MiRNAs as Noninvasive Biomarkers and Therapeutic Agents of Pituitary Adenomas



 , ,
, , 
Abstract
:1. Introduction
2. The Current State of miRNA Research in PAs
3. Circulating miRNAs and PA
4. miRNAs and PA Therapy
5. Challenges of miRNAs as Diagnostic/Therapeutic Tools
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TRAIL | The tumor necrosis factor-related apoptosis-inducing ligand |
| mTOR | The mammalian target of rapamycin |
| cRGD | Cyclic Arg-Gly-Asp peptide |
| MMQ | Prolactin secreting rat tumor cell line |
| NCI-H727, CNDT2 | Neuroendocrine tumor cell lines. |
References
- Ho, D.; Liu, H. Prolactin-Containing Pituitary Adenomas. Pathol. Res. Pr. 1992, 188, 296–302. [Google Scholar] [CrossRef]
- Lake, M.G.; Krook, L.S.; Cruz, S.V. Pituitary adenomas: An overview. Am. Fam. Physician 2013, 88, 319–327. [Google Scholar] [PubMed]
- Nadhamuni, V.S.; Korbonits, M. Novel insights into Pituitary Tumorigenesis: Genetic and Epigenetic Mechanisms. Endocr. Rev. 2020. [Google Scholar] [CrossRef] [Green Version]
- Mikhaleva, L.M.; Davydov, A.I.; Patsap, O.I.; Mikhaylenko, E.V.; Nikolenko, V.N.; Neganova, M.E.; Klochkov, S.G.; Somasundaram, S.G.; Kirkland, C.E.; Aliev, G. Malignant Transformation and Associated Biomarkers of Ovarian Endometriosis: A Narrative Review. Adv. Ther. 2020, 37, 2580–2603. [Google Scholar] [CrossRef] [PubMed]
- Fung, E.C.; Butt, A.N.; Eastwood, J.; Swaminathan, R.; Sodi, R. Circulating microRNA in cardiovascular disease. In Advances in Applied Microbiology; Elsevier BV: Amsterdam, The Netherlands, 2019; Volume 91, pp. 99–122. [Google Scholar]
- Russo, F.; Di Bella, S.; Vannini, F.; Berti, G.; Scoyni, F.; Cook, H.; Santos, A.; Nigita, G.; Bonnici, V.; Laganà, A.; et al. miRandola 2017: A curated knowledge base of non-invasive biomarkers. Nucleic Acids Res. 2018, 46, D354–D359. [Google Scholar] [CrossRef] [Green Version]
- Meigs, T.E.; Lyakhovich, A.; Shim, H.; Dupré, D.J.; Hébert, T.E.; Blumer, J.B.; Tall, G.G.; Vaillancourt, R.; Spilker, A.C.; Park, M.; et al. G630042G04. In Encyclopedia of Signaling Molecules; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2012; p. 722. [Google Scholar]
- Feng, Y.; Mao, Z.-G.; Wang, X.; Du, Q.; Jian, M.; Zhu, D.; Xiao, Z.; Wang, H.-J.; Zhu, Y.-H. MicroRNAs and Target Genes in Pituitary Adenomas. Horm. Metab. Res. 2018, 50, 179–192. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev. Boil. 2007, 302, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Swier, L.J.Y.M.; Dzikiewicz-Krawczyk, A.; Winkle, M.; Berg, A.V.D.; Kluiver, J. Intricate crosstalk between MYC and non-coding RNA s regulates hallmarks of cancer. Mol. Oncol. 2018, 13, 26–45. [Google Scholar] [CrossRef] [Green Version]
- Acunzo, M.; Visone, R.; Romano, G.; Veronese, A.; Lovat, F.; Palmieri, D.; Bottoni, A.; Garofalo, M.; Gasparini, P.; Condorelli, G.; et al. miR-130a targets MET and induces TRAIL-sensitivity in NSCLC by downregulating miR-221 and 222. Oncogene 2011, 31, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.-B.; Xue, L.; Yang, J.; Ma, A.-H.; Zhao, J.; Xu, M.; Tepper, C.G.; Evans, C.P.; Kung, H.-J.; White, R.W.D.V. An androgen-regulated miRNA suppresses Bak1 expression and induces androgen-independent growth of prostate cancer cells. Proc. Natl. Acad. Sci. USA 2007, 104, 19983–19988. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Liu, Z.; Zhao, Y.; Ding, Y.; Liu, H.; Xi, Y.; Xiong, W.; Li, G.; Lu, J.; Fodstad, O.; et al. MicroRNA-125b Confers the Resistance of Breast Cancer Cells to Paclitaxel through Suppression of Pro-apoptotic Bcl-2 Antagonist Killer 1 (Bak1) Expression. J. Boil. Chem. 2010, 285, 21496–21507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lal, A.; Kim, H.H.; Abdelmohsen, K.; Kuwano, Y.; Pullmann, R.; Srikantan, S.; Subrahmanyam, R.; Martindale, J.L.; Yang, X.; Ahmed, F.; et al. p16INK4a Translation Suppressed by miR-24. PLoS ONE 2008, 3, e1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhas, A.; Saunders, N.J.; Vaux, D.J. The nuclear envelope can control gene expression and cell cycle progression via miRNA regulation. Cell Cycle 2010, 9, 531–539. [Google Scholar] [CrossRef]
- Hu, W.; Chan, C.S.; Wu, R.; Zhang, C.; Sun, Y.; Song, J.; Tang, L.H.; Levine, A.J.; Feng, Z. Negative Regulation of Tumor Suppressor p53 by MicroRNA miR-504. Mol. Cell 2010, 38, 689–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frixa, T.; Donzelli, S.; Blandino, G. Oncogenic MicroRNAs: Key Players in Malignant Transformation. Cancers 2015, 7, 2466–2485. [Google Scholar] [CrossRef]
- Grzywa, T.M.; Klicka, K.; Rak, B.; Mehlich, D.; Garbicz, F.; Zieliński, G.; Maksymowicz, M.; Sajjad, E.; Wlodarski, P. Lineage-dependent role of miR-410-3p as oncomiR in gonadotroph and corticotroph pituitary adenomas or tumor suppressor miR in somatotroph adenomas via MAPK, PTEN/AKT, and STAT3 signaling pathways. Endocrine 2019, 65, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Zhen, W.; Qiu, D.; Zhiyong, C.; Xin, W.; Mengyao, J.; Dimin, Z.; Chonghui, H.; Haijun, W.; Yonghong, Z. MicroRNA-524-5p Functions as a Tumor Suppressor in a Human Pituitary Tumor-Derived Cell Line. Horm. Metab. Res. 2017, 49, 550–557. [Google Scholar] [CrossRef]
- Butz, H.; Németh, K.; Czenke, D.; Likó, I.; Czirják, S.; Zivkovic, V.; Baghy, K.; Korbonits, M.; Kovalszky, I.; Igaz, P. Systematic investigation of expression of G2/M transition genes reveals CDC25 alteration in nonfunctioning pituitary adenomas. Pathol. Oncol. Res. 2017, 23, 633–641. [Google Scholar] [CrossRef] [Green Version]
- Dénes, J.; Kasuki, L.; Trivellin, G.; Colli, L.M.; Takiya, C.M.; Stiles, C.E.; Barry, S.; De Castro, M.; Gadelha, M.R.; Korbonits, M. Regulation of Aryl Hydrocarbon Receptor Interacting Protein (AIP) Protein Expression by MiR-34a in Sporadic Somatotropinomas. PLoS ONE 2015, 10, e0117107. [Google Scholar] [CrossRef]
- Müssnich, P.; Raverot, G.; Jaffrain-Rea, M.-L.; Fraggetta, F.; Wierinckx, A.; Trouillas, J.; Fusco, A.; D’Angelo, D. Downregulation of miR-410 targeting the cyclin B1 gene plays a role in pituitary gonadotroph tumors. Cell Cycle 2015, 14, 2590–2597. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Li, J.; Sun, F.; Cui, J.; Fang, H.; Sui, G. Expression and Clinical Significance of miR-26a and Pleomorphic Adenoma Gene 1 (PLAG1) in Invasive Pituitary Adenoma. Med. Sci. Monit. 2016, 22, 5101–5108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leone, V.; Langella, C.; D’Angelo, D.; Mussnich, P.; Wierinckx, A.; Terracciano, L.; Raverot, G.; Lachuer, J.; Rotondi, S.; Jaffrain-Rea, M.-L.; et al. miR-23b and miR-130b expression is downregulated in pituitary adenomas. Mol. Cell. Endocrinol. 2014, 390, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Zhang, Y.; Zhang, Z.; Yang, Y.; Song, T. Effect of miR-106b on Invasiveness of Pituitary Adenoma via PTEN-PI3K/AKT. Med. Sci. Monit. 2017, 23, 1277–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, M.; Zhang, M.; Liu, H.-F.; Wang, J.-P. Effects of microRNA-21 targeting PITX2 on proliferation and apoptosis of pituitary tumor cells. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2995–3004. [Google Scholar] [PubMed]
- Renjie, W.; Haiqian, L. MiR-132, miR-15a and miR-16 synergistically inhibit pituitary tumor cell proliferation, invasion and migration by targeting Sox5. Cancer Lett. 2015, 356, 568–578. [Google Scholar] [CrossRef]
- Liao, C.; Chen, W.; Fan, X.; Jiang, X.; Qiu, L.; Chen, C.; Zhu, Y.; Wang, H. MicroRNA-200c Inhibits Apoptosis in Pituitary Adenoma Cells by Targeting the PTEN/Akt Signaling Pathway. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2014, 21, 129–136. [Google Scholar] [CrossRef]
- Liang, H.-q.; Wang, R.-j.; Diao, C.-f.; Li, J.-w.; Su, J.-l.; Zhang, S. The PTTG1-targeting miRNAs miR-329, miR-300, miR-381, and miR-655 inhibit pituitary tumor cell tumorigenesis and are involved in a p53/PTTG1 regulation feedback loop. Oncotarget 2015, 6, 29413. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Zhou, C.; Liu, M.; Yao, Y.; Sun, J.; Xiao, J.; Ma, W.; Zhu, H.; Wang, R. MicroRNA involvement in a metastatic non-functioning pituitary carcinoma. Pituitary 2015, 18, 710–721. [Google Scholar] [CrossRef]
- Roche, M.; Wierinckx, A.; Croze, S.; Rey, C.; Legras-Lachuer, C.; Morel, A.-P.; Fusco, A.; Raverot, G.; Trouillas, J.; Lachuer, J. Deregulation of miR-183 and KIAA0101 in Aggressive and Malignant Pituitary Tumors. Front. Med. 2015, 2, 54. [Google Scholar] [CrossRef] [Green Version]
- Morello, V.; Cabodi, S.; Sigismund, S.; Camacho-Leal, M.P.; Repetto, D.; Volante, M.; Papotti, M.; Turco, E.; Defilippi, P. β1 integrin controls EGFR signaling and tumorigenic properties of lung cancer cells. Oncogene 2011, 30, 4087–4096. [Google Scholar] [CrossRef] [Green Version]
- Dunn, B.K.; Wagner, P.D.; Anderson, D.; Greenwald, P. Molecular Markers for Early Detection. In Seminars in Oncology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 224–242. [Google Scholar]
- Ghotme, K.A.G.; Barreto, G.E.; Moran, V.; Gonzalez, J.; Bustos, R.H.; Sanchez, M.; Leszek, J.; Yarla, N.S.; Gomez, R.M.; Tarasov, V.V.; et al. Gliomas: New Perspectives in Diagnosis, Treatment and Prognosis. Curr. Top. Med. Chem. 2017, 17, 1438–1447. [Google Scholar] [CrossRef] [PubMed]
- Perge, P.; Butz, H.; Pezzani, R.; Bancos, I.; Nagy, Z.; Pálóczi, K.; Nyírő, G.; Decmann, Á.; Pap, E.; Luconi, M.; et al. Evaluation and diagnostic potential of circulating extracellular vesicle-associated microRNAs in adrenocortical tumors. Sci. Rep. 2017, 7, 5474. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Liu, X.; Zhang, Y.; Li, J.; Chen, S.; Zheng, H.; Reng, R.; Zhang, C.; Chen, J.; Chen, L. Circulating microRNA124-3p, microRNA9-3p and microRNA196b-5p may be potential signatures for differential diagnosis of thyroid nodules. Oncotarget 2016, 7, 84165–84177. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Song, Q.; Li, H.; Lou, Y.; Wang, L. Circulating miR-25-3p and miR-451a may be potential biomarkers for the diagnosis of papillary thyroid carcinoma. PLoS ONE 2015, 10, e0132403. [Google Scholar]
- Patel, D.; Boufraqech, M.; Jain, M.; Zhang, L.; He, M.; Gesuwan, K.; Gulati, N.; Nilubol, N.; Fojo, T.; Kebebew, E. MiR-34a and miR-483-5p are candidate serum biomarkers for adrenocortical tumors. Surgery 2013, 154, 1224–1229. [Google Scholar] [CrossRef] [Green Version]
- Decmann, A.; Bancos, I.; Khanna, A.; Thomas, M.A.; Turai, P.; Perge, P.; Pintér, J.Z.; Toth, M.; Patócs, A.; Igaz, P. Comparison of plasma and urinary microRNA-483-5p for the diagnosis of adrenocortical malignancy. J. Biotechnol. 2019, 297, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Wang, H.; Yao, X.; Zhang, D.; Xie, Y.; Cui, R.; Zhang, X. Circulating MicroRNAs in Cancer: Potential and Challenge. Front. Genet. 2019, 10, 626. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G. Circulating miRNAs: Roles in cancer diagnosis, prognosis and therapy. Adv. Drug Deliv. Rev. 2015, 81, 75–93. [Google Scholar] [CrossRef]
- Dallapiazza, R.F.; Grober, Y.; Starke, R.M.; Laws, E.R., Jr.; Jane, J.A., Jr. Long-term results of endonasal endoscopic transsphenoidal resection of nonfunctioning pituitary macroadenomas. Neurosurgery 2015, 76, 42–52. [Google Scholar] [CrossRef]
- Koutourousiou, M.; Paluzzi, A.; Tormenti, M.; Pinheiro-Neto, C.; Fernandez-Miranda, J.; Wang, E.; Snyderman, C.; Gardner, P. Giant Pituitary Adenomas: Advantages and Limitations of Endoscopic Endonasal Surgery. J. Neurol. Surg. Part B Skull Base 2012, 73, A119. [Google Scholar] [CrossRef]
- Raverot, G.; Burman, P.; McCormack, A.; Heaney, A.; Petersenn, S.; Popovic, V.; Trouillas, J.; Dekkers, O.M.; The European Society of Endocrinology. European Society of Endocrinology Clinical Practice Guidelines for the management of aggressive pituitary tumours and carcinomas. Eur. J. Endocrinol. 2018, 178, G1–G24. [Google Scholar] [CrossRef] [PubMed]
- Di Ieva, A.; Rotondo, F.; Syro, L.V.; Cusimano, M.D.; Kovacs, K. Aggressive pituitary adenomas—Diagnosis and emerging treatments. Nat. Rev. Endocrinol. 2014, 10, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Ríos, G.G.A.; Portocarrero-Ortiz, L.; Rojas-Guerrero, C.; Terrones-Lozano, A.; Ortíz-Plata, A.; Reza-Albarrán, A.A. Nonfunctioning Pituitary Adenoma That Changed to a Functional Gonadotropinoma. Case Rep. Endocrinol. 2018, 2018, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Németh, K.; Darvasi, O.; Likó, I.; Szücs, N.; Czirják, S.; Reiniger, L.; Szabó, B.; Krokker, L.; Pállinger, É.; Igaz, P.; et al. Comprehensive Analysis of Circulating miRNAs in the Plasma of Patients With Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2019, 104, 4151–4168. [Google Scholar] [CrossRef] [PubMed]
- Das, A.V.; Pillai, R.M. Implications of miR cluster 143/145 as universal anti-oncomiRs and their dysregulation during tumorigenesis. Cancer Cell Int. 2015, 15, 92. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Li, Z.; Zhu, J.; Wang, P.; Fan, G.; Dai, Y.; Zheng, Z.; Liu, Y. Serum expression levels of microRNA-382-3p,-598-3p,-1246 and-184 in breast cancer patients. Oncol. Lett. 2016, 12, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Dai, G.; Yu, L.; Hu, Q.; Chen, J.; Guo, W. miR-143-3p inhibits the proliferation, migration and invasion in osteosarcoma by targeting FOSL2. Sci. Rep. 2018, 8, 606. [Google Scholar] [CrossRef] [Green Version]
- Amaral, F.C.; Saggioro, F.; Neder, L.; De Castro, M.; Torres, N.; Machado, H.R.; Jr, W.A.S.; Moreira, A.C. MicroRNAs Differentially Expressed in ACTH-Secreting Pituitary Tumors. J. Clin. Endocrinol. Metab. 2009, 94, 320–323. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ma, D.; Liu, H.; Wang, J.; Fan, J.; Li, X. MicroRNA-143 shows tumor suppressive effects through inhibition of oncogenic K-Ras in pituitary tumor. Int. J. Clin. Exp. Pathol. 2017, 10, 10969–10978. [Google Scholar]
- Zhao, S.; Li, J.; Feng, J.; Li, Z.; Liu, Q.; Lv, P.; Wang, F.; Hua, G.; Zhang, Y. Identification of Serum miRNA-423-5p Expression Signature in Somatotroph Adenomas. Int. J. Endocrinol. 2019, 8516858-12. [Google Scholar] [CrossRef] [Green Version]
- Kelly, B.; Haverstick, D.; Lee, J.; Thorner, M.; Vance, M.; Xin, W.; Bruns, D. İnsan büyüme hormonu yönetim hastalara bir biyomarker olarak mikroRNA dolaşan. Uyuşturucu Testi Anal. 2014, 6, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Lutsenko, A.S.; Belaya, Z.E.; Przhiyalkovskaya, E.G.; Nikitin, A.G.; Koshkin, P.A.; Lapshina, A.M.; Khandaeva, P.; Melnichenko, G.A. Expression of plasma microRNA in patients with acromegaly. Probl. Endocrinol. 2019, 65, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, I.; Nahmias, N.; Persky, M.N.; Greenfield, C.; Goldman-Wohl, D.; Hurwitz, A.; Haimov-Kochman, R.; Yagel, S.; Imbar, T. Elevated circulating micro-ribonucleic acid (miRNA)-200b and miRNA-429 levels in anovulatory women. Fertil. Steril. 2017, 107, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-X.; Li, Q.; Wang, C.; Su, Z.-P.; Li, W.-Q.; Chen, X.; Wu, Z.-B. [Differential expression analysis of prolactinoma-related microRNAs]. Zhonghua Yi Xue Za Zhi 2012, 92, 320–323. [Google Scholar]
- Wang, Y.; Yin, X.; Zhao, L.; Li, S.; Duan, J.; Kuang, R.; Duan, J. MicroRNA-200b inhibits pituitary tumor cell proliferation and invasion by targeting PKCα. Exp. Ther. Med. 2017, 14, 1706–1714. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Lu, X.; Gong, L.; Jin, F. TCTP promotes epithelial–mesenchymal transition in lung adenocarcinoma. OncoTargets Ther. 2019, 12, 1641–1653. [Google Scholar] [CrossRef] [Green Version]
- Krützfeldt, J. Strategies to use microRNAs as therapeutic targets. Best Pr. Res. Clin. Endocrinol. Metab. 2016, 30, 551–561. [Google Scholar] [CrossRef]
- Biswas, S. MicroRNAs as Therapeutic Agents: The Future of the Battle Against Cancer. Curr. Top. Med. Chem. 2019, 18, 2544–2554. [Google Scholar] [CrossRef]
- Jászai, J.; Schmidt, M.H. Trends and Challenges in Tumor Anti-Angiogenic Therapies. Cells 2019, 8, 1102. [Google Scholar] [CrossRef] [Green Version]
- Corlan, A.S.; Cimpean, A.M.; Melnic, E.; Raica, M.; Sarb, S. VEGF, VEGF165b and EG-VEGF expression is specifically related with hormone profile in pituitary adenomas. Eur. J. Histochem. 2019, 63, 1. [Google Scholar] [CrossRef] [Green Version]
- Nör, J.E.; Christensen, J.; Mooney, D.J.; Polverini, P.J. Vascular Endothelial Growth Factor (VEGF)-Mediated Angiogenesis Is Associated with Enhanced Endothelial Cell Survival and Induction of Bcl-2 Expression. Am. J. Pathol. 1999, 154, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Oh, M.-H.; Schwarz, E.M.; LaRue, C.T.; Sivaguru, M.; Imai, B.S.; Yau, P.M.; Ort, D.R.; Huber, S.C. Lysine Acetylation Is a Widespread Protein Modification for Diverse Proteins in Arabidopsis. Plant. Physiol. 2011, 155, 1769–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, S.; Majeti, B.K.; Acevedo, L.M.; Murphy, E.A.; Mukthavaram, R.; Scheppke, L.; Huang, M.; Shields, D.J.; Lindquist, J.N.; Lapinski, P.E.; et al. MicroRNA-132–mediated loss of p120RasGAP activates the endothelium to facilitate pathological angiogenesis. Nat. Med. 2010, 16, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-Q.; Song, W.-J.; Sun, T.-M.; Zhang, P.-Z.; Wang, J. Targeted Delivery of Antisense Inhibitor of miRNA for Antiangiogenesis Therapy Using cRGD-Functionalized Nanoparticles. Mol. Pharm. 2010, 8, 250–259. [Google Scholar] [CrossRef]
- Ben-Shlomo, A.; Cooper, O. The role of tyrosine kinase inhibitors in the treatment of pituitary tumors: From bench to bedside. Curr. Opin. Endocrinol. Diabetes Obes. 2017, 24, 301. [Google Scholar] [CrossRef]
- Lee, F.Y.; Vallim, T.Q.D.A.; Chong, H.K.; Zhang, Y.; Liu, Y.; Jones, S.A.; Osborne, T.F.; Edwards, P.A. Activation of the farnesoid X receptor provides protection against acetaminophen-induced hepatic toxicity. Mol. Endocrinol. 2010, 24, 1626–1636. [Google Scholar] [CrossRef] [Green Version]
- Raeder, J.; Larson, D.; LiiD, W.; Kepko, E.L.; Fuller-Rowell, T. OpenGGCM Simulations for the THEMIS Mission. Space Sci. Rev. 2008, 141, 535–555. [Google Scholar] [CrossRef]
- Switon, K.; Kotulska, K.; Janusz-Kaminska, A.; Zmorzynska, J.; Jaworski, J. Molecular neurobiology of mTOR. Neuroscience 2017, 341, 112–153. [Google Scholar] [CrossRef] [Green Version]
- Laplante, M.; Sabatini, D.M. mTOR Signaling in Growth Control and Disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [Green Version]
- Monsalves, E.; Juraschka, K.; Tateno, T.; Agnihotri, S.; Asa, S.L.; Ezzat, S.; Zadeh, G. The PI3K/AKT/mTOR pathway in the pathophysiology and treatment of pituitary adenomas. Endocr. Relat. Cancer 2014, 21, R331–R344. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Fan, Y.-D.; Wu, P.-F.; Duysenbi, S.; Feng, Z.-H.; Du, G.-J.; Zhang, T.-R. MicroRNA-145 inhibits the activation of the mTOR signaling pathway to suppress the proliferation and invasion of invasive pituitary adenoma cells by targeting AKT3 in vivo and in vitro. OncoTargets Ther. 2017, 10, 1625–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.B.; Li, W.Q.; Lin, S.J.; De Wang, C.; Cai, L.; Lu, J.L.; Chen, Y.X.; Su, Z.P.; Shang, H.B.; Yang, W.L.; et al. MicroRNA expression profile of bromocriptine-resistant prolactinomas. Mol. Cell. Endocrinol. 2014, 395, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Vickers, N.J. Animal Communication: When I’m Calling You, Will You Answer Too? Curr. Boil. 2017, 27, R713–R715. [Google Scholar] [CrossRef] [PubMed]
- Jian, M.; Du, Q.; Zhu, D.; Mao, Z.; Wang, X.; Feng, Y.; Xiao, Z.; Wang, H.; Zhu, Y. Tumor suppressor miR-145-5p sensitizes prolactinoma to bromocriptine by downregulating TPT1. J. Endocrinol. Investig. 2018, 42, 639–652. [Google Scholar] [CrossRef]
- Taboada, G.F.; Luque, R.M.; Bastos, W.; Guimaraes, R.F.; Marcondes, J.B.; Chimelli, L.M.; Fontes, R.; Mata, P.J.; Niemeyer Filho, P.; Carvalho, D.P. Quantitative analysis of somatostatin receptor subtype (SSTR1–5) gene expression levels in somatotropinomas and non-functioning pituitary adenomas. Eur. J. Endocrinol. 2007, 156, 65–74. [Google Scholar] [CrossRef]
- Saveanu, A.; Gunz, G.; Dufour, H.; Caron, P.; Fina, F.; Ouafik, L.; Culler, M.; Moreau, J.; Enjalbert, A.; Jaquet, P. Bim-23244, a somatostatin receptor subtype 2-and 5-selective analog with enhanced efficacy in suppressing growth hormone (GH) from octreotide-resistant human GH-secreting adenomas. J. Clin. Endocrinol. Metab. 2001, 86, 140–145. [Google Scholar]
- Mao, Z.-G.; He, D.-S.; Zhou, J.; Yao, B.; Xiao, W.-W.; Chen, C.-H.; Zhu, Y.-H.; Wang, H.-J. Differential expression of microRNAs in GH-secreting pituitary adenomas. Diagn. Pathol. 2010, 5, 79. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Mao, Z.; He, D.; Liao, C.; Jiang, X.; Lei, N.; Hu, B.; Wang, X.; Li, Z.; Lin, Y.; et al. Expression of somatostatin receptor subtype 2 in growth hormone-secreting pituitary adenoma and the regulation of miR-185. J. Endocrinol. Investig. 2015, 38, 1117–1128. [Google Scholar] [CrossRef]
- Butz, H.; Patócs, A. MicroRNAs in endocrine tumors. Ejifcc 2019, 30, 146–164. [Google Scholar]
- Ludwig, N.; Whiteside, T.L.; Reichert, T.E. Challenges in Exosome Isolation and Analysis in Health and Disease. Int. J. Mol. Sci. 2019, 20, 4684. [Google Scholar] [CrossRef] [Green Version]
- Takov, K.; Yellon, D.M.; Davidson, S.M. Comparison of small extracellular vesicles isolated from plasma by ultracentrifugation or size-exclusion chromatography: Yield, purity and functional potential. J. Extracell. Vesicles 2018, 8, 1560809. [Google Scholar] [CrossRef]
- Mincheva-Nilsson, L.; Baranov, V.; Nagaeva, O.; Dehlin, E. Isolation and Characterization of Exosomes from Cultures of Tissue Explants and Cell Lines. Curr. Protoc. Immunol. 2016, 115, 14.42.1–14.42.21. [Google Scholar] [CrossRef]


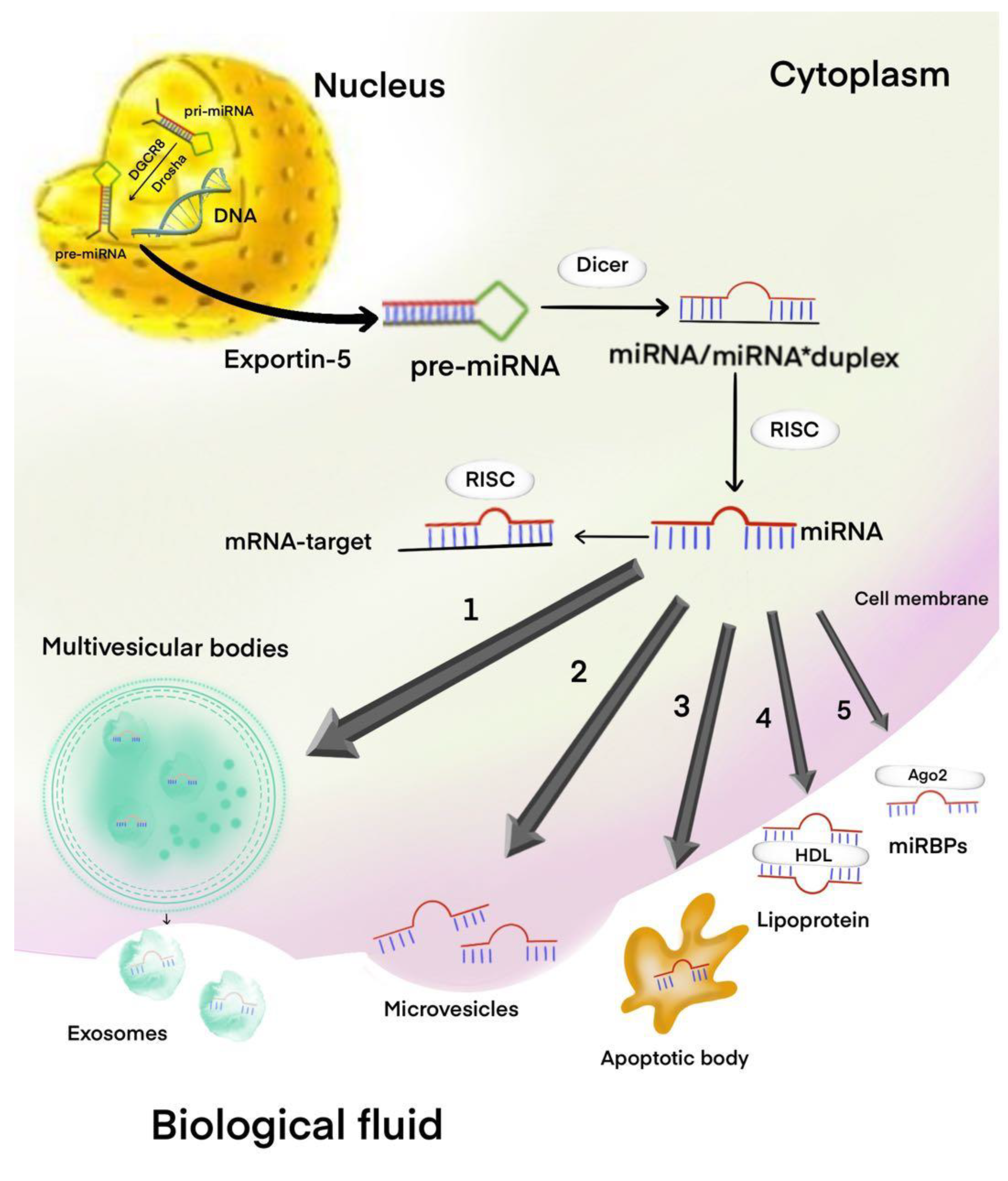{kind=link}
| miRNA | Gene Target | Type of PA | Biological Function | Regulation | Phenotype | Ref. |
|---|---|---|---|---|---|---|
| miR-524-5p | PTTG1/PBF | NFA | Inhibition of tumor cell proliferation, migration, and invasion | Down | Tumor suppressor | [19] |
| miR-424, miR-503 | CDC25A | NFA | Inhibition of tumor cell growth | Down | Tumor suppressor | [20] |
| miR-34 | AIP | GH | Induces invasive properties in tumor cells | Up | OncomiR | [21] |
| miR-410 | CCNB1 | FSH /LH | Increased expression of cyclin A and D protein affecting the G1-S phase of the cell cycle. Inhibition of tumor cell proliferation | Down | Tumor suppressor | [22] |
| miR-26a | PLAG1 | NFA, GH, ACTH, PRL | Induces invasive properties in tumor cells | Up | OncomiR | [23] |
| miR-23b | HMGA2 | NFA, FSH /LH, GH | Inhibition of tumor cell proliferation, delaying cell division in the G1 phase of the cell cycle | Down | Tumor suppressor | [24] |
| miR-130b | CCNA2 | NFA, FSH /LH, GH | Inhibition of tumor cell proliferation, delaying cell division in the G2 phase of the cell cycle | Down | Tumor suppressor | [24] |
| miR-106b | PTEN | Invasive (NFA, GH, ACTH, PRL) | Induces invasive properties in tumor cells | Up | OncomiR | [25] |
| miR-21 | PITX2 | Invasive PA | Suppresses PA tumor cell proliferation and facilitates their apoptosis | Down | Tumor suppressor | [26] |
| miR-132, miR-15a, miR-16 | SOX5 | Invasive (NFA, GH, ACTH, PRL) | Inhibition of tumor cell proliferation, migration, and invasion | Down | Tumor suppressor | [27] |
| miR-200c | PTEN | PRL | Tumor cell apoptosis reduction | Up | OncomiR | [28] |
| miR-329, miR-300, miR-381, miR-655 | PTTG1 | PRL, GH | Inhibition of tumor cell growth | Down | Tumor suppressor | [29] |
| miR-20a, miR-17-5p | PTEN, TIMP2 | NFA | Carcinoma metastasis | Up | OncomiR | [27] |
| miR-106b | PTEN-PI3K/AKT | NFA | Activation of invasive properties and migration of tumor cells; carcinoma metastasis | Up | OncomiR | [27,30] |
| miR-183 | KIAA0101 | PRL | Inhibition of tumor cell proliferation | Down | Tumor suppressor | [31] |
| miR-15, miR-16, miR-26a, miR-196a2 and Let-7a | HMGA1 and HMGA2 | GH, PRL | Inhibition of tumor cell proliferation | Up | Tumor suppressor | [32] |
| miRNA. | Tumor Type | Sample | Comparison | Sensitivity % | Specificity % | AUC | Regulation | Ref. |
|---|---|---|---|---|---|---|---|---|
| Exosomal miR-101 and miR-483-5p | ACA and ACC | Plasma | ACC vs. ACA | 68.75 and 87.5 | 83.33 and 94.44 | 0.766 and 0.965 | Up | [35] |
| miR-124-3p, miR-9-3p and miR-196b-5p | PTC and benign thyroid nodules | Plasma | PTC vs. benign lesion/PTC vs. healthy control | 88, 80, and 74 | 78.8, 73.7, and 66 | 0.859, 0.823, and 0.781 | Up | [36] |
| miR-25-3p and miR-451a | PTC and benign thyroid nodules | Plasma | PTC vs. benign lesion | 92.8 and 88.9 | 68.8 and 66.7 | 0.835 and 0.857 | Up | [37] |
| miR-34a and miR-483-5p | ACA and ACC | Serum | ACC vs. ACA | / | / | 0.81 and 0.74 | Up | [38] |
| miR-483-5p | ACA and ACC | Plasma | ACC vs. ACA | 87 | 78.3 | 0.88 | Up | [39] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beylerli, O.; Beeraka, N.M.; Gareev, I.; Pavlov, V.; Yang, G.; Liang, Y.; Aliev, G. MiRNAs as Noninvasive Biomarkers and Therapeutic Agents of Pituitary Adenomas. Int. J. Mol. Sci. 2020, 21, 7287. https://doi.org/10.3390/ijms21197287
Beylerli O, Beeraka NM, Gareev I, Pavlov V, Yang G, Liang Y, Aliev G. MiRNAs as Noninvasive Biomarkers and Therapeutic Agents of Pituitary Adenomas. International Journal of Molecular Sciences. 2020; 21(19):7287. https://doi.org/10.3390/ijms21197287
Chicago/Turabian StyleBeylerli, Ozal, Narasimha M. Beeraka, Ilgiz Gareev, Valentin Pavlov, Guang Yang, Yanchao Liang, and Gjumrakch Aliev. 2020. "MiRNAs as Noninvasive Biomarkers and Therapeutic Agents of Pituitary Adenomas" International Journal of Molecular Sciences 21, no. 19: 7287. https://doi.org/10.3390/ijms21197287
APA StyleBeylerli, O., Beeraka, N. M., Gareev, I., Pavlov, V., Yang, G., Liang, Y., & Aliev, G. (2020). MiRNAs as Noninvasive Biomarkers and Therapeutic Agents of Pituitary Adenomas. International Journal of Molecular Sciences, 21(19), 7287. https://doi.org/10.3390/ijms21197287