The Class III Peroxidase Encoding Gene AtPrx62 Positively and Spatiotemporally Regulates the Low pH-Induced Cell Death in Arabidopsis thaliana Roots

 ,
, 
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cell Death, CIII Prx Activity and ROS Distribution Colocalized in Wild-Type Roots Exposed to Low pH
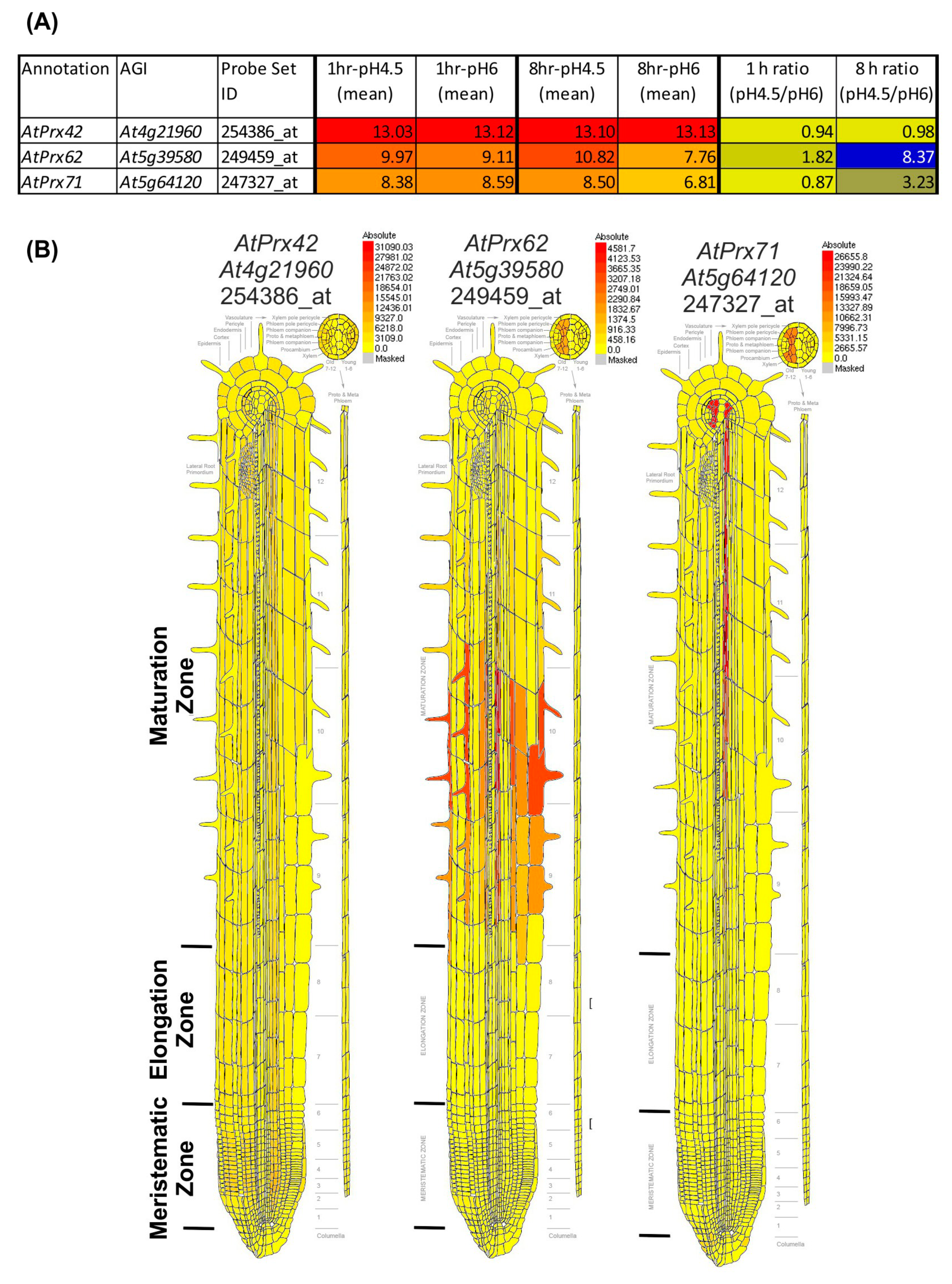
2.2. Data Mining of Published Transcriptomics Data Searching for CIII Prx Genes Expression upon Low pH Treatment: Identification of Candidate Genes of Interest
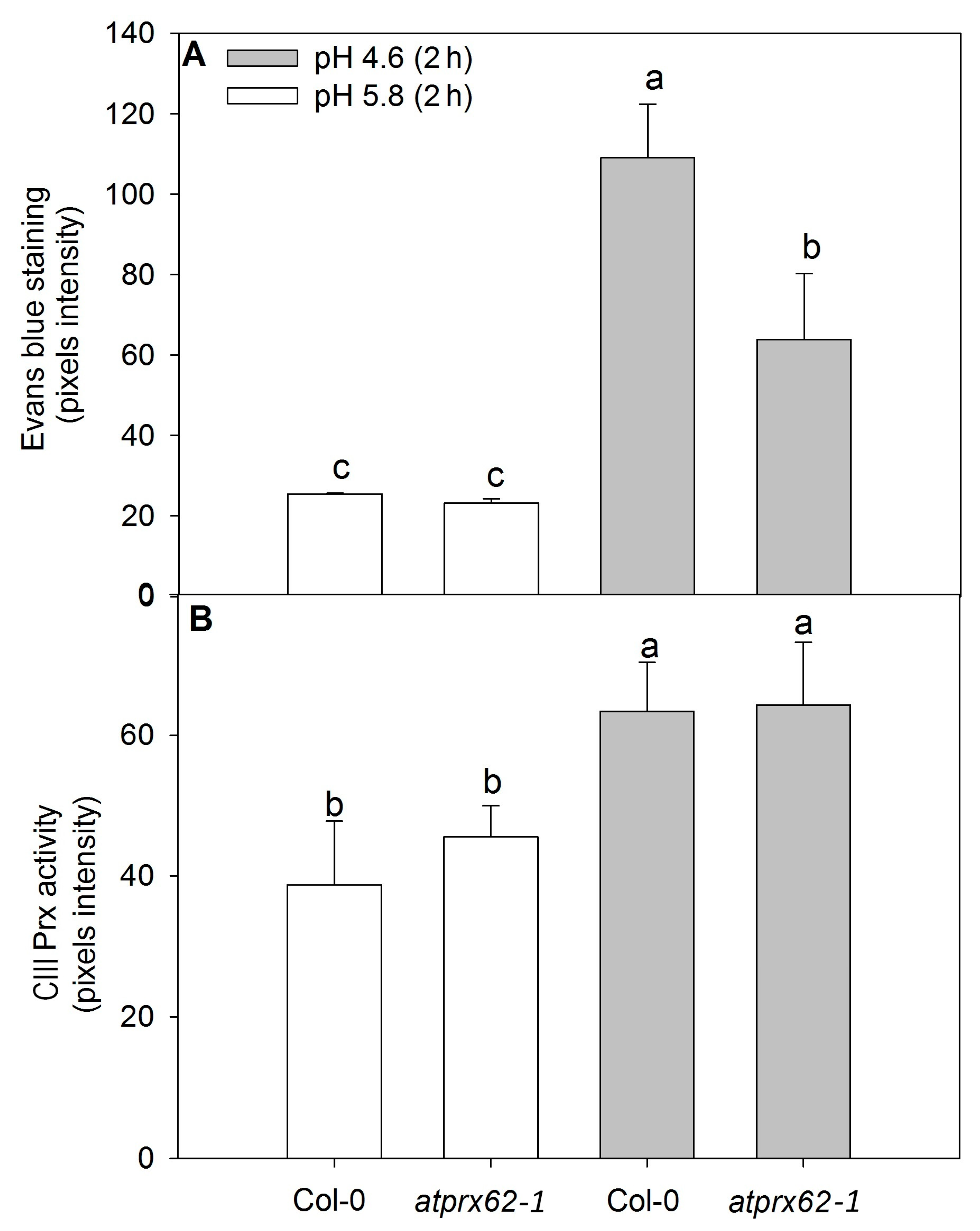
2.3. Cell Viability and Total CIII Prx Activity in Atprx62 and Atprx71 Mutants Exposed to Low pH
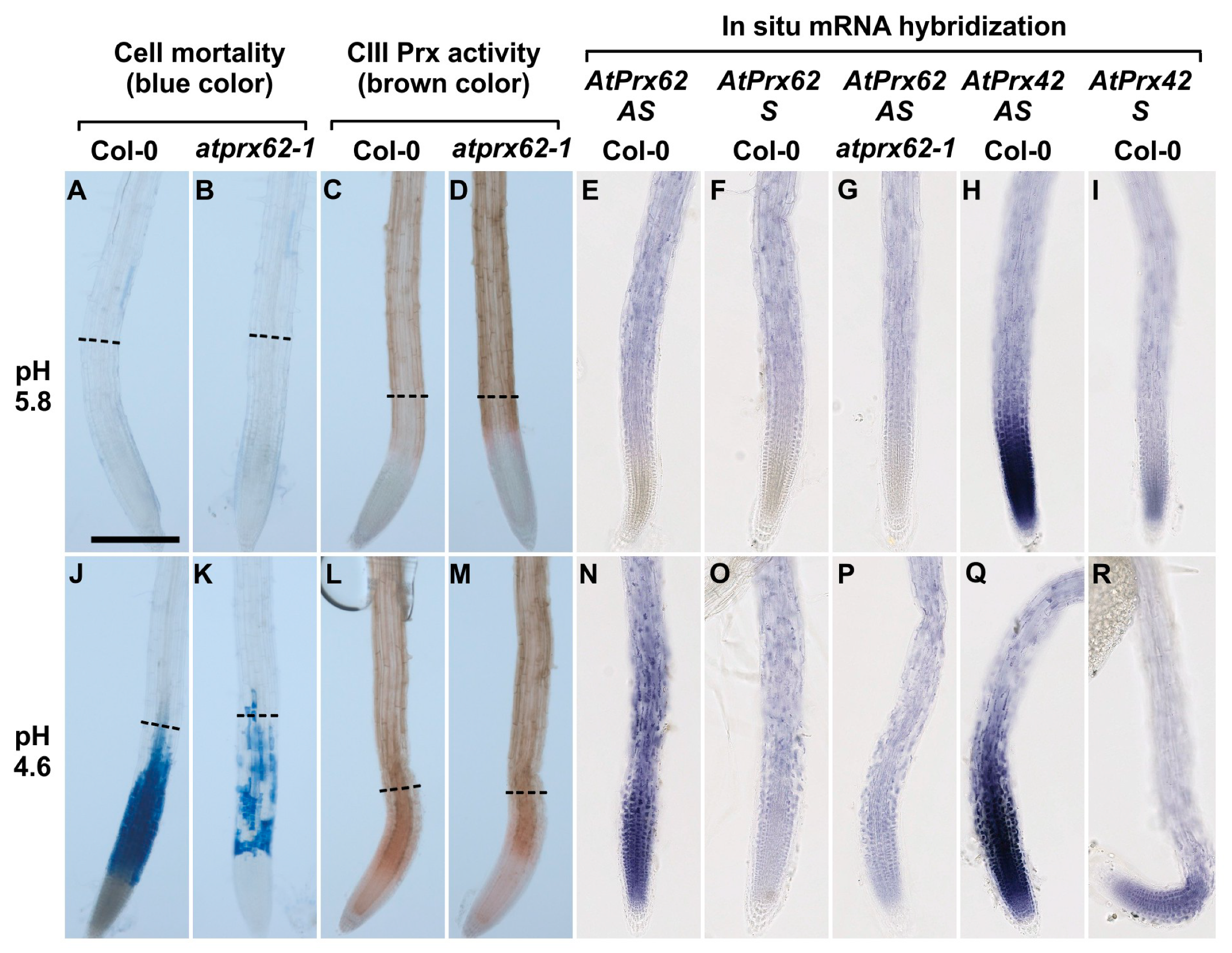
2.4. Tissue-Specific Expression Patterns of AtPrx62 and AtPrx42 in Roots upon Low pH Treatment
3. Discussion
3.1. AtPrx62 Expression Is Spatiotemporally Correlated to Low pH-Induced Cell Death in Roots
3.2. Low pH Disrupts the O2●−/H2O2 Homeostasis in Roots
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Evaluation of Total CIII Prx Activity and ROS Distribution in Roots Exposed to Low pH
4.3. Transcriptomic Data Mining for CIII Prx Genes Involved in Low pH Response
4.4. Evaluation of Cell Death
4.5. Image Analysis
4.6. Whole Mount In Situ mRNA Hybridization
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CIII Prx | Class III Peroxidase |
| CW | Cell Wall |
| CWD | Cell Wall Disturbance |
| CWI | Cell Wall Integrity |
| EZ | Elongation zone |
| MZ | Meristematic zone |
| PCD | Programmed Cell Death |
| ROS | Reactive Oxygen Species |
| TZ | Transition zone |
References
- Von Uexküll, H.R.; Mutert, E. Global extent, development and economic impact of acid soils. Plant Soil. 1995, 171, 1–15. [Google Scholar] [CrossRef]
- Koyama, H.; Toda, T.; Hara, T. Brief exposure to low-pH stress causes irreversible damage to the growing root in Arabidopsis thaliana: Pectin-Ca interaction may play an important role in proton rhizotoxicity. J. Exp. Bot. 2001, 52, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Kochian, L.V. Cellular mechanisms of aluminum toxicity and resistance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 237–260. [Google Scholar] [CrossRef]
- Kochian, L.V.; Piñeros, M.A.; Liu, J.; Magalhães, J.V. Plant adaption to acid soils: The molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef] [PubMed]
- Graças, J.P.; Ruiz-Romero, R.; Figueiredo, L.D.; Mattiello, L.; Peres, L.E.P.; Vitorello, V.A. Root growth restraint can be an acclimatory response to low pH and is associated with reduced cell mortality: A possible role of class III peroxidases and NADPH oxidases. Plant Biol. 2016, 18, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Kinraide, T.B.; Ryan, P.R.; Kochian, L.V. Interactive effects of Al3+, H+, and other cations on root elongation considered in terms of cell-surface electrical potential. Plant Physiol. 1992, 99, 1461–1468. [Google Scholar] [CrossRef] [Green Version]
- Bibikova, T.N.; Jacob, T.; Dahse, I.; Gilroy, S. Localized changes in apoplastic and cytoplasmic pH are associated with root hair development in Arabidopsis thaliana. Development 1998, 125, 2925–2934. [Google Scholar]
- Kobayashi, Y.; Kobayashi, Y.; Watanabe, T.; Shaff, J.E.; Ohta, H.; Kochian, L.V.; Wagatsuma, T.; Kinraide, T.B.; Koyama, H. Molecular and physiological analysis of Al3+ and H+ rhizotoxicities at moderately acidic conditions. Plant Physiol. 2013, 163, 180–192. [Google Scholar] [CrossRef] [Green Version]
- Graças, J.P.; Lima, J.E.; Peres, L.E.P.; Jamet, E.; Dunand, C.; Vitorello, V.A.; Chervin, C. Ethylene signaling causing tolerance of Arabidopsis thaliana roots to low pH stress is linked to class III peroxidase activity. J. Plant Growth Regul. 2020. [Google Scholar] [CrossRef]
- Verbelen, J.-P.; Cnodder, T.; Le, J.; Vissemberg, K.; Baluska, F. The root apex of Arabidopsis thaliana consists of four distinct zones of growth activities. Plant Signal. Behav. 2006, 6, 296–304. [Google Scholar] [CrossRef] [Green Version]
- Baluska, F.; Volkmann, D.; Barlow, P.W. Specialized zones of development in roots: View from cellular level. Plant Physiol. 1996, 112, 3–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somssich, M.; Khan, G.A.; Persson, S. Cell wall heterogeneity in root development of Arabidopsis. Front. Plant Sci. 2016, 7, 1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monshausen, G.B.; Bibikova, T.N.; Messerli, M.A.; Shi, C.; Gilroy, S. Oscillations in extracellular pH and reactive oxygen species modulate tip growth of Arabidopsis root hairs. Proc. Natl. Acad. Sci. USA 2007, 104, 20996–21001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanimoto, E.; Fujii, S.; Yamamoto, R.; Inanaga, S. Measurement of viscoelastic properties of root cell walls affected by low pH in lateral roots of Pisum sativum L. Plant Soil. 2000, 226, 21–28. [Google Scholar] [CrossRef]
- Willats, W.G.T.; McCartney, L.; Mackie, W.; Knox, J.P. Pectin: Cell biology and prospects for functional analysis. Plant Mol. Biol. 2001, 47, 9–27. [Google Scholar] [CrossRef]
- Voxeur, A.; Hofte, H. Cell wall integrity signaling in plants: “To grow or not to grow that’s the question”. Glycobiology 2016, 26, 950–960. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Ohyama, Y.; Ito, H.; Iuchi, S.; Fujita, M.; Zhao, C.R.; Tanveer, T.; Ganesan, M.; Kobayashi, M.; Koyama, H. STOP2 activates transcription of several genes for Al- and low pH-tolerance that are regulated by STOP1 in Arabidopsis. Mol. Plant 2014, 7, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Lager, I.; Andreasson, O.; Dunbar, T.L.; Andreasson, E.; Escobar, M.A.; Rasmusson, A.G. Changes in external pH rapidly alter plant gene expression and modulate auxin and elicitor responses. Plant Cell Environ. 2010, 33, 1513–1528. [Google Scholar] [CrossRef] [Green Version]
- Graças, J.P. The cell wall is crucial for cellular sensitivity to low pH: The role of class III peroxidases and ethylene in cell death in Arabidopsis thaliana roots. Ph.D. Thesis, Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo, Butantã, São Paulo, Brazil, 2018. [Google Scholar] [CrossRef] [Green Version]
- Francoz, E.; Ranocha, P.; Nguyen-Kim, H.; Jamet, E.; Burlat, V.; Dunand, C. Roles of cell wall peroxidases in plant development. Phytochemistry 2015, 112, 15–21. [Google Scholar] [CrossRef]
- Valerio, L.; De Meyer, M.; Penel, C.; Dunand, C. Expression analysis of the Arabidopsis peroxidase multigenic family. Phytochemistry 2004, 65, 1331–1342. [Google Scholar] [CrossRef]
- Podgorska, A.; Burian, M.; Szal, B. Extra-cellular but extra-ordinarily important for cells: Apoplastic reactive oxygen species metabolism. Front. Plant Sci. 2017, 8, 1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.F.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.G.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Marino, D.; Dunand, C.; Puppo, A.; Pauly, N. A burst of plant NADPH oxidase. Trends Plant Sci. 2012, 17, 9–15. [Google Scholar] [CrossRef]
- Passardi, F.; Penel, C.; Dunand, C. Performing the paradoxical: How plant peroxidases modify the cell wall. Trends Plant Sci. 2004, 9, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.X.; Schopfer, P. Hydroxyl-radical production in physiological reactions—A novel function of peroxidase. Eur. J. Biochem. 1999, 260, 726–735. [Google Scholar] [CrossRef]
- Dunand, C.; Crevecoeur, M.; Penel, C. Distribution of superoxide and hydrogen peroxide in Arabidopsis root and their influence on root development: Possible interaction with peroxidases. New Phytol. 2007, 174, 332–341. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.J.; Oo, T.L.; He, H.Y.; Wang, A.Q.; Zhan, J.; Li, C.Z.; Wei, S.Q.; He, L.F. Aluminum induces rapidly mitochondria-dependent programmed cell death in Al-sensitive peanut root tips. Bot. Stud. 2014, 55, 67. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.J.; Yang, X.D.; Yao, S.C.; LwinOo, T.; He, H.Y.; Wang, A.Q.; Li, C.Z.; He, L.F. Reactive oxygen species burst induced by aluminum stress triggers mitochondria-dependent programmed cell death in peanut root tip cells. Plant Physiol. Biochem. 2014, 82, 76–84. [Google Scholar] [CrossRef]
- Simonovicova, M.; Huttova, J.; Mistrik, I.; Siroka, B.; Tamas, L. Root growth inhibition by aluminum is probably caused by cell death due to peroxidase-mediated hydrogen peroxide production. Protoplasma 2004, 224, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Kim, H.; Clemente, H.S.; Balliau, T.; Zivy, M.; Dunand, C.; Albenne, C.; Jamet, E. Arabidopsis thaliana root cell wall proteomics: Increasing the proteome coverage using a combinatorial peptide ligand library and description of unexpected Hyp in peroxidase amino acid sequences. Proteomics 2016, 16, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Tognolli, M.; Penel, C.; Greppin, H.; Simon, P. Analysis and expression of the class III peroxidase large gene family in Arabidopsis thaliana. Gene 2002, 288, 129–138. [Google Scholar] [CrossRef]
- Brady, S.M.; Orlando, D.A.; Lee, J.Y.; Wang, J.Y.; Koch, J.; Dinneny, J.R.; Mace, D.; Ohler, U.; Benfey, P.N. A high-resolution root spatiotemporal map reveals dominant expression patterns. Science 2007, 318, 801–816. [Google Scholar] [CrossRef]
- Jemmat, M.A.; Ranocha, P.; Le Ru, A.; Neel, M.; Jauneau, A.; Raggi, S.; Ferrari, S.; Burlat, V.; Dunand, C. Coordination of five class III peroxidase-encoding genes for early germination events of Arabidopsis thaliana. Plant Sci. 2020, 298, 110565. [Google Scholar] [CrossRef]
- Redoxibase. Available online: http://peroxibase.toulouse.inra.fr/ (accessed on 28 September 2020).
- Arabidopsis electronic fluorescent pictographic (eFP) browser. Available online: http://bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi?dataSource=Root (accessed on 28 September 2020).
- Sawaki, Y.; Iuchi, S.; Kobayashi, Y.; Ikka, T.; Sakurai, N.; Fujita, M.; Shinozaki, K.; Shibata, D.; Kobayashi, M.; Koyama, H. STOP1 regulates multiple genes that protect Arabidopsis from proton and aluminum toxicities. Plant Physiol. 2009, 150, 281–294. [Google Scholar] [CrossRef] [Green Version]
- Iuchi, S.; Koyama, H.; Iuchi, A.; Kobayashi, Y.; Kitabayashi, S.; Ikka, T.; Hirayama, T.; Shinozaki, K.; Kobayashi, M. Zinc finger protein STOP1 is critical for proton tolerance in Arabidopsis and coregulates a key gene in aluminum tolerance. Proc. Natl. Acad. Sci. USA 2007, 104, 9900–9905. [Google Scholar] [CrossRef] [Green Version]
- Kumari, M.; Taylor, G.J.; Deyholos, M.K. Transcriptomic responses to aluminum stress in roots of Arabidopsis thaliana. Mol. Genet. Genom. 2008, 279, 339–357. [Google Scholar] [CrossRef]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a Swiss army knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef]
- Raggi, S.; Ferrarini, A.; Delledonne, M.; Dunand, C.; Ranocha, P.; De Lorenzo, G.; Cervone, F.; Ferrari, S. The Arabidopsis class III peroxidase AtPRX71 negatively regulates growth under physiological conditions and in response to cell wall damage. Plant Physiol. 2015, 169, 2513–2525. [Google Scholar] [CrossRef] [Green Version]
- Urade, R. Cellular response to unfolded proteins in the endoplasmic reticulum of plants. FEBS J. 2007, 274, 1152–1171. [Google Scholar] [CrossRef] [PubMed]
- Martinez, I.M.; Chrispeels, M.J. Genomic analysis of the unfolded protein response in Arabidopsis shows its connection to important cellular processes. Plant Cell. 2003, 15, 561–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brutus, A.; Sicilia, F.; Macone, A.; Cervone, F.; Lorenzo, G.D. A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides. Proc. Natl. Acad. Sci. USA 2010, 107, 9452–9457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidsson, P.; Broberg, M.; Kariola, T.; Sipari, N.; Pirhonen, M.; Palva, E.T. Short oligogalacturonides induce pathogen resistance-associated gene expression in Arabidopsis thaliana. BMC Plant Biol. 2017, 19, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S.; Greiner, S. Growth control by cell wall pectins. Protoplasma 2012, 249, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Moscatiello, R.; Mariani, P.; Sanders, D.; Maathuis, F.J.M. Transcriptional analysis of calcium-dependent and calcium-independent signalling pathways induced by oligogalacturonides. J. Exp. Bot. 2006, 57, 2847–2865. [Google Scholar] [CrossRef]
- Yang, C.Y.; Hsu, F.C.; Li, J.P.; Wang, N.N.; Shih, M.C. The AP2/ERF transcription factor AtERF73/HRE1 modulates ethylene responses during hypoxia in Arabidopsis. Plant Physiol. 2011, 156, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Waszczak, C.; Carmody, M.; Kangasjarvi, J. Reactive oxygen species in plant signaling. Ann. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- European Arabidopsis Stock Center. Available online: http://arabidopsis.info/ (accessed on 28 September 2020).
- Shah, K.; Penel, C.; Gagnon, J.; Dunand, C. Purification and identification of a Ca2+-pectate binding peroxidase from Arabidopsis leaves. Phytochemistry 2004, 65, 307–312. [Google Scholar] [CrossRef]
- NASC’s International Affymetrix Service. Available online: http://arabidopsis.info/affy (accessed on 28 September 2020).
- Affymetrix. Available online: http://www.affymetrix.com (accessed on 28 September 2020).
- Microsoft Excell. Available online: https://www.microsoft.com (accessed on 28 September 2020).
- Francoz, E.; Ranocha, P.; Pernot, C.; Le Ru, A.; Pacquit, V.; Dunand, C.; Burlat, V. Complementarity of medium-throughput in situ RNA hybridization and tissue-specific transcriptomics: Case study of Arabidopsis seed development kinetics. Sci. Rep. 2016, 6, 24644. [Google Scholar] [CrossRef]
- Baker, C.J.; Mock, N.M. An improved method for monitoring cell-death in cell-suspension and leaf disc assays using Evans blue. Plant Cell Tissue Organ. Cult. 1994, 39, 7–12. [Google Scholar] [CrossRef]
- ImageJ software. Available online: https://imagej.nih.gov/ij/docs/guide/index.html (accessed on 28 September 2020).
- Hejatko, J.; Blilou, I.; Brewer, P.B.; Friml, J.; Scheres, B.; Benkova, E. In situ hybridization technique for mRNA detection in whole mount Arabidopsis samples. Nat. Protoc. 2006, 1, 1939–1946. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graças, J.P.; Ranocha, P.; Vitorello, V.A.; Savelli, B.; Jamet, E.; Dunand, C.; Burlat, V. The Class III Peroxidase Encoding Gene AtPrx62 Positively and Spatiotemporally Regulates the Low pH-Induced Cell Death in Arabidopsis thaliana Roots. Int. J. Mol. Sci. 2020, 21, 7191. https://doi.org/10.3390/ijms21197191
Graças JP, Ranocha P, Vitorello VA, Savelli B, Jamet E, Dunand C, Burlat V. The Class III Peroxidase Encoding Gene AtPrx62 Positively and Spatiotemporally Regulates the Low pH-Induced Cell Death in Arabidopsis thaliana Roots. International Journal of Molecular Sciences. 2020; 21(19):7191. https://doi.org/10.3390/ijms21197191
Chicago/Turabian StyleGraças, Jonathas Pereira, Philippe Ranocha, Victor Alexandre Vitorello, Bruno Savelli, Elisabeth Jamet, Christophe Dunand, and Vincent Burlat. 2020. "The Class III Peroxidase Encoding Gene AtPrx62 Positively and Spatiotemporally Regulates the Low pH-Induced Cell Death in Arabidopsis thaliana Roots" International Journal of Molecular Sciences 21, no. 19: 7191. https://doi.org/10.3390/ijms21197191
APA StyleGraças, J. P., Ranocha, P., Vitorello, V. A., Savelli, B., Jamet, E., Dunand, C., & Burlat, V. (2020). The Class III Peroxidase Encoding Gene AtPrx62 Positively and Spatiotemporally Regulates the Low pH-Induced Cell Death in Arabidopsis thaliana Roots. International Journal of Molecular Sciences, 21(19), 7191. https://doi.org/10.3390/ijms21197191