Develop a High-Throughput Screening Method to Identify C-P4H1 (Collagen Prolyl 4-Hydroxylase 1) Inhibitors from FDA-Approved Chemicals



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
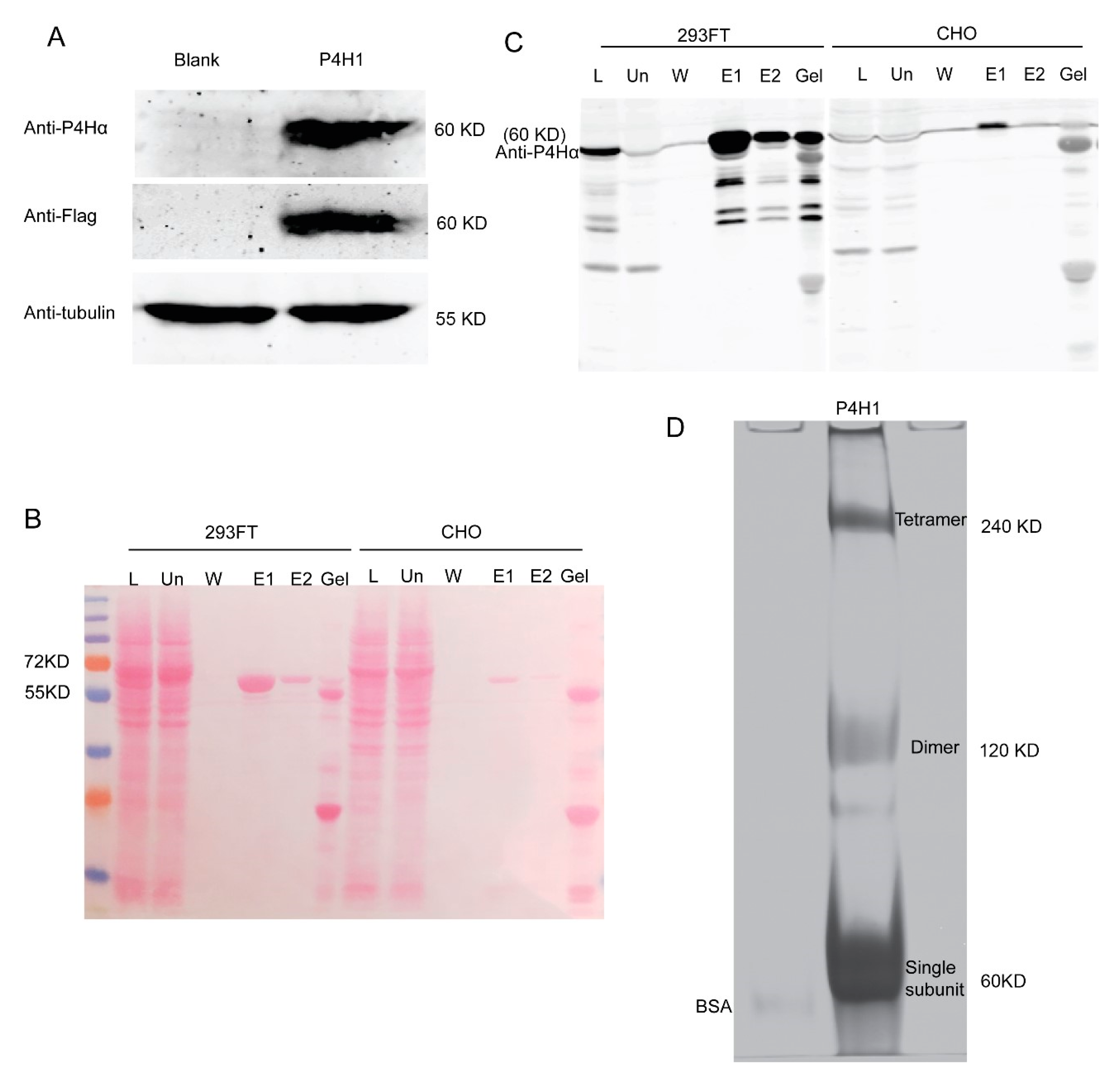
2.1. Protein Expression and Purification
2.2. Screening Method Confirmation
2.3. Inhibitory Screening and Verification
3. Discussion
4. Martials and Methods
4.1. Cell Culture and 3D Culture Assay
4.2. Cell Viability Assay
4.3. CP4H1 Expression and Purification
4.4. Western Blot
4.5. Measuring Human CP4H1 Activity with Hydroxyproline Colorimetric Assay and Succinate-GloTM Hydroxylase Assay
4.5.1. Human CP4H1 Activity Assay
4.5.2. Hydroxyproline Colorimetric Assay
4.5.3. Succinate-GloTM Hydroxylase Assay
4.6. High-Throughput Screening
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Myllyharju, J. Collagens, modifying enzymes and their mutations in humans, flies and worms. Trends Genet. 2004, 20, 33–43. [Google Scholar] [CrossRef]
- Levental, K.R.; Yu, H.; Kass, L.; Lakins, J.N.; Egeblad, M.; Erler, J.; Fong, S.F.; Csiszar, K.; Giaccia, A.; Weninger, W.; et al. Matrix crosslinking forces tumor progression by enhancing integrin signaling. Cell 2009, 139, 891–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provenzano, P.P.; Inman, D.R.; Eliceiri, K.W.; Knittel, J.G.; Yan, L.; Rueden, C.T.; White, J.; Keely, P.J. Collagen density promotes mammary tumor initiation and progression. BMC Med. 2008, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Specks, U.; Nerlich, A.G.; Colby, T.V.; Wiest, I.; Timpl, R. Increased expression of type VI collagen in lung fibrosis. Am. J. Respir. Crit. Care Med. 1995, 151, 1956–1964. [Google Scholar] [CrossRef] [PubMed]
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Boil. 2010, 3, a004978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, C.L.; Bretscher, L.E.; Guzei, I.A.; Raines, R.T. Effect of 3-hydroxyproline residues on collagen stability. J. Am. Chem. Soc. 2003, 125, 6422–6427. [Google Scholar] [CrossRef] [PubMed]
- Vasta, J.D.; Choudhary, A.; Jensen, K.H.; McGrath, N.A.; Raines, R.T. Prolyl 4-hydroxylase: Substrate isosteres in which an (E)—Or (Z)-alkene replaces the prolyl peptide bond. Biochemistry 2016, 56, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anantharajan, J.; Koski, M.K.; Kursula, P.; Hieta, R.; Bergmann, U.; Myllyharju, J.; Wierenga, R.K. The structural motifs for substrate binding and dimerization of the α subunit of collagen prolyl 4-hydroxylase. Structure 2013, 21, 2107–2118. [Google Scholar] [CrossRef] [Green Version]
- Annunen, P.; Autio-Harmainen, H.; Kivirikko, K.I. The novel type II prolyl 4-hydroxylase is the main enzyme form in chondrocytes and capillary endothelial cells, whereas the type I enzyme predominates in most cells. J. Boil. Chem. 1998, 273, 5989–5992. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.-H.; Ji, C.-D.; Xiao, H.-L.; Zhao, H.-B.; Cui, Y.-H.; Bian, X.-W. Reorganized collagen in the tumor microenvironment of gastric cancer and its association with prognosis. J. Cancer 2017, 8, 1466–1476. [Google Scholar] [CrossRef] [Green Version]
- Cloos, P.; Christgau, S.; Lyubimova, N.; Body, J.-J.; Qvist, P.; Christiansen, C. Breast cancer patients with bone metastases are characterised by increased levels of nonisomerised type I collagen fragments. Breast Cancer Res. 2003, 5, R103–R109. [Google Scholar] [CrossRef]
- Ewald, J.A.; Downs, T.M.; Cetnar, J.P.; Ricke, W.A. Expression microarray meta-analysis identifies genes associated with Ras/MAPK and related pathways in progression of muscle-invasive bladder transition cell carcinoma. PLoS ONE 2013, 8, e55414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Li, L.; Ye, H.; Tao, H.; He, H. Role of COL6A3 in colorectal cancer. Oncol. Rep. 2018, 39, 2527–2536. [Google Scholar] [CrossRef] [PubMed]
- Carafoli, F.; Hohenester, E. Collagen recognition and transmembrane signalling by discoidin domain receptors. Biochim. Biophys. Acta (BBA) 2012, 1834, 2187–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Fredericks, T.I.; Xiong, G.; Qi, Y.; Rychahou, P.; Li, J.-D.; Pihlajaniemi, T.; Xu, W.; Xu, R. Membrane associated collagen XIII promotes cancer metastasis and enhances anoikis resistance. Breast Cancer Res. 2018, 20, 116. [Google Scholar] [CrossRef] [PubMed]
- Hanker, A.B.; Estrada, M.V.; Bianchini, G.; Moore, P.D.; Zhao, J.; Cheng, F.; Koch, J.P.; Gianni, L.; Tyson, D.R.; Sanchez, V.; et al. Extracellular matrix/integrin signaling promotes resistance to combined inhibition of HER2 and PI3K in HER2+ breast cancer. Cancer Res. 2017, 77, 3280–3292. [Google Scholar] [CrossRef] [Green Version]
- Lai, S.L.; Tan, M.L.; Hollows, R.J.; Robinson, M.; Ibrahim, M.; Margielewska, S.; Parkinson, E.K.; Ramanathan, A.; Zain, R.B.; Mehanna, H.; et al. Collagen induces a more proliferative, migratory and chemoresistant phenotype in head and neck cancer via ddr1. Cancers 2019, 11, 1766. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-C.; Kurtova, A.V.; Xiao, J.; Nikolos, F.; Hayashi, K.; Tramel, Z.; Jain, A.; Chen, F.; Chokshi, M.; Lee, C.M.; et al. Collagen-rich airway smooth muscle cells are a metastatic niche for tumor colonization in the lung. Nat. Commun. 2019, 10, 2131. [Google Scholar] [CrossRef]
- Xiong, G.; Stewart, R.L.; Chen, J.; Gao, T.; Scott, T.L.; Samayoa, L.M.; O’Connor, K.; Lane, A.N.; Xu, R. Collagen prolyl 4-hydroxylase 1 is essential for HIF-1α stabilization and TNBC chemoresistance. Nat. Commun. 2018, 9, 4456. [Google Scholar] [CrossRef] [Green Version]
- Bickel, M.; Baringhaus, K.-H.; Gerl, M.; Günzler, V.; Kanta, J.; Schmidts, L.; Stapf, M.; Tschank, G.; Weidmann, K.; Werner, U. Selective inhibition of hepatic collagen accumulation in experimental liver fibrosis in rats by a new prolyl 4-hydroxylase inhibitor. Hepatology 1998, 28, 404–411. [Google Scholar] [CrossRef]
- Cao, X.-Q.; Liu, X.-X.; Li, M.-M.; Zhang, Y.; Chen, L.; Wang, L.; Di, M.-X.; Zhang, M. Overexpression of prolyl-4-hydroxylase-α1 stabilizes but increases shear stress-induced atherosclerotic plaque in apolipoprotein E-deficient mice. Dis. Markers 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gorres, K.L.; Raines, R.T. Direct and continuous assay for prolyl 4-hydroxylase. Anal. Biochem. 2009, 386, 181–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasta, J.D.; Andersen, K.A.; Deck, K.M.; Nizzi, C.P.; Eisenstein, R.S.; Raines, R.T. Selective inhibition of collagen prolyl 4-hydroxylase in human cells. ACS Chem. Boil. 2015, 11, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasta, J.D.; Raines, R.T. Selective inhibition of prolyl 4-hydroxylases by bipyridinedicarboxylates. Bioorg. Med. Chem. 2015, 23, 3081–3090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koivu, J.; Myllylä, R. Protein disulfide-isomerase retains procollagen prolyl 4-hydroxylase structure in its native conformation. Biochemistry 1986, 25, 5982–5986. [Google Scholar] [CrossRef]
- Vuori, K.; Pihlajaniemi, T.; Marttila, M.; Kivirikko, K.I. Characterization of the human prolyl 4-hydroxylase tetramer and its multifunctional protein disulfide-isomerase subunit synthesized in a baculovirus expression system. Proc. Natl. Acad. Sci. USA 1992, 89, 7467–7470. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, A.; Neubauer, P.; Myllyharju, J. High-level production of human collagen prolyl 4-hydroxylase in Escherichia coli. Matrix Boil. 2005, 24, 59–68. [Google Scholar] [CrossRef]
- Baldi, L.; Hacker, D.L.; Adam, M.; Wurm, F.M. Recombinant protein production by large-scale transient gene expression in mammalian cells: State of the art and future perspectives. Biotechnol. Lett. 2007, 29, 677–684. [Google Scholar] [CrossRef] [Green Version]
- Neuman, R.E.; Logan, M.A. The determination of hydroxyproline. J. Boil. Chem. 1950, 184, 299–306. [Google Scholar]
- Stegemann, H.; Stalder, K. Determination of hydroxyproline. Clin. Chim. Acta 1967, 18, 267–273. [Google Scholar] [CrossRef]
- Edwards, C.; O’Brien, W. Modified assay for determination of hydroxyproline in a tissue hydrolyzate. Clin. Chim. Acta 1980, 104, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.-F.; Cho, E.J.; Devkota, A.K.; Chen, Y.; Russell, W.; Phillips, G.N.; Yamauchi, M.; Dalby, K.N.; Kurie, J.M. A scalable lysyl hydroxylase 2 expression system and luciferase-based enzymatic activity assay. Arch. Biochem. Biophys. 2017, 618, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamberg, A.; Pihlajaniemi, T.; Kivirikko, I.K. Site-directed mutagenesis of the alpha subunit of human prolyl 4-hydroxylase. Identification of three histidine residues critical for catalytic activity. J. Boil. Chem. 1995, 270, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majamaa, K.; Günzler, V.; Hanauske-Abel, H.M.; Myllylä, R.; Kivirikko, K.I. Partial identity of the 2-oxoglutarate and ascorbate binding sites of prolyl 4-hydroxylase. J. Boil. Chem. 1986, 261, 7819–7823. [Google Scholar]
- Franklin, T.J.; Hitchen, M. Inhibition of collagen hydroxylation by 2,7,8-trihydroxyanthraquinone in embryonic-chick tendon cells. Biochem. J. 1989, 261, 127–130. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, J.J.; Pryszlak, M.; Smith, L.; Yanchus, C.; Kurji, N.; Shahani, V.M.; Molinski, S.V. Giving drugs a second chance: Overcoming regulatory and financial hurdles in repurposing approved drugs as cancer therapeutics. Front. Oncol. 2017, 7, 273. [Google Scholar] [CrossRef] [Green Version]
- Xiong, G.; Deng, L.; Zhu, J.; Rychahou, P.; Xu, R. Prolyl-4-hydroxylase α subunit 2 promotes breast cancer progression and metastasis by regulating collagen deposition. BMC Cancer 2014, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Chen, J.; Xiong, G.; Clair, D.K.S.; Xu, W.; Xu, R. Increased ROS production in non-polarized mammary epithelial cells induces monocyte infiltration in 3D culture. J. Cell Sci. 2016, 130, 190–202. [Google Scholar] [CrossRef] [Green Version]
- Kenny, P.A.; Lee, G.Y.; Myers, C.A.; Neve, R.M.; Semeiks, J.R.; Spellman, P.T.; Lorenz, K.; Lee, E.H.; Barcellos-Hoff, M.H.; Petersen, O.W.; et al. The morphologies of breast cancer cell lines in three-dimensional assays correlate with their profiles of gene expression. Mol. Oncol. 2007, 1, 84–96. [Google Scholar] [CrossRef]
- Kivirikko, K.I.; Kishida, Y.; Sakakibara, S.; Prockop, D.J. Hydroxylation of (X-Pro-Gly)n by protocollagen proline hydroxylase effect of chain length, helical conformation and amino acid sequence in the substrate. Biochim. Biophys. Acta (BBA) Protein Struct. 1972, 271, 347–356. [Google Scholar] [CrossRef]
- Manon--Jensen, T.; Kjeld, N.G.; Karsdal, M.A. Collagen-mediated hemostasis. J. Thromb. Haemost. 2016, 14, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, G.E.; Semus, S.F. 3D QSAR in modern drug design. Exp. Suppl. 2003, 93, 223–241. [Google Scholar] [CrossRef]
- Verma, J.; Khedkar, V.; Coutinho, E.C. 3D-QSAR in Drug Design—A Review. Curr. Top. Med. Chem. 2010, 10, 95–115. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Lee, K.-H.; Araujo, N.V.; Zhan, C.-G.; Rangnekar, V.M.; Xu, R. Develop a High-Throughput Screening Method to Identify C-P4H1 (Collagen Prolyl 4-Hydroxylase 1) Inhibitors from FDA-Approved Chemicals. Int. J. Mol. Sci. 2020, 21, 6613. https://doi.org/10.3390/ijms21186613
Wang S, Lee K-H, Araujo NV, Zhan C-G, Rangnekar VM, Xu R. Develop a High-Throughput Screening Method to Identify C-P4H1 (Collagen Prolyl 4-Hydroxylase 1) Inhibitors from FDA-Approved Chemicals. International Journal of Molecular Sciences. 2020; 21(18):6613. https://doi.org/10.3390/ijms21186613
Chicago/Turabian StyleWang, Shike, Kuo-Hao Lee, Nathalia Victoria Araujo, Chang-Guo Zhan, Vivek M. Rangnekar, and Ren Xu. 2020. "Develop a High-Throughput Screening Method to Identify C-P4H1 (Collagen Prolyl 4-Hydroxylase 1) Inhibitors from FDA-Approved Chemicals" International Journal of Molecular Sciences 21, no. 18: 6613. https://doi.org/10.3390/ijms21186613
APA StyleWang, S., Lee, K. -H., Araujo, N. V., Zhan, C. -G., Rangnekar, V. M., & Xu, R. (2020). Develop a High-Throughput Screening Method to Identify C-P4H1 (Collagen Prolyl 4-Hydroxylase 1) Inhibitors from FDA-Approved Chemicals. International Journal of Molecular Sciences, 21(18), 6613. https://doi.org/10.3390/ijms21186613