The Influence of Radiation on Bone and Bone Cells—Differential Effects on Osteoclasts and Osteoblasts


 ,
,  and
and 
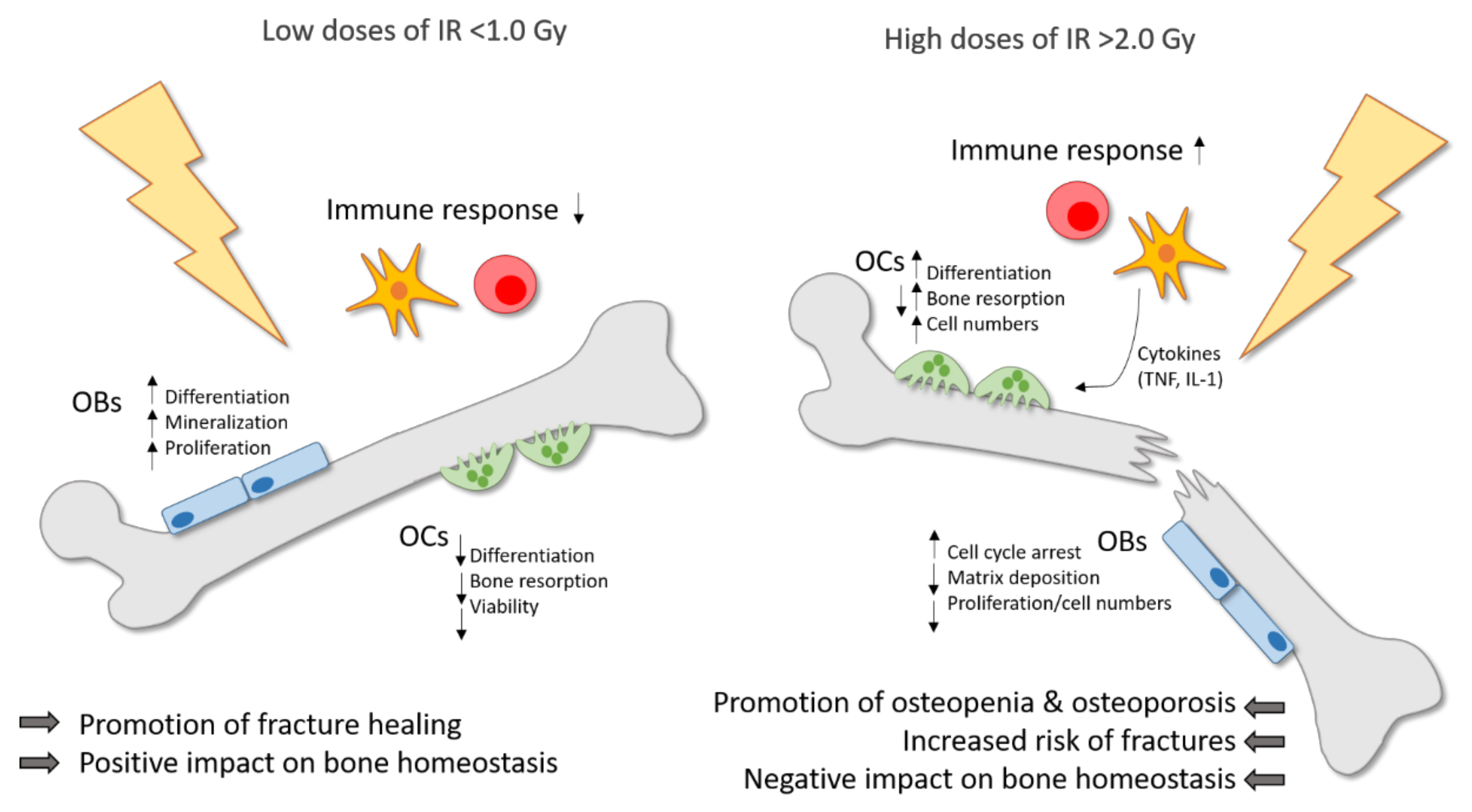{kind=link}
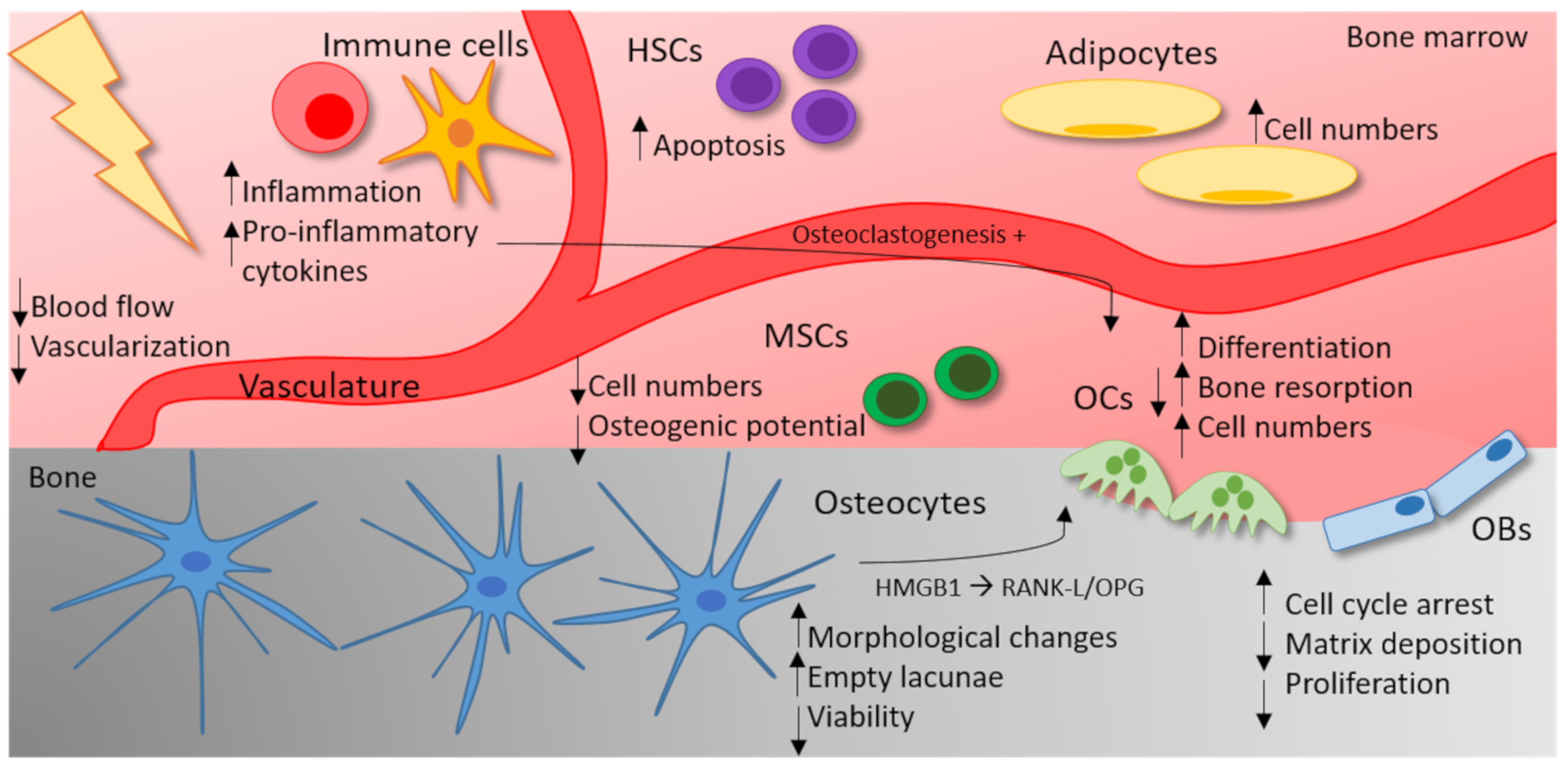{kind=link}
Abstract
:1. Introduction
1.1. Osteoclast Differentiation and Function
1.2. Effects of Ionizing Radiation on Cells
2. High Dose Radiation—IR in Cancer Therapy and the Influence on Bone Cells
3. Low-Dose Radiation—IR for Degenerative and Inflammatory Diseases and the Influence on Bone Cells
4. IR in Space Travel and the Influence on Bone Cells
5. The Influence of IR on the Bone Microenvironment
6. Conclusion and Outlook
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| OC | Osteoclast |
| OB | Osteoblast |
| Cts | Cathepsin |
| MMP | Matrixmetalloproteinase |
| HSC | Hematopoietic stem cell |
| CMP | Common myeloid progenitor |
| RANK-L | Receptor activator of nuclear factor kappa B ligand |
| NF-ƙB | Nuclear factor kappa-light-chain-enhancer of activated B-cells |
| NFATc1 | Nuclear factor of activated T cells 1 |
| M-CSF | Macrophage colony-stimulating factor |
| DC-STAMP | Dendritic cell-specific transmembrane protein |
| TRAP | Tartrate-resistant acid phosphatase |
| CtsK | Cathepsin K |
| OC-STAMP | Osteoclast stimulatory transmembrane protein |
| CCL2 | C-C motif chemokine ligand 2 |
| IR | Ionizing radiation |
| ROS | Reactive oxygen species |
| DAMPs | Damage-associated molecular patterns |
| RT | Radiotherapy |
| Gy | Gray |
| TGF-β | Transforming growth factor β |
| TNF-α | Tumor necrosis factor α |
| IL | Interleukin |
| LD-RT | Low-dose radiation therapy |
| NO | Nitric oxide |
| LET | Linear energy transfer |
| OPG | Osteoprotegerin |
| HMGB1 | High mobility group box 1 |
| VEGF | Vascular endothelial growth factor |
References
- Buckwalter, J.A.; Glimcher, M.J.; Cooper, R.R.; Recker, R. Bone biology. I: Structure, blood supply, cells, matrix, and mineralization. Instr. Course Lect. 1996, 45, 371–386. [Google Scholar] [PubMed]
- Okamoto, K.; Nakashima, T.; Shinohara, M.; Negishi-Koga, T.; Komatsu, N.; Terashima, A.; Sawa, S.; Nitta, T.; Takayanagi, H. Osteoimmunology: The Conceptual Framework Unifying the Immune and Skeletal Systems. Physiol. Rev. 2017, 97, 1295–1349. [Google Scholar] [CrossRef]
- Parfitt, A.M. High bone turnover is intrinsically harmful: Two paths to a similar conclusion. The Parfitt view. J. Bone Miner. Res. 2002, 17, 1558–1559. [Google Scholar] [CrossRef] [PubMed]
- Lees, R.L.; Sabharwal, V.K.; Heersche, J.N. Resorptive state and cell size influence intracellular pH regulation in rabbit osteoclasts cultured on collagen-hydroxyapatite films. Bone 2001, 28, 187–194. [Google Scholar] [CrossRef]
- Yagi, M.; Miyamoto, T.; Sawatani, Y.; Iwamoto, K.; Hosogane, N.; Fujita, N.; Morita, K.; Ninomiya, K.; Suzuki, T.; Miyamoto, K.; et al. DC-STAMP is essential for cell-cell fusion in osteoclasts and foreign body giant cells. J. Exp. Med. 2005, 202, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, B. Normal bone anatomy and physiology. Clin. J. Am. Soc. Nephrol. 2008, 3 (Suppl. 3), S131–S139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaananen, H.K.; Zhao, H.; Mulari, M.; Halleen, J.M. The cell biology of osteoclast function. J. Cell Sci. 2000, 113, 377–381. [Google Scholar]
- Henriksen, K.; Bollerslev, J.; Everts, V.; Karsdal, M.A. Osteoclast activity and subtypes as a function of physiology and pathology—Implications for future treatments of osteoporosis. Endocr. Rev. 2011, 32, 31–63. [Google Scholar] [CrossRef] [Green Version]
- Ono, T.; Nakashima, T. Recent advances in osteoclast biology. Histochem. Cell Biol. 2018, 149, 325–341. [Google Scholar] [CrossRef]
- Kim, N.; Takami, M.; Rho, J.; Josien, R.; Choi, Y. A novel member of the leukocyte receptor complex regulates osteoclast differentiation. J. Exp. Med. 2002, 195, 201–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.; Petretto, E.; Gordon, S.; Bassett, J.H.D.; Williams, G.R.; Behmoaras, J. Common signalling pathways in macrophage and osteoclast multinucleation. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capulli, M.; Paone, R.; Rucci, N. Osteoblast and osteocyte: Games without frontiers. Arch. Biochem. Biophys. 2014, 561, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Uda, Y.; Azab, E.; Sun, N.; Shi, C.; Pajevic, P.D. Osteocyte Mechanobiology. Curr. Osteoporos. Rep. 2017, 15, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Weitzmann, M.N. The Role of Inflammatory Cytokines, the RANKL/OPG Axis, and the Immunoskeletal Interface in Physiological Bone Turnover and Osteoporosis. Scientifica 2013, 2013, 125705. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Laurent, M.R.; Dubois, V.; Claessens, F.; O’Brien, C.A.; Bouillon, R.; Vanderschueren, D.; Manolagas, S.C. Estrogens and Androgens in Skeletal Physiology and Pathophysiology. Physiol. Rev. 2017, 97, 135–187. [Google Scholar] [CrossRef]
- Rachner, T.D.; Khosla, S.; Hofbauer, L.C. Osteoporosis: Now and the future. Lancet 2011, 377, 1276–1287. [Google Scholar] [CrossRef] [Green Version]
- Prisby, R.D. Mechanical, hormonal and metabolic influences on blood vessels, blood flow and bone. J. Endocrinol. 2017, 235, R77–R100. [Google Scholar] [CrossRef] [Green Version]
- Shanmugarajan, S.; Zhang, Y.; Moreno-Villanueva, M.; Clanton, R.; Rohde, L.H.; Ramesh, G.T.; Sibonga, J.D.; Wu, H. Combined Effects of Simulated Microgravity and Radiation Exposure on Osteoclast Cell Fusion. Int. J. Mol. Sci. 2017, 18, 2443. [Google Scholar] [CrossRef] [Green Version]
- Baxter, N.N.; Habermann, E.B.; Tepper, J.E.; Durham, S.B.; Virnig, B.A. Risk of pelvic fractures in older women following pelvic irradiation. JAMA 2005, 294, 2587–2593. [Google Scholar] [CrossRef] [Green Version]
- WHO. Fact Sheet: Ionizing Radiation, Health Effects and Protective Measures. Available online: https://www.who.int/news-room/fact-sheets/detail/ionizing-radiation-health-effects-and-protective-measures (accessed on 4 June 2020).
- BfS. Was Ist Ionisierende Strahlung? Available online: https://www.bfs.de/DE/themen/ion/einfuehrung/einfuehrung_node.html (accessed on 4 June 2020).
- Balagamwala, E.H.; Stockham, A.; Macklis, R.; Singh, A.D. Introduction to radiotherapy and standard teletherapy techniques. Dev. Ophthalmol. 2013, 52, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Derer, A.; Deloch, L.; Rubner, Y.; Fietkau, R.; Frey, B.; Gaipl, U.S. Radio-Immunotherapy-Induced Immunogenic Cancer Cells as Basis for Induction of Systemic Anti-Tumor Immune Responses—Pre-Clinical Evidence and Ongoing Clinical Applications. Front. Immunol. 2015, 6, 505. [Google Scholar] [CrossRef] [Green Version]
- Ruckert, M.; Deloch, L.; Fietkau, R.; Frey, B.; Hecht, M.; Gaipl, U.S. Immune modulatory effects of radiotherapy as basis for well-reasoned radioimmunotherapies. Strahlenther. Onkol. 2018, 194, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Z.; Wu, A.; Nie, J.; Pei, H.; Hu, W.; Wang, B.; Shang, P.; Li, B.; Zhou, G. Differences in responses to X-ray exposure between osteoclast and osteoblast cells. J. Radiat. Res. 2017, 58, 791–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaney, G.; Jacob, S.; Featherstone, C.; Barton, M. The role of radiotherapy in cancer treatment: Estimating optimal utilization from a review of evidence-based clinical guidelines. Cancer 2005, 104, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Haussmann, J.; Tamaskovics, B.; Bolke, E.; Djiepmo-Njanang, F.J.; Kammers, K.; Corradini, S.; Hautmann, M.; Ghadjar, P.; Maas, K.; Schuler, P.J.; et al. Addition of chemotherapy to hyperfractionated radiotherapy in advanced head and neck cancer-a meta-analysis. Strahlenther. Onkol. 2019, 195, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.K.; Poortmans, P.; Chalmers, A.J.; Faivre-Finn, C.; Hall, E.; Huddart, R.A.; Lievens, Y.; Sebag-Montefiore, D.; Coles, C.E. Practice-changing radiation therapy trials for the treatment of cancer: Where are we 150 years after the birth of Marie Curie? Br. J. Cancer 2018, 119, 389–407. [Google Scholar] [CrossRef] [Green Version]
- Gouk, S.S.; Lim, T.M.; Teoh, S.H.; Sun, W.Q. Alterations of human acellular tissue matrix by gamma irradiation: Histology, biomechanical property, stability, in vitro cell repopulation, and remodeling. J. Biomed. Mater. Res. B Appl. Biomater. 2008, 84, 205–217. [Google Scholar] [CrossRef]
- Fideler, B.M.; Vangsness, C.T., Jr.; Lu, B.; Orlando, C.; Moore, T. Gamma irradiation: Effects on biomechanical properties of human bone-patellar tendon-bone allografts. Am. J. Sports Med. 1995, 23, 643–646. [Google Scholar] [CrossRef]
- Saito, M.; Marumo, K. Collagen cross-links as a determinant of bone quality: A possible explanation for bone fragility in aging, osteoporosis, and diabetes mellitus. Osteoporos. Int. 2010, 21, 195–214. [Google Scholar] [CrossRef]
- Darzy, K.H.; Shalet, S.M. Hypopituitarism after cranial irradiation. J. Endocrinol. Investig. 2005, 28, 78–87. [Google Scholar]
- Langlands, A.O.; Souter, W.A.; Samuel, E.; Redpath, A.T. Radiation osteitis following irradiation for breast cancer. Clin. Radiol. 1977, 28, 93–96. [Google Scholar] [CrossRef]
- Banfi, A.; Bianchi, G.; Galotto, M.; Cancedda, R.; Quarto, R. Bone marrow stromal damage after chemo/radiotherapy: Occurrence, consequences and possibilities of treatment. Leuk. Lymphoma 2001, 42, 863–870. [Google Scholar] [CrossRef]
- Ergun, H.; Howland, W.J. Postradiation atrophy of mature bone. CRC Crit. Rev. Diagn. Imaging 1980, 12, 225–243. [Google Scholar] [PubMed]
- Sakurai, T.; Sawada, Y.; Yoshimoto, M.; Kawai, M.; Miyakoshi, J. Radiation-induced reduction of osteoblast differentiation in C2C12 cells. J. Radiat. Res. 2007, 48, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Gal, T.J.; Munoz-Antonia, T.; Muro-Cacho, C.A.; Klotch, D.W. Radiation effects on osteoblasts in vitro: A potential role in osteoradionecrosis. Arch. Otolaryngol. Head Neck Surg. 2000, 126, 1124–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymczyk, K.H.; Shapiro, I.M.; Adams, C.S. Ionizing radiation sensitizes bone cells to apoptosis. Bone 2004, 34, 148–156. [Google Scholar] [CrossRef]
- Chandra, A.; Lin, T.; Tribble, M.B.; Zhu, J.; Altman, A.R.; Tseng, W.J.; Zhang, Y.; Akintoye, S.O.; Cengel, K.; Liu, X.S.; et al. PTH1-34 alleviates radiotherapy-induced local bone loss by improving osteoblast and osteocyte survival. Bone 2014, 67, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Chandra, A.; Lin, T.; Zhu, J.; Tong, W.; Huo, Y.; Jia, H.; Zhang, Y.; Liu, X.S.; Cengel, K.; Xia, B.; et al. PTH1-34 blocks radiation-induced osteoblast apoptosis by enhancing DNA repair through canonical Wnt pathway. J. Biol. Chem. 2015, 290, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Wright, L.E.; Buijs, J.T.; Kim, H.S.; Coats, L.E.; Scheidler, A.M.; John, S.K.; She, Y.; Murthy, S.; Ma, N.; Chin-Sinex, H.J.; et al. Single-Limb Irradiation Induces Local and Systemic Bone Loss in a Murine Model. J. Bone Miner. Res. 2015, 30, 1268–1279. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Li, C.; Yang, K.; Kang, H.; Xu, X.; Xu, X.; Deng, L. Increased EZH2 and decreased osteoblastogenesis during local irradiation-induced bone loss in rats. Sci. Rep. 2016, 6, 31318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, P.; Baumstark-Khan, C.; Hellweg, C.E.; Reitz, G. X-irradiation-induced cell cycle delay and DNA double-strand breaks in the murine osteoblastic cell line OCT-1. Radiat. Environ. Biophys. 2010, 49, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Xu, L.; Chen, M.; Mao, Y.T.; Xie, Z.G.; Wu, S.L.; Dong, Q.R. The effects of low dose X-irradiation on osteoblastic MC3T3-E1 cells in vitro. BMC Musculoskelet. Disord. 2012, 13, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Huang, Q.; Xu, W.; She, C.; Xie, Z.G.; Mao, Y.T.; Dong, Q.R.; Ling, M. Low-dose X-ray irradiation promotes osteoblast proliferation, differentiation and fracture healing. PLoS ONE 2014, 9, e104016. [Google Scholar] [CrossRef] [PubMed]
- Park, S.S.; Kim, K.A.; Lee, S.Y.; Lim, S.S.; Jeon, Y.M.; Lee, J.C. X-ray radiation at low doses stimulates differentiation and mineralization of mouse calvarial osteoblasts. BMB Rep. 2012, 45, 571–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zur Nieden, N.I.; Kempka, G.; Ahr, H.J. In vitro differentiation of embryonic stem cells into mineralized osteoblasts. Differentiation 2003, 71, 18–27. [Google Scholar] [CrossRef]
- Sawajiri, M.; Mizoe, J.; Tanimoto, K. Changes in osteoclasts after irradiation with carbon ion particles. Radiat. Environ. Biophys. 2003, 42, 219–223. [Google Scholar] [CrossRef]
- Goblirsch, M.; Lynch, C.; Mathews, W.; Manivel, J.C.; Mantyh, P.W.; Clohisy, D.R. Radiation treatment decreases bone cancer pain through direct effect on tumor cells. Radiat. Res. 2005, 164, 400–408. [Google Scholar] [CrossRef]
- Vit, J.P.; Ohara, P.T.; Tien, D.A.; Fike, J.R.; Eikmeier, L.; Beitz, A.; Wilcox, G.L.; Jasmin, L. The analgesic effect of low dose focal irradiation in a mouse model of bone cancer is associated with spinal changes in neuro-mediators of nociception. Pain 2006, 120, 188–201. [Google Scholar] [CrossRef]
- Anderson, N.D.; Colyer, R.A.; Riley, L.H., Jr. Skeletal changes during prolonged external irradiation: Alterations in marrow, growth plate and osteoclast populations. Johns Hopkins Med. J. 1979, 145, 73–83. [Google Scholar]
- Scheven, B.A.; Burger, E.H.; Kawilarang-de Haas, E.W.; Wassenaar, A.M.; Nijweide, P.J. Effects of ionizing irradiation on formation and resorbing activity of osteoclasts in vitro. Lab. Investig. 1985, 53, 72–79. [Google Scholar] [PubMed]
- Willey, J.S.; Lloyd, S.A.; Nelson, G.A.; Bateman, T.A. Ionizing Radiation and Bone Loss: Space Exploration and Clinical Therapy Applications. Clin. Rev. Bone Miner. Metab. 2011, 9, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alwood, J.S.; Shahnazari, M.; Chicana, B.; Schreurs, A.S.; Kumar, A.; Bartolini, A.; Shirazi-Fard, Y.; Globus, R.K. Ionizing Radiation Stimulates Expression of Pro-Osteoclastogenic Genes in Marrow and Skeletal Tissue. J. Interferon Cytokine Res. 2015, 35, 480–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, H.; Searby, N.D.; Mojarrab, R.; Phillips, J.; Alwood, J.; Yumoto, K.; Almeida, E.A.; Limoli, C.L.; Globus, R.K. Total-body irradiation of postpubertal mice with (137)Cs acutely compromises the microarchitecture of cancellous bone and increases osteoclasts. Radiat. Res. 2009, 171, 283–289. [Google Scholar] [CrossRef]
- Yang, B.; Zhou, H.; Zhang, X.D.; Liu, Z.; Fan, F.Y.; Sun, Y.M. Effect of radiation on the expression of osteoclast marker genes in RAW264.7 cells. Mol. Med. Rep. 2012, 5, 955–958. [Google Scholar] [CrossRef] [Green Version]
- Willey, J.S.; Lloyd, S.A.; Robbins, M.E.; Bourland, J.D.; Smith-Sielicki, H.; Bowman, L.C.; Norrdin, R.W.; Bateman, T.A. Early increase in osteoclast number in mice after whole-body irradiation with 2 Gy X rays. Radiat. Res. 2008, 170, 388–392. [Google Scholar] [CrossRef] [Green Version]
- Rodel, F.; Frey, B.; Gaipl, U.; Keilholz, L.; Fournier, C.; Manda, K.; Schollnberger, H.; Hildebrandt, G.; Rodel, C. Modulation of inflammatory immune reactions by low-dose ionizing radiation: Molecular mechanisms and clinical application. Curr. Med. Chem. 2012, 19, 1741–1750. [Google Scholar] [CrossRef]
- Schmid-Monnard, C. Über Heilung des Gelenkrheumatismus durch Röntgenstrahlung bei Kindern. Fortschr. Auf Dem Gebiet Röntgenstrahlung 1898, 1, 209. [Google Scholar]
- Schoen, R.; Böni, A.; Miehlke, K. Klinik der Rheumatischen Erkrankungen; Springer: Berlin/Heidelberg, Germany, 1970. [Google Scholar]
- Ott, O.J.; Niewald, M.; Weitmann, H.D.; Jacob, I.; Adamietz, I.A.; Schaefer, U.; Keilholz, L.; Heyd, R.; Muecke, R.; German Cooperative Group on Radiotherapy for Benign Diseases. DEGRO guidelines for the radiotherapy of non-malignant disorders. Part II: Painful degenerative skeletal disorders. Strahlenther. Onkol. 2015, 191, 1–6. [Google Scholar] [CrossRef]
- Ruhle, P.F.; Klein, G.; Rung, T.; Tiep Phan, H.; Fournier, C.; Fietkau, R.; Gaipl, U.S.; Frey, B. Impact of radon and combinatory radon/carbon dioxide spa on pain and hypertension: Results from the explorative RAD-ON01 study. Mod. Rheumatol. 2019, 29, 165–172. [Google Scholar] [CrossRef]
- Mucke, R.; Seegenschmiedt, M.H.; Heyd, R.; Schafer, U.; Prott, F.J.; Glatzel, M.; Micke, O.; German Cooperative Group on Radiotherapy for Benign Diseases. Radiotherapy in painful gonarthrosis. Results of a national patterns-of-care study. Strahlenther. Onkol. 2010, 186, 7–17. [Google Scholar] [CrossRef]
- Hautmann, M.G.; Rechner, P.; Neumaier, U.; Suss, C.; Dietl, B.; Putz, F.J.; Behr, M.; Kolbl, O.; Steger, F. Radiotherapy for osteoarthritis-an analysis of 295 joints treated with a linear accelerator. Strahlenther. Onkol. 2020, 196, 715–724. [Google Scholar] [CrossRef]
- Feldmann, M.; Brennan, F.M.; Maini, R.N. Rheumatoid arthritis. Cell 1996, 85, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Kullmann, M.; Ruhle, P.F.; Harrer, A.; Donaubauer, A.; Becker, I.; Sieber, R.; Klein, G.; Fournier, C.; Fietkau, R.; Gaipl, U.S.; et al. Temporarily increased TGFbeta following radon spa correlates with reduced pain while serum IL-18 is a general predictive marker for pain sensitivity. Radiat. Environ. Biophys. 2019, 58, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Rodel, F.; Keilholz, L.; Herrmann, M.; Sauer, R.; Hildebrandt, G. Radiobiological mechanisms in inflammatory diseases of low-dose radiation therapy. Int. J. Radiat. Biol. 2007, 83, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Wunderlich, R.; Ernst, A.; Rodel, F.; Fietkau, R.; Ott, O.; Lauber, K.; Frey, B.; Gaipl, U.S. Low and moderate doses of ionizing radiation up to 2 Gy modulate transmigration and chemotaxis of activated macrophages, provoke an anti-inflammatory cytokine milieu, but do not impact upon viability and phagocytic function. Clin. Exp. Immunol. 2015, 179, 50–61. [Google Scholar] [CrossRef] [Green Version]
- Deloch, L.; Derer, A.; Hueber, A.J.; Herrmann, M.; Schett, G.A.; Wolfelschneider, J.; Hahn, J.; Ruhle, P.F.; Stillkrieg, W.; Fuchs, J.; et al. Low-Dose Radiotherapy Ameliorates Advanced Arthritis in hTNF-alpha tg Mice by Particularly Positively Impacting on Bone Metabolism. Front. Immunol. 2018, 9, 1834. [Google Scholar] [CrossRef] [Green Version]
- Liebmann, A.; Hindemith, M.; Jahns, J.; Madaj-Sterba, P.; Weisheit, S.; Kamprad, F.; Hildebrandt, G. Low-dose X-irradiation of adjuvant-induced arthritis in rats. Efficacy of different fractionation schedules. Strahlenther. Onkol. 2004, 180, 165–172. [Google Scholar] [CrossRef]
- Schaue, D.; Jahns, J.; Hildebrandt, G.; Trott, K.R. Radiation treatment of acute inflammation in mice. Int. J. Radiat. Biol. 2005, 81, 657–667. [Google Scholar] [CrossRef]
- Arenas, M.; Gil, F.; Gironella, M.; Hernandez, V.; Jorcano, S.; Biete, A.; Pique, J.M.; Panes, J. Anti-inflammatory effects of low-dose radiotherapy in an experimental model of systemic inflammation in mice. Int. J. Radiat. Oncol. Biol. Phys. 2006, 66, 560–567. [Google Scholar] [CrossRef]
- Frey, B.; Gaipl, U.S.; Sarter, K.; Zaiss, M.M.; Stillkrieg, W.; Rodel, F.; Schett, G.; Herrmann, M.; Fietkau, R.; Keilholz, L. Whole body low dose irradiation improves the course of beginning polyarthritis in human TNF-transgenic mice. Autoimmunity 2009, 42, 346–348. [Google Scholar] [CrossRef] [PubMed]
- Ruhle, P.F.; Wunderlich, R.; Deloch, L.; Fournier, C.; Maier, A.; Klein, G.; Fietkau, R.; Gaipl, U.S.; Frey, B. Modulation of the peripheral immune system after low-dose radon spa therapy: Detailed longitudinal immune monitoring of patients within the RAD-ON01 study. Autoimmunity 2017, 50, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, S.; Eberle, B.; Vogt, D.R.; Meier, E.; Moser, L.; Gomez Ordonez, S.; Desborough, S.; Riesterer, O.; Takacs, I.; Hasler, P.; et al. Prospective Evaluation of Changes in Pain Levels, Quality of Life and Functionality After Low Dose Radiotherapy for Epicondylitis, Plantar Fasciitis, and Finger Osteoarthritis. Front. Med. 2020, 7. [Google Scholar] [CrossRef]
- Donaubauer, A.J.; Zhou, J.G.; Ott, O.J.; Putz, F.; Fietkau, R.; Keilholz, L.; Gaipl, U.S.; Frey, B.; Weissmann, T. Low Dose Radiation Therapy, Particularly with 0.5 Gy, Improves Pain in Degenerative Joint Disease of the Fingers: Results of a Retrospective Analysis. Int. J. Mol. Sci. 2020, 21, 5854. [Google Scholar] [CrossRef]
- Zhou, X.Z.; Zhang, G.; Dong, Q.R.; Chan, C.W.; Liu, C.F.; Qin, L. Low-dose X-irradiation promotes mineralization of fracture callus in a rat model. Arch. Orthop. Trauma Surg. 2009, 129, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Deloch, L.; Ruckert, M.; Fietkau, R.; Frey, B.; Gaipl, U.S. Low-Dose Radiotherapy Has No Harmful Effects on Key Cells of Healthy Non-Inflamed Joints. Int. J. Mol. Sci. 2018, 19, 3197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pramojanee, S.N.; Pratchayasakul, W.; Chattipakorn, N.; Chattipakorn, S.C. Low-dose dental irradiation decreases oxidative stress in osteoblastic MC3T3-E1 cells without any changes in cell viability, cellular proliferation and cellular apoptosis. Arch. Oral Biol. 2012, 57, 252–256. [Google Scholar] [CrossRef]
- Kim, M.S.; Yang, Y.M.; Son, A.; Tian, Y.S.; Lee, S.I.; Kang, S.W.; Muallem, S.; Shin, D.M. RANKL-mediated reactive oxygen species pathway that induces long lasting Ca2+ oscillations essential for osteoclastogenesis. J. Biol. Chem. 2010, 285, 6913–6921. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.J.; Kim, S.E.; Yun, Y.P.; Hwang, Y.S.; Bang, J.B.; Park, J.H.; Kwon, I.K. Simvastatin inhibits osteoclast differentiation by scavenging reactive oxygen species. Exp. Mol. Med. 2011, 43, 605–612. [Google Scholar] [CrossRef]
- Gravallese, E.M.; Harada, Y.; Wang, J.T.; Gorn, A.H.; Thornhill, T.S.; Goldring, S.R. Identification of cell types responsible for bone resorption in rheumatoid arthritis and juvenile rheumatoid arthritis. Am. J. Pathol. 1998, 152, 943–951. [Google Scholar]
- Cucu, A.; Shreder, K.; Kraft, D.; Ruhle, P.F.; Klein, G.; Thiel, G.; Frey, B.; Gaipl, U.S.; Fournier, C. Decrease of Markers Related to Bone Erosion in Serum of Patients with Musculoskeletal Disorders after Serial Low-Dose Radon Spa Therapy. Front. Immunol. 2017, 8, 882. [Google Scholar] [CrossRef]
- Townsend, L.W.; Fry, R.J. Radiation protection guidance for activities in low-Earth orbit. Adv. Space Res. 2002, 30, 957–963. [Google Scholar] [CrossRef]
- Durante, M.; Cucinotta, F.A. Heavy ion carcinogenesis and human space exploration. Nat. Rev. Cancer 2008, 8, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.W. Cancer and radiation therapy: Current advances and future directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chancellor, J.C.; Scott, G.B.; Sutton, J.P. Space Radiation: The Number One Risk to Astronaut Health beyond Low Earth Orbit. Life 2014, 4, 491–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alwood, J.S.; Ronca, A.E.; Mains, R.C.; Shelhamer, M.J.; Smith, J.D.; Goodwin, T.J. From the bench to exploration medicine: NASA life sciences translational research for human exploration and habitation missions. NPJ Microgravity 2017, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Moreno-Villanueva, M.; Krieger, S.; Ramesh, G.T.; Neelam, S.; Wu, H. Transcriptomics, NF-kappaB Pathway, and Their Potential Spaceflight-Related Health Consequences. Int. J. Mol. Sci. 2017, 18, 1166. [Google Scholar] [CrossRef]
- Grimm, D.; Grosse, J.; Wehland, M.; Mann, V.; Reseland, J.E.; Sundaresan, A.; Corydon, T.J. The impact of microgravity on bone in humans. Bone 2016, 87, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Willey, J.S.; Livingston, E.W.; Robbins, M.E.; Bourland, J.D.; Tirado-Lee, L.; Smith-Sielicki, H.; Bateman, T.A. Risedronate prevents early radiation-induced osteoporosis in mice at multiple skeletal locations. Bone 2010, 46, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, S.A.; Pecaut, M.J.; Gridley, D.S.; Travis, N.D.; Bandstra, E.R.; Willey, J.S.; Nelson, G.A.; Bateman, T.A. A murine model for bone loss from therapeutic and space-relevant sources of radiation. J. Appl. Physiol. (1985) 2006, 101, 789–793. [Google Scholar] [CrossRef]
- Bandstra, E.R.; Pecaut, M.J.; Anderson, E.R.; Willey, J.S.; De Carlo, F.; Stock, S.R.; Gridley, D.S.; Nelson, G.A.; Levine, H.G.; Bateman, T.A. Long-term dose response of trabecular bone in mice to proton radiation. Radiat. Res. 2008, 169, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Bandstra, E.R.; Thompson, R.W.; Nelson, G.A.; Willey, J.S.; Judex, S.; Cairns, M.A.; Benton, E.R.; Vazquez, M.E.; Carson, J.A.; Bateman, T.A. Musculoskeletal changes in mice from 20–50 cGy of simulated galactic cosmic rays. Radiat. Res. 2009, 172, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Willey, J.S.; Lloyd, S.A.; Nelson, G.A.; Bateman, T.A. Space Radiation and Bone Loss. Gravit. Space Biol. Bull. 2011, 25, 14–21. [Google Scholar] [PubMed]
- Yumoto, K.; Globus, R.K.; Mojarrab, R.; Arakaki, J.; Wang, A.; Searby, N.D.; Almeida, E.A.; Limoli, C.L. Short-term effects of whole-body exposure to (56)fe ions in combination with musculoskeletal disuse on bone cells. Radiat. Res. 2010, 173, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Sambandam, Y.; Baird, K.L.; Stroebel, M.; Kowal, E.; Balasubramanian, S.; Reddy, S.V. Microgravity Induction of TRAIL Expression in Preosteoclast Cells Enhances Osteoclast Differentiation. Sci. Rep. 2016, 6, 25143. [Google Scholar] [CrossRef] [Green Version]
- Chatani, M.; Mantoku, A.; Takeyama, K.; Abduweli, D.; Sugamori, Y.; Aoki, K.; Ohya, K.; Suzuki, H.; Uchida, S.; Sakimura, T.; et al. Microgravity promotes osteoclast activity in medaka fish reared at the international space station. Sci. Rep. 2015, 5, 14172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes-Fulford, M.; Tjandrawinata, R.; Fitzgerald, J.; Gasuad, K.; Gilbertson, V. Effects of microgravity on osteoblast growth. Gravit. Space Biol. Bull. 1998, 11, 51–60. [Google Scholar]
- Kondo, H.; Yumoto, K.; Alwood, J.S.; Mojarrab, R.; Wang, A.; Almeida, E.A.; Searby, N.D.; Limoli, C.L.; Globus, R.K. Oxidative stress and gamma radiation-induced cancellous bone loss with musculoskeletal disuse. J. Appl. Physiol. (1985) 2010, 108, 152–161. [Google Scholar] [CrossRef] [Green Version]
- Macias, B.R.; Lima, F.; Swift, J.M.; Shirazi-Fard, Y.; Greene, E.S.; Allen, M.R.; Fluckey, J.; Hogan, H.A.; Braby, L.; Wang, S.; et al. Simulating the Lunar Environment: Partial Weightbearing and High-LET Radiation-Induce Bone Loss and Increase Sclerostin-Positive Osteocytes. Radiat. Res. 2016, 186, 254–263. [Google Scholar] [CrossRef]
- Choy, M.H.V.; Wong, R.M.Y.; Chow, S.K.H.; Li, M.C.; Chim, Y.N.; Li, T.K.; Ho, W.T.; Cheng, J.C.Y.; Cheung, W.H. How much do we know about the role of osteocytes in different phases of fracture healing? A systematic review. J. Orthop. Translat. 2020, 21, 111–121. [Google Scholar] [CrossRef]
- He, F.; Bai, J.; Wang, J.; Zhai, J.; Tong, L.; Zhu, G. Irradiation-induced osteocyte damage promotes HMGB1-mediated osteoclastogenesis in vitro. J. Cell Physiol. 2019, 234, 17314–17325. [Google Scholar] [CrossRef] [PubMed]
- Mendes, E.M.; Irie, M.S.; Rabelo, G.D.; Borges, J.S.; Dechichi, P.; Diniz, R.S.; Soares, P.B.F. Effects of ionizing radiation on woven bone: Influence on the osteocyte lacunar network, collagen maturation, and microarchitecture. Clin. Oral Investig. 2019. [Google Scholar] [CrossRef] [PubMed]
- Bellido, T. Osteocyte-driven bone remodeling. Calcif. Tissue Int. 2014, 94, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villars, F.; Guillotin, B.; Amedee, T.; Dutoya, S.; Bordenave, L.; Bareille, R.; Amedee, J. Effect of HUVEC on human osteoprogenitor cell differentiation needs heterotypic gap junction communication. Am. J. Physiol. Cell Physiol. 2002, 282, C775–C785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewing, J. Radiation osteitis. Acta Radiol. 1926, 6, 399–412. [Google Scholar] [CrossRef]
- Pitkanen, M.A.; Hopewell, J.W. Functional changes in the vascularity of the irradiated rat femur. Implications for late effects. Acta Radiol. Oncol. 1983, 22, 253–256. [Google Scholar] [CrossRef] [Green Version]
- Okunieff, P.; Wang, X.; Rubin, P.; Finkelstein, J.N.; Constine, L.S.; Ding, I. Radiation-induced changes in bone perfusion and angiogenesis. Int. J. Radiat. Oncol. Biol. Phys. 1998, 42, 885–889. [Google Scholar] [CrossRef]
- Kaigler, D.; Wang, Z.; Horger, K.; Mooney, D.J.; Krebsbach, P.H. VEGF scaffolds enhance angiogenesis and bone regeneration in irradiated osseous defects. J. Bone Miner. Res. 2006, 21, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Michel, G.; Blery, P.; Pilet, P.; Guicheux, J.; Weiss, P.; Malard, O.; Espitalier, F. Micro-CT Analysis of Radiation-Induced Osteopenia and Bone Hypovascularization in Rat. Calcif. Tissue Int. 2015, 97, 62–68. [Google Scholar] [CrossRef]
- Akmansu, M.; Unsal, D.; Bora, H.; Elbeg, S. Influence of locoregional radiation treatment on tumor necrosis factor-alpha and interleukin-6 in the serum of patients with head and neck cancer. Cytokine 2005, 31, 41–45. [Google Scholar] [CrossRef]
- Xu, R. Semaphorin 3A: A new player in bone remodeling. Cell Adh. Migr. 2014, 8, 5–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayanagi, H. Osteoimmunology: Shared mechanisms and crosstalk between the immune and bone systems. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Kostenuik, P.J.; Shalhoub, V. Osteoprotegerin: A physiological and pharmacological inhibitor of bone resorption. Curr. Pharm. Des. 2001, 7, 613–635. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Huang, Y.; Collin-Osdoby, P.; Osdoby, P. CCR1 chemokines promote the chemotactic recruitment, RANKL development, and motility of osteoclasts and are induced by inflammatory cytokines in osteoblasts. J. Bone Miner. Res. 2004, 19, 2065–2077. [Google Scholar] [CrossRef]
- Kim, M.S.; Day, C.J.; Morrison, N.A. MCP-1 is induced by receptor activator of nuclear factor-{kappa}B ligand, promotes human osteoclast fusion, and rescues granulocyte macrophage colony-stimulating factor suppression of osteoclast formation. J. Biol. Chem. 2005, 280, 16163–16169. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Zhang, Q.; Yuan, Y.; Xin, N.; He, K.; Huang, Y.; Tang, H.; Gong, P. Sema3a inhibits the differentiation of Raw264.7 cells to osteoclasts under 2Gy radiation by reducing inflammation. PLoS ONE 2018, 13, e0200000. [Google Scholar] [CrossRef] [Green Version]
- Iaquinta, M.R.; Mazzoni, E.; Bononi, I.; Rotondo, J.C.; Mazziotta, C.; Montesi, M.; Sprio, S.; Tampieri, A.; Tognon, M.; Martini, F. Adult Stem Cells for Bone Regeneration and Repair. Front. Cell Dev. Biol. 2019, 7, 268. [Google Scholar] [CrossRef] [Green Version]
- Shupp, A.B.; Kolb, A.D.; Mukhopadhyay, D.; Bussard, K.M. Cancer Metastases to Bone: Concepts, Mechanisms, and Interactions with Bone Osteoblasts. Cancers 2018, 10, 182. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; So, Y.H.; Cui, J.; Ma, K.; Xu, X.; Zhao, Y.; Cai, L.; Li, W. The low-dose ionizing radiation stimulates cell proliferation via activation of the MAPK/ERK pathway in rat cultured mesenchymal stem cells. J. Radiat. Res. 2011, 52, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Kwong, D.L.; Chan, G.C. The effects of various irradiation doses on the growth and differentiation of marrow-derived human mesenchymal stromal cells. Pediatr. Transplant. 2007, 11, 379–387. [Google Scholar] [CrossRef]
- Russu, I.Z.; Rodionova, N.K.; Bilko, D.I.; Bilko, N.M. Mesenchymal stem and progenitor cells of rats’ bone marrow under chronic action of ionizing radiation. Probl. Radiac. Med. Radiobiol. 2017, 22, 224–230. [Google Scholar] [CrossRef]
- Muralidharan, S.; Sasi, S.P.; Zuriaga, M.A.; Hirschi, K.K.; Porada, C.D.; Coleman, M.A.; Walsh, K.X.; Yan, X.; Goukassian, D.A. Ionizing Particle Radiation as a Modulator of Endogenous Bone Marrow Cell Reprogramming: Implications for Hematological Cancers. Front. Oncol. 2015, 5, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Wang, G.; Cui, J.; Xue, L.; Cai, L. Low-dose radiation (LDR) induces hematopoietic hormesis: LDR-induced mobilization of hematopoietic progenitor cells into peripheral blood circulation. Exp. Hematol. 2004, 32, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- Schaue, D.; McBride, W.H. Links between innate immunity and normal tissue radiobiology. Radiat. Res. 2010, 173, 406–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchwald, Z.S.; Aurora, R. Osteoclasts and CD8 T cells form a negative feedback loop that contributes to homeostasis of both the skeletal and immune systems. Clin. Dev. Immunol. 2013, 2013, 429373. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.; Hong, W.; Zhou, Y.; Ding, Q.; Wang, J.; Jin, W.; Gao, J.; Hua, G.; Xu, X. Bone marrow stem cell dysfunction in radiation-induced abscopal bone loss. J. Orthop. Surg. Res. 2016, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Conget, P.A.; Minguell, J.J. Phenotypical and functional properties of human bone marrow mesenchymal progenitor cells. J. Cell Physiol. 1999, 181, 67–73. [Google Scholar] [CrossRef]
- Meirow, D.; Nugent, D. The effects of radiotherapy and chemotherapy on female reproduction. Hum. Reprod. Update 2001, 7, 535–543. [Google Scholar] [CrossRef] [Green Version]
- Hui, S.K.; Khalil, A.; Zhang, Y.; Coghill, K.; Le, C.; Dusenbery, K.; Froelich, J.; Yee, D.; Downs, L. Longitudinal assessment of bone loss from diagnostic computed tomography scans in gynecologic cancer patients treated with chemotherapy and radiation. Am. J. Obstet. Gynecol. 2010, 203, 353.e1–353.e7. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donaubauer, A.-J.; Deloch, L.; Becker, I.; Fietkau, R.; Frey, B.; Gaipl, U.S. The Influence of Radiation on Bone and Bone Cells—Differential Effects on Osteoclasts and Osteoblasts. Int. J. Mol. Sci. 2020, 21, 6377. https://doi.org/10.3390/ijms21176377
Donaubauer A-J, Deloch L, Becker I, Fietkau R, Frey B, Gaipl US. The Influence of Radiation on Bone and Bone Cells—Differential Effects on Osteoclasts and Osteoblasts. International Journal of Molecular Sciences. 2020; 21(17):6377. https://doi.org/10.3390/ijms21176377
Chicago/Turabian StyleDonaubauer, Anna-Jasmina, Lisa Deloch, Ina Becker, Rainer Fietkau, Benjamin Frey, and Udo S. Gaipl. 2020. "The Influence of Radiation on Bone and Bone Cells—Differential Effects on Osteoclasts and Osteoblasts" International Journal of Molecular Sciences 21, no. 17: 6377. https://doi.org/10.3390/ijms21176377
APA StyleDonaubauer, A. -J., Deloch, L., Becker, I., Fietkau, R., Frey, B., & Gaipl, U. S. (2020). The Influence of Radiation on Bone and Bone Cells—Differential Effects on Osteoclasts and Osteoblasts. International Journal of Molecular Sciences, 21(17), 6377. https://doi.org/10.3390/ijms21176377