Molecular Investigation on a Triple Negative Breast Cancer Xenograft Model Exposed to Proton Beams

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  , , ,
, , ,  , ,
, , 
 and
and 
Abstract
:
1. Introduction
2. Results
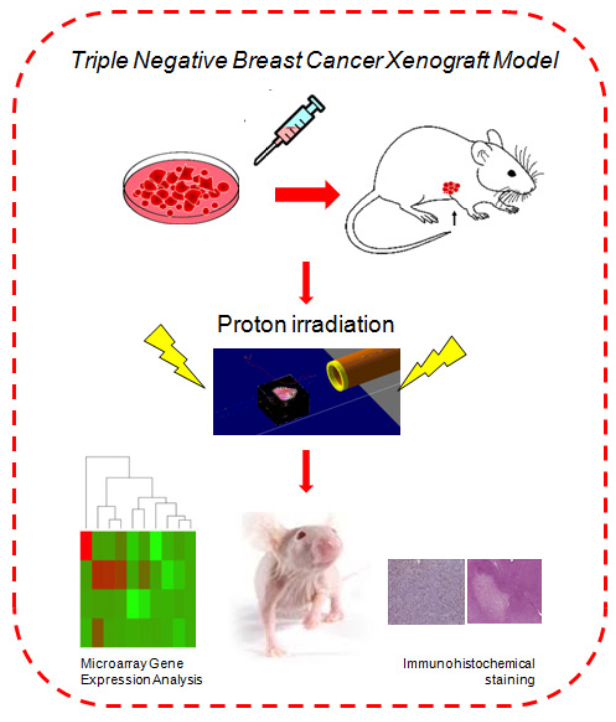
2.1. Monte Carlo Simulation Depth Dose Profile and LET (Linear Energy Transfer) Assessment
2.2. Immunohistochemical Evaluation of Key Markers
High Power Fields (HPF)
2.3. Overview of cDNA Microarray Gene Expression Analyses
- (i)
- MDA-MB-231 xenograft mice_2Gy_PT
- (ii)
- MDA-MB-231 xenograft mice_6Gy_PT
- (iii)
- MDA-MB-231 xenograft mice_9Gy_PT
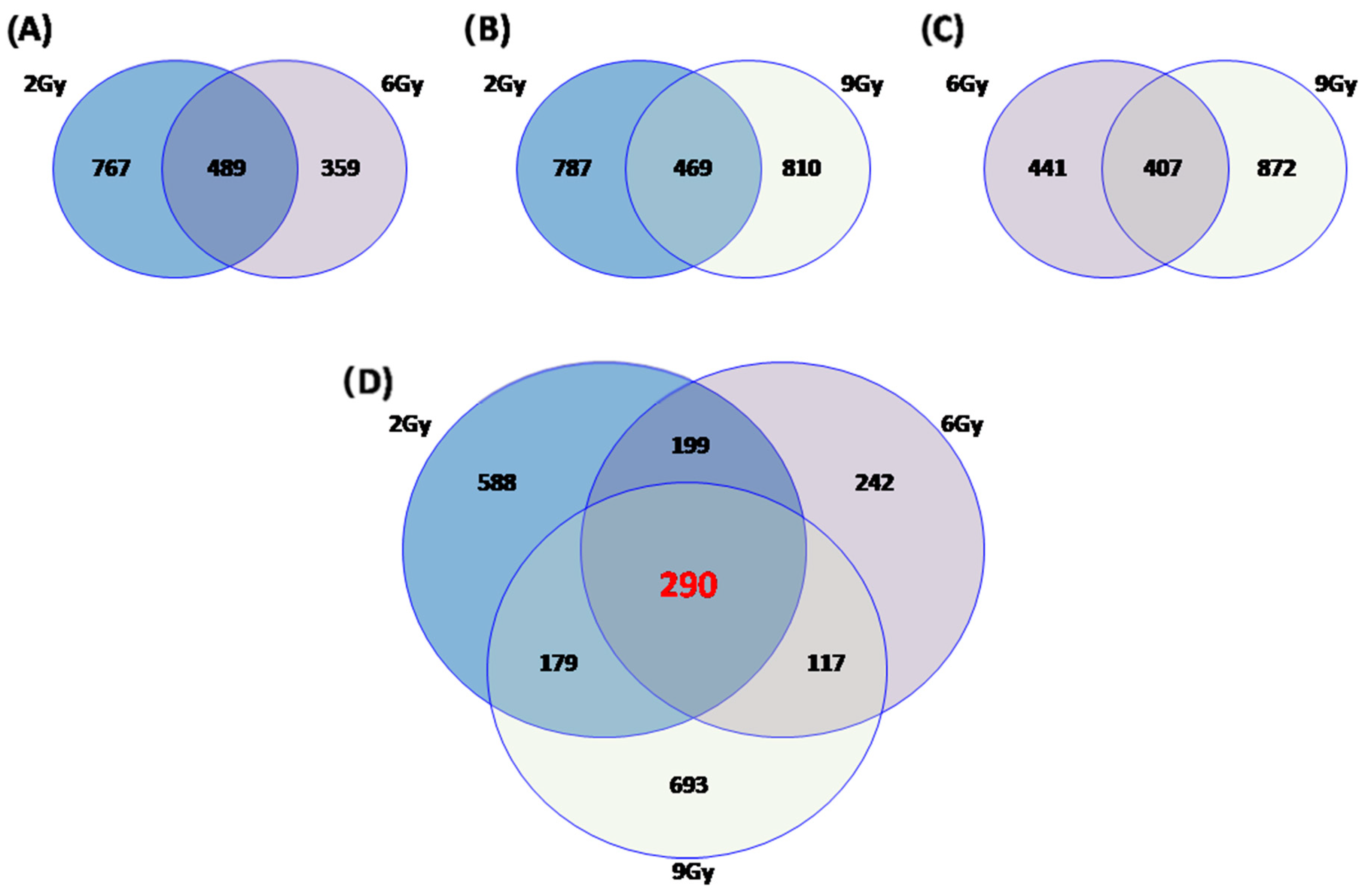
2.4. Commonly Deregulated Genes and Pathways among the PT Schedules
3. Discussion
4. Material and Methods
4.1. Dose Evaluation and Distribution by Monte Carlo GEANT4 Toolkit
4.2. Animal Model
Ethics Statement and Animal Model
4.3. Immunohistochemistry (IHC)
4.4. Apoptotic Count
4.5. Whole Genome cDNA Microarray Expression Analysis
4.6. qRT-PCR Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bravatà, V.; Cava, C.; Minafra, L.; Cammarata, F.P.; Russo, G.; Gilardi, M.C.; Castiglioni, I.; Forte, G.I. Radiation-Induced Gene Expression Changes in High and Low Grade Breast Cancer Cell Types. Int. J. Mol. Sci. 2018, 19, 1084. [Google Scholar] [CrossRef] [Green Version]
- Nicolini, A.; Ferrari, P.; Duffy, M.J. Prognostic and predictive biomarkers in breast cancer: Past, present and future. Semin. Cancer Biol. 2018, 52, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.; Park, S.; Cho, W.K.; Choi, D.H. Cyclin D1 is Associated with Radiosensitivity of Triple-Negative Breast Cancer Cells to Proton Beam Irradiation. Int. J. Mol. Sci. 2019, 20, 4943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Park, H.J.; Shin, K.H.; Kim, J.H.; Choi, O.H.; Park, W.; Ahn, S.D.; Kim, S.S.; Kim, D.Y.; Kim, T.H.; et al. Breast Conservation Therapy Versus Mastectomy in Patients with T1-2N1 Triple-Negative Breast Cancer: Pooled Analysis of KROG 14-18 and 14-23. Cancer Res. Treat. 2018, 50, 1316–1323. [Google Scholar] [CrossRef]
- Wang, J.; Xie, X.; Wang, X.; Tang, J.; Pan, Q.; Zhang, Y.; Di, M. Locoregional and distant recurrences after breast conserving therapy in patients with triple-negative breast cancer: A meta-analysis. Surg. Oncol. 2013, 22, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Abdulkarim, B.; Cuartero, J.; Hanson, J.; Deschênes, J.; Lesniak, D.; Sabri, S. Increased Risk of Locoregional Recurrence for Women with T1-2N0 Triple-Negative Breast Cancer Treated With Modified Radical Mastectomy Without Adjuvant Radiation Therapy Compared With Breast-Conserving Therapy. J. Clin. Oncol. 2011, 29, 2852–2858. [Google Scholar] [CrossRef]
- Bravatà, V.; Cammarata, F.P.; Minafra, L.; Pisciotta, P.; Scazzone, C.; Manti, L.; Savoca, G.; Petringa, G.; Cirrone, G.A.P.; Cuttone, G.; et al. Proton-irradiated breast cells: Molecular points of view. J. Radiat. Res. 2019, 60, 451–465. [Google Scholar] [CrossRef]
- Bravatà, V.; Minafra, L.; Cammarata, F.P.; Pisciotta, P.; Lamia, D.; Marchese, V.; Petringa, G.; Manti, L.; Ap Cirrone, G.; Gilardi, M.C.; et al. Gene expression profiling of breast cancer cell lines treated with proton and electron radiations. Br. J. Radiol. 2018, 91, 1089. [Google Scholar] [CrossRef]
- Tommasino, F.; Durante, M.; D’Avino, V.; Liuzzi, R.; Conson, M.; Farace, P.; Palma, G.; Schwarz, M.; Cella, L.; Pacelli, R. Model-based approach for quantitative estimates of skin, heart, and lung toxicity risk for left-side photon and proton irradiation after breast-conserving surgery. Acta Oncol. 2017, 56, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Spetz, J.; Moslehi, J.; Sarosiek, K. Radiation-Induced Cardiovascular Toxicity: Mechanisms, Prevention, and Treatment. Curr. Treat. Options Cardiovasc. Med. 2018, 20, 31. [Google Scholar] [CrossRef]
- Chowdhary, M.; Lee, A.; Gao, S.; Wang, D.; Barry, P.N.; Diaz, R.; Bagadiya, N.R.; Park, H.S.; Yu, J.B.; Wilson, L.D.; et al. Is Proton Therapy a “Pro” for Breast Cancer? A Comparison of Proton vs. Non-proton Radiotherapy Using the National Cancer Database. Front. Oncol. 2019, 8, 678. [Google Scholar] [CrossRef] [Green Version]
- Cuaron, J.; Macdonald, S.M.; Cahlon, O. Novel applications of proton therapy in breast carcinoma. Chin. Clin. Oncol. 2016, 5, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbin, K.S.; Mutter, R.W. Proton therapy for breast cancer: Progress & pitfalls. Breast Cancer Manag. 2018, 7, BMT06. [Google Scholar] [CrossRef]
- Orecchia, R.; Fossati, P.; Zurrida, S.; Krengli, M. New frontiers in proton therapy. Curr. Opin. Oncol. 2015, 27, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Parshad, R.; Suhani; Kazi, M.; Seenu, V.; Mathur, S.; Dattagupta, S.; Haresh, K.P. Triple-negative breast cancers: Are they always different from nontriple-negative breast cancers? An experience from a tertiary center in India. Indian J. Cancer 2018, 54, 658–663. [Google Scholar] [CrossRef]
- Gong, C.; Ma, G.; Hu, X.; Zhang, Y.; Wang, Z.-H.; Zhang, J.; Zhao, Y.; Li, Y.; Xie, Y.; Yang, Z.; et al. Pretreatment 18 F-FDG Uptake Heterogeneity Predicts Treatment Outcome of First-Line Chemotherapy in Patients with Metastatic Triple-Negative Breast Cancer. Oncologist 2018, 23, 1144–1152. [Google Scholar] [CrossRef] [Green Version]
- Russo, G.; Pisciotta, P.; Cirrone, G.; Romano, F.; Cammarata, F.; Marchese, V.; Forte, G.; Lamia, D.; Minafra, L.; Bravatá, V.; et al. Preliminary study for small animal preclinical hadrontherapy facility. Nucl. Instrum. Methods Phys. Res. Sect. 2017, 846, 126–134. [Google Scholar] [CrossRef]
- Pauwels, A.-M.; Trost, M.; Beyaert, R.; Hoffmann, E. Patterns, Receptors, and Signals: Regulation of Phagosome Maturation. Trends Immunol. 2017, 38, 407–422. [Google Scholar] [CrossRef] [Green Version]
- Iozzo, R.V.; Sanderson, R.D. Proteoglycans in cancer biology, tumour microenvironment and angiogenesis. J. Cell. Mol. Med. 2011, 15, 1013–1031. [Google Scholar] [CrossRef]
- Tommasino, F.; Durante, M. Proton Radiobiology. Cancers 2015, 7, 353–381. [Google Scholar] [CrossRef]
- Pisciotta, P.; Costantino, A.; Cammarata, F.P.; Torrisi, F.; Marchese, V.; Cirrone, G.A.P.; Petringa, G.; Forte, G.I.; Minafra, L.; Bravatà, V.; et al. Evaluation of Proton Beam Radiation-induced Skin injury in a murine model by end-part of a SOBP. PLoS ONE 2020, 22, e0233258. [Google Scholar]
- Workman, P.; An ad hoc committee of the National Cancer Research Institute; Aboagye, E.O.; Balkwill, F.; Balmain, A.; Bruder, G.; Chaplin, D.J.; Double, J.A.; Everitt, J.; Farningham, D.A.H.; et al. Guidelines for the welfare and use of animals in cancer research. Br. J. Cancer 2010, 102, 1555–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Zhang, Z.; Zhou, L.; Hu, L.; Yin, C.; Qing, D.; Huang, S.; Cai, X.; Chen, Y.; Qin, J. Cancer associated fibroblasts-derived exosomes contribute to radioresistance through promoting colorectal cancer stem cells phenotype. Exp. Cell Res. 2020, 391, 111956. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Shankar, B.S. Radio resistance in breast cancer cells is mediated through TGF-β signalling, hybrid epithelial-mesenchymal phenotype and cancer stem cells. Biomed. Pharmacother. 2019, 111, 119–130. [Google Scholar] [CrossRef]
- Langlands, F.E.; Horgan, K.; Dodwell, D.D.; Smith, L. Breast cancer subtypes: Response to radiotherapy and potential radiosensitisation. Br. J. Radiol. 2013, 86, 20120601. [Google Scholar] [CrossRef]
- Willers, H.; Azzoli, C.G.; Santivasi, W.L.; Xia, F. Basic Mechanisms of Therapeutic Resistance to Radiation and Chemotherapy in Lung Cancer. Cancer J. 2013, 19, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Césaire, M.; Ghosh, U.; Austry, J.-B.; Muller, E.; Cammarata, F.P.; Guillamin, M.; Caruso, M.; Castéra, L.; Petringa, G.; Cirrone, G.A.P.; et al. Sensitization of chondrosarcoma cells with PARP inhibitor and high-LET radiation. J. Bone Oncol. 2019, 17, 100246. [Google Scholar] [CrossRef]
- Niazi, M.K.K.; Senaras, C.; Pennell, M.L.; Arole, V.; Tozbikian, G.; Gurcan, M.N. Relationship between the Ki67 index and its area based approximation in breast cancer. BMC Cancer 2018, 18, 867. [Google Scholar] [CrossRef]
- Niazi, M.K.K.; Downs-Kelly, E.; Gurcan, M.N. Hot spot detection for breast cancer in Ki-67 stained slides: Image dependent filtering approach. In Medical Imaging 2014: Digital Pathology; SPIE: Bellingham, WA, USA, 2014. [Google Scholar]
- Di Maggio, F.M.; Minafra, L.; Forte, G.I.; Cammarata, F.P.; Lio, D.; Messa, C.; Gilardi, M.C.; Bravatà, V. Portrait of inflammatory response to ionizing radiation treatment. J. Inflamm. 2015, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Minafra, L.; Bravatà, V. Cell and molecular response to IORT treatment. Transl. Cancer Res. 2014, 3, 32–47. [Google Scholar]
- Aggarwal, B.B.; Sethi, G.; Ahn, K.S.; Sandur, S.K.; Pandey, M.K.; Kunnumakkara, A.B.; Sung, B.; Ichikawa, H. Targeting signal-transducer-and-activator-of-transcription-3 for prevention and therapy of cancer: Modern target but ancient solution. Ann. N. Y. Acad. Sci. 2006, 1091, 151–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimamura, H.; Terada, Y.; Okado, T.; Tanaka, H.; Inoshita, S.; Sasaki, S. The PI3-kinase-Akt pathway promotes mesangial cell survival and inhibits apoptosis in vitro via NF-kappa B and Bad. J. Am. Soc. Nephrol. 2003, 14, 1427–1434. [Google Scholar] [CrossRef] [Green Version]
- Bravatà, V.; Cammarata, F.P.; Minafra, L.; Musso, R.; Pucci, G.; Spada, M.; Fazio, I.; Russo, G.; Forte, G.I.; Cammarata, P.F. Gene Expression Profiles Induced by High-dose Ionizing Radiation in MDA-MB-231 Triple-negative Breast Cancer Cell Line. Cancer Genom. Proteom. 2019, 16, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Martínez, P.; Gómez-López, G.; Pisano, D.G.; Flores, J.M.; Blasco, M.A. A genetic interaction between RAP1 and telomerase reveals an unanticipated role for RAP1 in telomere maintenance. Aging Cell 2016, 15, 1113–1125. [Google Scholar] [CrossRef]
- Lin, H.-J.; Liu, H.-H.; Lin, C.-D.; Kao, M.-C.; Chen, Y.-A.; Chiang-Ni, C.; Jiang, Z.-P.; Huang, M.-Z.; Lin, C.-J.; Lo, U.-G.; et al. Cytolethal Distending Toxin Enhances Radiosensitivity in Prostate Cancer Cells by Regulating Autophagy. Front. Microbiol. 2017, 7, 223. [Google Scholar] [CrossRef] [PubMed]
- Gueulette, J.; Blattmann, H.; Pedroni, E.; Coray, A.; De Coster, B.M.; Mahy, P.; Wambersie, A.; Goitein, G. Relative biologic effectiveness determination in mouse intestine for scanning proton beam at Paul Scherrer Institute, Switzerland. Influence of motion. Int. J. Radiat. Oncol. 2005, 62, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Pisciotta, P.; Russo, G.; Marchese, V.; Cirrone, G.; Romano, F.; Lamia, D.; Cuttone, G.; Gilardi, M. Preliminary dosimetric study for preclinical small animal hadrontherapy. Phys. Medica 2016, 32, 53. [Google Scholar] [CrossRef]
- Broggi, G.; Musumeci, G.; Puzzo, L.; Russo, A.; Reibaldi, M.; Ragusa, M.; Longo, A.; Caltabiano, R. Immunohistochemical Expression of ABCB5 as a Potential Prognostic Factor in Uveal Melanoma. Appl. Sci. 2019, 9, 1316. [Google Scholar] [CrossRef] [Green Version]
- Salgado, R.; Denkert, C.; DeMaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; Eynden, G.V.D.; Baehner, F.L.; Penault-Llorca, F.; et al. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. 2015, 26, 259–271. [Google Scholar] [CrossRef]
- Bravatà, V.; Minafra, L.; Russo, G.; Forte, G.I.; Cammarata, F.P.; Ripamonti, M.; Casarino, C.; Augello, G.; Costantini, F.; Barbieri, G.; et al. High-dose Ionizing Radiation Regulates Gene Expression Changes in the MCF7 Breast Cancer Cell Line. Anticancer. Res. 2015, 35, 2577–2591. [Google Scholar]
- Minafra, L.; Bravatà, V.; Russo, G.; Forte, G.I.; Cammarata, F.P.; Ripamonti, M.; Candiano, G.; Cervello, M.; Giallongo, A.; Perconti, G.; et al. Gene Expression Profiling of MCF10A Breast Epithelial Cells Exposed to IOERT. Anticancer. Res. 2015, 35, 3223–3234. [Google Scholar] [PubMed]
- Minafra, L.; Porcino, N.; Bravatà, V.; Gaglio, D.; Bonanomi, M.; Amore, E.; Cammarata, F.P.; Russo, G.; Militello, C.; Savoca, G.; et al. Radiosensitizing effect of curcumin-loaded lipid nanoparticles in breast cancer cells. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, K.; Hosack, D.A.; Dennis, G.; Lempicki, R.A.; Bright, T.J.; Cheadle, C.; Engel, J. PubMatrix: A tool for multiplex literature mining. BMC Bioinform. 2003, 4, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minafra, L.; Bravatà, V.; Cammarata, F.P.; Russo, G.; Gilardi, M.C.; Forte, G.I. Radiation Gene-expression Signatures in Primary Breast Cancer Cells. Anticancer. Res. 2018, 38, 2707–2715. [Google Scholar] [CrossRef] [PubMed]
- Cirrone, G.; Margarone, D.; Maggiore, M.; Anzalone, A.; Borghesi, M.; Jia, S.B.; Bulanov, S.S.; Bulanov, S.; Carpinelli, M.; Cavallaro, S.; et al. ELIMED: A new hadron therapy concept based on laser driven ion beams. In Laser Acceleration of Electrons, Protons, and Ions II; and Medical Applications of Laser-Generated Beams of Particles II; and Harnessing Relativistic Plasma Waves III; International Society for Optics and Photonics: Bellingham, WA, USA, 2013. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
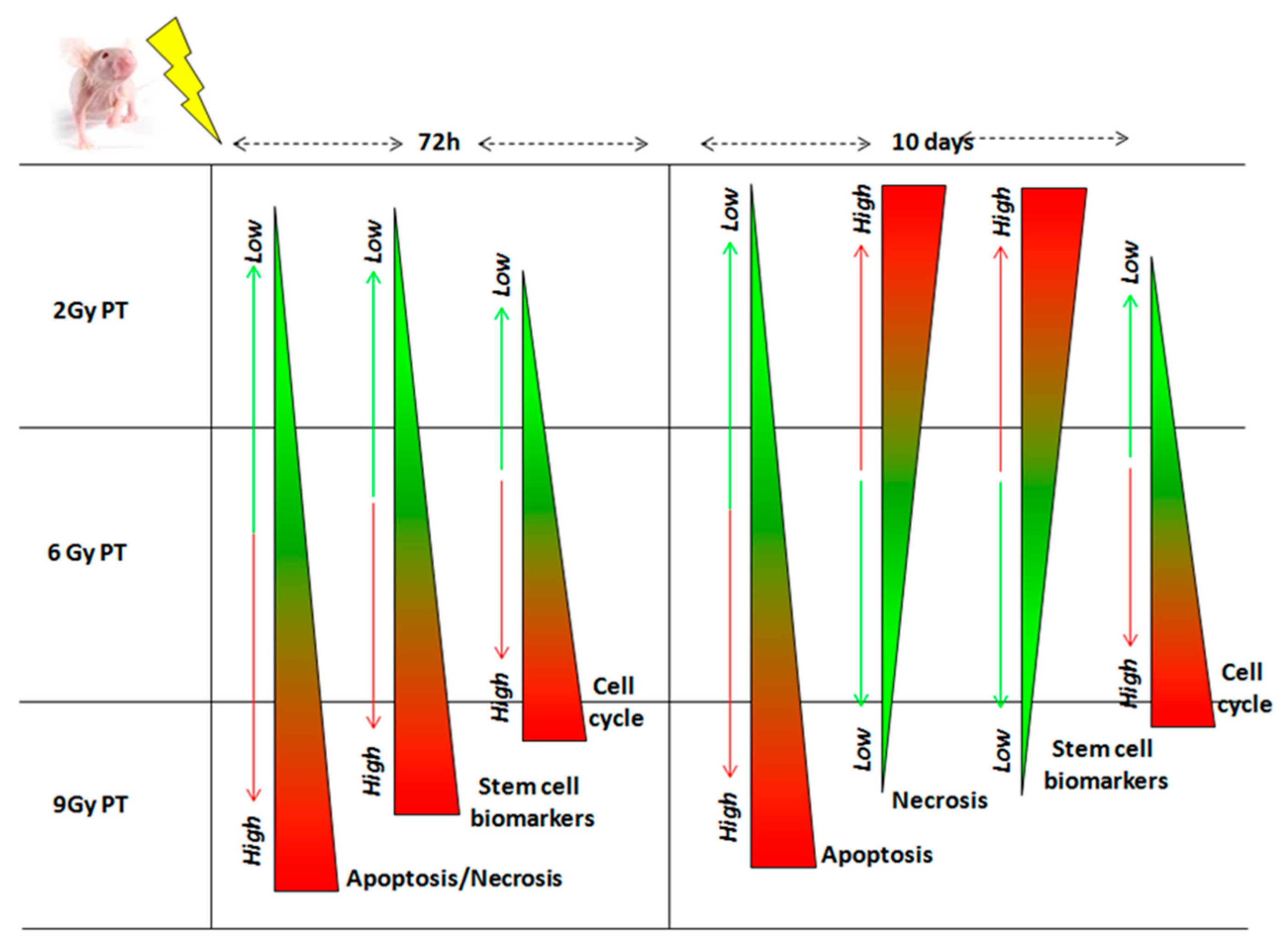{kind=link}
| Samples | Number of Necrotic Foci | Extension of Necrosis (%) | Caspase-3 | CD133 | Ki67 | CD68 | Cyclin D1 | Apoptotic Count (10 High Power Fields) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IS | ES | IRS | IS | ES | IRS | |||||||
| Untreated (72 h) | 0 | 0 | 1 | 1 | 1 | 2 | 1 | 2 | 60% | Negative | Negative | 10 |
| Untreated (10 days) | 4 | 0.45 | 2 | 4 | 8 | 3 | 3 | 9 | 70% | Negative | Negative | 11 |
| MDA-MB-231 Xenograft mice exposed to 2 Gy (72 h) | 1 | 10% | 1 | 2 | 2 | 2 | 2 | 4 | 50% | Negative | Positive (+IS 2; ES 2; IRS 4) | 12 |
| MDA-MB-231 Xenograft mice exposed to 2 Gy (10 days) | 3 | 20% | 2 | 3 | 6 | 3 | 3 | 9 | 65% | Positive (+5–20%) | Positive (+IS 1; ES 2; IRS 2) | 16 |
| MDA-MB-231 Xenograft mice exposed to 6 Gy (72 h) | 3 | 25% | 2 | 4 | 8 | 3 | 3 | 9 | 50% | Negative | Positive (+IS 2; ES 2; IRS 4) | 14 |
| MDA-MB-231 Xenograft mice exposed to 6 Gy (10 days) | 2 | 15% | 2 | 2 | 4 | 2 | 3 | 6 | 70% | Positive (+5–20%) | Positive (+IS 2; ES 2; IRS 4) | 14 |
| MDA-MB-231 Xenograft mice exposed to 9 Gy (72 h) | 2 | 50% | 2 | 4 | 8 | 3 | 3 | 9 | 60% | Negative | Positive (+IS 2; ES 3; IRS 6) | 15 |
| MDA-MB-231 Xenograft mice exposed to 9 Gy (10 days) | 1 | 5% | 1 | 2 | 2 | 2 | 2 | 4 | 60% | Positive (++20–50%) | Positive (+IS 1; ES 3; IRS 3) | 14 |
| Top 5 Molecular Pathways of Differentially Expressed Genes of MDA-MB-231 Xenograft Mice Exposed to 2 Gy of Proton Irradiations | ||||
|---|---|---|---|---|
| Term | Count | p Value | Genes | |
| 1 | Graft-versus-host disease | 12 | 8.1 × 10−7 | HLA-DQB1, HLA-DRB1, HLA-A, IL1B, FASLG, HLA-C, HLA-DPA1, HLA-B, HLA-DPB1, HLA-DOA, HLA-E, HLA-DRA |
| 2 | Allograft rejection | 11 | 2.1 × 10−5 | HLA-DQB1, HLA-DRB1, HLA-A, FASLG, HLA-C, HLA-DPA1, HLA-B, HLA-DPB1, HLA-DOA, HLA-E, HLA-DRA |
| 3 | Antigen processing and presentation | 15 | 5.8 × 10−5 | CIITA, HLA-DQB1, HLA-DRB1, HLA-A, HSPA1A, HLA-C, HLA-B, HLA-E, CD74, HSPA6, KIR3DL3, HLA-DPA1, HLA-DPB1, HLA-DOA, HLA-DRA |
| 4 | Phagosome | 21 | 2.0 × 10−4 | HLA-DQB1, NOS1, HLA-DRB1, HLA-A, HLA-C, HLA-B, SFTPA1, ITGB3, HLA-E, CLEC4M, FCAR, CD209, COMP, TUBAL3, HLA-DPA1, SCARB1, HLA-DPB1, HLA-DOA, ATP6V0D2, TUBB4A, HLA-DRA |
| 5 | Cell adhesion molecules (CAMs) | 19 | 7.7 × 10−4 | PVR, HLA-DQB1, HLA-DRB1, CLDN5, HLA-A, NLGN1, HLA-C, HLA-B, HLA-E, CLDN15, ALCAM, NCAM2, SDC1, CD2, MADCAM1, HLA-DPA1, HLA-DPB1, HLA-DOA, HLA-DRA |
| Top 5 Molecular Pathways of Differentially Expressed Genes of MDA-MB-231 Xenograft Mice Exposed to 6 Gy of Proton Irradiations | ||||
|---|---|---|---|---|
| Term | Count | p Value | Genes | |
| 1 | Proteoglycans in cancer | 22 | 1.2 × 10−5 | NANOG, ERBB4, ROCK2, HCLS1, ERBB2, FASLG, IGF2, FZD3, HGF, DCN, ITGB3, MMP2, PXN, KDR, CTNNB1, SMO, MAPK13, HPSE, PLCG2, HSPB2, PRKACA, TWIST1 |
| 2 | Leukocyte transendothelial migration | 11 | 9.5 × 10−3 | ITGAL, ROCK2, MAPK13, PLCG2, CLDN5, CTNND1, MYLPF, JAM2, MMP2, PXN, CTNNB1 |
| 3 | Graft-versus-host disease | 5 | 3.2 × 10−2 | HLA-DQB1, HLA-DRB1, FASLG, HLA-DPA1, HLA-DPB1 |
| 4 | Allograft rejection | 5 | 4.6 × 10−2 | HLA-DQB1, HLA-DRB1, FASLG, HLA-DPA1, HLA-DPB1 |
| 5 | Phagosome | 11 | 4.9 × 10−2 | HLA-DQB1, TUBA8, CD36, HLA-DRB1, TUBAL3, HLA-DPA1, SFTPA1, COLEC11, HLA-DPB1, ITGB3, TUBB4A |
| Top 15 Molecular Pathways of Differentially Expressed Genes of MDA-MB-231 Xenograft Mice Exposed to 9 Gy of Proton Irradiations | ||||
|---|---|---|---|---|
| Term | Count | p Value | Genes | |
| 1 | Proteoglycans in cancer | 26 | 8.4 × 10−5 | ERBB2, ITGB3, DCN, MMP2, GPC3, ANK2, HPSE, PPP1R12A, PIK3R5, PIK3R3, PIK3CG, NANOG, WNT10B, ROCK2, ITGA2, IGF2, FZD3, PRKCG, FZD5, FLNB, KDR, WNT2B, EIF4B, MAPK13, VEGFA, WNT11 |
| 2 | Rap1 signaling pathway | 26 | 0.0002 | PRKCZ, FGFR3, RAP1GAP, TLN2, CTNND1, LPAR3, FGF13, ITGB3, ITGAM, RAC3, RASGRP2, RAPGEF4, PIK3R5, PIK3R3, ANGPT2, PLCB2, PIK3CG, FYB, GNAO1, GRIN1, PRKCG, KDR, DOCK4, MAPK13, VEGFA, PDGFRA |
| 3 | Pathways in cancer | 40 | 0.0002 | FGFR3, APC2, PTGS2, ERBB2, GNG13, CXCL8, LPAR3, FGF13, MMP2, SUFU, AGTR1, CDKN2A, RAC3, CASP8, RASGRP2, PIK3R5, HHIP, PIK3R3, PLCB2, PIK3CG, CEBPA, PTGER1, COL4A3, WNT10B, HSP90AA1, BCR, VHL, RALBP1, ROCK2, TGFBR2, BRCA2, ITGA2, PRKCG, FZD3, FZD5, STAT1, WNT2B, VEGFA, PDGFRA, WNT11 |
| 4 | Signaling pathways regulating pluripotency of stem cells | 19 | 0.0006 | PIK3CG, NANOG, WNT10B, FGFR3, ONECUT1, APC2, PAX6, FZD3, FZD5, WNT2B, RIF1, HAND1, MAPK13, PIK3R5, WNT11, JAK3, SKIL, BMPR1B, PIK3R3 |
| 5 | VEGF signaling pathway | 11 | 0.002 | PIK3CG, PTGS2, RAC3, MAPK13, VEGFA, PPP3R2, PRKCG, NOS3, PIK3R5, PIK3R3, KDR |
| 6 | Platelet activation | 16 | 0.005 | PIK3CG, PRKCZ, ROCK2, TLN2, COL3A1, ITGA2, ITGB3, PRKG1, MAPK13, RASGRP2, PPP1R12A, PIK3R5, NOS3, PIK3R3, PLCB2, COL11A1 |
| 7 | Phagosome | 17 | 0.008 | HLA-DQB1, NOS1, HLA-DRB1, ITGA2, C1R, SFTPA1, ITGB3, HLA-DQA2, ITGAM, TUBA8, ATP6V0E2, CD209, COMP, TUBAL3, HLA-DPA1, HLA-DPB1, HLA-DRA |
| 8 | Antigen processing and presentation | 11 | 0.008 | HLA-DQB1, HSP90AA1, HSPA2, HLA-DRB1, CD8A, KIR3DL3, HLA-DPA1, HLA-DPB1, HLA-DQA2, CD74, HLA-DRA |
| 9 | Graft-versus-host disease | 7 | 0.008 | HLA-DQB1, CD86, HLA-DRB1, HLA-DPA1, HLA-DPB1, HLA-DQA2, HLA-DRA |
| 10 | Focal adhesion | 21 | 0.009 | PIK3CG, COL4A3, ROCK2, TLN2, ERBB2, COL3A1, ITGA10, ITGA2, PRKCG, ITGB3, FLNB, KDR, RAC3, COMP, VEGFA, PPP1R12A, PDGFRA, PIK3R5, PIK3R3, COL11A1, SHC4 |
| 11 | Ras signaling pathway | 22 | 0.01 | PIK3CG, PLD1, PLA2G16, FGFR3, RALBP1, GRIN1, GNG13, PRKCG, FGF13, RGL1, KDR, RAC3, RASGRP2, VEGFA, PDGFRA, ZAP70, PIK3R5, SYNGAP1, PIK3R3, PLA2G2D, ANGPT2, SHC4 |
| 12 | Allograft rejection | 7 | 0.01 | HLA-DQB1, CD86, HLA-DRB1, HLA-DPA1, HLA-DPB1, HLA-DQA2, HLA-DRA |
| 13 | Chemokine signaling pathway | 18 | 0.03 | CXCL1, PIK3CG, PRKCZ, FGR, ROCK2, GNG13, CXCL8, CCL4L2, STAT1, CCL15, CCL26, CCR3, RASGRP2, PIK3R5, JAK3, PIK3R3, PLCB2, SHC4 |
| 14 | HIF-1 signaling pathway | 11 | 0.04 | PIK3CG, VHL, PFKFB3, HMOX1, ERBB2, VEGFA, PRKCG, NOS3, PIK3R5, PIK3R3, ANGPT2 |
| 15 | Wnt signaling pathway | 14 | 0.04 | WNT10B, APC2, ROCK2, PPP3R2, FZD3, PRKCG, FZD5, WNT2B, CSNK2A1, SOST, SFRP1, RAC3, WNT11, PLCB2 |
| Gene Expression Analyses of Key Genes of TNBC Xenograft Mice Exposed to PT | |||||||
|---|---|---|---|---|---|---|---|
| Gene Symbol | Description | 2 Gy PRT | 6 Gy PRT | 9 Gy PRT | |||
| Array | qRT-PCR | Array | qRT-PCR | Array | qRT-PCR | ||
| BAX | BCL2 associated X, apoptosis regulator | −1.25 | 0.51 | −1.12 | 0.32 | 1.14 | 2.26 |
| BCL2 | BCL2 apoptosis regulator | −1.20 | 0.56 | −1.20 | 0.28 | 1.18 | 1.87 |
| CCNA2 | Cyclin A2 | −1.24 | 0.62 | −2.73 | 0.30 | Und. | 2.24 |
| CD24 | CD24 molecule | 1.43 | 1.24 | 1.23 | Und. | Und. | 2.92 |
| CD44 | CD44 molecule | 1.37 | 0.97 | 1.07 | Und. | 1.24 | 3.16 |
| CDC20 | Cell division cycle 20 | −1.11 | 0.48 | −1.11 | 0.83 | 1.04 | 3.83 |
| CDC25 | Cell division cycle 25 | −1.19 | 0.38 | Und | 0.51 | 1.11 | 1.51 |
| FOS | Fos proto-oncogene, AP-1 transcription factor subunit | 4.95 | 3.35 | 1.12 | Und. | 1.91 | 30.56 |
| FOSB | FosB proto-oncogene, AP-1 transcription factor subunit | 2.51 | 1.07 | 1.34 | 0.57 | 1.30 | 6.35 |
| GDF15 | Growth differentiation factor 15 | 1.86 | 1.64 | 1.72 | 1.23 | 2.50 | 10.27 |
| MMP9 | Matrix metallopeptidase 9 | −1.16 | 0.21 | Und | 0.38 | 1.33 | 2.31 |
| RRAD | Ras related glycolysis inhibitor and calcium channel regulator | 1.50 | 3.78 | 1.54 | 1.95 | 1.71 | 6.51 |
| TAF7L | TATA-box binding protein associated factor 7 like | 1.14 | 1.27 | Und | Und. | 1.18 | 1.80 |
| TNF | Tumor necrosis factor | 1.34 | 4.14 | Und | 1.31 | 1.17 | 5.02 |
| TP53 | Tumor protein p53 | n.a. | 3.11 | 1.33 | 1.09 | 1.25 | 3.28 |
| TP53INP1 | Tumor protein p53 inducible nuclear protein 1 | 1.04 | 1.40 | 1.23 | 1.76 | Und. | 4.53 |
| WNT5A | Wnt family member 5A | 1.17 | 2.12 | 1.26 | 2.64 | 1.14 | 9.28 |
| Top 5 Molecular Pathways of Differentially Expressed Genes Shared Between MDA-MB-231 Xenograft Mice Exposed to 2, 6 and 9 Gy of Proton Irradiations | ||||
|---|---|---|---|---|
| Term | Count | p Value | Genes | |
| 1 | Antigen processing and presentation | 6 | 0.003 | HLA-DQB1, HLA-DRB1, KIR3DL3, HLA-DPA1, HLA-DPB1, CD74 |
| 2 | Graft-versus-host disease | 4 | 0.009 | HLA-DQB1, HLA-DRB1, HLA-DPA1, HLA-DPB1 |
| 3 | Allograft rejection | 4 | 0.01 | HLA-DQB1, HLA-DRB1, HLA-DPA1, HLA-DPB1 |
| 4 | Phagosome | 6 | 0.04 | HLA-DQB1, HLA-DRB1, TUBAL3, HLA-DPA1, HLA-DPB1, ITGB3 |
| 5 | Complement and coagulation cascades | 4 | 0.05 | F5, CD46, F13A1, C1S |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cammarata, F.P.; Forte, G.I.; Broggi, G.; Bravatà, V.; Minafra, L.; Pisciotta, P.; Calvaruso, M.; Tringali, R.; Tomasello, B.; Torrisi, F.; et al. Molecular Investigation on a Triple Negative Breast Cancer Xenograft Model Exposed to Proton Beams. Int. J. Mol. Sci. 2020, 21, 6337. https://doi.org/10.3390/ijms21176337
Cammarata FP, Forte GI, Broggi G, Bravatà V, Minafra L, Pisciotta P, Calvaruso M, Tringali R, Tomasello B, Torrisi F, et al. Molecular Investigation on a Triple Negative Breast Cancer Xenograft Model Exposed to Proton Beams. International Journal of Molecular Sciences. 2020; 21(17):6337. https://doi.org/10.3390/ijms21176337
Chicago/Turabian StyleCammarata, Francesco P., Giusi I. Forte, Giuseppe Broggi, Valentina Bravatà, Luigi Minafra, Pietro Pisciotta, Marco Calvaruso, Roberta Tringali, Barbara Tomasello, Filippo Torrisi, and et al. 2020. "Molecular Investigation on a Triple Negative Breast Cancer Xenograft Model Exposed to Proton Beams" International Journal of Molecular Sciences 21, no. 17: 6337. https://doi.org/10.3390/ijms21176337
APA StyleCammarata, F. P., Forte, G. I., Broggi, G., Bravatà, V., Minafra, L., Pisciotta, P., Calvaruso, M., Tringali, R., Tomasello, B., Torrisi, F., Petringa, G., Cirrone, G. A. P., Cuttone, G., Acquaviva, R., Caltabiano, R., & Russo, G. (2020). Molecular Investigation on a Triple Negative Breast Cancer Xenograft Model Exposed to Proton Beams. International Journal of Molecular Sciences, 21(17), 6337. https://doi.org/10.3390/ijms21176337