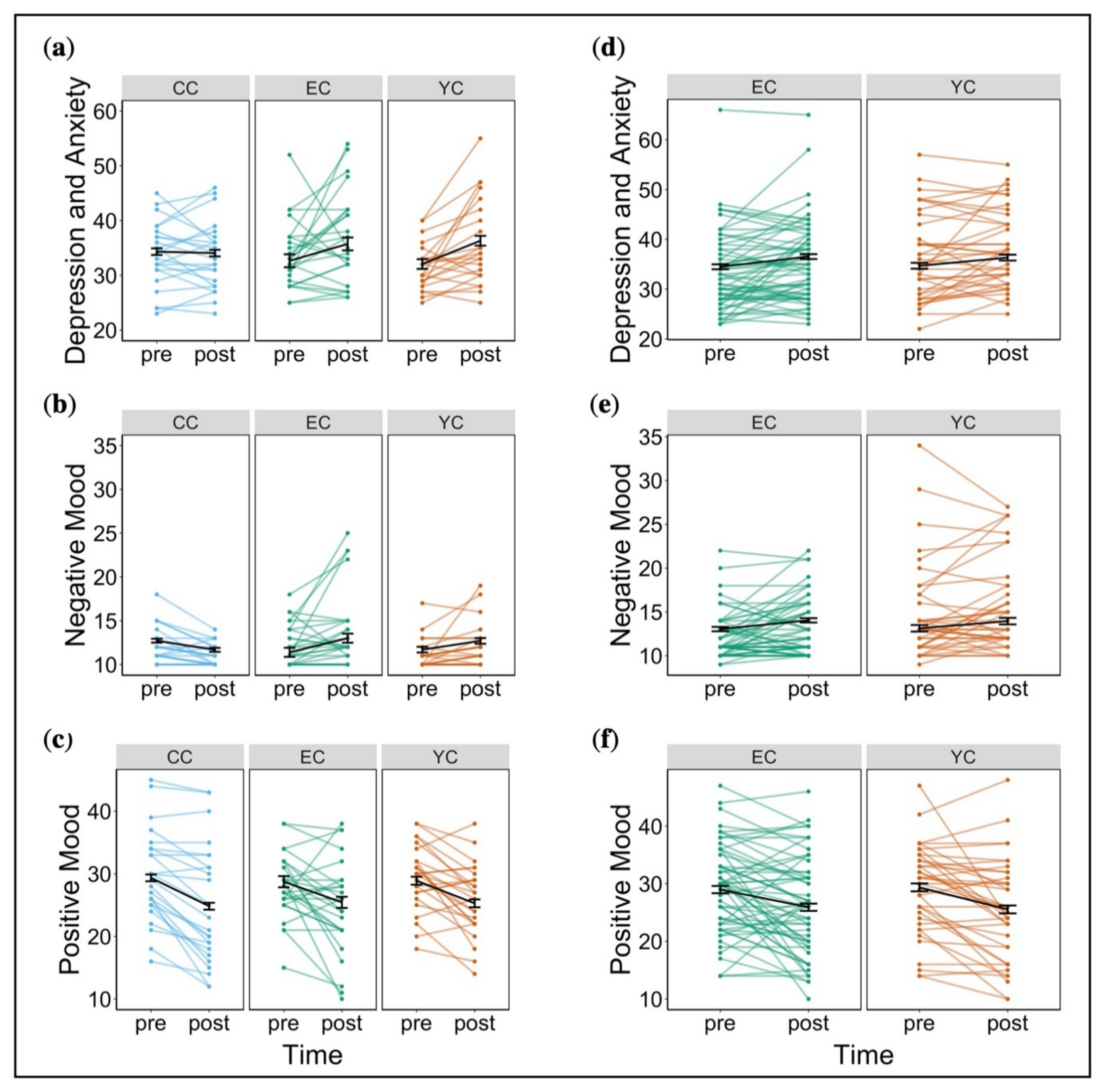
2.1.3. Affective State, Escape Behaviour, and Cognitive Performance during Subsequent (Post Stress) Assessment Changes in Affective State
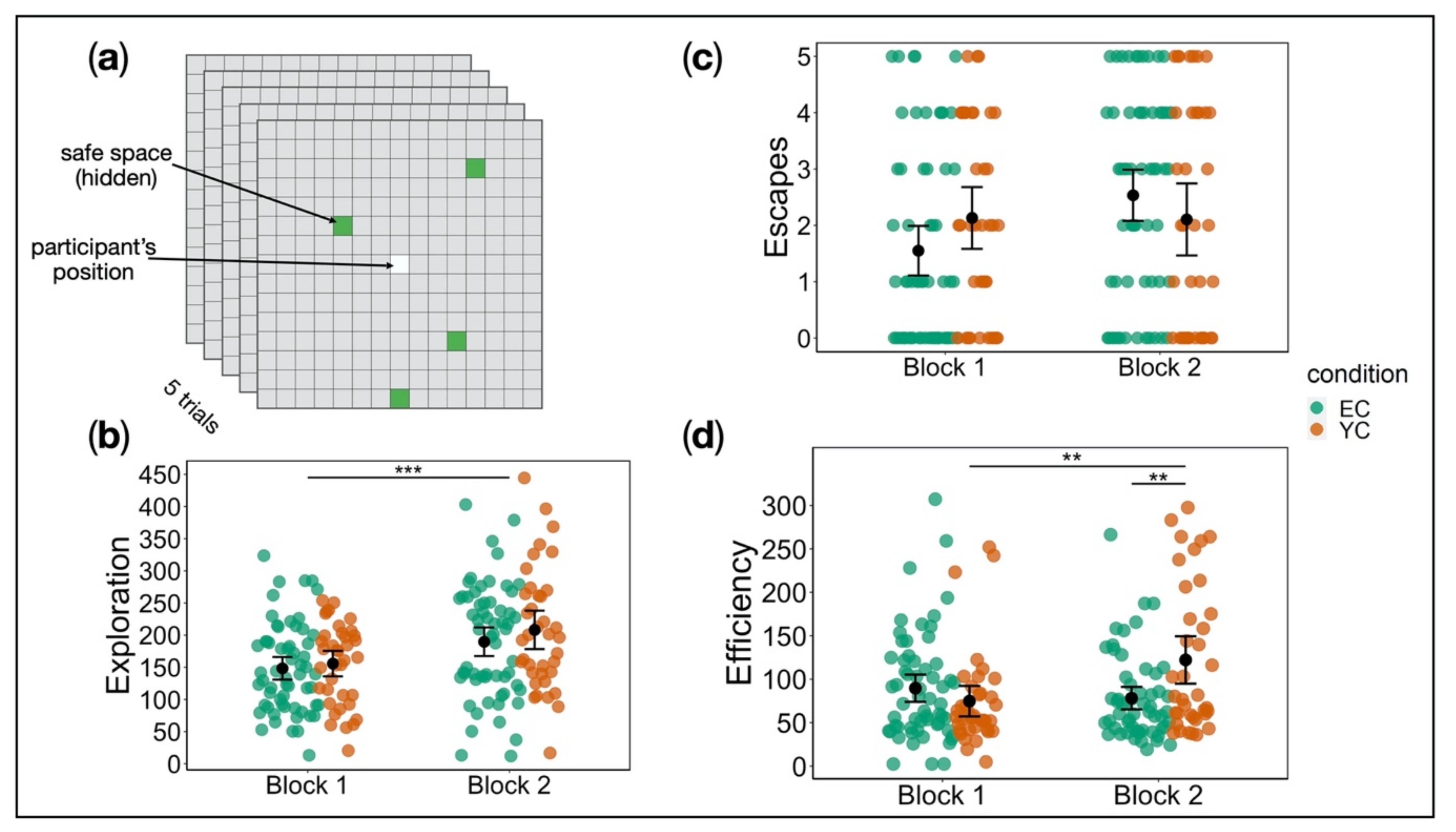
Escape Behaviour
Working Memory
2.2. Study 2: Results of the Improved Paradigm
2.2.1. A Priori Group Comparisons and Manipulation Check
2.2.2. Ratings and RT during Acute Stress Exposure
2.2.3. Affective State and Escape Behaviour during Subsequent (Post Stress) Assessment
Changes in Affective State
Escape Behaviour
3. Discussion
4. Materials and Methods
4.1. Study 1: Initial Paradigm
4.1.1. Participants
4.1.2. Procedure
4.1.3. Stress Induction
4.1.4. Measurements
Ratings and RT during Stress Exposure
Pre-Post Affect Questionnaires
Escape Behaviour Test
Working Memory
4.1.5. Statistical Analysis
4.2. Study 2: Improved Paradigm
4.2.1. Participants
4.2.2. Procedure
4.2.3. Stress Induction
4.2.4. Measurements
4.2.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| LH | Learned helplessness |
| EC | Escapable stress condition |
| YC | Yoked inescapable stress condition |
| CC | Control condition |
References
- Steptoe, A.; Poole, L. Control and Stress. In Stress: Concepts, Cognition, Emotion, and Behavior; Fink, G., Ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 1, pp. 73–80. [Google Scholar] [CrossRef]
- Maier, S.F.; Seligman, M.E.P. Learned helplessness at fifty: Insights from neuroscience. Psychol. Rev. 2016, 123, 349–367. [Google Scholar] [CrossRef]
- Pryce, C.R.; Azzinnari, D.; Spinelli, S.; Seifritz, E.; Tegethoff, M.; Meinlschmidt, G. Helplessness: A systematic translational review of theory and evidence for its relevance to understanding and treating depression. Pharmacol. Therapeut. 2011, 132, 242–267. [Google Scholar] [CrossRef]
- Seligman, M.E.; Maier, S.F. Failure to escape traumatic shock. J. Exp. Psychol. 1967, 74, 1–9. [Google Scholar] [CrossRef]
- Chourbaji, S.; Zacher, C.; Sanchis-Segura, C.; Dormann, C.; Vollmayr, B.; Gass, P. Learned helplessness: Validity and reliability of depressive-like states in mice. Brain Res. Protoc. 2005, 16, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Overmier, J.B.; Seligman, M.E. Effects of inescapable shock upon subsequent escape and avoidance responding. J. Comp. Physiol. Psychol. 1967, 63, 28–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollini, A.M.; Walker, E.F.; Hamann, S.; Kestler, L. The influence of perceived control and locus of control on the cortisol and subjective responses to stress. Biol. Psychol. 2004, 67, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Havranek, M.M.; Bolliger, B.; Roos, S.; Pryce, C.R.; Quednow, B.B.; Seifritz, E. Uncontrollable and unpredictable stress interacts with subclinical depression and anxiety scores in determining anxiety response. Stress 2016, 19, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Bräscher, A.-K.; Becker, S.; Hoeppli, M.-E.; Schweinhardt, P. Different brain circuitries mediating controllable and uncontrollable pain. J. Neurosci. 2016, 36, 5013–5025. [Google Scholar] [CrossRef]
- Henderson, R.K.; Snyder, H.R.; Gupta, T.; Banich, M.T. When does stress help or harm? The effects of stress controllability and subjective stress response on stroop performance. Front. Psychol. 2012, 3, 179:1–179:15. [Google Scholar] [CrossRef] [Green Version]
- Hartley, C.A.; Gorun, A.; Reddan, M.C.; Ramirez, F.; Phelps, E.A. Stressor controllability modulates fear extinction in humans. Neurobiol. Learn. Mem. 2014, 113, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Hiroto, D.S. Locus of control and learned helplessnss. J. Exp. Psychol. 1974, 102, 187–193. [Google Scholar] [CrossRef]
- Hiroto, D.S.; Seligman, M.E. Generality of learned helplessness in man. J. Pers. Soc. Psychol. 1975, 31, 311–327. [Google Scholar] [CrossRef]
- Thornton, J.W.; Jacobs, P.D. Learned helplessness in human subjects. J. Exp. Psychol. 1971, 87, 367–372. [Google Scholar] [CrossRef]
- Alloy, L.B.; Peterson, C.; Abramson, L.Y.; Seligman, M.E. Attributional style and the generality of learned helplessness. J. Pers. Soc. Psychol. 1984, 46, 681. [Google Scholar] [CrossRef] [PubMed]
- Bongard, S. Mental effort during active and passive coping: A dual-task analysis. Psychophysiology 1995, 32, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Diener, C.; Struve, M.; Balz, N.; Kuehner, C.; Flor, H. Exposure to uncontrollable stress and the postimperative negative variation (PINV): Prior control matters. Biol. Psychol. 2009, 80, 189–195. [Google Scholar] [CrossRef]
- Reznik, S.J.; Nusslock, R.; Pornpattananangkul, N.; Abramson, L.Y.; Coan, J.A.; Harmon-Jones, E. Laboratory-induced learned helplessness attenuates approach motivation as indexed by posterior versus frontal theta activity. Cogn. Affect. Behav. Neurosci. 2017, 17, 904–916. [Google Scholar] [CrossRef]
- Kerr, D.L.; McLaren, D.G.; Mathy, R.M.; Nitschke, J.B. Controllability modulates the anticipatory response in the human ventromedial prefrontal cortex. Front. Psychol. 2012, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Salomons, T.V.; Nusslock, R.; Detloff, A.; Johnstone, T.; Davidson, R.J. Neural emotion regulation circuitry underlying anxiolytic effects of perceived control over pain. J. Cogn. Neurosci. 2015, 27, 222–233. [Google Scholar] [CrossRef] [Green Version]
- Bauer, H.; Pripfl, J.; Lamm, C.; Prainsack, C.; Taylor, N. Functional neuroanatomy of learned helplessness. Neuroimage 2003, 20, 927–939. [Google Scholar] [CrossRef]
- Jacobs, N.; Myin-Germeys, I.; Derom, C.; Delespaul, P.; Van Os, J.; Nicolson, N.A. A momentary assessment study of the relationship between affective and adrenocortical stress responses in daily life. Biol. Psychol. 2007, 74, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Eckenrode, J. Impact of chronic and acute stressors on daily reports of mood. J. Pers Soc. Psychol. 1984, 46, 907. [Google Scholar] [CrossRef] [PubMed]
- Shields, G.S.; Sazma, M.A.; Yonelinas, A.P. The effects of acute stress on core executive functions: A meta-analysis and comparison with cortisol. Neurosci. Biobehav. Rev. 2016, 68, 651–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, A.R.; Raio, C.M.; Chiang, A.; Phelps, E.A.; Daw, N.D. Working-memory capacity protects model-based learning from stress. Proc. Natl. Acad. Sci. USA 2013, 110, 20941–20946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanke, N.; Schwabe, L. Subjective uncontrollability over aversive events reduces working memory performance and related large-scale network interactions. Cereb. Cortex 2020, 30, 3116–3129. [Google Scholar] [CrossRef]
- Kalisch, R.; Baker, D.G.; Basten, U.; Boks, M.P.; Bonanno, G.A.; Brummelman, E.; Chmitorz, A.; Fernàndez, G.; Fiebach, C.J.; Galatzer-Levy, I.; et al. The resilience framework as a strategy to combat stress-related disorders. Nat. Hum. Behav. 2017, 1, 784–790. [Google Scholar] [CrossRef] [Green Version]
- Chmitorz, A.; Kurth, K.; Mey, L.K.; Wenzel, M.; Lieb, K.; Tüscher, O.; Kubiak, T.; Kalisch, R. Assessment of microstressors in adults: questionnaire development and ecological validation of the mainz inventory of microstressors. JMIR Ment. Health 2020, 7, e14566:1–e14566:12. [Google Scholar] [CrossRef]
- Moscarello, J.M.; Hartley, C.A. Agency and the calibration of motivated behavior. Trends Cogn. Sci. 2017, 21, 725–735. [Google Scholar] [CrossRef]
- Huys, Q.J.; Dayan, P. A Bayesian formulation of behavioral control. Cognition 2009, 113, 314–328. [Google Scholar] [CrossRef]
- Guitart-Masip, M.; Huys, Q.J.; Fuentemilla, L.; Dayan, P.; Duzel, E.; Dolan, R.J. Go and no-go learning in reward and punishment: Interactions between affect and effect. Neuroimage 2012, 62, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Radenbach, C.; Reiter, A.M.; Engert, V.; Sjoerds, Z.; Villringer, A.; Heinze, H.J.; Deserno, L.; Schlagenhauf, F. The interaction of acute and chronic stress impairs model-based behavioral control. Psychoneuroendocrinology 2015, 53, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Voon, V.; Reiter, A.; Sebold, M.; Groman, S. Model-based control in dimensional psychiatry. Biol. Psychiatry 2017, 82, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.I.; Gagnon, S.A.; Wagner, A.D. Stress disrupts human hippocampal-prefrontal function during prospective spatial navigation and hinders flexible behavior. Curr. Biol. 2020, 30, 1821–1833. [Google Scholar] [CrossRef]
- Fineberg, N.A.; Chamberlain, S.R.; Goudriaan, A.E.; Stein, D.J.; Vanderschuren, L.J.; Gillan, C.M.; Shekar, S.; Gorwood, P.A.P.M.; Voon, V.; Morein-Zamir, S.; et al. New developments in human neurocognition: Clinical, genetic, and brain imaging correlates of impulsivity and compulsivity. CNS Spectr. 2014, 19, 69–89. [Google Scholar] [CrossRef]
- Ousdal, O.T.; Huys, Q.J.; Milde, A.M.; Craven, A.R.; Ersland, L.; Endestad, T.; Dolan, R.J. The impact of traumatic stress on pavlovian biases. Psychol. Med. 2018, 48, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schönfeld, P.; Preusser, F.; Margraf, J. Costs and benefits of self-efficacy: Differences of the stress response and clinical implications. Neurosci. Biobehav. Rev. 2017, 75, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Bzdok, D.; Meyer-Lindenberg, A. Machine learning for precision psychiatry: Opportunities and challenges. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Richter-Levin, G.; Stork, O.; Schmidt, M.V. Animal models of PTSD: A challenge to be met. Mol. Psychiatry 2019, 24, 1135–1156. [Google Scholar] [CrossRef] [Green Version]
- Hooijmans, C.R.; Ritskes-Hoitinga, M. Progress in using systematic reviews of animal studies to improve translational research. PLoS Med. 2013, 10, e1001482:1–e1001482:4. [Google Scholar] [CrossRef] [Green Version]
- Mak, I.W.; Evaniew, N.; Ghert, M. Lost in translation: Animal models and clinical trials in cancer treatment. Am. J. Transl. Res. 2014, 6, 114–118. [Google Scholar]
- Gururajan, A.; Reif, A.; Cryan, J.F.; Slattery, D.A. The future of rodent models in depression research. Nat. Rev. Neurosci. 2019, 20, 686–701. [Google Scholar] [CrossRef]
- Amat, J.; Aleksejev, R.M.; Paul, E.; Watkins, L.R.; Maier, S.F. Behavioral control over shock blocks behavioral and neurochemical effects of later social defeat. Neuroscience 2010, 165, 1031–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baratta, M.V.; Lucero, T.R.; Amat, J.; Watkins, L.R.; Maier, S.F. Role of the ventral medial prefrontal cortex in mediating behavioral control-induced reduction of later conditioned fear. Learn. Mem. 2008, 15, 84–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, S.F.; Watkins, L.R. Stressor controllability and learned helplessness: The roles of the dorsal raphe nucleus, serotonin, and corticotropin-releasing factor. Neurosci. Biobehav. Rev. 2005, 29, 829–841. [Google Scholar] [CrossRef] [PubMed]
- Amat, J.; Dolzani, S.D.; Tilden, S.; Christianson, J.P.; Kubala, K.H.; Bartholomay, K.; Sperr, K.; Ciancio, N.; Watkins, L.R.; Maier, S.F. Previous ketamine produces an enduring blockade of neurochemical and behavioral effects of uncontrollable stress. J. Neurosci. 2016, 36, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amat, J.; Paul, E.; Watkins, L.R.; Maier, S.F. Activation of the ventral medial prefrontal cortex during an uncontrollable stressor reproduces both the immediate and long-term protective effects of behavioral control. Neuroscience 2008, 154, 1178–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meine, L.E.; Meier, J.; Schueler, K.; Wessa, M. Why trust is fine, but control is better. Neural correlates of stressor controllability in humans. In Proceedings of the 5th International Symposium on Resilience Research, Mainz, Germany, 25 September 2019. [Google Scholar]
- Dickerson, S.S.; Kemeny, M.E. Acute stressors and cortisol responses: A theoretical integration and synthesis of laboratory research. Psychol. Bull. 2004, 130, 355–391. [Google Scholar] [CrossRef] [Green Version]
- Lucas, M.; Ilin, Y.; Anunu, R.; Kehat, O.; Xu, L.; Desmedt, A.; Richter-Levin, G. Long-term effects of controllability or the lack of it on coping abilities and stress resilience in the rat. Stress 2014, 17, 423–430. [Google Scholar] [CrossRef]
- Dulawa, S.C.; Hen, R. Recent advances in animal models of chronic antidepressant effects: The novelty-induced hypophagia test. Neurosci. Biobehav.Rev. 2005, 29, 771–783. [Google Scholar] [CrossRef]
- Yin, X.; Guven, N.; Dietis, N. Stress-based animal models of depression: Do we actually know what we are doing? Brain Res. 2016, 1652, 30–42. [Google Scholar] [CrossRef]
- Lupien, S.J.; Juster, R.P.; Raymond, C.; Marin, M.F. The effects of chronic stress on the human brain: From neurotoxicity, to vulnerability, to opportunity. Front. Neuroendocrin. 2018, 49, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Gururajan, A.; Kos, A.; Lopez, J.P. Preclinical stress research: Where are we headed? An early career investigator’s perspective. Stress 2018, 21, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Leiner, D. SoSci Survey (Version 3.1.06), 2019. Available online: http://www.soscisurvey.com (accessed on 18 August 2020).
- Petermann, F. WAIS-IV. Wechsler Adult Intelligence Scale–Fourth Edition. Deutschsprachige Adaptation der WAIS-IV von D. Wechsler; Pearson Assessment: Frankfurt/Main, Germany, 2012; Available online: https://www.pearsonclinical.de/pub/media/wysiwyg/Pearson_DE/WAIS_IV_Manual_1.pdf (accessed on 18 August 2020).
- Zimmermann, P.; Fimm, B. Testbatterie zur Aufmerksamkeitsprüfung (TAP) Version 1.7; PSYTEST Verlag: Herzogenrath, Germany, 2002. [Google Scholar]
- Van Rossum, G.; Drake, F.L. The Python Library Reference, Release 2.7; Python Software Foundation: Wolfeboro Falls, NH, USA, 2010; Available online: www.python.org (accessed on 20 August 2020).
- Peirce, J.; Gray, J.R.; Simpson, S.; MacAskill, M.; Höchenberger, R.; Sogo, H.; Kastman, E.; Lindeløv, J.K. PsychoPy2: Experiments in behavior made easy. Behav. Res. Methods 2019, 51, 195–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laux, L.; Hock, M.; Bergner-Köther, R.; Hodapp, V.; Renner, K.H. Stadi: State-Trait-Angst-Depressions-Inventar; Hogrefe: Göttingen, Germany, 2013. [Google Scholar]
- Geue, K.; Strauß, B.; Brähler, E. Diagnostische Verfahren in der Psychotherapie; Hogrefe: Göttingen, Germany, 2016; p. 482. [Google Scholar]
- Krohne, H.W.; Egloff, B.; Kohlmann, C.-W.; Tausch, A. Positive and negative affect schedule—german version. Diagnostica 1996. [Google Scholar] [CrossRef]
- Seligman, M.E.P. Learned helplessness. Annu. Rev. Med. 1972, 23, 407–412. [Google Scholar] [CrossRef]
- von der Malsburg, T. Py-Span.-Task–A Software for Testing Working Memory Span; CERN: Meyrin, Switzerland, 2015. [Google Scholar] [CrossRef]
- Conway, A.R.A.; Kane, M.J.; Bunting, M.F.; Hambrick, D.Z.; Wilhelm, O.; Engle, R.W. Working memory span tasks: A methodological review and user’s guide. Psychon. Bull. Rev. 2005, 12, 769–786. [Google Scholar] [CrossRef]
- Daneman, M.; Carpenter, P.A. Individual differences in working memory and reading. J. Verbal Learning Verbal Behav. 1980, 19, 450–466. [Google Scholar] [CrossRef]
- Holm, S.A. Simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Cohen, J. Eta-Squared and partial eta-squared in fixed factor anova designs. Educ. and Psychol. Meas. 1973, 33, 107–112. [Google Scholar] [CrossRef]
- Tomczak, M.; Tomczak, E. The need to report effect size estimates revisited. An overview of some recommended measures of effect size. Trends Sport Sci. 2014, 1, 19–25. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.r-project.org (accessed on 18 August 2020).
- Kassambara, A. rstatix: Pipe-Friendly Framework for Basic Statistical Tests; R package version 0.5.0, 2020. Available online: https://CRAN.R-project.org/package=rstatix (accessed on 18 August 2020).
- Singmann, H.; Bolker, B.; Westfall, J.; Aust, F.; Ben-Shachar, M.S. afex: Analysis of Factorial Experiments. R package version 0.26-0, 2020. Available online: https://CRAN.R-project.org/package=afex (accessed on 18 August 2020).
- Lenth, R.V. Least-Squares means: The r package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Greiner, B. Subject pool recruitment procedures: Organizing experiments with ORSEE. J. Econ. Sci. Assoc. 2015, 1, 114–125. [Google Scholar] [CrossRef]
- Hadad-Ophir, O.; Ardi, Z.; Brande-Eilat, N.; Kehat, O.; Anunu, R.; Richter-Levin, G. Exposure to prolonged controllable or uncontrollable stress affects GABAergic function in sub-regions of the hippocampus and the amygdala. Neurobiol. Learn. Mem. 2017, 138, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Mair, P.; Wilcox, R.R. Robust statistical methods in R using the WRS2 package. Behav. Res. Methods 2020, 52, 464–488. [Google Scholar] [CrossRef]
- Algina, J.; Keselman, H.J.; Penfield, R.D. An alternative to Cohen’s standardized mean difference effect size: A robust parameter and confidence interval in the two independent groups case. Psychol. Methods 2005, 10, 317–328. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; L. Erlbaum Associates: Hillsdale, NJ, USA, 1988. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Group | Statistical Test | p | Effect Size | ||
|---|---|---|---|---|---|---|
| EC | YC | CC | ||||
| n1 | 27 | 26 | 27 | |||
| Sex (% Female) | 56 | 58 | 56 | Chi-Squared | 0.984 | X2 = 0.03 |
| Mean Age (SD) | 23.48 (3.74) | 25 (4.26) | 25.85 (4.3) | Kruskal–Wallis | <0.05 | η2H = 0.06 |
| Mean Working Memory IQ (SD) | 100.89 (9.55) | 104.58 (11.53) | 104.3 (13.99) | One-Way ANOVA | 0.45 | Partial η2 = 0.02 |
| Mean Alertness (SD) | 45.3 (6.31) | 43.96 (6.99) | 42.11 (6.31) | One-Way ANOVA | 0.24 | Partial η2 = 0.04 |
| Variable | Group | Statistical Test | p | Effect Size | |
|---|---|---|---|---|---|
| EC | YC | ||||
| n1 | 62 | 38 | |||
| Sex (% Female) | 53 | 58 | Chi-Squared | 0.80 | X2 = 0.06 |
| Mean Age (SD) | 22.77 (3.99) | 24.26 (4.2) | t-test | 0.08 | d = −0.36 |
| Mean Level of Education (SD) | 7.84 (1.43) | 8.03 (1.55) | Wilcoxon Rank Sum | 0.51 | r = 0.07 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meine, L.E.; Schüler, K.; Richter-Levin, G.; Scholz, V.; Wessa, M. A Translational Paradigm to Study the Effects of Uncontrollable Stress in Humans. Int. J. Mol. Sci. 2020, 21, 6010. https://doi.org/10.3390/ijms21176010
Meine LE, Schüler K, Richter-Levin G, Scholz V, Wessa M. A Translational Paradigm to Study the Effects of Uncontrollable Stress in Humans. International Journal of Molecular Sciences. 2020; 21(17):6010. https://doi.org/10.3390/ijms21176010
Chicago/Turabian StyleMeine, Laura E., Katja Schüler, Gal Richter-Levin, Vanessa Scholz, and Michele Wessa. 2020. "A Translational Paradigm to Study the Effects of Uncontrollable Stress in Humans" International Journal of Molecular Sciences 21, no. 17: 6010. https://doi.org/10.3390/ijms21176010
APA StyleMeine, L. E., Schüler, K., Richter-Levin, G., Scholz, V., & Wessa, M. (2020). A Translational Paradigm to Study the Effects of Uncontrollable Stress in Humans. International Journal of Molecular Sciences, 21(17), 6010. https://doi.org/10.3390/ijms21176010