Applying Single-Cell Analysis to Gonadogenesis and DSDs (Disorders/Differences of Sex Development)


Abstract
:
1. Introduction
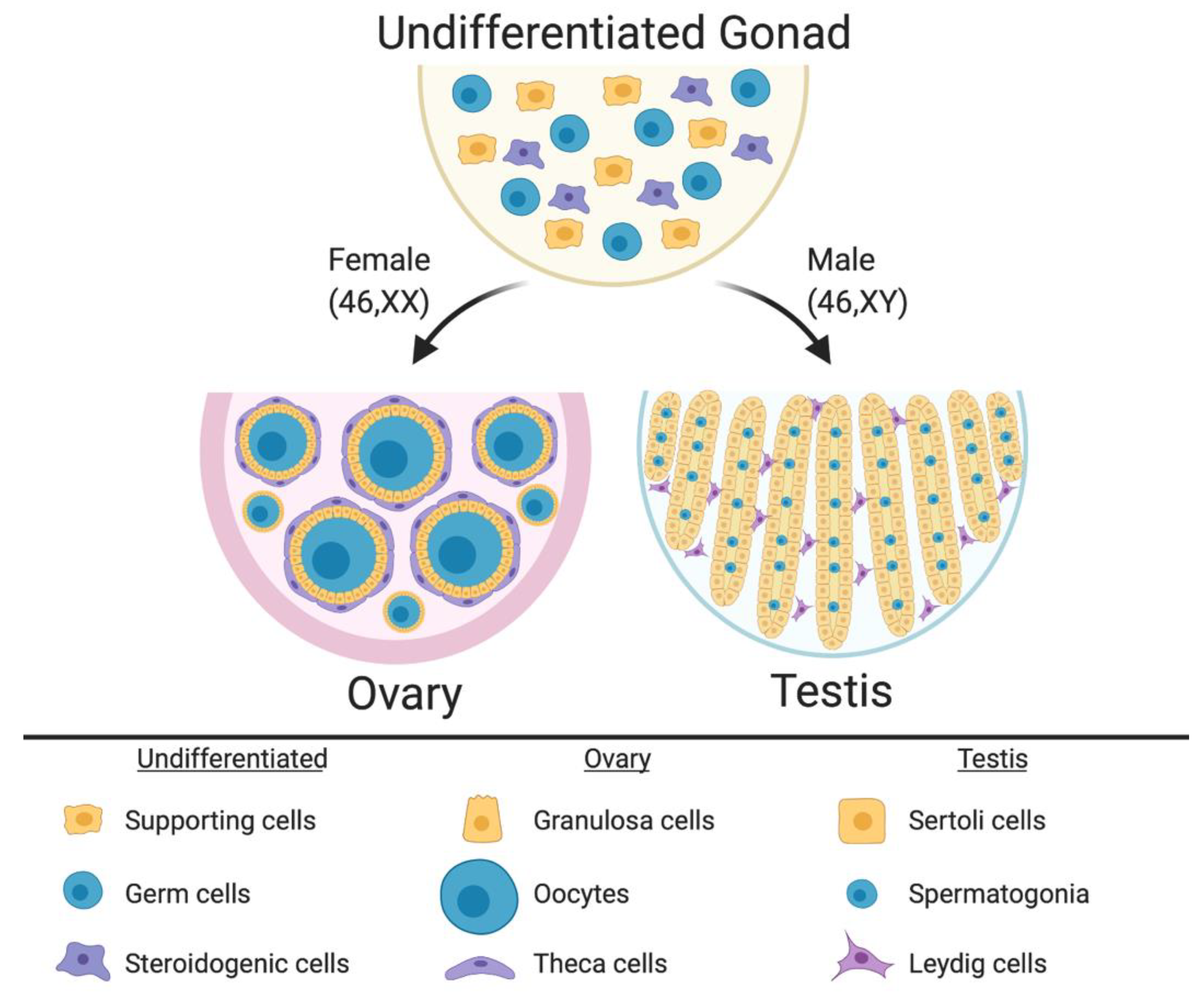
2. Gonadal Sex Differentiation
3. Disorders of Sex Development
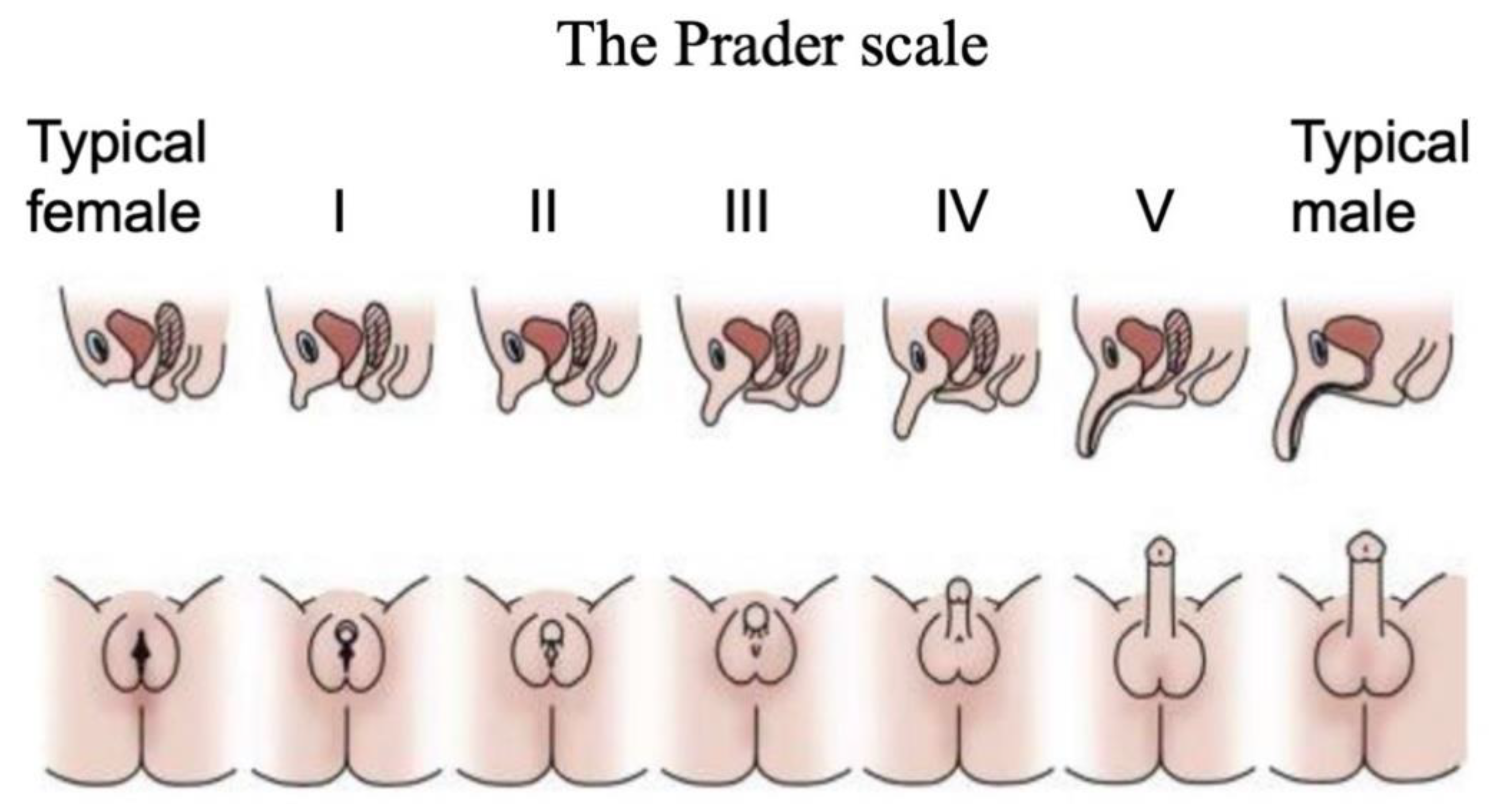
4. Diagnosis of DSD
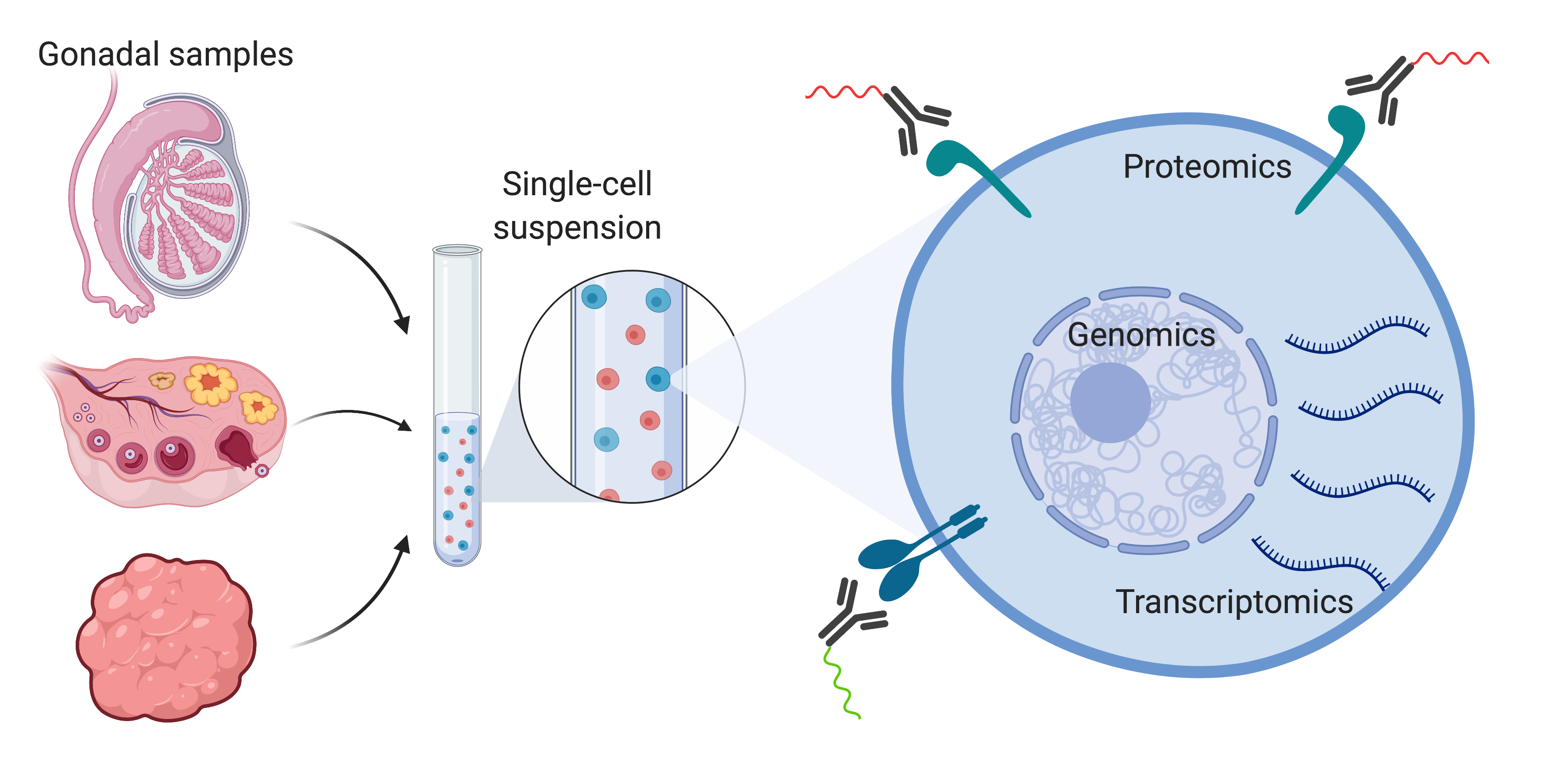
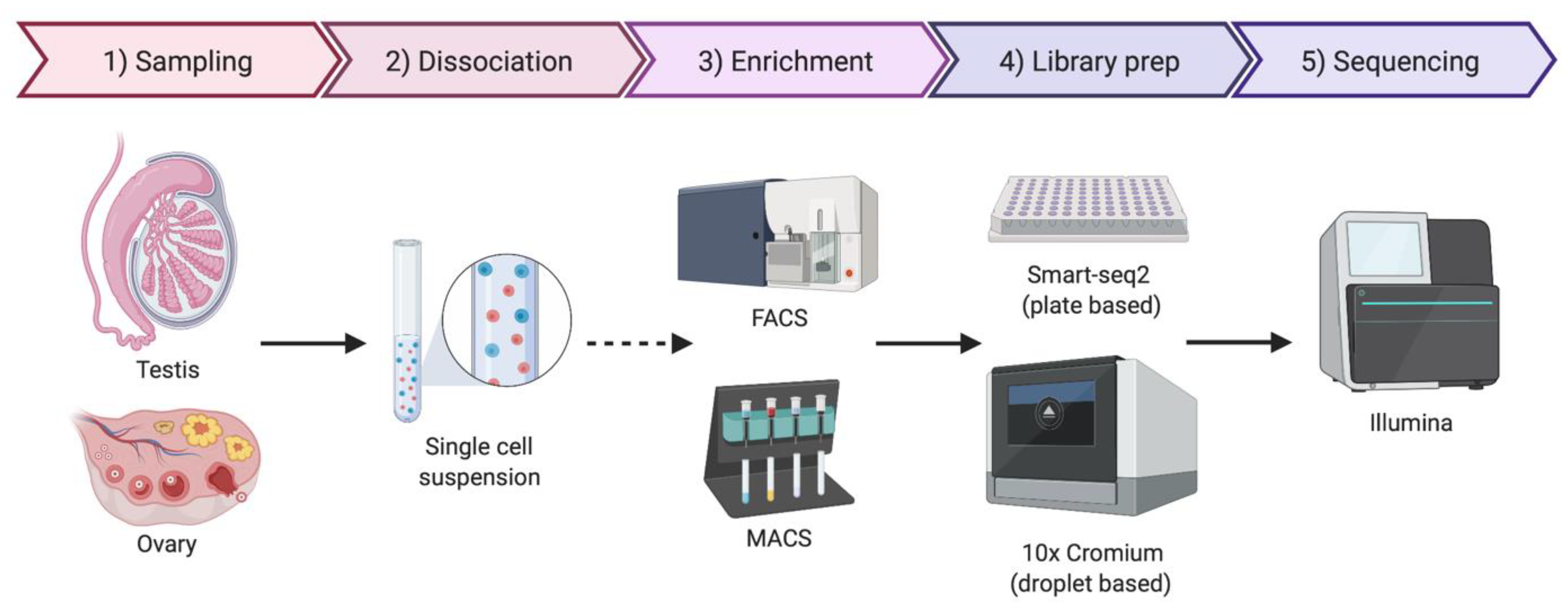
5. Single-Cell Sequencing Technologies
6. Single-Cell -Omics in Gonadal Development and Maturation
6.1. Early Gonadal Development and Differentiation at the Single-Cell Level
6.2. Testicular Maturation and Single-Cell Transcriptomics
6.3. Ovarian Maturation and Single-Cell Transcriptomics
7. Single-Cell -Omics in Gonadal Disease
7.1. DSDs
7.2. Detection of Infertility by Single-Cell Sequencing
7.3. Single-Cell Sequencing in Cancer Biology
8. Considerations
9. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 17-OHP | 17-hydroxy-progesterone |
| AMH | Anti-Müllerian Hormone |
| AR | Androgen Receptor |
| CAH | Congenital Adrenal Hyperplasia |
| CGH | Comparative Genomic Hybridization |
| CNV | Copy Number Variants |
| DSD | Disorders of Sex Development |
| E | Embryonic day |
| FACS | Fluorescence-Activated Cell Sorting |
| NGS | Next-Generation Sequencing |
| iPS cells | induced Pluripotent Stem cells |
| LH | Luteinzing Hormone |
| MACS | Magnetic Activated Cell Sorting |
| PBMCs | Peripheral Blood Mononuclear cells |
| P | Postnatal Day |
| PCOS | Polycystic Ovary Syndrome |
| scRNA-seq | Single-Cell RNA Sequencing |
| SNP | Single Nucleotide Polymorphisms |
| t-SNE | t-distributed Stochastic Neighbor Embedding |
| UMAP | Uniform Manifold Approximation and Projection |
| yo | Years Old |
References
- Gubbay, J.; Collignon, J.; Koopman, P.; Capel, B.; Economou, A.; Munsterberg, A.; Vivian, N.; Goodfellow, P.; Lovell-Badge, R. A gene mapping to the sex-determining region of the mouse Y chromosome is a member of a novel family of embryonically expressed genes. Nature 1990, 346, 245–250. [Google Scholar] [CrossRef]
- Koopman, P.; Gubbay, J.; Vivian, N.; Goodfellow, P.; Lovell-Badge, R. Male development of chromosomally female mice transgenic for Sry. Nature 1991, 351, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, A.H.; Berta, P.; Palmer, M.S.; Hawkins, J.R.; Griffiths, B.L.; Smith, M.J.; Foster, J.W.; Frischauf, A.M.; Lovell-Badge, R.; Goodfellow, P.N. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 1990, 346, 240–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilhelm, D.; Palmer, S.; Koopman, P. Sex determination and gonadal development in mammals. Physiol. Rev. 2007, 87, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekido, R.; Lovell-Badge, R. Sex determination involves synergistic action of SRY and SF1 on a specific Sox9 enhancer. Nature 2008, 453, 930–934. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, M.; Lau, Y.F. The sex-determining factors SRY and SOX9 regulate similar target genes and promote testis cord formation during testicular differentiation. Cell Rep. 2014, 8, 723–733. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.D.; George, F.W.; Renfree, M.B. The endocrine role in mammalian sexual differentiation. Recent Prog. Horm. Res. 1995, 50, 349–364. [Google Scholar] [CrossRef]
- Hughes, I.A.; Houk, C.; Ahmed, S.F.; Lee, P.A.; Lawson Wilkins Pediatric Endocrine Society/European Society for Paediatric Endocrinology Consensus Group. Consensus statement on management of intersex disorders. J. Pediatr. Urol. 2006, 2, 148–162. [Google Scholar] [CrossRef]
- Baetens, D.; Verdin, H.; De Baere, E.; Cools, M. Update on the genetics of differences of sex development (DSD). Best Pract. Res. Clin. Endocrinol. Metab. 2019, 33, 101271. [Google Scholar] [CrossRef]
- Barseghyan, H.; Delot, E.; Vilain, E. New genomic technologies: An aid for diagnosis of disorders of sex development. Horm. Metab. Res. 2015, 47, 312–320. [Google Scholar] [CrossRef]
- Parivesh, A.; Barseghyan, H.; Delot, E.; Vilain, E. Translating genomics to the clinical diagnosis of disorders/differences of sex development. Curr. Top. Dev. Biol. 2019, 134, 317–375. [Google Scholar] [CrossRef]
- Croft, B.; Ayers, K.; Sinclair, A.; Ohnesorg, T. Review disorders of sex development: The evolving role of genomics in diagnosis and gene discovery. Birth Defects Res. C Embryo Today 2016, 108, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Kent, J.; Wheatley, S.C.; Andrews, J.E.; Sinclair, A.H.; Koopman, P. A male-specific role for SOX9 in vertebrate sex determination. Development 1996, 122, 2813–2822. [Google Scholar] [PubMed]
- Chassot, A.A.; Gregoire, E.P.; Magliano, M.; Lavery, R.; Chaboissier, M.C. Genetics of ovarian differentiation: Rspo1, a major player. Sex. Dev. 2008, 2, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.; Siggers, P.; Corrochano, S.; Warr, N.; Sagar, D.; Grimes, D.T.; Suzuki, M.; Burdine, R.D.; Cong, F.; Koo, B.K.; et al. ZNRF3 functions in mammalian sex determination by inhibiting canonical WNT signaling. Proc. Natl. Acad. Sci. USA 2018, 115, 5474–5479. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, D.; Yang, J.X.; Thomas, P. Mammalian sex determination and gonad development. Curr. Top. Dev. Biol. 2013, 106, 89–121. [Google Scholar] [CrossRef]
- Nef, S.; Stevant, I.; Greenfield, A. Characterizing the bipotential mammalian gonad. Curr. Top. Dev. Biol. 2019, 134, 167–194. [Google Scholar] [CrossRef]
- Stevant, I.; Nef, S. Genetic Control of Gonadal Sex Determination and Development. Trends Genet. 2019, 35, 346–358. [Google Scholar] [CrossRef]
- Karl, J.; Capel, B. Sertoli cells of the mouse testis originate from the coelomic epithelium. Dev. Biol. 1998, 203, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Svingen, T.; Koopman, P. Building the mammalian testis: Origins, differentiation, and assembly of the component cell populations. Genes Dev. 2013, 27, 2409–2426. [Google Scholar] [CrossRef] [Green Version]
- Stevant, I.; Neirijnck, Y.; Borel, C.; Escoffier, J.; Smith, L.B.; Antonarakis, S.E.; Dermitzakis, E.T.; Nef, S. Deciphering Cell Lineage Specification during Male Sex Determination with Single-Cell RNA Sequencing. Cell Rep. 2018, 22, 1589–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebourcet, D.; O’Shaughnessy, P.J.; Pitetti, J.L.; Monteiro, A.; O’Hara, L.; Milne, L.; Tsai, Y.T.; Cruickshanks, L.; Riethmacher, D.; Guillou, F.; et al. Sertoli cells control peritubular myoid cell fate and support adult Leydig cell development in the prepubertal testis. Development 2014, 141, 2139–2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, M.K.; Tung, P.S.; Fritz, I.B. Cooperativity between Sertoli cells and testicular peritubular cells in the production and deposition of extracellular matrix components. J. Cell Biol. 1985, 100, 1941–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tung, P.S.; Skinner, M.K.; Fritz, I.B. Cooperativity between Sertoli cells and peritubular myoid cells in the formation of the basal lamina in the seminiferous tubule. Ann. N. Y. Acad. Sci. 1984, 438, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Brennan, J.; Tilmann, C.; Capel, B. Pdgfr-alpha mediates testis cord organization and fetal Leydig cell development in the XY gonad. Genes Dev. 2003, 17, 800–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.M.; Garland, K.K.; Russell, L.D. Desert hedgehog (Dhh) gene is required in the mouse testis for formation of adult-type Leydig cells and normal development of peritubular cells and seminiferous tubules. Biol. Reprod. 2000, 63, 1825–1838. [Google Scholar] [CrossRef] [Green Version]
- Svechnikov, K.; Landreh, L.; Weisser, J.; Izzo, G.; Colon, E.; Svechnikova, I.; Soder, O. Origin, development and regulation of human Leydig cells. Horm Res. Paediatr 2010, 73, 93–101. [Google Scholar] [CrossRef]
- Wainwright, E.N.; Wilhelm, D. The game plan: Cellular and molecular mechanisms of mammalian testis development. Curr. Top. Dev. Biol. 2010, 90, 231–262. [Google Scholar] [CrossRef]
- Uhlenhaut, N.H.; Treier, M. Foxl2 function in ovarian development. Mol. Genet. Metab. 2006, 88, 225–234. [Google Scholar] [CrossRef]
- Tomizuka, K.; Horikoshi, K.; Kitada, R.; Sugawara, Y.; Iba, Y.; Kojima, A.; Yoshitome, A.; Yamawaki, K.; Amagai, M.; Inoue, A.; et al. R-spondin1 plays an essential role in ovarian development through positively regulating Wnt-4 signaling. Hum. Mol. Genet. 2008, 17, 1278–1291. [Google Scholar] [CrossRef] [Green Version]
- Wear, H.M.; Eriksson, A.; Yao, H.H.; Watanabe, K.H. Cell-based computational model of early ovarian development in mice. Biol. Reprod. 2017, 97, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicol, B.; Grimm, S.A.; Gruzdev, A.; Scott, G.J.; Ray, M.K.; Yao, H.H. Genome-wide identification of FOXL2 binding and characterization of FOXL2 feminizing action in the fetal gonads. Hum. Mol. Genet. 2018, 27, 4273–4287. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, K.H.; Eicher, E.M. Evidence that Sry is expressed in pre-Sertoli cells and Sertoli and granulosa cells have a common precursor. Dev. Biol. 2001, 240, 92–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, W.; Spradling, A.C. Two distinct pathways of pregranulosa cell differentiation support follicle formation in the mouse ovary. Proc. Natl. Acad. Sci. USA 2020. [Google Scholar] [CrossRef] [PubMed]
- Rotgers, E.; Jorgensen, A.; Yao, H.H. At the Crossroads of Fate-Somatic Cell Lineage Specification in the Fetal Gonad. Endocr. Rev. 2018, 39, 739–759. [Google Scholar] [CrossRef]
- Liu, C.; Peng, J.; Matzuk, M.M.; Yao, H.H. Lineage specification of ovarian theca cells requires multicellular interactions via oocyte and granulosa cells. Nat. Commun. 2015, 6, 6934. [Google Scholar] [CrossRef] [Green Version]
- Honda, A.; Hirose, M.; Hara, K.; Matoba, S.; Inoue, K.; Miki, H.; Hiura, H.; Kanatsu-Shinohara, M.; Kanai, Y.; Kono, T.; et al. Isolation, characterization, and in vitro and in vivo differentiation of putative thecal stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 12389–12394. [Google Scholar] [CrossRef] [Green Version]
- Vilain, E. The genetics of ovotesticular disorders of sex development. Adv. Exp. Med. Biol. 2011, 707, 105–106. [Google Scholar] [CrossRef]
- Baxter, R.M.; Vilain, E. Translational genetics for diagnosis of human disorders of sex development. Annu. Rev. Genom. Hum. Genet. 2013, 14, 371–392. [Google Scholar] [CrossRef] [Green Version]
- Eggers, S.; Sadedin, S.; Van den Bergen, J.A.; Robevska, G.; Ohnesorg, T.; Hewitt, J.; Lambeth, L.; Bouty, A.; Knarston, I.M.; Tan, T.Y.; et al. Disorders of sex development: Insights from targeted gene sequencing of a large international patient cohort. Genome. Biol. 2016, 17, 243. [Google Scholar] [CrossRef] [Green Version]
- Bennecke, E.; Kohler, B.; Rohle, R.; Thyen, U.; Gehrmann, K.; Lee, P.; Nordenstrom, A.; Cohen-Kettenis, P.; Bouvattier, C.; Wiesemann, C. Disorders or Differences of Sex Development? Views of Affected Individuals on DSD Terminology. J. Sex Res. 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Audi, L.; Ahmed, S.F.; Krone, N.; Cools, M.; McElreavey, K.; Holterhus, P.M.; Greenfield, A.; Bashamboo, A.; Hiort, O.; Wudy, S.A.; et al. Genetics in endocrinology: Approaches to molecular genetic diagnosis in the management of differences/disorders of sex development (DSD): Position paper of EU COST Action BM 1303 ‘DSDnet’. Eur. J. Endocrinol. 2018, 179, R197–R206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, T.K.; Chowdhury, M.Z.; Mili, F.; Hutson, J.M.; Banu, T. Gender identity shows a high correlation with Prader score in patients with disorders of sex development (DSD) presenting in mid childhood. Pediatr. Surg. Int. 2014, 30, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Berta, P.; Hawkins, J.R.; Sinclair, A.H.; Taylor, A.; Griffiths, B.L.; Goodfellow, P.N.; Fellous, M. Genetic evidence equating SRY and the testis-determining factor. Nature 1990, 348, 448–450. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.W.; Dominguez-Steglich, M.A.; Guioli, S.; Kwok, C.; Weller, P.A.; Stevanovic, M.; Weissenbach, J.; Mansour, S.; Young, I.D.; Goodfellow, P.N.; et al. Campomelic dysplasia and autosomal sex reversal caused by mutations in an SRY-related gene. Nature 1994, 372, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.; Wirth, J.; Meyer, J.; Zabel, B.; Held, M.; Zimmer, J.; Pasantes, J.; Bricarelli, F.D.; Keutel, J.; Hustert, E.; et al. Autosomal sex reversal and campomelic dysplasia are caused by mutations in and around the SRY-related gene SOX9. Cell 1994, 79, 1111–1120. [Google Scholar] [CrossRef]
- Knower, K.C.; Kelly, S.; Ludbrook, L.M.; Bagheri-Fam, S.; Sim, H.; Bernard, P.; Sekido, R.; Lovell-Badge, R.; Harley, V.R. Failure of SOX9 regulation in 46XY disorders of sex development with SRY, SOX9 and SF1 mutations. PLoS ONE 2011, 6, e17751. [Google Scholar] [CrossRef] [Green Version]
- Ohnesorg, T.; Vilain, E.; Sinclair, A.H. The genetics of disorders of sex development in humans. Sex Dev. 2014, 8, 262–272. [Google Scholar] [CrossRef]
- Larney, C.; Bailey, T.L.; Koopman, P. Switching on sex: Transcriptional regulation of the testis-determining gene Sry. Development 2014, 141, 2195–2205. [Google Scholar] [CrossRef] [Green Version]
- Lourenco, D.; Brauner, R.; Rybczynska, M.; Nihoul-Fekete, C.; McElreavey, K.; Bashamboo, A. Loss-of-function mutation in GATA4 causes anomalies of human testicular development. Proc. Natl. Acad. Sci. USA 2011, 108, 1597–1602. [Google Scholar] [CrossRef] [Green Version]
- Allali, S.; Muller, J.B.; Brauner, R.; Lourenco, D.; Boudjenah, R.; Karageorgou, V.; Trivin, C.; Lottmann, H.; Lortat-Jacob, S.; Nihoul-Fekete, C.; et al. Mutation analysis of NR5A1 encoding steroidogenic factor 1 in 77 patients with 46, XY disorders of sex development (DSD) including hypospadias. PLoS ONE 2011, 6, e24117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achermann, J.C.; Ito, M.; Ito, M.; Hindmarsh, P.C.; Jameson, J.L. A mutation in the gene encoding steroidogenic factor-1 causes XY sex reversal and adrenal failure in humans. Nat. Genet. 1999, 22, 125–126. [Google Scholar] [CrossRef] [PubMed]
- Pearlman, A.; Loke, J.; Le Caignec, C.; White, S.; Chin, L.; Friedman, A.; Warr, N.; Willan, J.; Brauer, D.; Farmer, C.; et al. Mutations in MAP3K1 cause 46,XY disorders of sex development and implicate a common signal transduction pathway in human testis determination. Am. J. Hum. Genet. 2010, 87, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Kremen, J.; Chan, Y.M.; Swartz, J.M. Recent findings on the genetics of disorders of sex development. Curr. Opin. Urol. 2017, 27, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Looijenga, L.H.; Hersmus, R.; Oosterhuis, J.W.; Cools, M.; Drop, S.L.; Wolffenbuttel, K.P. Tumor risk in disorders of sex development (DSD). Best Pract. Res. Clin. Endocrinol. Metab. 2007, 21, 480–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, M.; Harley, V.R. Disorders of sex development: New genes, new concepts. Nat. Rev. Endocrinol. 2013, 9, 79–91. [Google Scholar] [CrossRef]
- Eggers, S.; Sinclair, A. Mammalian sex determination-insights from humans and mice. Chromosome Res. 2012, 20, 215–238. [Google Scholar] [CrossRef] [Green Version]
- Parma, P.; Radi, O.; Vidal, V.; Chaboissier, M.C.; Dellambra, E.; Valentini, S.; Guerra, L.; Schedl, A.; Camerino, G. R-spondin1 is essential in sex determination, skin differentiation and malignancy. Nat. Genet. 2006, 38, 1304–1309. [Google Scholar] [CrossRef]
- McElreavey, K.; Vilain, E.; Abbas, N.; Herskowitz, I.; Fellous, M. A regulatory cascade hypothesis for mammalian sex determination: SRY represses a negative regulator of male development. Proc. Natl. Acad. Sci. USA 1993, 90, 3368–3372. [Google Scholar] [CrossRef] [Green Version]
- Batista, R.L.; Costa, E.M.F.; Rodrigues, A.S.; Gomes, N.L.; Faria, J.A., Jr.; Nishi, M.Y.; Arnhold, I.J.P.; Domenice, S.; Mendonca, B.B. Androgen insensitivity syndrome: A review. Arch. Endocrinol. Metab. 2018, 62, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Tadokoro-Cuccaro, R.; Hughes, I.A. Androgen insensitivity syndrome. Curr. Opin. Endocrinol. Diabetes Obes. 2014, 21, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, B.; Beitel, L.K.; Nadarajah, A.; Paliouras, M.; Trifiro, M. The androgen receptor gene mutations database: 2012 update. Hum. Mutat. 2012, 33, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Okeigwe, I.; Kuohung, W. 5-Alpha reductase deficiency: A 40-year retrospective review. Curr. Opin. Endocrinol. Diabetes Obes. 2014, 21, 483–487. [Google Scholar] [CrossRef]
- Rafigh, M.; Salmaninejad, A.; Sorouri Khorashad, B.; Arabi, A.; Milanizadeh, S.; Hiradfar, M.; Abbaszadegan, M.R. Novel Deleterious Mutation in Steroid-5alpha-Reductase-2 in 46, XY Disorders of Sex Development: Case Report Study. Fetal Pediatr. Pathol. 2020, 1–8. [Google Scholar] [CrossRef] [PubMed]
- El-Maouche, D.; Arlt, W.; Merke, D.P. Congenital adrenal hyperplasia. Lancet 2017, 390, 2194–2210. [Google Scholar] [CrossRef]
- Speiser, P.W.; Arlt, W.; Auchus, R.J.; Baskin, L.S.; Conway, G.S.; Merke, D.P.; Meyer-Bahlburg, H.F.L.; Miller, W.L.; Murad, M.H.; Oberfield, S.E.; et al. Congenital Adrenal Hyperplasia Due to Steroid 21-Hydroxylase Deficiency: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2018, 103, 4043–4088. [Google Scholar] [CrossRef]
- Delot, E.C.; Papp, J.C.; Workgroup, D.-T.G.; Sandberg, D.E.; Vilain, E. Genetics of Disorders of Sex Development: The DSD-TRN Experience. Endocrinol. Metab. Clin. N. Am. 2017, 46, 519–537. [Google Scholar] [CrossRef]
- Kilberg, M.J.; McLoughlin, M.; Pyle, L.C.; Vogiatzi, M.G. Endocrine Management of Ovotesticular DSD, an Index Case and Review of the Literature. Pediatr. Endocrinol. Rev. 2019, 17, 110–116. [Google Scholar] [CrossRef]
- Witchel, S.F. Disorders of sex development. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 48, 90–102. [Google Scholar] [CrossRef]
- Garcia-Acero, M.; Moreno, O.; Suarez, F.; Rojas, A. Disorders of Sexual Development: Current Status and Progress in the Diagnostic Approach. Curr. Urol. 2020, 13, 169–178. [Google Scholar] [CrossRef]
- Mouriquand, P.D.; Gorduza, D.B.; Gay, C.L.; Meyer-Bahlburg, H.F.; Baker, L.; Baskin, L.S.; Bouvattier, C.; Braga, L.H.; Caldamone, A.C.; Duranteau, L.; et al. Surgery in disorders of sex development (DSD) with a gender issue: If (why), when, and how? J. Pediatr. Urol. 2016, 12, 139–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearsey, I.; Hutson, J.M. Disorders of sex development (DSD): Not only babies with ambiguous genitalia. A practical guide for surgeons. Pediatr. Surg. Int. 2017, 33, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Gothwal, M.; Yadav, G.; Singh, K. Complete Androgen Insensitivity Syndrome: Dilemmas for Further Management after Gonadectomy. J. Hum. Reprod. Sci. 2019, 12, 348–350. [Google Scholar] [CrossRef]
- Hemesath, T.P.; De Paula, L.C.P.; Carvalho, C.G.; Leite, J.C.L.; Guaragna-Filho, G.; Costa, E.C. Controversies on Timing of Sex Assignment and Surgery in Individuals with Disorders of Sex Development: A Perspective. Front. Pediatr. 2018, 6, 419. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, A.B.; Tishelman, A.C. Psychological perspectives to early surgery in the management of disorders/differences of sex development. Curr. Opin. Pediatr. 2019, 31, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, A.B.; Batista, R.L.; Costa, E.M.F.; Finlayson, C.; Sircili, M.H.P.; Denes, F.T.; Domenice, S.; Mendonca, B.B. Management of 46,XY Differences/Disorders of Sex Development (DSD) Throughout Life. Endocr. Rev. 2019, 40, 1547–1572. [Google Scholar] [CrossRef]
- Shnorhavorian, M. Navigating Differences/Disorders of Sex Development in 2020. J. Urol. 2020. [Google Scholar] [CrossRef] [PubMed]
- McNamara, E.R.; Swartz, J.M.; Diamond, D.A. Initial Management of Disorders of Sex Development in Newborns. Urology 2017, 101, 1–8. [Google Scholar] [CrossRef]
- Singh, R.J. Quantitation of 17-OH-progesterone (OHPG) for diagnosis of congenital adrenal hyperplasia (CAH). Methods Mol. Biol. 2010, 603, 271–277. [Google Scholar] [CrossRef]
- Day, D.J.; Speiser, P.W.; Schulze, E.; Bettendorf, M.; Fitness, J.; Barany, F.; White, P.C. Identification of non-amplifying CYP21 genes when using PCR-based diagnosis of 21-hydroxylase deficiency in congenital adrenal hyperplasia (CAH) affected pedigrees. Hum. Mol. Genet. 1996, 5, 2039–2048. [Google Scholar] [CrossRef] [Green Version]
- Audi, L.; Camats, N.; Fernandez-Cancio, M.; Granada, M.L. Development of Laboratory Investigations in Disorders of Sex Development. Sex. Dev. 2018, 12, 7–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanciotti, L.; Cofini, M.; Leonardi, A.; Bertozzi, M.; Penta, L.; Esposito, S. Different Clinical Presentations and Management in Complete Androgen Insensitivity Syndrome (CAIS). Int. J. Environ. Res. Public Health 2019, 16, 1268. [Google Scholar] [CrossRef] [Green Version]
- Hannema, S.E.; De Rijke, Y.B. Improving Laboratory Assessment in Disorders of Sex Development through a Multidisciplinary Network. Sex Dev. 2018, 12, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Ledig, S.; Hiort, O.; Scherer, G.; Hoffmann, M.; Wolff, G.; Morlot, S.; Kuechler, A.; Wieacker, P. Array-CGH analysis in patients with syndromic and non-syndromic XY gonadal dysgenesis: Evaluation of array CGH as diagnostic tool and search for new candidate loci. Hum. Reprod. 2010, 25, 2637–2646. [Google Scholar] [CrossRef] [Green Version]
- White, S.; Ohnesorg, T.; Notini, A.; Roeszler, K.; Hewitt, J.; Daggag, H.; Smith, C.; Turbitt, E.; Gustin, S.; Van den Bergen, J.; et al. Copy number variation in patients with disorders of sex development due to 46,XY gonadal dysgenesis. PLoS ONE 2011, 6, e17793. [Google Scholar] [CrossRef] [Green Version]
- Jordan, B.K.; Mohammed, M.; Ching, S.T.; Delot, E.; Chen, X.N.; Dewing, P.; Swain, A.; Rao, P.N.; Elejalde, B.R.; Vilain, E. Up-regulation of WNT-4 signaling and dosage-sensitive sex reversal in humans. Am. J. Hum. Genet. 2001, 68, 1102–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallet, D.; Bretones, P.; Michel-Calemard, L.; Dijoud, F.; David, M.; Morel, Y. Gonadal dysgenesis without adrenal insufficiency in a 46, XY patient heterozygous for the nonsense C16X mutation: A case of SF1 haploinsufficiency. J. Clin. Endocrinol. Metab. 2004, 89, 4829–4832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.J.; Willatt, L.; Homfray, T.; Woods, C.G. A SOX9 duplication and familial 46,XX developmental testicular disorder. N. Eng. J. Med. 2011, 364, 91–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barseghyan, H.; Delot, E.C.; Vilain, E. New technologies to uncover the molecular basis of disorders of sex development. Mol. Cell Endocrinol. 2018, 468, 60–69. [Google Scholar] [CrossRef]
- Bashamboo, A.; McElreavey, K. The role of next generation sequencing in understanding male and female sexual development: Clinical implications. Expert Rev. Endocrinol. Metab. 2016, 11, 433–443. [Google Scholar] [CrossRef]
- Gkountela, S.; Clark, A.T. RNA-Seq Library Generation from Rare Human Cells Isolated by FACS. Bio Protoc. 2013, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beverdam, A.; Koopman, P. Expression profiling of purified mouse gonadal somatic cells during the critical time window of sex determination reveals novel candidate genes for human sexual dysgenesis syndromes. Hum. Mol. Genet. 2006, 15, 417–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Ning, B.; Shi, T. Single-Cell RNA-Seq Technologies and Related Computational Data Analysis. Front. Genet. 2019, 10, 317. [Google Scholar] [CrossRef]
- La Manno, G. From single-cell RNA-seq to transcriptional regulation. Nat. Biotechnol. 2019, 37, 1421–1422. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.S.; Li, Y.; Mitra, A.K.; Bi, L.; Abyzov, A.; Van Wijnen, A.J.; Baughn, L.B.; Van Ness, B.; Rajkumar, V.; Kumar, S.; et al. Molecular signatures of multiple myeloma progression through single cell RNA-Seq. Blood Cancer J. 2019, 9, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, K.; Wang, X.; Meng, C.; He, L.; Sang, X.; Zheng, Y.; Xu, H. The application of single-cell sequencing technology in the diagnosis and treatment of hepatocellular carcinoma. Ann. Transl. Med. 2019, 7, 790. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.R.; Yong, K.W.; Choi, J.Y.; Cowie, A.C. Single-Cell RNA Sequencing and Its Combination with Protein and DNA Analyses. Cells 2020, 9, 1130. [Google Scholar] [CrossRef] [PubMed]
- Paolillo, C.; Londin, E.; Fortina, P. Single-Cell Genomics. Clin. Chem. 2019, 65, 972–985. [Google Scholar] [CrossRef]
- Wang, X.; He, Y.; Zhang, Q.; Ren, X.; Zhang, Z. Direct Comparative Analysis of 10X Genomics Chromium and Smart-seq2. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Ziegenhain, C.; Vieth, B.; Parekh, S.; Reinius, B.; Guillaumet-Adkins, A.; Smets, M.; Leonhardt, H.; Heyn, H.; Hellmann, I.; Enard, W. Comparative Analysis of Single-Cell RNA Sequencing Methods. Mol. Cell 2017, 65, 631–643.e4. [Google Scholar] [CrossRef] [Green Version]
- Becht, E.; McInnes, L.; Healy, J.; Dutertre, C.A.; Kwok, I.W.H.; Ng, L.G.; Ginhoux, F.; Newell, E.W. Dimensionality reduction for visualizing single-cell data using UMAP. Nat. Biotechnol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Efremova, M.; Vento-Tormo, R.; Park, J.E.; Teichmann, S.A.; James, K.R. Immunology in the Era of Single-Cell Technologies. Annu. Rev. Immunol. 2020, 38, 727–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strzelecka, P.M.; Ranzoni, A.M.; Cvejic, A. Dissecting human disease with single-cell omics: Application in model systems and in the clinic. Dis. Model. Mech. 2018, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pott, S.; Lieb, J.D. Single-cell ATAC-seq: Strength in numbers. Genome Biol. 2015, 16, 172. [Google Scholar] [CrossRef] [Green Version]
- Jia, G.; Preussner, J.; Chen, X.; Guenther, S.; Yuan, X.; Yekelchyk, M.; Kuenne, C.; Looso, M.; Zhou, Y.; Teichmann, S.; et al. Single cell RNA-seq and ATAC-seq analysis of cardiac progenitor cell transition states and lineage settlement. Nat. Commun. 2018, 9, 4877. [Google Scholar] [CrossRef] [Green Version]
- Stahl, P.L.; Salmen, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; Giacomello, S.; Asp, M.; Westholm, J.O.; Huss, M.; et al. Visualization and analysis of gene expression in tissue sections by spatial transcriptomics. Science 2016, 353, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Finck, A.; Fan, R. Single-Cell Omics Analyses Enabled by Microchip Technologies. Annu. Rev. Biomed. Eng. 2019, 21, 365–393. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; An, Q.; Sheu, K.; Trejo, B.; Fan, S.; Guo, Y. Single Cell Multi-Omics Technology: Methodology and Application. Front. Cell Dev. Biol. 2018, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Stoeckius, M.; Hafemeister, C.; Stephenson, W.; Houck-Loomis, B.; Chattopadhyay, P.K.; Swerdlow, H.; Satija, R.; Smibert, P. Simultaneous epitope and transcriptome measurement in single cells. Nat. Methods 2017, 14, 865–868. [Google Scholar] [CrossRef] [Green Version]
- Stoeckius, M.; Zheng, S.; Houck-Loomis, B.; Hao, S.; Yeung, B.Z.; Mauck, W.M., 3rd; Smibert, P.; Satija, R. Cell Hashing with barcoded antibodies enables multiplexing and doublet detection for single cell genomics. Genome Biol. 2018, 19, 224. [Google Scholar] [CrossRef] [Green Version]
- Regev, A.; Teichmann, S.A.; Lander, E.S.; Amit, I.; Benoist, C.; Birney, E.; Bodenmiller, B.; Campbell, P.; Carninci, P.; Clatworthy, M.; et al. The Human Cell Atlas. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Botting, R.A.; Dominguez Conde, C.; Popescu, D.M.; Lavaert, M.; Kunz, D.J.; Goh, I.; Stephenson, E.; Ragazzini, R.; Tuck, E.; et al. A cell atlas of human thymic development defines T cell repertoire formation. Science 2020, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aizarani, N.; Saviano, A.; Mailly, L.; Durand, S.; Herman, J.S.; Pessaux, P.; Baumert, T.F.; Grun, D. A human liver cell atlas reveals heterogeneity and epithelial progenitors. Nature 2019, 572, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Grow, E.J.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Nie, X.; Guo, Y.; Takei, Y.; Yun, J.; Cai, L.; et al. The adult human testis transcriptional cell atlas. Cell Res. 2018, 28, 1141–1157. [Google Scholar] [CrossRef]
- Guo, J.; Nie, X.; Giebler, M.; Mlcochova, H.; Wang, Y.; Grow, E.J.; DonorConnect; Kim, R.; Tharmalingam, M.; Matilionyte, G.; et al. The Dynamic Transcriptional Cell Atlas of Testis Development during Human Puberty. Cell Stem Cell 2020, 26, 262–276.e4. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Yan, L.; Guo, H.; Li, L.; Hu, B.; Zhao, Y.; Yong, J.; Hu, Y.; Wang, X.; Wei, Y.; et al. The Transcriptome and DNA Methylome Landscapes of Human Primordial Germ Cells. Cell 2015, 161, 1437–1452. [Google Scholar] [CrossRef] [Green Version]
- Tang, F.; Barbacioru, C.; Wang, Y.; Nordman, E.; Lee, C.; Xu, N.; Wang, X.; Bodeau, J.; Tuch, B.B.; Siddiqui, A.; et al. mRNA-Seq whole-transcriptome analysis of a single cell. Nat. Methods 2009, 6, 377–382. [Google Scholar] [CrossRef]
- Lindeman, R.E.; Gearhart, M.D.; Minkina, A.; Krentz, A.D.; Bardwell, V.J.; Zarkower, D. Sexual cell-fate reprogramming in the ovary by DMRT1. Curr. Biol. 2015, 25, 764–771. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Dong, J.; Yan, L.; Yong, J.; Liu, X.; Hu, Y.; Fan, X.; Wu, X.; Guo, H.; Wang, X.; et al. Single-Cell RNA-Seq Analysis Maps Development of Human Germline Cells and Gonadal Niche Interactions. Cell Stem Cell 2017, 20, 891–892. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zheng, Y.; Gao, Y.; Lin, Z.; Yang, S.; Wang, T.; Wang, Q.; Xie, N.; Hua, R.; Liu, M.; et al. Single-cell RNA-seq uncovers dynamic processes and critical regulators in mouse spermatogenesis. Cell Res. 2018, 28, 879–896. [Google Scholar] [CrossRef] [Green Version]
- Green, C.D.; Ma, Q.; Manske, G.L.; Shami, A.N.; Zheng, X.; Marini, S.; Moritz, L.; Sultan, C.; Gurczynski, S.J.; Moore, B.B.; et al. A Comprehensive Roadmap of Murine Spermatogenesis Defined by Single-Cell RNA-Seq. Dev. Cell 2018, 46, 651–667.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermann, B.P.; Cheng, K.; Singh, A.; Roa-De La Cruz, L.; Mutoji, K.N.; Chen, I.C.; Gildersleeve, H.; Lehle, J.D.; Mayo, M.; Westernstroer, B.; et al. The Mammalian Spermatogenesis Single-Cell Transcriptome, from Spermatogonial Stem Cells to Spermatids. Cell Rep. 2018, 25, 1650–1667.e8. [Google Scholar] [CrossRef] [Green Version]
- Vertesy, A.; Arindrarto, W.; Roost, M.S.; Reinius, B.; Torrens-Juaneda, V.; Bialecka, M.; Moustakas, I.; Ariyurek, Y.; Kuijk, E.; Mei, H.; et al. Parental haplotype-specific single-cell transcriptomics reveal incomplete epigenetic reprogramming in human female germ cells. Nat. Commun. 2018, 9, 1873. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, X.; Chang, G.; Chen, Y.; An, G.; Yan, L.; Gao, S.; Xu, Y.; Cui, Y.; Dong, J.; et al. Single-Cell RNA Sequencing Analysis Reveals Sequential Cell Fate Transition during Human Spermatogenesis. Cell Stem Cell 2018, 23, 599–614.e4. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yan, Z.; Qin, Q.; Nisenblat, V.; Chang, H.M.; Yu, Y.; Wang, T.; Lu, C.; Yang, M.; Yang, S.; et al. Transcriptome Landscape of Human Folliculogenesis Reveals Oocyte and Granulosa Cell Interactions. Mol. Cell 2018, 72, 1021–1034.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, C.; Eling, N.; Martinez-Jimenez, C.P.; Marioni, J.C.; Odom, D.T. Staged developmental mapping and X chromosome transcriptional dynamics during mouse spermatogenesis. Nat. Commun. 2019, 10, 1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Bialecka, M.; Moustakas, I.; Lam, E.; Torrens-Juaneda, V.; Borggreven, N.V.; Trouw, L.; Louwe, L.A.; Pilgram, G.S.K.; Mei, H.; et al. Single-cell reconstruction of follicular remodeling in the human adult ovary. Nat. Commun. 2019, 10, 3164. [Google Scholar] [CrossRef]
- Grive, K.J.; Hu, Y.; Shu, E.; Grimson, A.; Elemento, O.; Grenier, J.K.; Cohen, P.E. Dynamic transcriptome profiles within spermatogonial and spermatocyte populations during postnatal testis maturation revealed by single-cell sequencing. PLoS Genet. 2019, 15, e1007810. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Liu, S.; Wu, Q.; Zhang, L.; Guo, F. Integrative single-cell analysis of transcriptome, DNA methylome and chromatin accessibility in mouse oocytes. Cell Res. 2019, 29, 110–123. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Li, L.; Li, J.; Wu, X.; Hu, B.; Zhu, P.; Wen, L.; Tang, F. Single-cell multi-omics sequencing of mouse early embryos and embryonic stem cells. Cell Res. 2017, 27, 967–988. [Google Scholar] [CrossRef]
- Law, N.C.; Oatley, M.J.; Oatley, J.M. Developmental kinetics and transcriptome dynamics of stem cell specification in the spermatogenic lineage. Nat. Commun. 2019, 10, 2787. [Google Scholar] [CrossRef] [Green Version]
- Sohni, A.; Tan, K.; Song, H.W.; Burow, D.; De Rooij, D.G.; Laurent, L.; Hsieh, T.C.; Rabah, R.; Hammoud, S.S.; Vicini, E.; et al. The Neonatal and Adult Human Testis Defined at the Single-Cell Level. Cell Rep. 2019, 26, 1501–1517.e4. [Google Scholar] [CrossRef] [Green Version]
- Stevant, I.; Kuhne, F.; Greenfield, A.; Chaboissier, M.C.; Dermitzakis, E.T.; Nef, S. Dissecting Cell Lineage Specification and Sex Fate Determination in Gonadal Somatic Cells Using Single-Cell Transcriptomics. Cell Rep. 2019, 26, 3272–3283.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estermann, M.A.; Williams, S.; Hirst, C.E.; Roly, Z.Y.; Serralbo, O.; Adhikari, D.; Powell, D.; Major, A.T.; Smith, C.A. Insights into Gonadal Sex Differentiation Provided by Single-Cell Transcriptomics in the Chicken Embryo. Cell Rep. 2020, 31, 107491. [Google Scholar] [CrossRef] [PubMed]
- Lau, X.; Munusamy, P.; Ng, M.J.; Sangrithi, M. Single-Cell RNA Sequencing of the Cynomolgus Macaque Testis Reveals Conserved Transcriptional Profiles during Mammalian Spermatogenesis. Dev. Cell 2020. [Google Scholar] [CrossRef] [PubMed]
- Shami, A.N.; Zheng, X.; Munyoki, S.K.; Ma, Q.; Manske, G.L.; Green, C.D.; Sukhwani, M.; Orwig, K.E.; Li, J.Z.; Hammoud, S.S. Single-Cell RNA Sequencing of Human, Macaque, and Mouse Testes Uncovers Conserved and Divergent Features of Mammalian Spermatogenesis. Dev. Cell 2020. [Google Scholar] [CrossRef]
- Tan, K.; Song, H.W.; Wilkinson, M.F. Single-cell RNAseq analysis of testicular germ and somatic cell development during the perinatal period. Development 2020, 147. [Google Scholar] [CrossRef]
- Wagner, M.; Yoshihara, M.; Douagi, I.; Damdimopoulos, A.; Panula, S.; Petropoulos, S.; Lu, H.; Pettersson, K.; Palm, K.; Katayama, S.; et al. Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells. Nat. Commun. 2020, 11, 1147. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zheng, Y.; Li, J.; Yu, Y.; Zhang, W.; Song, M.; Liu, Z.; Min, Z.; Hu, H.; Jing, Y.; et al. Single-Cell Transcriptomic Atlas of Primate Ovarian Aging. Cell 2020, 180, 585–600.e19. [Google Scholar] [CrossRef]
- Xia, B.; Yan, Y.; Baron, M.; Wagner, F.; Barkley, D.; Chiodin, M.; Kim, S.Y.; Keefe, D.L.; Alukal, J.P.; Boeke, J.D.; et al. Widespread Transcriptional Scanning in the Testis Modulates Gene Evolution Rates. Cell 2020, 180, 248–262.e21. [Google Scholar] [CrossRef]
- She, Z.Y.; Yang, W.X. Sry and SoxE genes: How they participate in mammalian sex determination and gonadal development? Semin. Cell Dev. Biol. 2017, 63, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wang, T.; Leng, L.; Zheng, W.; Huang, J.; Fang, F.; Yang, L.; Chen, F.; Lin, G.; Wang, W.J.; et al. Single-cell RNA-seq reveals distinct dynamic behavior of sex chromosomes during early human embryogenesis. Mol. Reprod. Dev. 2019, 86, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Workman, S.; Wilson, M. The molecular pathways underlying early gonadal development. J. Mol. Endocrinol. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeays-Ward, K.; Hoyle, C.; Brennan, J.; Dandonneau, M.; Alldus, G.; Capel, B.; Swain, A. Endothelial and steroidogenic cell migration are regulated by WNT4 in the developing mammalian gonad. Development 2003, 130, 3663–3670. [Google Scholar] [CrossRef] [Green Version]
- Rey, R.; Josso, N.; Racine, C. Sexual Differentiation. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., Kopp, P., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Kashimada, K.; Koopman, P. Sry: The master switch in mammalian sex determination. Development 2010, 137, 3921–3930. [Google Scholar] [CrossRef] [Green Version]
- Lovell-Badge, R.; Goodfellow, P.N. SRY and sex determination in mammals. Ann. Rev. Genet. 1993, 27, 71–92. [Google Scholar]
- Koopman, P. Sry and Sox9: Mammalian testis-determining genes. Cell Mol. Life Sci. 1999, 55, 839–856. [Google Scholar] [CrossRef]
- Gonen, N.; Lovell-Badge, R. The regulation of Sox9 expression in the gonad. Curr. Top. Dev. Biol. 2019, 134, 223–252. [Google Scholar] [CrossRef]
- Barrionuevo, F.; Bagheri-Fam, S.; Klattig, J.; Kist, R.; Taketo, M.M.; Englert, C.; Scherer, G. Homozygous inactivation of Sox9 causes complete XY sex reversal in mice. Biol. Reprod. 2006, 74, 195–201. [Google Scholar] [CrossRef]
- Barrionuevo, F.J.; Hurtado, A.; Kim, G.J.; Real, F.M.; Bakkali, M.; Kopp, J.L.; Sander, M.; Scherer, G.; Burgos, M.; Jimenez, R. Sox9 and Sox8 protect the adult testis from male-to-female genetic reprogramming and complete degeneration. eLife 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Chassot, A.A.; Gillot, I.; Chaboissier, M.C. R-spondin1, WNT4, and the CTNNB1 signaling pathway: Strict control over ovarian differentiation. Reproduction 2014, 148, R97–R110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannetier, M.; Chassot, A.A.; Chaboissier, M.C.; Pailhoux, E. Involvement of FOXL2 and RSPO1 in Ovarian Determination, Development, and Maintenance in Mammals. Sex. Dev. 2016, 10, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Gustin, S.E.; Hogg, K.; Stringer, J.M.; Rastetter, R.H.; Pelosi, E.; Miles, D.C.; Sinclair, A.H.; Wilhelm, D.; Western, P.S. WNT/beta-catenin and p27/FOXL2 differentially regulate supporting cell proliferation in the developing ovary. Dev. Biol. 2016, 412, 250–260. [Google Scholar] [CrossRef]
- Zheng, W.; Zhang, H.; Gorre, N.; Risal, S.; Shen, Y.; Liu, K. Two classes of ovarian primordial follicles exhibit distinct developmental dynamics and physiological functions. Hum. Mol. Genet. 2014, 23, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Chue, J.; Smith, C.A. Sex determination and sexual differentiation in the avian model. FEBS J. 2011, 278, 1027–1034. [Google Scholar] [CrossRef]
- Lambeth, L.S.; Morris, K.; Ayers, K.L.; Wise, T.G.; O’Neil, T.; Wilson, S.; Cao, Y.; Sinclair, A.H.; Cutting, A.D.; Doran, T.J.; et al. Overexpression of Anti-Mullerian Hormone Disrupts Gonadal Sex Differentiation, Blocks Sex Hormone Synthesis, and Supports Cell Autonomous Sex Development in the Chicken. Endocrinology 2016, 157, 1258–1275. [Google Scholar] [CrossRef] [Green Version]
- Hirst, C.E.; Major, A.T.; Smith, C.A. Sex determination and gonadal sex differentiation in the chicken model. Int. J. Dev. Biol. 2018, 62, 153–166. [Google Scholar] [CrossRef]
- Smith, C.A.; Roeszler, K.N.; Bowles, J.; Koopman, P.; Sinclair, A.H. Onset of meiosis in the chicken embryo; evidence of a role for retinoic acid. BMC Dev. Biol. 2008, 8, 85. [Google Scholar] [CrossRef] [Green Version]
- Matson, C.K.; Murphy, M.W.; Sarver, A.L.; Griswold, M.D.; Bardwell, V.J.; Zarkower, D. DMRT1 prevents female reprogramming in the postnatal mammalian testis. Nature 2011, 476, 101–104. [Google Scholar] [CrossRef]
- Uhlenhaut, N.H.; Jakob, S.; Anlag, K.; Eisenberger, T.; Sekido, R.; Kress, J.; Treier, A.C.; Klugmann, C.; Klasen, C.; Holter, N.I.; et al. Somatic sex reprogramming of adult ovaries to testes by FOXL2 ablation. Cell 2009, 139, 1130–1142. [Google Scholar] [CrossRef] [Green Version]
- Geyer, C.B. Setting the Stage: The First Round of Spermatogenesis. In The Biology of Mammalian Spermatogonia; Oatley, J.M., Griswold, M.D., Eds.; Springer: New York, NY, USA, 2017; pp. 39–63. [Google Scholar] [CrossRef]
- Hu, S.L. Non-human primate models for AIDS vaccine research. Curr. Drug Targets Infect. Disord. 2005, 5, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edson, M.A.; Nagaraja, A.K.; Matzuk, M.M. The mammalian ovary from genesis to revelation. Endocr. Rev. 2009, 30, 624–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gershon, E.; Dekel, N. Newly Identified Regulators of Ovarian Folliculogenesis and Ovulation. Int. J. Mol. Sci. 2020, 21, 4565. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.C.; White, Y.A.; Tilly, J.L. Purification of oogonial stem cells from adult mouse and human ovaries: An assessment of the literature and a view toward the future. Reprod. Sci. 2013, 20, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Grieve, K.M.; McLaughlin, M.; Dunlop, C.E.; Telfer, E.E.; Anderson, R.A. The controversial existence and functional potential of oogonial stem cells. Maturitas 2015, 82, 278–281. [Google Scholar] [CrossRef] [Green Version]
- Zarate-Garcia, L.; Lane, S.I.; Merriman, J.A.; Jones, K.T. FACS-sorted putative oogonial stem cells from the ovary are neither DDX4-positive nor germ cells. Sci. Rep. 2016, 6, 27991. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.L.; Kikuchi, K.; Sun, Q.Y.; Schatten, H. Oocyte aging: Cellular and molecular changes, developmental potential and reversal possibility. Hum. Reprod. Update 2009, 15, 573–585. [Google Scholar] [CrossRef] [Green Version]
- Stringer, J.M.; Winship, A.; Zerafa, N.; Wakefield, M.; Hutt, K. Oocytes can efficiently repair DNA double-strand breaks to restore genetic integrity and protect offspring health. Proc. Natl. Acad. Sci. USA 2020, 117, 11513–11522. [Google Scholar] [CrossRef]
- Liu, C.; Rodriguez, K.; Yao, H.H. Mapping lineage progression of somatic progenitor cells in the mouse fetal testis. Development 2016, 143, 3700–3710. [Google Scholar] [CrossRef] [Green Version]
- Winterhoff, B.J.; Maile, M.; Mitra, A.K.; Sebe, A.; Bazzaro, M.; Geller, M.A.; Abrahante, J.E.; Klein, M.; Hellweg, R.; Mullany, S.A.; et al. Single cell sequencing reveals heterogeneity within ovarian cancer epithelium and cancer associated stromal cells. Gynecol. Oncol. 2017, 144, 598–606. [Google Scholar] [CrossRef] [Green Version]
- Vuong, N.H.; Cook, D.P.; Forrest, L.A.; Carter, L.E.; Robineau-Charette, P.; Kofsky, J.M.; Hodgkinson, K.M.; Vanderhyden, B.C. Single-cell RNA-sequencing reveals transcriptional dynamics of estrogen-induced dysplasia in the ovarian surface epithelium. PLoS Genet. 2018, 14, e1007788. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Huang, L.L.; Xu, J.; Ma, C.Q.; Chen, Z.H.; Zhang, Z.; Liao, C.H.; Zheng, S.X.; Huang, P.; Xu, W.M.; et al. Proteomics and single-cell RNA analysis of Akap4-knockout mice model confirm indispensable role of Akap4 in spermatogenesis. Dev. Biol. 2019, 454, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Wells, D.; Rusch, J.; Ahmad, S.; Marchini, J.; Myers, S.R.; Conrad, D.F. Unified single-cell analysis of testis gene regulation and pathology in five mouse strains. eLife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tang, D.; Zheng, F.; Xu, Y.; Guo, H.; Zhou, J.; Lin, L.; Xie, J.; Ou, M.; Dai, Y. Single-Cell Sequencing Reveals the Relationship between Phenotypes and Genotypes of Klinefelter Syndrome. Cytogenet. Genome Res. 2019, 159, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Izar, B.; Tirosh, I.; Stover, E.H.; Wakiro, I.; Cuoco, M.S.; Alter, I.; Rodman, C.; Leeson, R.; Su, M.J.; Shah, P.; et al. A single-cell landscape of high-grade serous ovarian cancer. Nat. Med. 2020, 26, 1271–1279. [Google Scholar] [CrossRef]
- Nelson, L.; Tighe, A.; Golder, A.; Littler, S.; Bakker, B.; Moralli, D.; Murtuza Baker, S.; Donaldson, I.J.; Spierings, D.C.J.; Wardenaar, R.; et al. A living biobank of ovarian cancer ex vivo models reveals profound mitotic heterogeneity. Nat. Commun. 2020, 11, 822. [Google Scholar] [CrossRef]
- Park, C.J.; Lin, P.C.; Zhou, S.; Barakat, R.; Bashir, S.T.; Choi, J.M.; Cacioppo, J.A.; Oakley, O.R.; Duffy, D.M.; Lydon, J.P.; et al. Progesterone Receptor Serves the Ovary as a Trigger of Ovulation and a Terminator of Inflammation. Cell Rep. 2020, 31, 107496. [Google Scholar] [CrossRef]
- Zitzmann, M.; Rohayem, J. Gonadal dysfunction and beyond: Clinical challenges in children, adolescents, and adults with 47,XXY Klinefelter syndrome. Am. J. Med. Genet. C Semin. Med. Genet. 2020, 184, 302–312. [Google Scholar] [CrossRef]
- Wu, H.; Uchimura, K.; Donnelly, E.L.; Kirita, Y.; Morris, S.A.; Humphreys, B.D. Comparative Analysis and Refinement of Human PSC-Derived Kidney Organoid Differentiation with Single-Cell Transcriptomics. Cell Stem Cell 2018, 23, 869–881.e8. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez Gutierrez, D.; Eid, W.; Biason-Lauber, A. A Human Gonadal Cell Model from Induced Pluripotent Stem Cells. Front. Genet. 2018, 9, 498. [Google Scholar] [CrossRef]
- Li, L.; Li, Y.; Sottas, C.; Culty, M.; Fan, J.; Hu, Y.; Cheung, G.; Chemes, H.E.; Papadopoulos, V. Directing differentiation of human induced pluripotent stem cells toward androgen-producing Leydig cells rather than adrenal cells. Proc. Natl. Acad. Sci. USA 2019, 116, 23274–23283. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Ding, S.; He, S.; He, L.; Gao, K.; Peng, S.; Shuai, C. Differentiation of primordial germ cells from premature ovarian insufficiency-derived induced pluripotent stem cells. Stem. Cell Res. Ther. 2019, 10, 156. [Google Scholar] [CrossRef]
- Sakib, S.; Goldsmith, T.; Voigt, A.; Dobrinski, I. Testicular organoids to study cell-cell interactions in the mammalian testis. Andrology 2020, 8, 835–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krausz, C.; Riera-Escamilla, A. Genetics of male infertility. Nat. Rev. Urol. 2018, 15, 369–384. [Google Scholar] [CrossRef] [PubMed]
- Houston, B.J.; Conrad, D.F.; O’Bryan, M.K. A framework for high-resolution phenotyping of candidate male infertility mutants: From human to mouse. Hum. Genet. 2020. [Google Scholar] [CrossRef]
- Chiba, K.; Enatsu, N.; Fujisawa, M. Management of non-obstructive azoospermia. Reprod. Med. Biol. 2016, 15, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Practice Committee of the American Society for Reproductive Medicine. Electronic address, a.a.o. Management of nonobstructive azoospermia: A committee opinion. Fertil. Steril. 2018, 110, 1239–1245. [Google Scholar] [CrossRef]
- Liu, Q.; Li, Y.; Feng, Y.; Liu, C.; Ma, J.; Li, Y.; Xiang, H.; Ji, Y.; Cao, Y.; Tong, X.; et al. Single-cell analysis of differences in transcriptomic profiles of oocytes and cumulus cells at GV, MI, MII stages from PCOS patients. Sci. Rep. 2016, 6, 39638. [Google Scholar] [CrossRef] [Green Version]
- Aiello, N.M.; Stanger, B.Z. Echoes of the embryo: Using the developmental biology toolkit to study cancer. Dis. Model. Mech. 2016, 9, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Lawson, D.A.; Kessenbrock, K.; Davis, R.T.; Pervolarakis, N.; Werb, Z. Tumour heterogeneity and metastasis at single-cell resolution. Nat. Cell Biol. 2018, 20, 1349–1360. [Google Scholar] [CrossRef]
- Landero-Huerta, D.A.; Vigueras-Villasenor, R.M.; Yokoyama-Rebollar, E.; Arechaga-Ocampo, E.; Rojas-Castaneda, J.C.; Jimenez-Trejo, F.; Chavez-Saldana, M. Epigenetic and risk factors of testicular germ cell tumors: A brief review. Front. Biosci. (Landmark Ed.) 2017, 22, 1073–1098. [Google Scholar] [CrossRef] [Green Version]
- Spoor, J.A.; Oosterhuis, J.W.; Hersmus, R.; Biermann, K.; Wolffenbuttel, K.P.; Cools, M.; Kazmi, Z.; Ahmed, S.F.; Looijenga, L.H.J. Histological Assessment of Gonads in DSD: Relevance for Clinical Management. Sex. Dev. 2018, 12, 106–122. [Google Scholar] [CrossRef] [Green Version]
- Hughes, I.A.; Nihoul-Fekete, C.; Thomas, B.; Cohen-Kettenis, P.T. Consequences of the ESPE/LWPES guidelines for diagnosis and treatment of disorders of sex development. Best Pract. Res. Clin. Endocrinol. Metab. 2007, 21, 351–365. [Google Scholar] [CrossRef]
- Gillam, L.H.; Hewitt, J.K.; Warne, G.L. Ethical principles for the management of infants with disorders of sex development. Horm. Res. Paediatr. 2010, 74, 412–418. [Google Scholar] [CrossRef]
- Diamond, D.A.; Swartz, J.; Tishelman, A.; Johnson, J.; Chan, Y.M. Management of pediatric patients with DSD and ambiguous genitalia: Balancing the child’s moral claims to self-determination with parental values and preferences. J. Pediatr. Urol. 2018, 14, 416.e1–416.e5. [Google Scholar] [CrossRef]
- Gomes, N.L.; Chetty, T.; Jorgensen, A.; Mitchell, R.T. Disorders of Sex Development-Novel Regulators, Impacts on Fertility, and Options for Fertility Preservation. Int. J. Mol. Sci. 2020, 21, 2282. [Google Scholar] [CrossRef] [Green Version]
- Guercio, G.; Costanzo, M.; Grinspon, R.P.; Rey, R.A. Fertility Issues in Disorders of Sex Development. Endocrinol. Metab. Clin. N. Am. 2015, 44, 867–881. [Google Scholar] [CrossRef]
- Denisenko, E.; Guo, B.B.; Jones, M.; Hou, R.; De Kock, L.; Lassmann, T.; Poppe, D.; Clement, O.; Simmons, R.K.; Lister, R.; et al. Systematic assessment of tissue dissociation and storage biases in single-cell and single-nucleus RNA-seq workflows. Genome. Biol. 2020, 21, 130. [Google Scholar] [CrossRef]
- Villacampa, E.G.; Larsson, L.; Kvastad, L.; Andersson, A.; Carlson, J.; Lundeberg, J. Genome-wide Spatial Expression Profiling in FFPE Tissues. bioRxiv 2020. [Google Scholar] [CrossRef]
- O’Flanagan, C.H.; Campbell, K.R.; Zhang, A.W.; Kabeer, F.; Lim, J.L.P.; Biele, J.; Eirew, P.; Lai, D.; McPherson, A.; Kong, E.; et al. Dissociation of solid tumor tissues with cold active protease for single-cell RNA-seq minimizes conserved collagenase-associated stress responses. Genome Biol. 2019, 20, 210. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
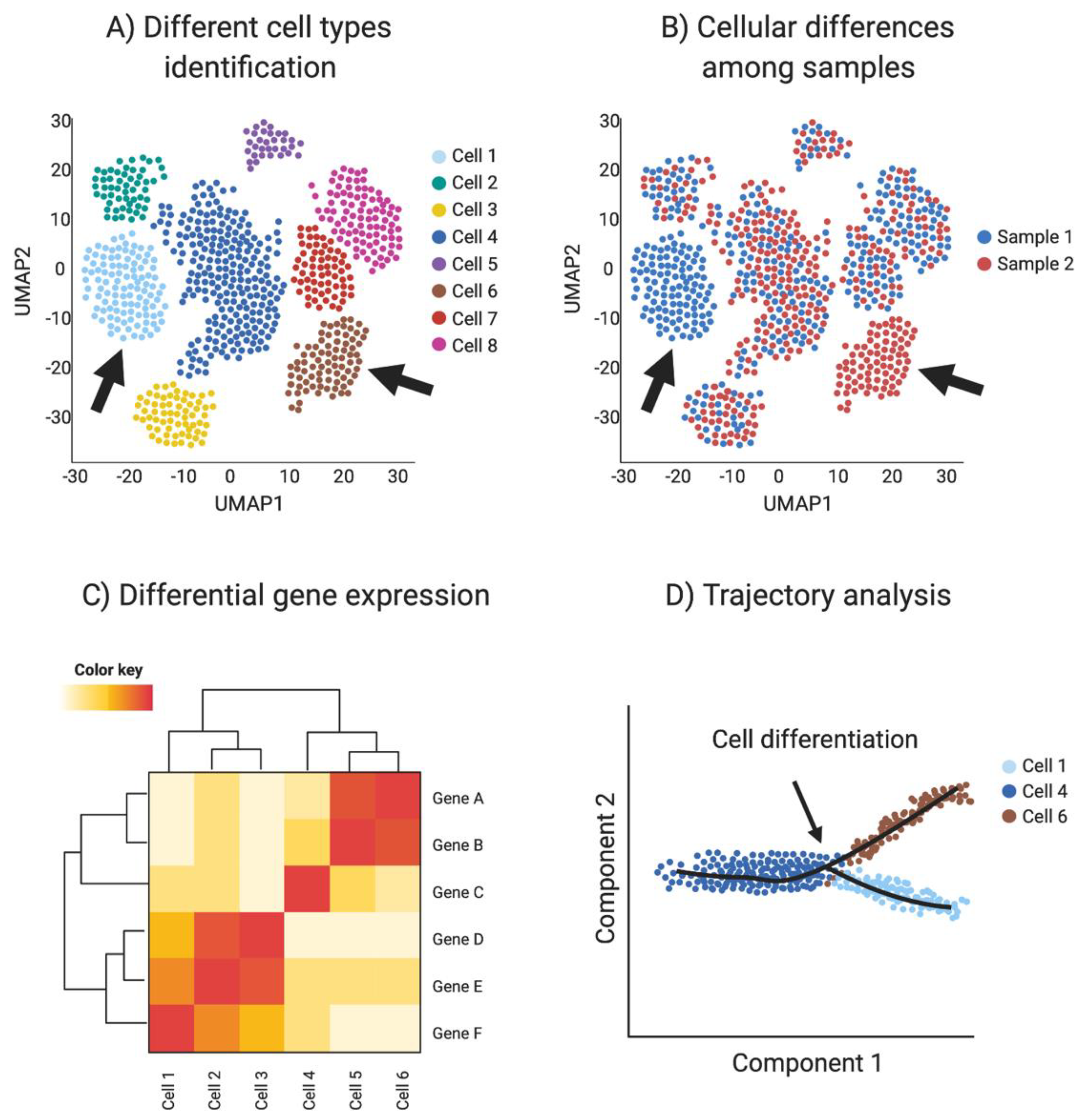{kind=link}
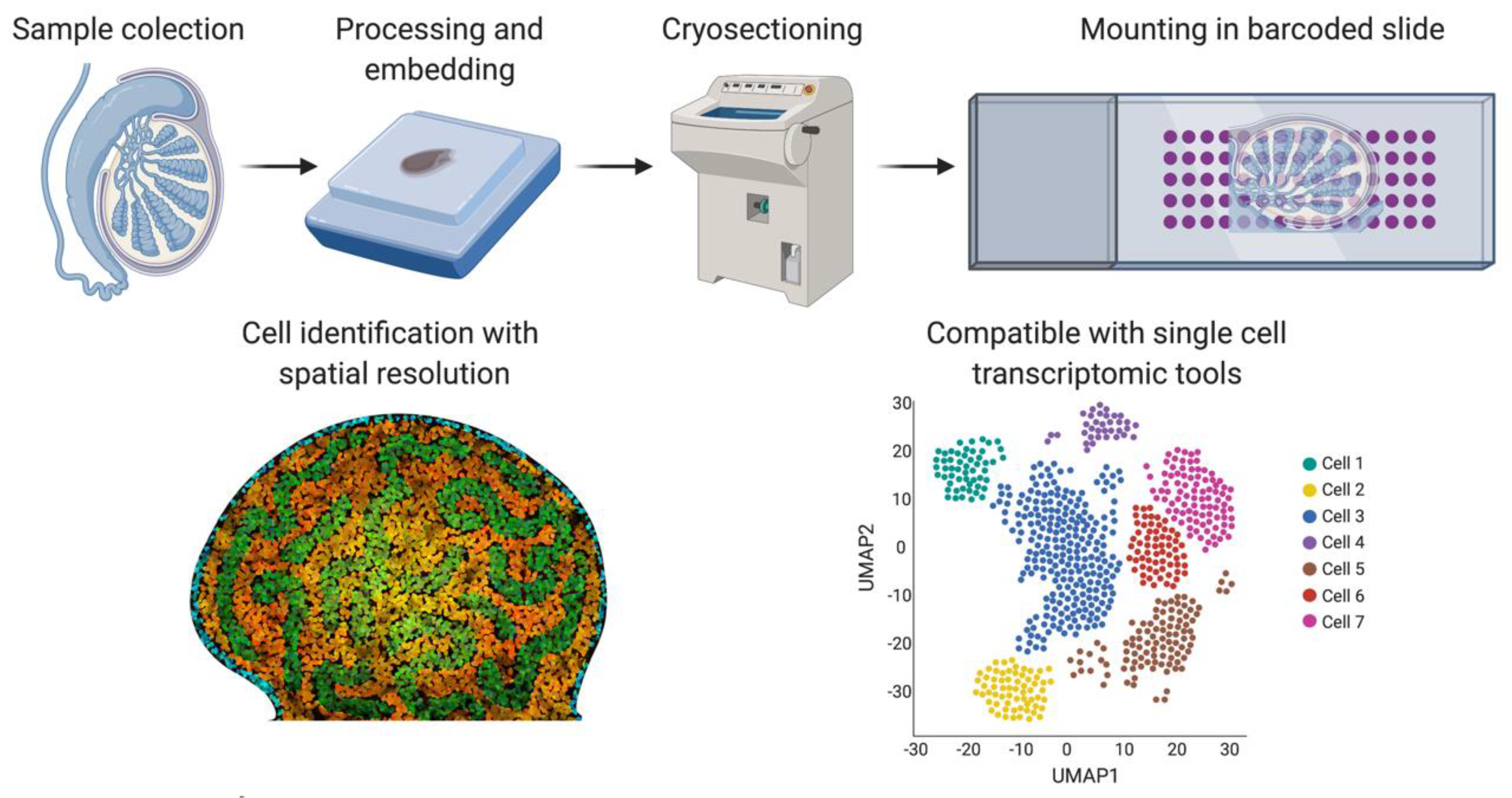{kind=link}
| Origin | 46,XY DSD | 46,XX DSD |
|---|---|---|
| DSD at the gonadal level | Gonadal dysgenesis (gonads are streaks of fibrous tissue) female Müllerian) ducts and variably feminized external genitalia Infertility | Gonads are testicular or partially testicular Male ducts Male or masculinized external genitalia Infertility |
| DSD at the hormonal level | Gonads typically testes Female ducts absent (AMH present) Partial or complete feminization of ducts and external genitalia due to androgen insensitivity (AIS) | Gonads typical ovaries Male ducts usually regressed External genitalia variably masculinized Elevated androgen levels, typically due to adrenal hyperplasia (CAH) |
| Methods of DSD diagnosis | -Prader scale for external genitalia -Ultrasound/laparoscopy of gonads and internal ducts -Karyotype analysis (46,XX, 46,XY, 46,X0, 46,XXY) -Biochemistry/endocrinology (e.g., steroid and peptide hormone levels) -Targeted gene mutation screening (e.g., SF1, SRY, CYP21A1, AR genes) -Whole genome or exome sequencing | |
| Year | Authors | Organism | Developmental Stage | Tissue | Technology | Cells | Data |
|---|---|---|---|---|---|---|---|
| 2015 | Guo et al. [116] | Human embryo (male and female) | Males: 4w, 7w, 10w, 11w and 19w. Females: 4w, 8w, 10w, 11w and 17w | PGCs and somatic cells | MACS, FACS, Tang et al. [117] | 319 | GSE63818 |
| 2015 | Lindeman et al. [118] | CAG-Stop-Dmrt1-Gfp; Sf1-Cre mice (female) | P17 | GFP sorted supporting cells | FACS, SMARTer (C1) | 68 | GSE64960 |
| 2017 | Li et al. [119] | Human embryo (male and female) | Males: 4w, 9w, 10w, 12w, 19w, 20w, 21w and 25w. Females: 5w, 7w, 8w, 10w, 11w, 12w, 14w, 18w, 20w, 23w, 24w and 26w | PGCs and somatic cells | MACS, FACS and Smart-seq2 | 2167 | GSE86146 |
| 2018 | Chen et al. [120] | Tg(Vasa-dTomato; Lin28-YFP) mice (male) | Unspecified | Spermatogenic cells | FACS, modified Smart-seq2 | 1136 | GSE107644 |
| 2018 | Green et al. [121] | C57BL/6J mice, Gfra1CreERT2; RosamT/mG, Amh-cre; RosamT/mG and Sox9-eGFP mice (male) | Adult | Whole testis, interstitial cells, spermatogonia and Sertoli cell enrichment, haploid depletion. | FACS, Drop-Seq | 34,644 | GSE112393 |
| 2018 | Guo et al. [114] | Human (male) | Adult (17, 24 and 25 yo) and infant (13 months old) | Whole testis | Chromium | 7790 | GSE120508 |
| 2018 | Hermann et al. [122] | Human and mice (male) | Human adults (mean = 41.6 yo). Mouse P6 and adults | Spermatogenic cells | FACS, Chromium & SMARTer (C1) | 62,000 | GSE108970, GSE108974, GSE108977, GSE109049, GSE109033 and GSE109037 |
| 2018 | Stévant et al. [21] | Tg(Nr5a1-GFP) mice (male) | E10.5, E11.5, E12.5, E13.5 and E16.5 | Nr5a1+ gonadal somatic cells | FACS, SMARTer (C1) | 400 | GSE97519 |
| 2018 | Vértesy et al. [123] | Human (female) | 8 to 14.4 weeks of development | Whole fetal gonads and adrenal germ cells | Smart-seq2 | 108 | GSE79280 |
| 2018 | Wang et al. [124] | Human adult (male) | Normal: 30 and 60 yo. Obstructive azoospermia: 27, 29, 34, 39, 41, 43 and 44 yo | Whole testis | Smart-seq2 | 2854 | GSE106487 |
| 2018 | Zhang et al. [125] | Human (female) | 24 to 32yo | Oocytes and granulosa cells | Tang et al. [117] | 151 | GSE107746 |
| 2019 | Ernst et al. [126] | C57BL/6J and TC1 mice (male) | P5, P10, P15, P20, P25, P30, P35, Adult (8–9 weeks) | Whole testis | Chromium | 53,510 | E-MTAB-6946 |
| 2019 | Fan et al. [127] | Human (female) | Adult | Ovarian inner cortex | Chromium | 20,676 | GSE118127 |
| 2019 | Grive et al. [128] | B6D2F1/J mice (male) | Postnatal days 6, 14, 18, 25, 30 and 8 weeks old | Whole testis, spermatogonia cells enrichment. | MACS, Chromium | 15,493 | GSE121904 |
| 2019 | Gu et al. [129] | ICR Mouse (female) | 14 days old | Mouse ES cells, MII oocytes and growing oocytes | scCOOL-seq [130] and Tang et al. [117] | 1070 | GSE114822 |
| 2019 | Law et al. [131] | Blimp1-CreTg;Id4-eGFPTg;Rosa26tdTomato-fl_STOP_fl/LacZ mice (male) | E16.5, P0, P3 and P6 | Germ cells | FACS, Chromium | 10,140 | GSE124904 |
| 2019 | Sohni et al. [132] | Human (male) | Neonatal (2 and 7 days old) and adult (37 and 42 yo) | Whole testis | Chromium | 33,585 | GSE124263 |
| 2019 | Stévant et al. [133] | Mouse Tg(Nr5a1-GFP) (female) | E10.5, E11.5, E12.5, E13.5, E16.5 and P6 | Nr5a1+ gonadal somatic cells | FACS, SMARTer (C1) | 563 | GSE119766 |
| 2020 | Estermann et al. [134] | Chicken embryo (male and female) | E4.5, E6.5, E8.5 and E10.5 | Whole gonads | Chromium | 33,247 | GSE143337 |
| 2020 | Guo et al. [115] | Human (male and transfemale) | Male: 7, 11, 13 and 14 yo. Transfemale: 26 and 50 yo | Whole testis | Chromium | 9836 | GSE134144 |
| 2020 | Lau et al. [135] | Macaque (male) | Infant (1 yo), Juvenile (2 yo) and Adult (4 yo) | Whole testis and sorted spermatogonia cells | FACS, Chromium | 16,932 | E-MTAB-8979 |
| 2020 | Niue et al. [34] | Mice (female) | E11.5, E12.5, E14.5, E16.5, E18.5, P1 and P5 | Whole Ovary | Chromium | 52,542 | GSE136441 |
| 2020 | Shami et al. [136] | Human and Macaque (male) | Adult human (20–40 yo), Adult Macaque (4–13 yo) | Whole testis | Drop-seq | 36,000 | GSE142585 |
| 2020 | Tan et al. [137] | C57BL/6 mice (male) | E18.5, P2 and P7 | Whole testis | Chromium | 50,859 | GSE130593 |
| 2020 | Wagner et al. [138] | Human adult (female) | 20–38 yo | Ovarian cortex and DDX4 sorted cell | FACS, Chromium and Smart-seq2 | 24,329 | E-MTAb−8381, E-MTAb−8403 |
| 2020 | Wang et al. [139] | Macaque (female) | Juvenile (4–5 yo) and Aged (18–20 yo) | Whole ovaries | STRT-seq | 2601 | GSE130664 |
| 2020 | Xia et al. [140] | C57BL/6J mice and human (male) | Human adult (40 and 45 yo); 4 months old mice | Whole testis, germ cell enrichment | InDrops | 4147 | GSE125372 |
| Year | Authors | Organism | Developmental Stage | Tissue | Technology | Cells | Data |
|---|---|---|---|---|---|---|---|
| 2016 | Liu et al. [171] | Human (female) with or without POCS | Non-POCS (28.5 ± 3.75 yo). POCS (27.4 ± 3.02 yo) | Oocytes and cumulus cells | Smart-seq2 | 48 | Unspecified |
| 2017 | Winterhoff et al. [172] | Human High-grade serous ovarian cancer (female) | Unspecified | Non-immune tumor cells (epithelial and stromal) | FACS, SMARTer (C1) | 66 | Unspecified |
| 2018 | Chen et al. [120] | Sox30tm1a(KOMP)Wtsi mice (male) | Adult | Spermatogenic cells | FACS sorted, modified Smart-seq2 | 85 | GSE107644 |
| 2018 | Vuong et al. [173] | FVB/N mice (female) | Unspecified | Ovarian surface epithelium cells, untreated or treated with estradiol | SMARTer (C1) | 589 | GSE121957 |
| 2018 | Wang et al. [124] | Human adult (male) | Nonobstructive azoospermia: 24 yo | Whole testis | Smart-seq2 | 174 | GSE106487 |
| 2019 | Fang et al. [174] | C57BL/6J and Akap4-KO mice (male) | 26 weeks old | Whole testis | Chromium | 6804 | SRR9107534 |
| 2019 | Jung et al. [175] | C57BL/6J, B6;CBA-Tg(Pou5f1-EGFP)2Mnn/J, C57BL/6J CNP-EGFP BAC-TRAP mice, Mlh3-/-, Hormad1-/-, Cul4-/- and C57BL/6J CNP eGFP BAC TRAP mice (male) | 11 to 38 weeks old | Whole testis, spermatocytes and spermatids cells enrichment | FACS and Drop-seq | 57,600 | GSE113293 |
| 2019 | Liu et al. [176] | Human male, female and Klinefelter syndrome | Adult | PBMCs | Chromium | 24,439 | GSE136353 |
| 2020 | Guo et al. [115] | Human (male and Transfemale) | Male: 7, 11, 13 and 14 yo. Transfemale: 26 and 50 yo | Whole testis | Chromium | 9836 | GSE134144 |
| 2020 | Izar et al. [177] | Human High-grade serous ovarian cancer (female) | Unspecified | High-grade serous ovarian cancer | FACS, Smart-seq2 and Chromium | 35,957 | GSE146026 |
| 2020 | Nelson et al. [178] | Human Ovarian cancer and stromal cells (female) | 43–81 yo | Cultured ovarian cancer and stromal cells (Biobank) | SMARTer (C1) and Chromium | Unspecified | E-MTAB-8559 |
| 2020 | Park et al. [179] | C57BL/6 and Esr2-PgrKO mice (female) | Unspecified | Whole ovaries | Chromium | 6421 | GSE145107 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estermann, M.A.; Smith, C.A. Applying Single-Cell Analysis to Gonadogenesis and DSDs (Disorders/Differences of Sex Development). Int. J. Mol. Sci. 2020, 21, 6614. https://doi.org/10.3390/ijms21186614
Estermann MA, Smith CA. Applying Single-Cell Analysis to Gonadogenesis and DSDs (Disorders/Differences of Sex Development). International Journal of Molecular Sciences. 2020; 21(18):6614. https://doi.org/10.3390/ijms21186614
Chicago/Turabian StyleEstermann, Martin A., and Craig A. Smith. 2020. "Applying Single-Cell Analysis to Gonadogenesis and DSDs (Disorders/Differences of Sex Development)" International Journal of Molecular Sciences 21, no. 18: 6614. https://doi.org/10.3390/ijms21186614
APA StyleEstermann, M. A., & Smith, C. A. (2020). Applying Single-Cell Analysis to Gonadogenesis and DSDs (Disorders/Differences of Sex Development). International Journal of Molecular Sciences, 21(18), 6614. https://doi.org/10.3390/ijms21186614