Protective Effect of CXCR4 Antagonist CX807 in a Rat Model of Hemorrhagic Stroke

Abstract
:1. Introduction
2. Results
2.1. In Vitro and In Vivo Characterization of CX807 and AMD3100
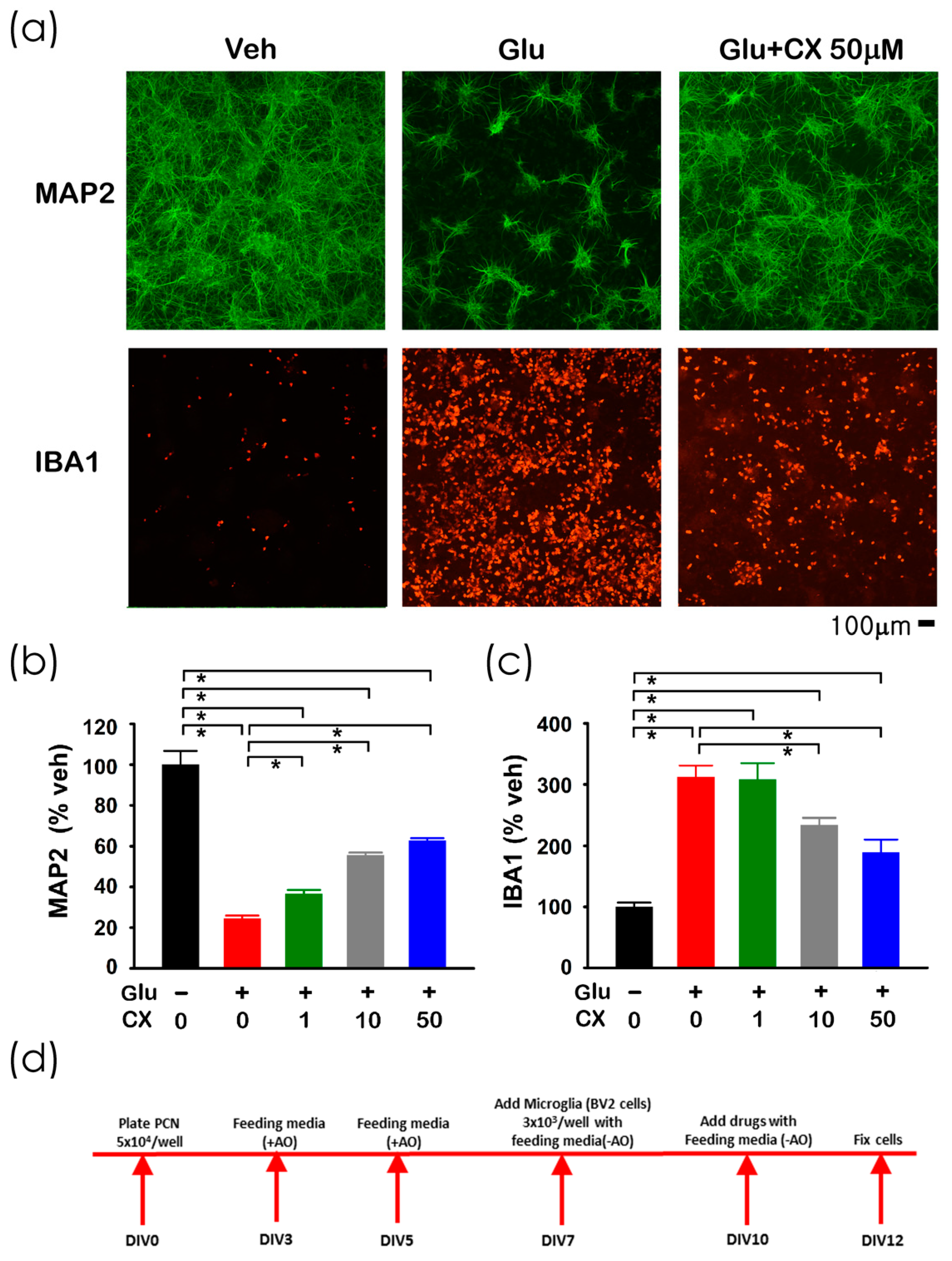
2.2. CX807-Mediated Neuroprotection in Primary Cortical Cells and Microglia Co-Cultures
2.3. CX807 Improved Locomotor Behavior in ICH Rats
2.4. CX807 Reduced ICH-Mediated Expression of CXCR4, TLR4, IL6, CD8, and TNFα
2.5. CX807 Mitigated IBA1, CD8 Immunoreactivity, and TUNEL in ICH Brains
3. Discussion
4. Materials and Methods
4.1. Animals and Material
4.2. CXCR4-Binding Assay
4.3. CXCL12-Induced Chemotaxis Assay
4.4. β-Arrestin Functional Assay
4.5. Pharmacokinetic Analysis and Toxicology Study
4.6. Primary Rat Cortical Neuron (PCN) and Microglia Co-Culture
4.7. Immunocytochemistry
4.8. Collagenase-Induced Intracerebral Hemorrhage
4.9. Locomotor Behavioral Measurement
4.10. Immunohistochemistry
4.11. TUNEL Labeling
4.12. Quantitative Reverse Transcription PCR (qRTPCR)
4.13. Statistics
5. Patents
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| β-Gal | β-galactosidase |
| CXCL12 | C-X-C motif ligand 12 |
| DIVs | days in vitro |
| EDTA | ethylene diamine tetraacetic acid |
| HACTV | horizontal activity |
| IBA1 | ionized calcium-binding adapter molecule 1 |
| ICH | intracerebral hemorrhage |
| MAP2 | microtubule-associated protein 2 |
| MOVTIME | horizontal movements |
| NK test | Newman–Keuls test |
| PCN | primary cortical neuron |
| TLR4 | toll-like receptor 4 |
| TOTDIST | total distance traveled |
| VACTV | vertical activity |
| VTIME | vertical movements time |
| VMOVNO | number of vertical movements |
Appendix A


References
- Tschoe, C.; Bushnell, C.D.; Duncan, P.W.; Alexander-Miller, M.A.; Wolfe, S.Q. Neuroinflammation after Intracerebral Hemorrhage and Potential Therapeutic Targets. J. Stroke 2020, 22, 29–46. [Google Scholar] [CrossRef] [Green Version]
- Lipfert, J.; Odemis, V.; Wagner, D.C.; Boltze, J.; Engele, J. CXCR4 and CXCR7 form a functional receptor unit for SDF-1/CXCL12 in primary rodent microglia. Neuropathol. Appl. Neurobiol. 2013, 39, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.B.; Banisadr, G.; Ren, D.; Chenn, A.; Miller, R.J. Chemokine receptor expression by neural progenitor cells in neurogenic regions of mouse brain. J. Comp. Neurol. 2007, 500, 1007–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stumm, R.K.; Rummel, J.; Junker, V.; Culmsee, C.; Pfeiffer, M.; Krieglstein, J.; Hollt, V.; Schulz, S. A dual role for the SDF-1/CXCR4 chemokine receptor system in adult brain: Isoform-selective regulation of SDF-1 expression modulates CXCR4-dependent neuronal plasticity and cerebral leukocyte recruitment after focal ischemia. J. Neurosci. 2002, 22, 5865–5878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robin, A.M.; Zhang, Z.G.; Wang, L.; Zhang, R.L.; Katakowski, M.; Zhang, L.; Wang, Y.; Zhang, C.; Chopp, M. Stromal cell-derived factor 1alpha mediates neural progenitor cell motility after focal cerebral ischemia. J. Cereb. Blood Flow Metab. 2006, 26, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Hill, W.D.; Hess, D.C.; Martin-Studdard, A.; Carothers, J.J.; Zheng, J.; Hale, D.; Maeda, M.; Fagan, S.C.; Carroll, J.E.; Conway, S.J. SDF-1 (CXCL12) is upregulated in the ischemic penumbra following stroke: Association with bone marrow cell homing to injury. J. Neuropathol. Exp. Neurol. 2004, 63, 84–96. [Google Scholar] [CrossRef] [Green Version]
- Walter, H.L.; van der Maten, G.; Antunes, A.R.; Wieloch, T.; Ruscher, K. Treatment with AMD3100 attenuates the microglial response and improves outcome after experimental stroke. J. Neuroinflamm. 2015, 12, 24. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.J.; Yu, S.J.; Shia, K.S.; Wu, C.H.; Song, J.S.; Kuan, H.H.; Yeh, K.C.; Chen, C.T.; Bae, E.; Wang, Y. A novel CXCR4 antagonist CX549 induces neuroprotection in stroke brain. Cell Transplant. 2017, 26, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Li, Y.; Tang, Y.; Tang, G.; Yang, G.Y.; Wang, Y. CXCR4 antagonist AMD3100 protects blood-brain barrier integrity and reduces inflammatory response after focal ischemia in mice. Stroke 2013, 44, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Ruscher, K.; Kuric, E.; Liu, Y.; Walter, H.L.; Issazadeh-Navikas, S.; Englund, E.; Wieloch, T. Inhibition of CXCL12 signaling attenuates the postischemic immune response and improves functional recovery after stroke. J. Cereb. Blood Flow Metab. 2013, 33, 1225–1234. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Chen, B.; Zheng, G.R.; Qiu, S.Z.; Yin, H.M.; Mao, W.; Wang, H.X.; Gao, J.B. Detection of high serum concentration of CXC chemokine ligand-12 in acute intracerebral hemorrhage. Clin. Chim. Acta 2017, 471, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.J.; Wu, K.J.; Bae, E.; Wang, Y.S.; Chiang, C.W.; Kuo, L.W.; Harvey, B.K.; Greig, N.H.; Wang, Y. Post-treatment with Posiphen reduces endoplasmic reticulum stress and neurodegeneration in stroke brain. iScience 2020, 23, 10866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Chen, G.J.; Harvey, B.K.; Bickford, P.L.; Wang, Y. Inosine reduces ischemic brain injury in rats. Stroke 2005, 36, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Globus, M.Y.; Alonso, O.; Dietrich, W.D.; Busto, R.; Ginsberg, M.D. Glutamate release and free radical production following brain injury: Effects of posttraumatic hypothermia. J. Neurochem. 1995, 65, 1704–1711. [Google Scholar] [CrossRef]
- Ohtani, Y.; Minami, M.; Kawaguchi, N.; Nishiyori, A.; Yamamoto, J.; Takami, S.; Satoh, M. Expression of stromal cell-derived factor-1 and CXCR4 chemokine receptor mRNAs in cultured rat glial and neuronal cells. Neurosci. Lett. 1998, 249, 163–166. [Google Scholar] [CrossRef]
- Lazarini, F.; Casanova, P.; Tham, T.N.; De, C.E.; Arenzana-Seisdedos, F.; Baleux, F.; Dubois-Dalcq, M. Differential signalling of the chemokine receptor CXCR4 by stromal cell-derived factor 1 and the HIV glycoprotein in rat neurons and astrocytes. Eur. J. Neurosci. 2000, 12, 117–125. [Google Scholar] [CrossRef]
- Bezzi, P.; Domercq, M.; Brambilla, L.; Galli, R.; Schols, D.; De, C.E.; Vescovi, A.; Bagetta, G.; Kollias, G.; Meldolesi, J.; et al. CXCR4-activated astrocyte glutamate release via TNFalpha: Amplification by microglia triggers neurotoxicity. Nat. Neurosci. 2001, 4, 702–710. [Google Scholar] [CrossRef]
- Rosenberg, G.A.; Mun-Bryce, S.; Wesley, M.; Kornfeld, M. Collagenase-induced intracerebral hemorrhage in rats. Stroke 1990, 21, 801–807. [Google Scholar] [CrossRef] [Green Version]
- Ajmo, C.T., Jr.; Vernon, D.O.; Collier, L.; Hall, A.A.; Garbuzova-Davis, S.; Willing, A.; Pennypacker, K.R. The spleen contributes to stroke-induced neurodegeneration. J. Neurosci. Res. 2008, 86, 2227–2234. [Google Scholar] [CrossRef] [Green Version]
- Yrjanheikki, J.; Tikka, T.; Keinanen, R.; Goldsteins, G.; Chan, P.H.; Koistinaho, J. A tetracycline derivative, minocycline, reduces inflammation and protects against focal cerebral ischemia with a wide therapeutic window. Proc. Natl. Acad. Sci. USA 1999, 96, 13496–13500. [Google Scholar] [CrossRef] [Green Version]
- Man, S.; Tucky, B.; Cotleur, A.; Drazba, J.; Takeshita, Y.; Ransohoff, R.M. CXCL12-induced monocyte-endothelial interactions promote lymphocyte transmigration across an in vitro blood-brain barrier. Sci. Transl. Med. 2012, 4, 119ra14. [Google Scholar]
- Lu, D.Y.; Tang, C.H.; Yeh, W.L.; Wong, K.L.; Lin, C.P.; Chen, Y.H.; Lai, C.H.; Chen, Y.F.; Leung, Y.M.; Fu, W.M. SDF-1alpha up-regulates interleukin-6 through CXCR4, PI3K/Akt, ERK, and NF-kappaB-dependent pathway in microglia. Eur. J. Pharmacol. 2009, 613, 146–154. [Google Scholar] [PubMed]
- Yao, L.; Kan, E.M.; Lu, J.; Hao, A.; Dheen, S.T.; Kaur, C.; Ling, E.A. Toll-like receptor 4 mediates microglial activation and production of inflammatory mediators in neonatal rat brain following hypoxia: Role of TLR4 in hypoxic microglia. J. Neuroinflamm. 2013, 10, 23. [Google Scholar]
- Yu, S.J.; Reiner, D.; Shen, H.; Wu, K.J.; Liu, Q.R.; Wang, Y. Time-dependent protection of CB2 receptor agonist in stroke. PLoS ONE 2015, 10, e0132487. [Google Scholar]
- Sang, H.; Qiu, Z.; Cai, J.; Lan, W.; Yu, L.; Zhang, H.; Li, M.; Xie, Y.; Guo, R.; Ye, R.; et al. Early Increased Bradykinin 1 Receptor Contributes to Hemorrhagic Transformation After Ischemic Stroke in Type 1 Diabetic Rats. Transl. Stroke Res. 2017, 8, 597–611. [Google Scholar] [PubMed]
- Huang, M.; Wan, Y.; Mao, L.; He, Q.W.; Xia, Y.P.; Li, M.; Li, Y.N.; Jin, H.J.; Hu, B. Inhibiting the Migration of M1 Microglia at Hyperacute Period Could Improve Outcome of tMCAO Rats. CNS Neurosci. Ther. 2017, 23, 222–232. [Google Scholar]
- van Der Lee, M.M.; Bras, M.; van Koppen, C.J.; Zaman, G.J. Beta-Arrestin recruitment assay for the identification of agonists of the sphingosine 1-phosphate receptor EDG1. J. Biomol. Screen. 2008, 13, 986–998. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p-Value | F Value | |||
|---|---|---|---|---|
| veh + ICH vs. | CX807 + ICH vs. | F2,105 = | ||
| no ICH | veh + ICH | no ICH | ||
| HACTV | <0.001 | <0.001 | 0.592 | 15.569 |
| MOVTIME | <0.001 | <0.001 | 0.803 | 18.172 |
| TOTDIST | <0.001 | <0.001 | 0.524 | 20.988 |
| VACTV | <0.001 | <0.001 | 0.916 | 13.322 |
| VTIME | 0.003 | <0.001 | 0.311 | 9.928 |
| VMOVNO | <0.001 | <0.001 | 0.401 | 13.499 |
| Gene | SYBR Green | TagMan | |
|---|---|---|---|
| Forward | Reverse | ||
| CXCR4 | ATCATCTCCAAGCTGTCACACTCC | GTGATGGAGATCCACTTGTGCAC | |
| CD8a | ACACTTCGCAAGGATGCTCT | GTTGCTGGTGATTGAGCAGA | |
| CD8b | GTCTTTGGGACAGGGACAAA | GAGGGATACCAGCAGAACCA | |
| TNF-α | CCACACCGTCAGCCGATT | TCCTTAGGGCAAGGGCTCTT | |
| IL-6 | GACCAAGACCATCCAACT | TAGGTTTGCCGAGTAGAC | |
| TLR4 | AGCTTTGGTCAGTTGGCTCT | CAGGATGACACCATTGAAGC | |
| β-Actin | Rn00667869_m1 | ||
| GAPDH | Rn01775763_g1 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.-J.; Wu, K.-J.; Wang, Y.-S.; Song, J.-S.; Wu, C.-H.; Jan, J.-J.; Bae, E.; Chen, H.; Shia, K.-S.; Wang, Y. Protective Effect of CXCR4 Antagonist CX807 in a Rat Model of Hemorrhagic Stroke. Int. J. Mol. Sci. 2020, 21, 7085. https://doi.org/10.3390/ijms21197085
Yu S-J, Wu K-J, Wang Y-S, Song J-S, Wu C-H, Jan J-J, Bae E, Chen H, Shia K-S, Wang Y. Protective Effect of CXCR4 Antagonist CX807 in a Rat Model of Hemorrhagic Stroke. International Journal of Molecular Sciences. 2020; 21(19):7085. https://doi.org/10.3390/ijms21197085
Chicago/Turabian StyleYu, Seong-Jin, Kuo-Jen Wu, Yu-Syuan Wang, Jen-Shin Song, Chien-Huang Wu, Jiing-Jyh Jan, Eunkyung Bae, Hsi Chen, Kak-Shan Shia, and Yun Wang. 2020. "Protective Effect of CXCR4 Antagonist CX807 in a Rat Model of Hemorrhagic Stroke" International Journal of Molecular Sciences 21, no. 19: 7085. https://doi.org/10.3390/ijms21197085
APA StyleYu, S. -J., Wu, K. -J., Wang, Y. -S., Song, J. -S., Wu, C. -H., Jan, J. -J., Bae, E., Chen, H., Shia, K. -S., & Wang, Y. (2020). Protective Effect of CXCR4 Antagonist CX807 in a Rat Model of Hemorrhagic Stroke. International Journal of Molecular Sciences, 21(19), 7085. https://doi.org/10.3390/ijms21197085