Identification of the Cytosolic Glucose-6-Phosphate Dehydrogenase Gene from Strawberry Involved in Cold Stress Response

 , , ,
, , ,  ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cloning and Characterization of G6PDH Genes in Strawberry
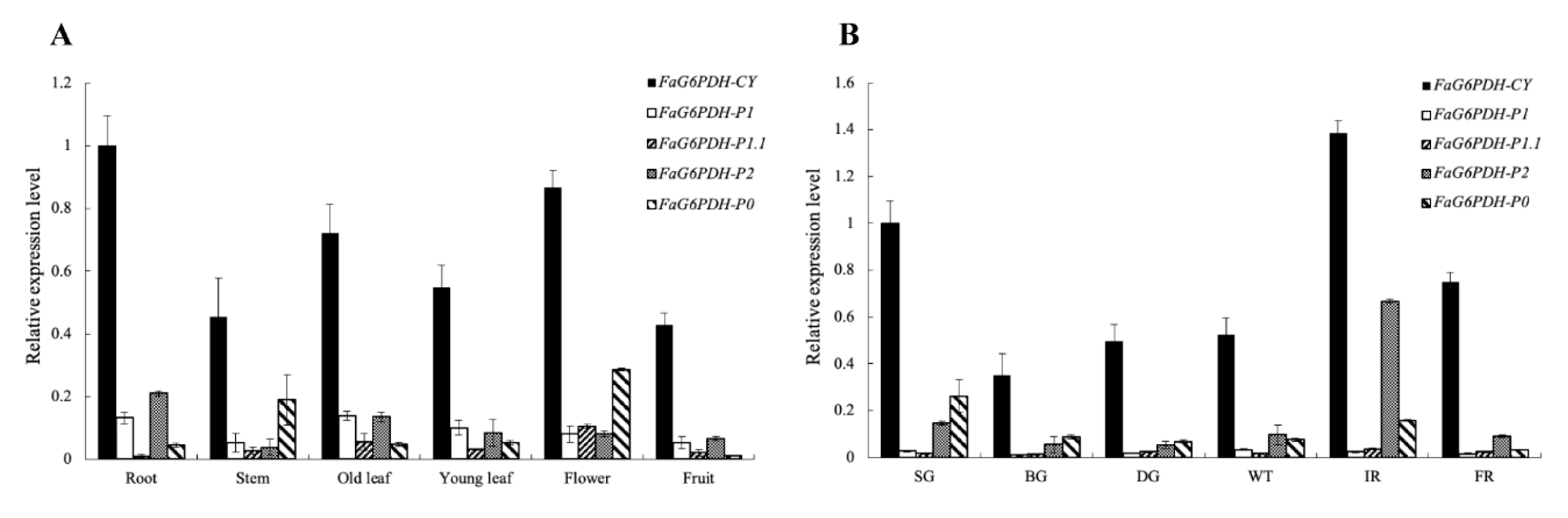
2.2. Expression Profile of FaG6PDHs in Different Tissues and during the Fruit Development
2.3. Effect of FaG6PDH Enzyme Activity on Related Physiological Indexes in Response to Cold Stress
2.4. Identification of FaG6PDH Genes Related to Cold Stress
2.5. Isolation and Cold Stress-Induced Activity Analysis of FaG6PDH-CY Promoter
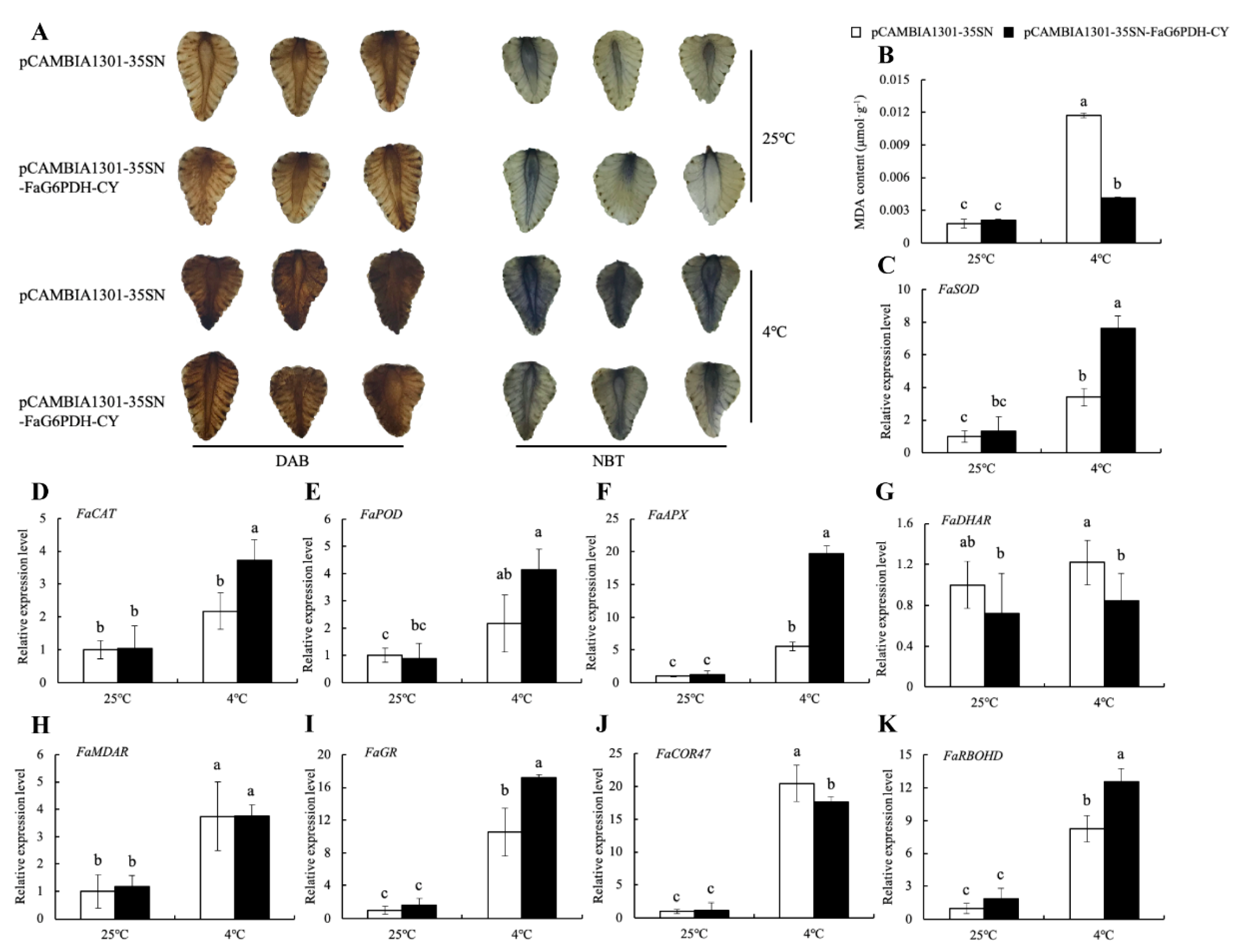
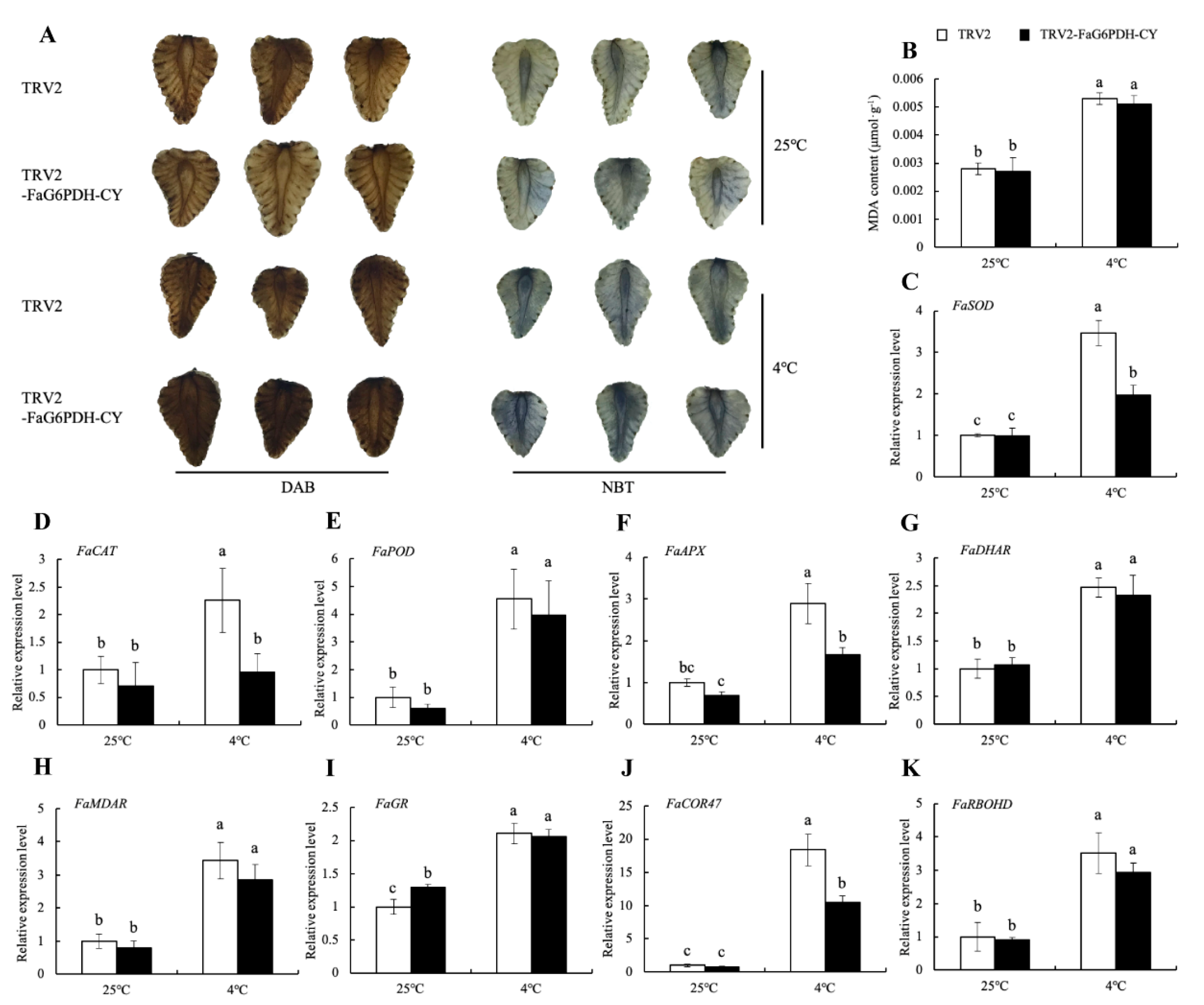
2.6. Effect of FaG6PDH-CY Transient Expression on Strawberries in Response to Cold Stress
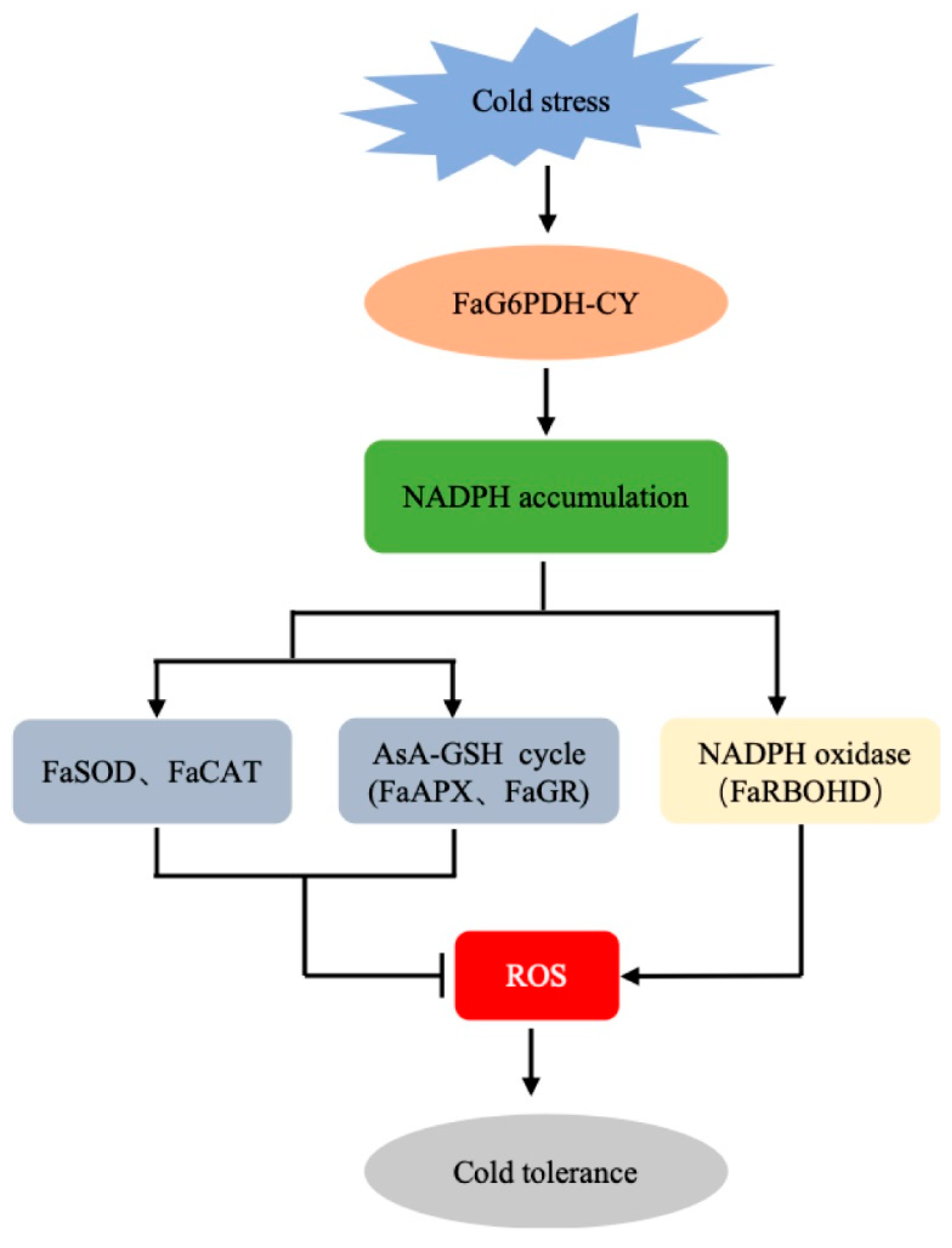
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Determination of G6PDH, SOD, CAT and POD Activities
4.3. Electrolyte Leakage and MDA Content Assay
4.4. Proline, Soluble Sugar and Soluble Protein Content Determination
4.5. RNA Extraction, First-Strand cDNA Synthesis and Genomic DNA Isolation
4.6. Cloning and Bioinformatic Analysis of G6PDH Genes
4.7. Quantitative Real-Time PCR
4.8. Vector Construction
4.9. Transient Expression and Cold-Resistance Assay in Tobacco Leaves and Strawberry Fruits
4.10. Histochemical Staining and Activity Assays for GUS
4.11. DAB and NBT Staining
4.12. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kruger, N.J.; Von Schaewen, A. The oxidative pentose phosphate pathway: Structure and organisation. Curr. Opin. Plant Boil. 2003, 6, 236–246. [Google Scholar] [CrossRef]
- Schnarrenberger, C.; Oeser, A.; Tolbert, N. Two isoenzymes each of glucose-6-phosphate dehydrogenase and 6-phosphogluconate dehydrogenase in spinach leaves. Arch. Biochem. Biophys. 1973, 154, 438–448. [Google Scholar] [CrossRef]
- Von Schaewen, A.; Langenkämper, G.; Graeve, K.; Wenderoth, I.; Scheibe, R. Molecular characterization of the plastidic glucose-6-phosphate dehydrogenase from potato in comparison to its cytosolic counterpart. Plant Physiol. 1995, 109, 1327–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, S.; Carfagna, S.; Massaro, G.; Vona, V.; Rigano, V.D.M. Glucose-6-phosphate dehydrogenase in barley roots: Kinetic properties and localisation of the isoforms. Planta 2001, 212, 627–634. [Google Scholar] [CrossRef]
- Graeve, K.; Schaewen, A.; Scheibe, R. Purification, characterization, and cDNA sequence of glucose-6-phosphate dehydrogenase from potato (Solatium tuberosum L.). Plant J. 1994, 5, 353–361. [Google Scholar] [CrossRef]
- Castiglia, D.; Cardi, M.; Landi, S.; Cafasso, D.; Esposito, S. Expression and characterization of a cytosolic glucose 6 phosphate dehydrogenase isoform from barley (Hordeum vulgare) roots. Protein Expr. Purif. 2015, 112, 8–14. [Google Scholar] [CrossRef]
- Wendt, U.K.; Wenderoth, I.; Tegeler, A.; Von Schaewen, A. Molecular characterization of a novel glucose-6-phosphate dehydrogenase from potato (Solanum tuberosum L.). Plant J. 2000, 23, 723–733. [Google Scholar] [CrossRef]
- Esposito, S.; Massaro, G.; Vona, V.; Rigano, V.D.M.; Carfagna, S. Glutamate synthesis in barley roots: The role of the plastidic glucose-6-phosphate dehydrogenase. Planta 2003, 216, 639–647. [Google Scholar] [CrossRef]
- Wakao, S.; Benning, C. Genome-wide analysis of glucose-6-phosphate dehydrogenases in Arabidopsis. Plant J. 2004, 41, 243–256. [Google Scholar] [CrossRef]
- Meyer, T.; Hölscher, C.; Schwöppe, C.; Von Schaewen, A. Alternative targeting of Arabidopsis plastidic glucose-6-phosphate dehydrogenase G6PD1 involves cysteine-dependent interaction with G6PD4 in the cytosol. Plant J. 2011, 66, 745–758. [Google Scholar] [CrossRef]
- Landi, S.; Nurcato, R.; De Lillo, A.; Lentini, M.; Grillo, S.; Esposito, S. Glucose-6-phosphate dehydrogenase plays a central role in the response of tomato (Solanum lycopersicum) plants to short and long-term drought. Plant Physiol. Biochem. 2016, 105, 79–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.W.; Tian, H.; Yu, X.; Zheng, L.P. Glucose-6-phosphate dehydrogenase plays critical role in artemisinin production of Artemisia annua under salt stress. Boil. Plant 2017, 61, 529–539. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, S.; Guo, H.; Zhang, Z.; Chen, X. Functional analysis of PsG6PDH, a cytosolic glucose-6-phosphate dehydrogenase gene from Populus suaveolens, and its contribution to cold tolerance improvement in tobacco plants. Biotechnol. Lett. 2013, 35, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Chen, G.; Li, F.; Wang, X.; Hu, Y.; Bi, Y. Involvement of G6PDH in heat stress tolerance in the calli from Przewalskia tangutica and Nicotiana tabacum. Biol. Plant 2012, 56, 422–430. [Google Scholar] [CrossRef]
- Ślaski, J.J.; Zhang, G.; Basu, U.; Stephens, J.L.; Taylor, G.J. Aluminum resistance in wheat (Triticum aestivum) is associated with rapid, Al-induced changes in activities of glucose-6-phosphate dehydrogenase and 6-phosphogluconate dehydrogenase in root apices. Physiol. Plant 1996, 98, 477–484. [Google Scholar] [CrossRef]
- Wang, J.; Nan, Z.; Christensen, M.J.; Li, C. Glucose-6-phosphate dehydrogenase plays a vital role in Achnatherum inebrians plants host to Epichloë gansuensis by improving growth under nitrogen deficiency. Plant Soil 2018, 430, 37–48. [Google Scholar] [CrossRef]
- Wei, M.; Ge, Y.; Li, C.; Han, X.; Qin, S.; Chen, Y.; Tang, Q.; Li, J. G6PDH regulated NADPH production and reactive oxygen species metabolism to enhance disease resistance against blue mold in apple fruit by acibenzolar-S-methyl. Postharvest Boil. Technol. 2019, 148, 228–235. [Google Scholar] [CrossRef]
- Santo, S.D.; Stampfl, H.; Krasensky, J.; Kempa, S.; Gibon, Y.; Petutschnig, E.; Rozhon, W.; Heuck, A.; Clausen, T.; Jonak, C. Stress-induced GSK3 regulates the redox stress response by phosphorylating glucose-6-phosphate dehydrogenase in Arabidopsis. Plant Cell 2012, 24, 3380–3392. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ma, Y.; Huang, C.; Wan, Q.; Li, N.; Bi, Y. Glucose-6-phosphate dehydrogenase plays a central role in modulating reduced glutathione levels in reed callus under salt stress. Planta 2007, 227, 611–623. [Google Scholar] [CrossRef]
- Krüger, A.; Grüning, N.-M.; Wamelink, M.M.; Kerick, M.; Kirpy, A.; Parkhomchuk, D.; Bluemlein, K.; Schweiger, M.-R.; Soldatov, A.; Lehrach, H.; et al. The pentose phosphate pathway is a metabolic redox sensor and regulates transcription during the antioxidant response. Antioxid. Redox Signal. 2011, 15, 311–324. [Google Scholar] [CrossRef]
- Caretto, S.; Linsalata, V.; Colella, G.; Mita, G.; Lattanzio, V. Carbon fluxes between primary metabolism and phenolic pathway in plant tissues under stress. Int. J. Mol. Sci. 2015, 16, 26378–26394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, S. Nitrogen assimilation, abiotic stress and glucose 6-phosphate dehydrogenase: The full circle of reductants. Plants 2016, 5, 24. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, Y.; Sasakuma, T. Specific expression of glucose-6-phosphate dehydrogenase (G6PDH) gene by salt stress in wheat (Triticum aestivum L.). Plant Sci. 2000, 158, 53–60. [Google Scholar] [CrossRef]
- Cardi, M.; Chibani, K.; Cafasso, D.; Rouhier, N.; Jacquot, J.-P.; Esposito, S. Abscisic acid effects on activity and expression of barley (Hordeum vulgare) plastidial glucose-6-phosphate dehydrogenase. J. Exp. Bot. 2011, 62, 4013–4023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, X.; He, B.; Fang, Y.; Tang, C. Identification and characterization of the glucose-6-phosphate dehydrogenase gene family in the para rubber tree, Hevea brasiliensis. Front. Plant Sci. 2016, 7, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Cui, Y.; Huang, S.; Yu, J.; Wang, X.; Xin, D.; Li, X.; Liu, Y.; Dai, Y.; Qi, Z.; et al. Genome-wide analysis of the glucose-6-phosphate dehydrogenase family in soybean and functional identification of GmG6PDH2 involvement in salt stress. Front. Plant Sci. 2020, 11, 214. [Google Scholar] [CrossRef] [Green Version]
- Singh, S. Regulation and properties of glucose-6-phosphate dehydrogenase: A review. Int. J. Plant Physiol. Biochem. 2012, 4, 1–19. [Google Scholar]
- Esposito, S.; Guerriero, G.; Vona, V.; Rigano, V.D.M.; Carfagna, S.; Rigano, C. Glutamate synthase activities and protein changes in relation to nitrogen nutrition in barley: The dependence on different plastidic glucose-6P dehydrogenase isoforms. J. Exp. Bot. 2004, 56, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.S.; Emes, M.J.; Debnam, P.M. Isolation and characterisation of a full-length genomic clone encoding a plastidic glucose 6-phosphate dehydrogenase from Nicotiana tabacum. Planta 2001, 212, 499–507. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, J.; Wang, X.; Bi, Y. Glucose-6-phosphate dehydrogenase acts as a regulator of cell redox balance in rice suspension cells under salt stress. Plant Growth Regul. 2012, 69, 139–148. [Google Scholar] [CrossRef]
- Yang, Y.; Fu, Z.; Su, Y.; Zhang, X.; Li, G.; Guo, J.; Que, Y.; Xu, L. A cytosolic glucose-6-phosphate dehydrogenase gene, ScG6PDH, plays a positive role in response to various abiotic stresses in sugarcane. Sci. Rep. 2014, 4, 7090. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ruan, M.; Wan, Q.; He, W.; Yang, L.; Liu, X.; He, L.; Yan, L.; Bi, Y. Nitric oxide and hydrogen peroxide increase glucose-6-phosphate dehydrogenase activities and expression upon drought stress in soybean roots. Plant Cell Rep. 2019, 39, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Asai, S.; Yoshioka, M.; Nomura, H.; Tone, C.; Nakajima, K.; Nakane, E.; Doke, N.; Yoshioka, H. A plastidic glucose-6-phosphate dehydrogenase is responsible for hypersensitive response cell death and reactive oxygen species production. J. Gen. Plant Pathol. 2011, 77, 152–162. [Google Scholar] [CrossRef]
- Hernandez-Garcia, C.M.; Finer, J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Sewelam, N.; Kazan, K.; Schenk, P.M. Global plant stress signaling: Reactive oxygen species at the cross-road. Front. Plant Sci. 2016, 7, 187. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.-H.; Hou, J.; Li, Y.; Zhang, Y.; Huang, J.; Liang, W. Nitric oxide-mediated cytosolic glucose-6-phosphate dehydrogenase is involved in aluminum toxicity of soybean under high aluminum concentration. Plant Soil 2017, 416, 39–52. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Al Mahmud, J.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wang, X.; Chang, N.; Nan, W.; Wang, S.; Ruan, M.; Sun, L.; Li, S.; Bi, Y. Cytosolic glucose-6-phosphate dehydrogenase is involved in seed germination and root growth under salinity in Arabidopsis. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Rajashekar, C.; Panda, M. Water stress is a component of cold acclimation process essential for inducing full freezing tolerance in strawberry. Sci. Hortic. 2014, 174, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Sagi, M.; Fluhr, R. Production of reactive oxygen species by plant NADPH oxidases. Plant Physiol. 2006, 141, 336–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, Y.; He, Y.; Hu, W.; Zhang, Y.; Wang, X.; Tang, H.-R. Identification of NADPH oxidase family members associated with cold stress in strawberry. FEBS Open Bio 2018, 8, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q. Experimental Tutorial of Plant Physiology; Science and Technology Press: Chengdu, China, 2003; pp. 40–49. [Google Scholar]
- Chen, Q.; Yu, H.; Tang, H.-R.; Wang, X. Identification and expression analysis of genes involved in anthocyanin and proanthocyanidin biosynthesis in the fruit of blackberry. Sci. Hortic. 2012, 141, 61–68. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Tang, Y.-M.; Hu, Y.-Y.; Wang, Y.; Sun, B.; Wang, X.; Tang, H.-R.; Chen, Q. FaTT12-1, a multidrug and toxin extrusion (MATE) member involved in proanthocyanidin transport in strawberry fruits. Sci. Hortic. 2018, 231, 158–165. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Luo, M.; Cheng, L.; Lin, Y.; Chen, Q.; Sun, B.; Gu, X.; Wang, Y.; Li, M.; Luo, Y.; et al. Identification of the Cytosolic Glucose-6-Phosphate Dehydrogenase Gene from Strawberry Involved in Cold Stress Response. Int. J. Mol. Sci. 2020, 21, 7322. https://doi.org/10.3390/ijms21197322
Zhang Y, Luo M, Cheng L, Lin Y, Chen Q, Sun B, Gu X, Wang Y, Li M, Luo Y, et al. Identification of the Cytosolic Glucose-6-Phosphate Dehydrogenase Gene from Strawberry Involved in Cold Stress Response. International Journal of Molecular Sciences. 2020; 21(19):7322. https://doi.org/10.3390/ijms21197322
Chicago/Turabian StyleZhang, Yunting, Mengwen Luo, Lijuan Cheng, Yuanxiu Lin, Qing Chen, Bo Sun, Xianjie Gu, Yan Wang, Mengyao Li, Ya Luo, and et al. 2020. "Identification of the Cytosolic Glucose-6-Phosphate Dehydrogenase Gene from Strawberry Involved in Cold Stress Response" International Journal of Molecular Sciences 21, no. 19: 7322. https://doi.org/10.3390/ijms21197322
APA StyleZhang, Y., Luo, M., Cheng, L., Lin, Y., Chen, Q., Sun, B., Gu, X., Wang, Y., Li, M., Luo, Y., Wang, X., Zhang, Y., & Tang, H. (2020). Identification of the Cytosolic Glucose-6-Phosphate Dehydrogenase Gene from Strawberry Involved in Cold Stress Response. International Journal of Molecular Sciences, 21(19), 7322. https://doi.org/10.3390/ijms21197322