Production of Antioxidant Molecules in Polygonum aviculare (L.) and Senecio vulgaris (L.) under Metal Stress: A Possible Tool in the Evaluation of Plant Metal Tolerance

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
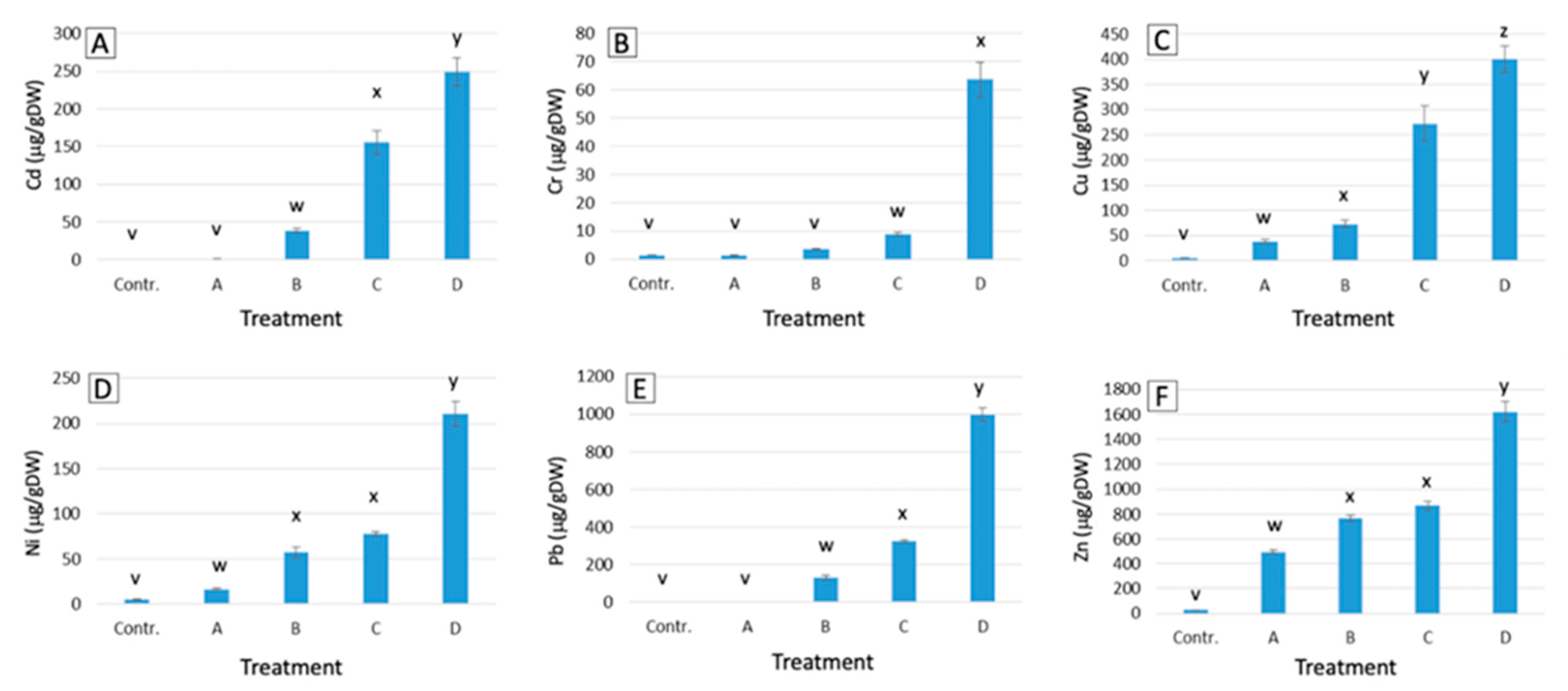
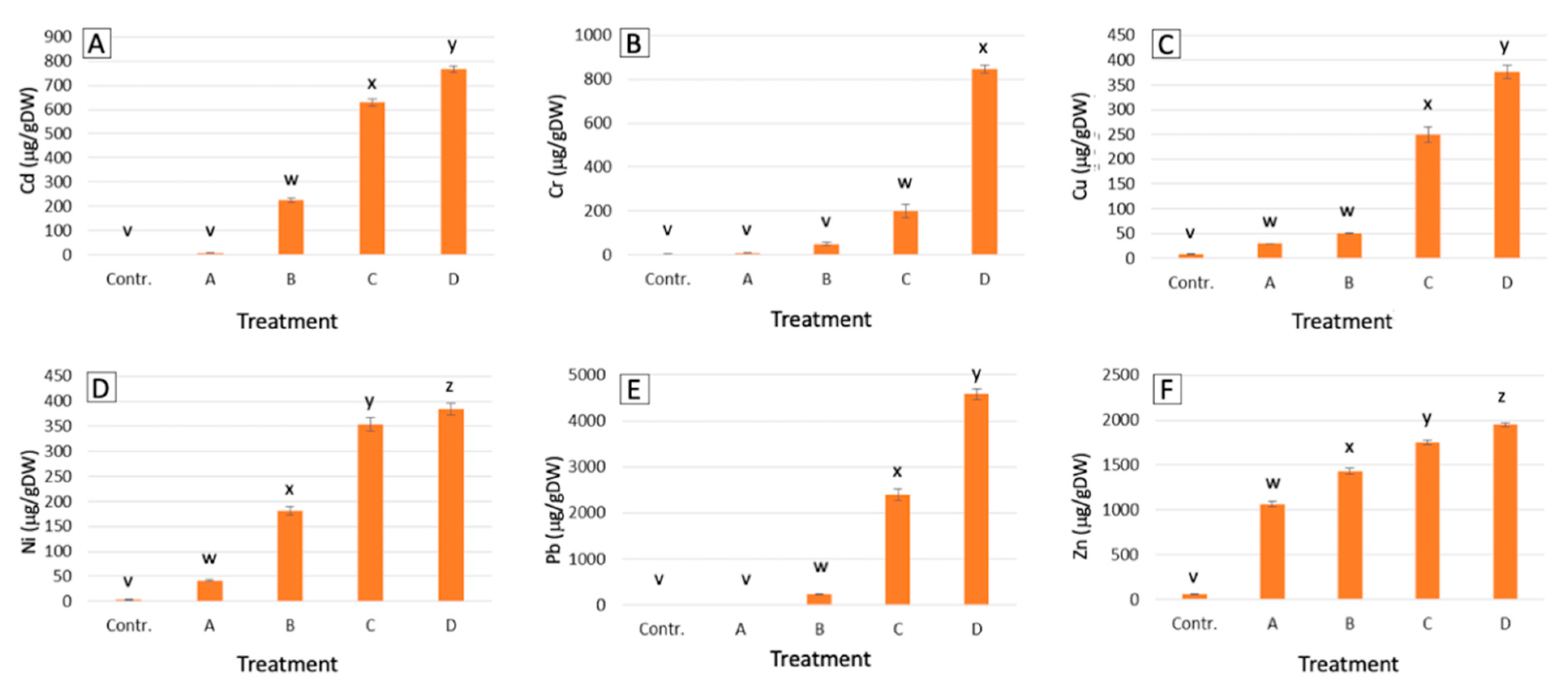
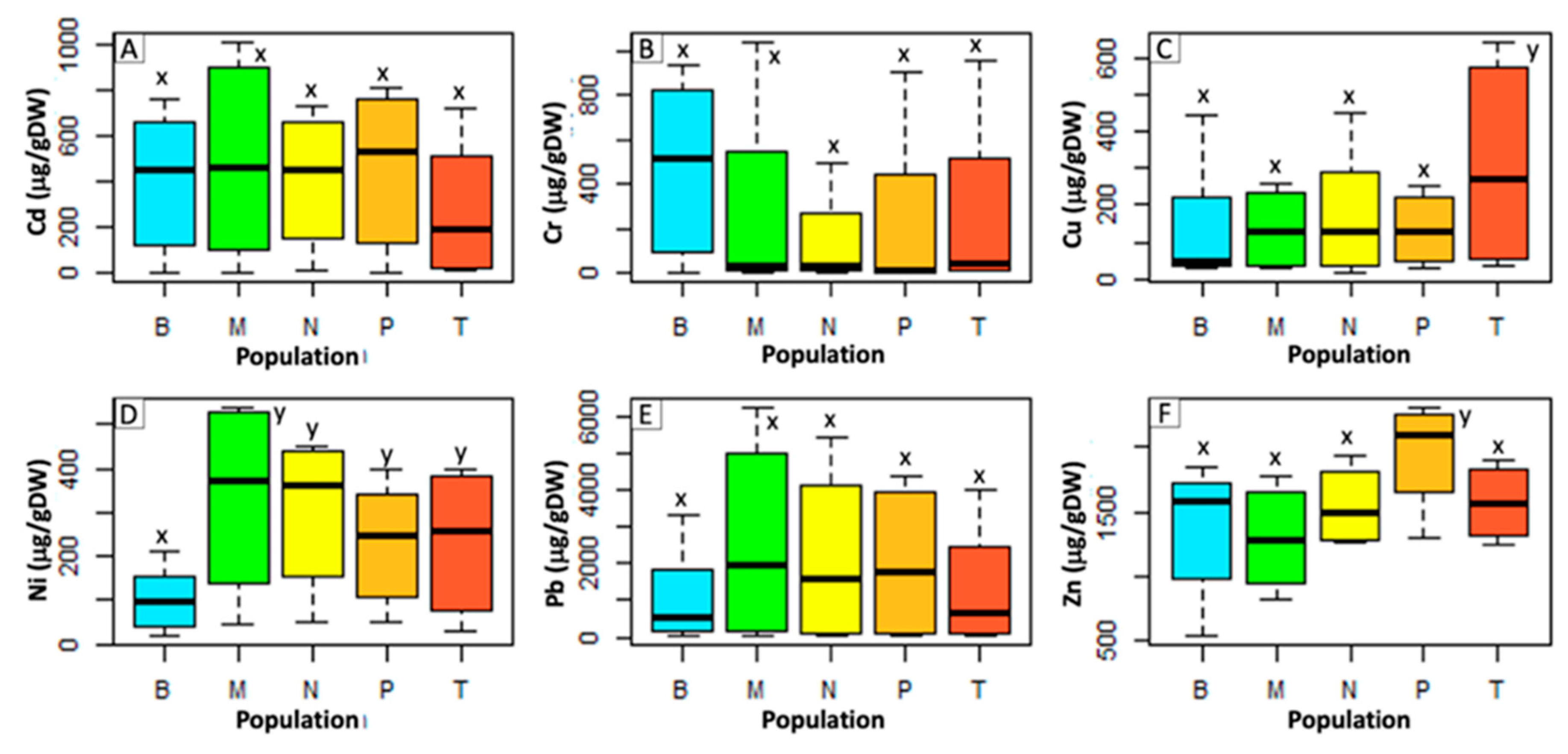
2.1. Metal Accumulation
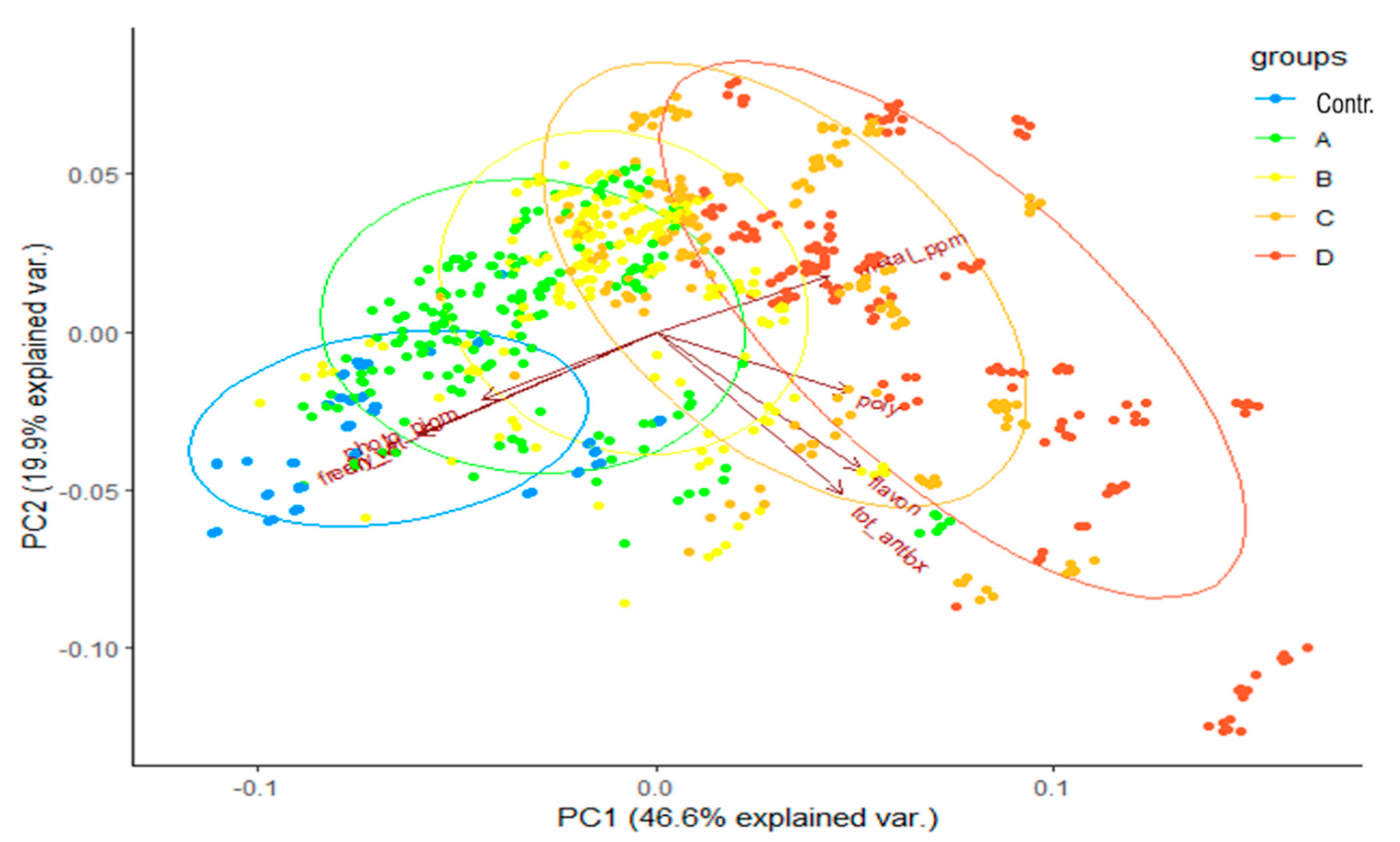
2.2. Effects of Metals on the Production of Antioxidant Metabolites and Photosynthetic Pigments
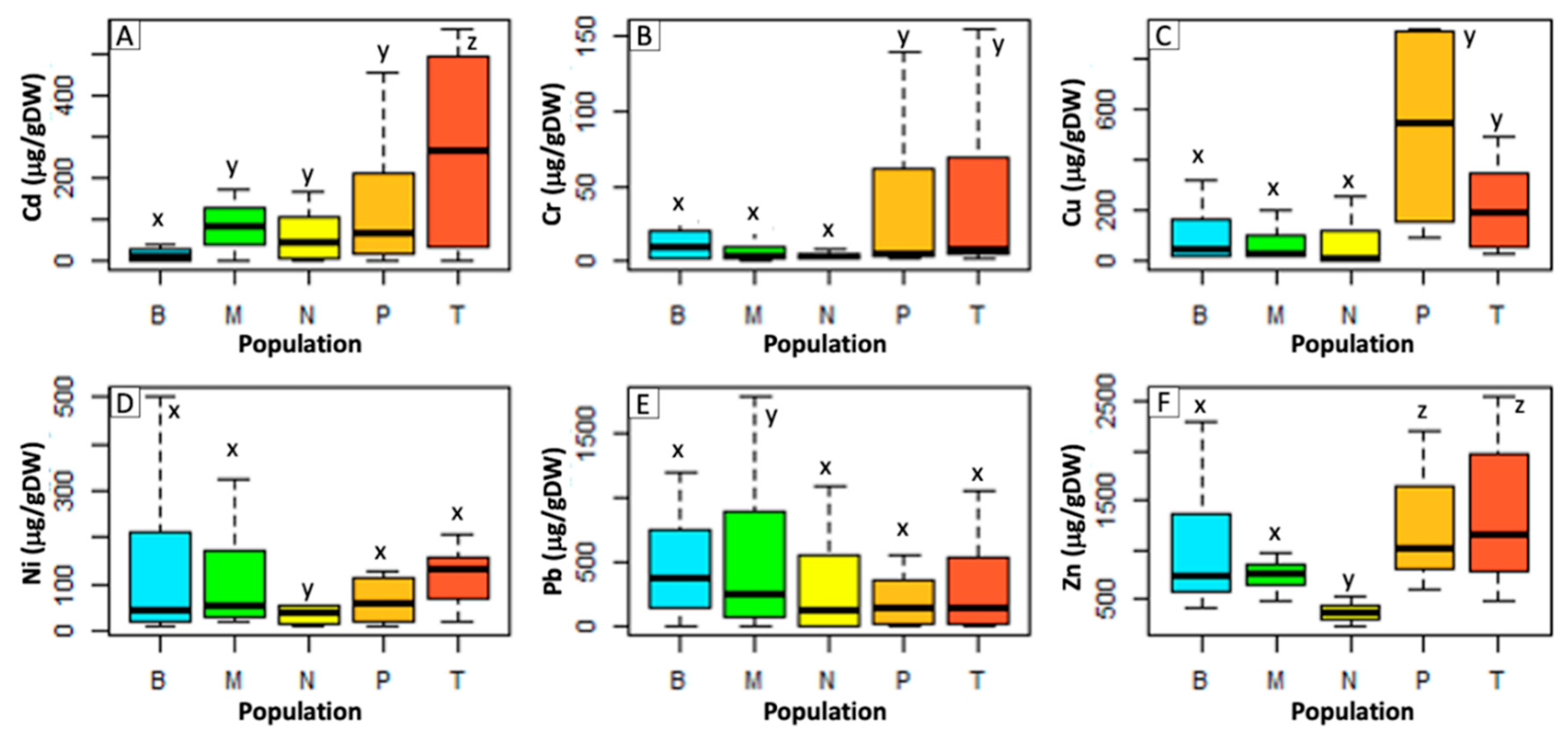
2.3. Population Differences
3. Discussion
4. Materials and Methods
4.1. Species Selection
4.2. Plant Cultivation
4.3. Sample Collection and Preparation
4.4. Heavy Metal Quantification
4.5. Spectrophotometric Analysis
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | ascorbic acid |
| ABTS | 2,2’-azino-bis(3-etilbenzotiazolin-6-sulfonic) acid |
| CAT | catechin |
| GA | gallic acid |
| HM | heavy metal |
| ROS | reactive oxygen species |
References
- Etesami, H. Bacterial mediated alleviation of heavy metal stress and decreased accumulation of metals in plant tissues: Mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 147, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Rout, G.R.; Das, P. Effect of metal toxicity on plant growth and metabolism: I. Zinc. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 873–884. [Google Scholar]
- Baker, A.J.M. Accumulators and excluders: Strategies in the response of plants to heavy metals. J. Plant. Nutr. 1981, 3, 643–665. [Google Scholar] [CrossRef]
- Viehweger, K. How plants cope with heavy metals. Bot. Stud. 2014, 55, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef]
- Patra, M.; Bhowmik, N.; Bandopadhyay, B.; Sharma, A. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ. Exp. Bot. 2004, 52, 199–223. [Google Scholar] [CrossRef]
- Dalvi, A.A.; Bhalerao, S.A. Response of plants towards heavy metal toxicity: An overview of avoidance, tolerance and uptake mechanism. Ann. Plant Sci. 2013, 2, 362–368. [Google Scholar]
- John, R.; Ahmad, P.; Gadgil, K.S.; Sharma, S. Heavy metal toxicity: Effect on plant growth, biochemical parameters and metal accumulation by Brassica juncea L. Int. J. Plant Prod. 2009, 3, 65–76. [Google Scholar] [CrossRef]
- Ovecka, M.; Takac, T. Managing heavy metal toxicity stress in plants: Biological and biotechnological tools. Biotechnol. Adv. 2014, 32, 73–86. [Google Scholar] [CrossRef]
- Kisa, D.; Elmastas, M.; Ozturk, L.; Kayir, O. Responses of the phenolic compounds of Zea mays under heavy metal stress. Appl. Biol. Chem. 2016, 59, 813–820. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef]
- Yadav, S.K. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef] [Green Version]
- González, A.; Gil-Díaz, M.; Pinilla, P.; Lobo, M. Impact of Cr and Zn on growth, biochemical and physiological parameters, and metal accumulation by wheat and barley plants. Water Air Soil Pollut. 2017, 228, 419. [Google Scholar] [CrossRef]
- Kisa, D. The responses of antioxidant system against the heavy metal-induced stress in tomato. J. Nat. Appl. Sci. 2018, 22, 1–6. [Google Scholar] [CrossRef]
- Pourrut, B.; Shahid, M.; Douay, F.; Dumat, C.; Pinelli, E. Molecular mechanisms involved in lead uptake, toxicity and detoxification in higher plants. In Heavy Metal Stress in Plants; Gupta, D., Corpas, F., Palma, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 121–147. [Google Scholar]
- Manara, A. Plant responses to heavy metal toxicity. In Plants and Heavy Metals; Furini, A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 27–53. [Google Scholar]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Keilig, K.; Ludwig-Müller, J. Effect of flavonoids on heavy metal tolerance in Arabidopsis thaliana seedlings. Bot. Stud. 2009, 50, 311–318. [Google Scholar]
- Pękal, A.; Biesaga, M.; Pyrzynska, K. Interaction of quercetin with copper ions: Complexation, oxidation and reactivity towards radicals. Biometals 2011, 24, 41–49. [Google Scholar] [CrossRef]
- Geipel, G.; Drewitz, S.; Viehweger, K. Detection of uranium (IV) by LIPAS in biologic relevant samples. In Annual Report 2010, Institute of Radiochemistry; Bernhard, G., Foerstendorf, H., Richter, A., Viehweger, K., Eds.; Helmholtz-Zentrum Dresden-Rossendorf e.V.: Dresden, Germany, 2011; p. 13. [Google Scholar]
- Salinitro, M.; van der Ent, A.; Tognacchini, A.; Tassoni, A. Stress responses and nickel and zinc accumulation in different accessions of Stellaria media (L.) Vill. in response to solution pH variation in hydroponic culture. Plant Physiol. Biochem. 2020, 148, 133–141. [Google Scholar] [CrossRef]
- Mysliwa-Kurdziel, B.; Prasad, M.; Strzalka, K. Photosynthesis in heavy metal stressed plants. In Heavy Metal Stress in Plants: From Biomolecules to Ecosystems, 2nd ed.; Prasad, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 146–181. [Google Scholar]
- Viehweger, K.; Geipel, G. Uranium accumulation and tolerance in Arabidopsis halleri under native versus hydroponic conditions. Environ. Exp. Bot. 2010, 69, 39–46. [Google Scholar] [CrossRef]
- Küpper, H.; Parameswaran, A.; Leitenmaier, B.; Trtilek, M.; Setlik, I. Cadmium-induced inhibition of photosynthesis and long-term acclimation to cadmium stress in the hyperaccumulator Thlaspi Caerulescens. New Phytol. 2007, 175, 655–674. [Google Scholar] [CrossRef]
- Chandra, R.; Kang, H. Mixed heavy metal stress on photosynthesis, transpiration rate, and chlorophyll content in poplar hybrids. For. Sci. Technol. 2016, 12, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, A.; Sharma, I.; Tripati, B.N.; Munjal, A.K.; Baunthiyal, M.; Sharma, V. Metal toxicity and photosynthesis. In Photosynthesis: Overviews on Recent Progress and Future Perspectives, 1st ed.; Itoh, S., Mohanty, P., Guruprasad, K.N., Eds.; IK International Publishing House Pvt. Ltd.: New Delhi, India, 2012; pp. 229–236. [Google Scholar]
- Van der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Salinitro, M.; Tassoni, A.; Casolari, S.; de Laurentiis, F.; Zappi, A.; Melucci, D. Heavy metals bioindication potential of the common weeds Senecio vulgaris L., Polygonum aviculare L. and Poa annua L. Molecules 2019, 24, 2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penarrubia, L.; Andres-Colas, N.; Moreno, J.; Puig, S. Regulation of copper transport in Arabidopsis thaliana: A biochemical oscillator? J. Biol. Inorg. Chem. 2010, 15, 29–36. [Google Scholar] [CrossRef]
- Shanker, A.K.; Cervantes, C.; Loza-Tavera, H.; Avudainayagam, S. Chromium toxicity in plants. Environ. Int. 2005, 31, 739–753. [Google Scholar] [CrossRef]
- Lavid, N.; Schwartz, A.; Yarden, O.; Tel-Or, A. The involvement of polyphenols and peroxidase activities in heavy metal accumulation by epidermal glands of waterlily (Nympheaceae). Planta 2001, 212, 323–331. [Google Scholar] [CrossRef]
- Kidd, P.S.; Llugany, M.; Poschenrieder, C.; Gunsé, B.; Barceló, J. The role of root exudates in aluminium resistance and silicon-induced amelioration of aluminium toxicity in three varieties of maize (Zea mays L.). J. Exp. Bot. 2001, 52, 1339–1352. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Bidar, G.; Garcon, G.; Pruvot, C.; Dewaele, D.; Cazier, F.; Douay, F.; Shirali, P. Behavior of Trifolium repens and Lolium perenne growing in a heavy metal contaminated field: Plant metal concentration and phytotoxicity. Environ. Pollut. 2007, 147, 546–553. [Google Scholar] [CrossRef]
- Vajpayee, L.P.; Tripathi, R.D.; Rai, U.N.; Ali, M.B.; Singh, S.N. Chromium (VI) accumulation reduces chlorophyll biosynthesis, nitrate reductase activity and protein content in Nymphaea alba. Chemosphere 2000, 41, 1075–1082. [Google Scholar] [CrossRef]
- Kosobrukhov, A.; Knyazeva, I.; Mudrik, V. Plantago major plants responses to increase content of lead in soil: Growth and photosynthesis. Plant Growth Regul. 2004, 42, 145–151. [Google Scholar] [CrossRef]
- Bilal Shakoor, M.; Ali, S.; Hameed, A.; Farid, M.; Hussain, S.; Yasmeen, T.; Najeeb, U.; Aslam Bharwana, S.; Hasan Abbasi, G. Citric acid improves lead (Pb) phytoextraction in Brassica napus L. by mitigating Pb-induced morphological and biochemical damages. Ecotoxicol. Environ. Saf. 2014, 109, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Jagodin, B.; Govorina, V.; Vinogradova, S.; Zamaraev, A.; Chapovskaja, G. Cadmium and lead accumulation in some agricultural crops, grown in podzolic soils. Izvestija TSHA 1995, 2, 85–99. [Google Scholar]
- Shu, W.S.; Ye, Z.H.; Lan, C.Y.; Zhang, Z.Q.; Wong, M.H. Lead, zinc and copper accumulation and tolerance in populations of Paspalum distichum and Cynodon dactylon. Environ. Pollut. 2002, 120, 445–453. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Murawski, D.A.; Nason, J.D. The influence of seed dispersal mechanisms on the genetic structure of tropical tree populations. Vegetatio 1993, 107, 281–297. [Google Scholar] [CrossRef]
- Waters, M.T.; Bussell, J.D.; Jost, R. Arabidopsis hydroponics and shoot branching assay. Bio-protocol 2012, 2, e264. [Google Scholar] [CrossRef]
- Huang, C.Y.; Schulte, E.E. Digestion of plant tissue for analysis by ICP emission spectroscopy. Commun. Soil Sci. Plant Anal. 1985, 16, 943–958. [Google Scholar] [CrossRef]
- Ferri, M.; Gianotti, A.; Tassoni, A. Optimisation of assay conditions for the determination of antioxidant capacity and polyphenols in cereal food components. J. Food Compos. Anal. 2013, 30, 94–101. [Google Scholar] [CrossRef]
- Radwan, D.E.M.; Fayez, K.A.; Mahmoud, S.Y.; Hamad, A.; Lu, G. Physiological and metabolic changes of Cucurbita pepo leaves in response to zucchini yellow mosaic virus (ZYMV) infection and salicylic acid treatments. Plant Physiol. Biochem. 2007, 45, 480–489. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An. R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Dinno, A. Dunn’s Test of Multiple Comparisons Using Rank Sums. R Package Version 1.3.5. Available online: https://CRAN.R-project.org/package=dunn.test (accessed on 11 June 2019).
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots. R Package Version 0.3.0. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 11 June 2019).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal | Treatment | BAF P. aviculare | BAF S. vulgaris |
|---|---|---|---|
| Cd | A | 46.3 | 39.5 |
| B | 37.4 | 20.4 | |
| C | 29.5 | 11.2 | |
| D | 16.9 | 7.7 | |
| Cr | A | 0.7 | 4.0 |
| B | 0.9 | 5.2 | |
| C | 1.8 | 10.7 | |
| D | 5.6 | 18.3 | |
| Cu | A | 4.5 | 3.6 |
| B | 4.4 | 3.1 | |
| C | 5.5 | 2.9 | |
| D | 5.9 | 3.1 | |
| Ni | A | 13.7 | 34.5 |
| B | 8.7 | 25.5 | |
| C | 5.3 | 11.2 | |
| D | 2.7 | 9.9 | |
| Pb | A | 0.3 | 1.6 |
| B | 2.4 | 4.4 | |
| C | 4.5 | 5.3 | |
| D | 6.2 | 8.8 | |
| Zn | A | 26.9 | 48.0 |
| B | 21.1 | 39.1 | |
| C | 11.9 | 24.1 | |
| D | 8.9 | 10.7 |

| Treatment | Zn (mM) | Pb (mM) | Cu (mM) | Ni (mM) | Cr (mM) | Cd (μM) |
|---|---|---|---|---|---|---|
| Control | (2 µM) | 0 | (0.5 µM) | (0.05 µM) | 0 | 0 |
| A | 0.28 | 0.025 | 0.13 | 0.025 | 0.036 | 0.0945 |
| B | 0.56 | 0.25 | 0.26 | 0.125 | 0.18 | 9.45 |
| C | 1.12 | 1.25 | 0.65 | 0.250 | 0.36 | 47.25 |
| D | 2.80 | 2.50 | 1.30 | 0.625 | 0.90 | 94.50 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salinitro, M.; Hoogerwerf, S.; Casolari, S.; Zappi, A.; Melucci, D.; Tassoni, A. Production of Antioxidant Molecules in Polygonum aviculare (L.) and Senecio vulgaris (L.) under Metal Stress: A Possible Tool in the Evaluation of Plant Metal Tolerance. Int. J. Mol. Sci. 2020, 21, 7317. https://doi.org/10.3390/ijms21197317
Salinitro M, Hoogerwerf S, Casolari S, Zappi A, Melucci D, Tassoni A. Production of Antioxidant Molecules in Polygonum aviculare (L.) and Senecio vulgaris (L.) under Metal Stress: A Possible Tool in the Evaluation of Plant Metal Tolerance. International Journal of Molecular Sciences. 2020; 21(19):7317. https://doi.org/10.3390/ijms21197317
Chicago/Turabian StyleSalinitro, Mirko, Sara Hoogerwerf, Sonia Casolari, Alessandro Zappi, Dora Melucci, and Annalisa Tassoni. 2020. "Production of Antioxidant Molecules in Polygonum aviculare (L.) and Senecio vulgaris (L.) under Metal Stress: A Possible Tool in the Evaluation of Plant Metal Tolerance" International Journal of Molecular Sciences 21, no. 19: 7317. https://doi.org/10.3390/ijms21197317
APA StyleSalinitro, M., Hoogerwerf, S., Casolari, S., Zappi, A., Melucci, D., & Tassoni, A. (2020). Production of Antioxidant Molecules in Polygonum aviculare (L.) and Senecio vulgaris (L.) under Metal Stress: A Possible Tool in the Evaluation of Plant Metal Tolerance. International Journal of Molecular Sciences, 21(19), 7317. https://doi.org/10.3390/ijms21197317