Genomic Analysis of Hematopoietic Stem Cell at the Single-Cell Level: Optimization of Cell Fixation and Whole Genome Amplification (WGA) Protocol

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
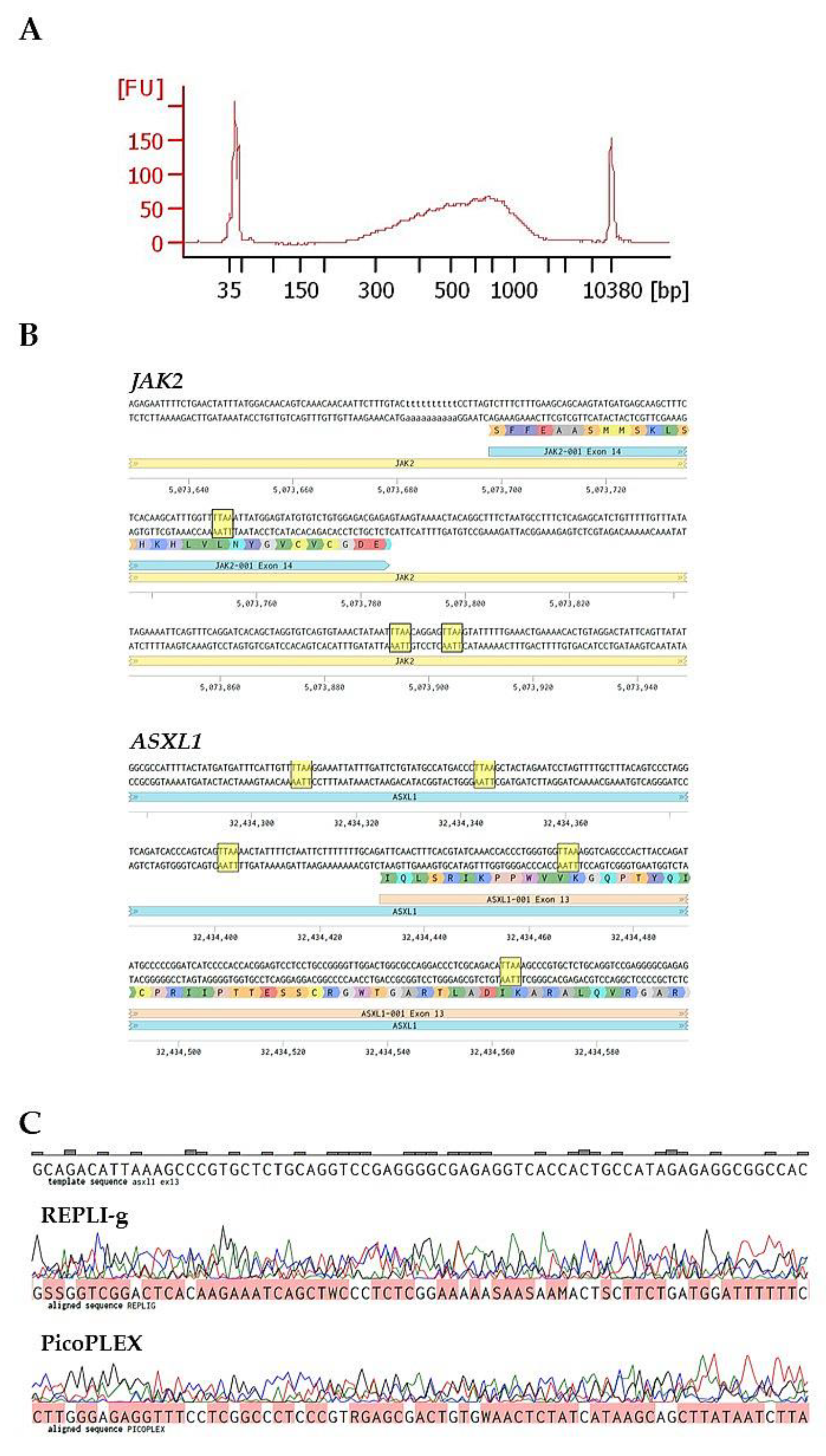
2.1. Cell Fixation Is Necessary for DEPArray Single-Cell Sorting but Can Affect WGA Efficiency
2.2. MetOH or Low-Concentration PFA Could Be Alternative Fixation Methods
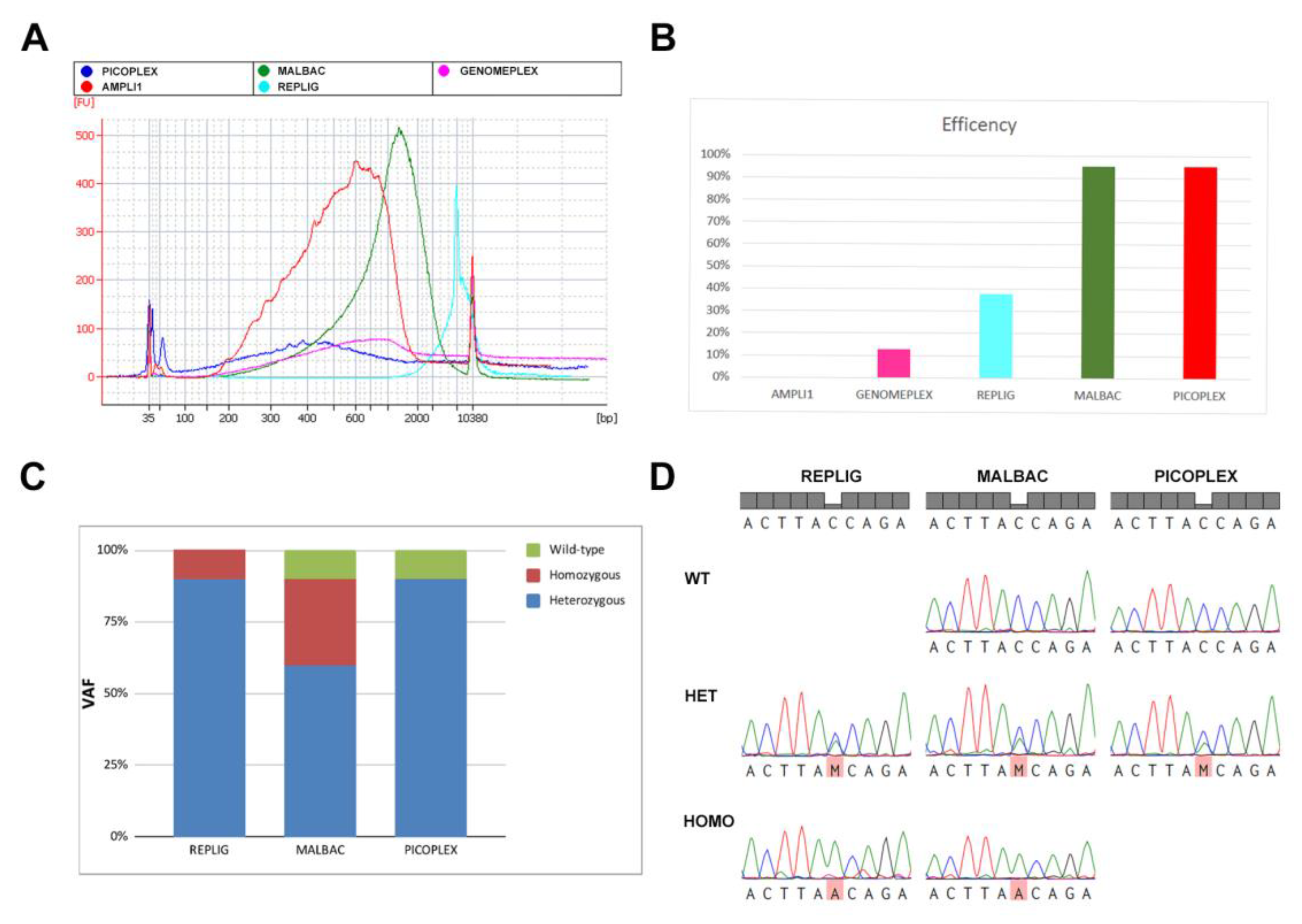
2.3. Comparison of ADO Effect in Single-Cell Whole Genome Amplification Kits
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. CD34+38- Cell Purification, Fixation and Immunostaining
4.3. CD34+38- Cell Sorting
4.4. Cell Lines Culture and Sorting
4.5. Single-Cell Whole Genome Amplification
4.6. Sanger Sequencing
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HSC | Hematopoietic Stem Cell |
| MPN | Myeloproliferative Neoplasm |
| scWGA | Single cell Whole Genome Amplification |
| FACS | Fluorescence-Activated Cell Sorting |
| WGA | Whole Genome Amplification |
| MDA | Multiple Displacement Amplification |
| DOP-PCR | Degenerative Oligonucleotide PCR |
| MALBAC | Multiple Annealing and Looping-Based Amplification |
| ADO | Allelic Dropout |
| PFA | Paraformaldehyde |
| MetOH | Methanol |
| NGS | Next Generation Sequencing |
References
- Navin, N.; Kendall, J.; Troge, J.; Andrews, P.; Rodgers, L.; McIndoo, J.; Cook, K.; Stepansky, A.; Levy, D.; Esposito, D.; et al. Tumour evolution inferred by single-cell sequencing. Nature 2011, 472, 90-U119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Hou, Y.; Yin, X.Y.; Bao, L.; Tang, A.F.; Song, L.T.; Li, F.Q.; Tsang, S.; Wu, K.; Wu, H.J.; et al. Single-Cell Exome Sequencing Reveals Single-Nucleotide Mutation Characteristics of a Kidney Tumor. Cell 2012, 148, 886–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.; Yu, J.; Yao, X.T.; Wu, W.K.K.; Lu, Y.Y.; Tang, S.W.; Li, X.C.; Bao, L.; Li, X.X.; Hou, Y.; et al. Discovery of biclonal origin and a novel oncogene SLC12A5 in colon cancer by single-cell sequencing. Cell Res. 2014, 24, 701–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, J.; Kendall, J.; McIndoo, J.; Rodgers, L.; Aboukhalil, R.; Levy, D.; Stepansky, A.; Sun, G.L.; Chobardjiev, L.; Riggs, M.; et al. Utility of Single-Cell Genomics in Diagnostic Evaluation of Prostate Cancer. Cancer Res. 2018, 78, 348–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuipers, J.; Jahn, K.; Beerenwinkel, N. Advances in understanding tumour evolution through single-cell sequencing. Biochim. Biophys. Acta Rev. Cancer 2017, 1867, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Paguirigan, A.L.; Smith, J.; Meshinchi, S.; Carroll, M.; Maley, C.; Radich, J.P. Single-cell genotyping demonstrates complex clonal diversity in acute myeloid leukemia. Sci. Transl. Med. 2015, 7, 281re282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, E.A.; Frey, B.L.; Smith, L.M.; Auble, D.T. Formaldehyde crosslinking: A tool for the study of chromatin complexes. J. Biol. Chem. 2015, 290, 26404–26411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, F.B.; Hosono, S.; Fang, L.; Wu, X.; Faruqi, A.F.; Bray-Ward, P.; Sun, Z.; Zong, Q.; Du, Y.; Du, J.; et al. Comprehensive human genome amplification using multiple displacement amplification. Proc. Natl. Acad. Sci. USA 2002, 99, 5261–5266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telenius, H.; Carter, N.P.; Bebb, C.E.; Nordenskjold, M.; Ponder, B.A.; Tunnacliffe, A. Degenerate oligonucleotide-primed PCR: General amplification of target DNA by a single degenerate primer. Genomics 1992, 13, 718–725. [Google Scholar] [CrossRef]
- Zong, C.; Lu, S.; Chapman, A.R.; Xie, X.S. Genome-wide detection of single-nucleotide and copy-number variations of a single human cell. Science 2012, 338, 1622–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortmann, C.A.; Kent, D.G.; Nangalia, J.; Silber, Y.; Wedge, D.C.; Grinfeld, J.; Baxter, E.J.; Massie, C.E.; Papaemmanuil, E.; Menon, S.; et al. Effect of mutation order on myeloproliferative neoplasms. N. Engl. J. Med. 2015, 372, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Trapani, M.; Manaresi, N.; Medoro, G. DEPArray system: An automatic image-based sorter for isolation of pure circulating tumor cells. Cytometry A 2018, 93, 1260–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, P.; Zhang, W.; Xin, H.; Deng, G. Single Cell Isolation and Analysis. Front. Cell Dev. Biol. 2016, 4, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passamonti, F.; Maffioli, M.; Caramazza, D.; Cazzola, M. Myeloproliferative neoplasms: From JAK2 mutations discovery to JAK2 inhibitor therapies. Oncotarget 2011, 2, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viny, A.D.; Levine, R.L. Genetics of myeloproliferative neoplasms. Cancer J. 2014, 20, 61–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobro, A.J.; Smith, N.I. An evaluation of fixation methods: Spatial and compositional cellular changes observed by Raman imaging. Vib. Spectrosc. 2017, 91, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Ediriwickrema, A.; Aleshin, A.; Reiter, J.G. Single-cell mutational profiling enhances the clinical evaluation of AML MRD (vol 4, pg 943, 2020). Blood Adv. 2020, 4, 1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunbar, A.J.; Rampal, R.K.; Levine, R. Leukemia secondary to myeloproliferative neoplasms. Blood 2020, 136, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Salati, S.; Genovese, E.; Carretta, C.; Zini, R.; Bartalucci, N.; Prudente, Z.; Pennucci, V.; Ruberti, S.; Rossi, C.; Rontauroli, S.; et al. Calreticulin Ins5 and Del52 mutations impair unfolded protein and oxidative stress responses in K562 cells expressing CALR mutants. Sci. Rep. 2019, 9, 10558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, A.; Schoendube, J.; Zimmermann, S.; Steeb, M.; Zengerle, R.; Koltay, P. Technologies for Single-Cell Isolation. Int. J. Mol. Sci. 2015, 16, 16897–16919. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WGA Method | Compatible with PFA Fixation (High Concentration and Long Time) | Compatible with PFA Fixation (Low Concentration and Short Time) | Compatible with MetOH Fixation | Efficiency (%) | ADO Effect | Sequence Quality |
|---|---|---|---|---|---|---|
| Ampli1 | Yes | Yes | Yes | 0 | - | - |
| Genomeplex | No | No | Yes | 12.5 | - | - |
| REPLI-g | No | No | Yes | 37.5 | Low | Low |
| PicoPLEX | No | Yes | Yes | 95 | Low | High |
| MALBAC | No | Yes | Yes | 95 | High | Medium |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carretta, C.; Mallia, S.; Genovese, E.; Parenti, S.; Rontauroli, S.; Bianchi, E.; Fantini, S.; Sartini, S.; Tavernari, L.; Tagliafico, E.; et al. Genomic Analysis of Hematopoietic Stem Cell at the Single-Cell Level: Optimization of Cell Fixation and Whole Genome Amplification (WGA) Protocol. Int. J. Mol. Sci. 2020, 21, 7366. https://doi.org/10.3390/ijms21197366
Carretta C, Mallia S, Genovese E, Parenti S, Rontauroli S, Bianchi E, Fantini S, Sartini S, Tavernari L, Tagliafico E, et al. Genomic Analysis of Hematopoietic Stem Cell at the Single-Cell Level: Optimization of Cell Fixation and Whole Genome Amplification (WGA) Protocol. International Journal of Molecular Sciences. 2020; 21(19):7366. https://doi.org/10.3390/ijms21197366
Chicago/Turabian StyleCarretta, Chiara, Selene Mallia, Elena Genovese, Sandra Parenti, Sebastiano Rontauroli, Elisa Bianchi, Sebastian Fantini, Stefano Sartini, Lara Tavernari, Enrico Tagliafico, and et al. 2020. "Genomic Analysis of Hematopoietic Stem Cell at the Single-Cell Level: Optimization of Cell Fixation and Whole Genome Amplification (WGA) Protocol" International Journal of Molecular Sciences 21, no. 19: 7366. https://doi.org/10.3390/ijms21197366
APA StyleCarretta, C., Mallia, S., Genovese, E., Parenti, S., Rontauroli, S., Bianchi, E., Fantini, S., Sartini, S., Tavernari, L., Tagliafico, E., & Manfredini, R. (2020). Genomic Analysis of Hematopoietic Stem Cell at the Single-Cell Level: Optimization of Cell Fixation and Whole Genome Amplification (WGA) Protocol. International Journal of Molecular Sciences, 21(19), 7366. https://doi.org/10.3390/ijms21197366