A SEMA3 Signaling Pathway-Based Multi-Biomarker for Prediction of Glioma Patient Survival

 , ,
, ,
Abstract
:1. Introduction
2. Results
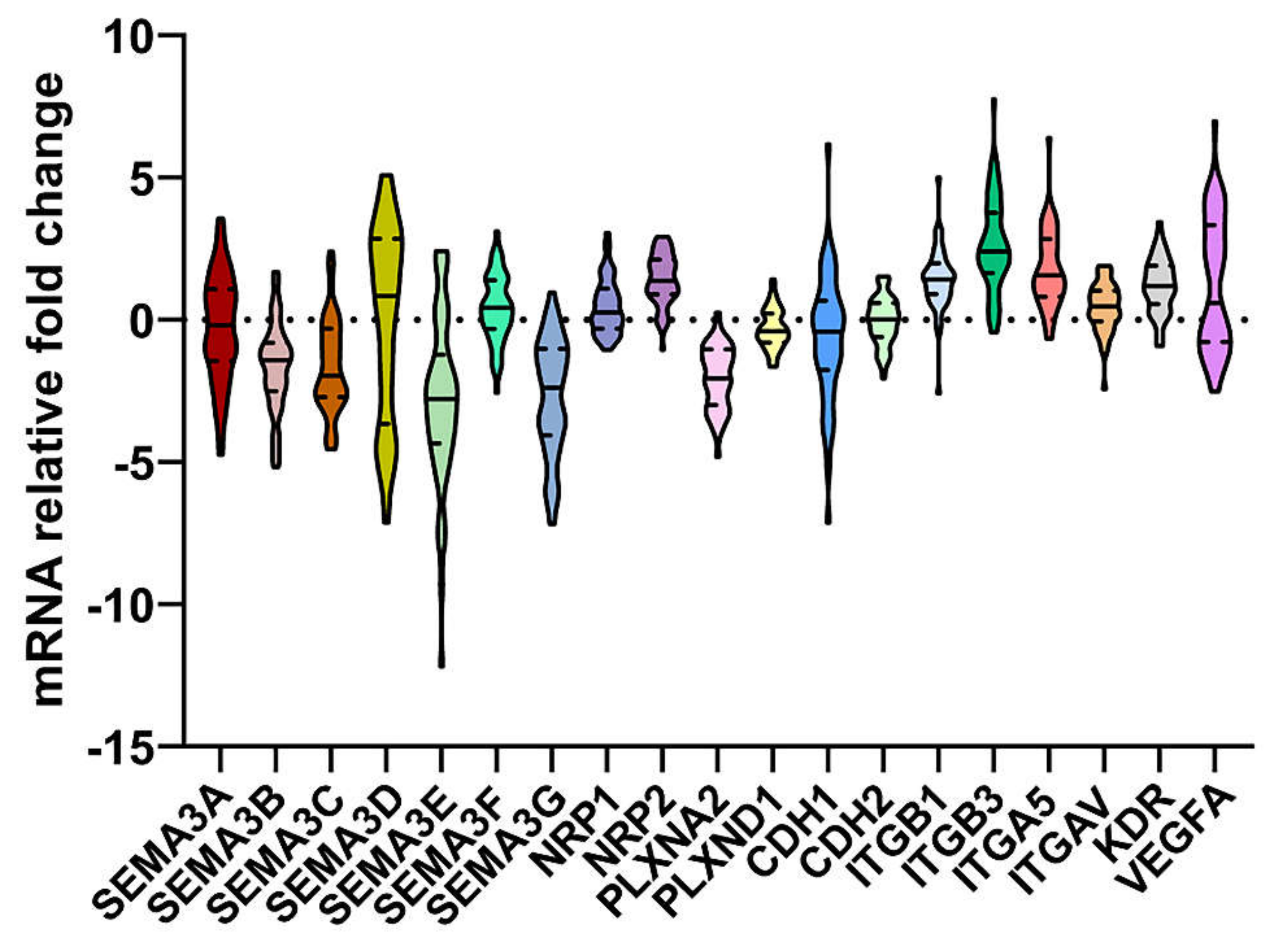
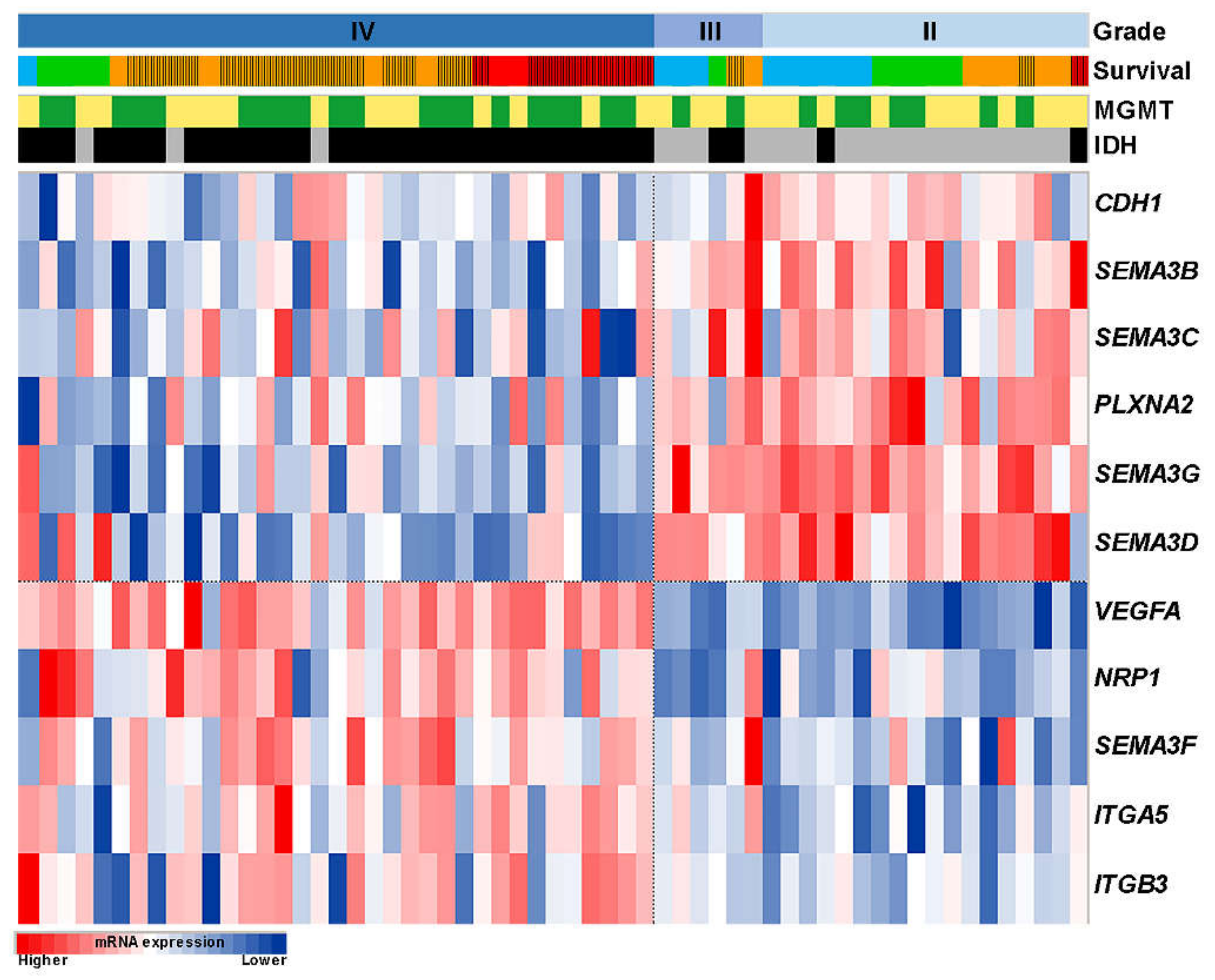
2.1. Associations of Expression Changes of Key SEMA3 Signaling Genes in Astrocytomas with Patient Clinical Data
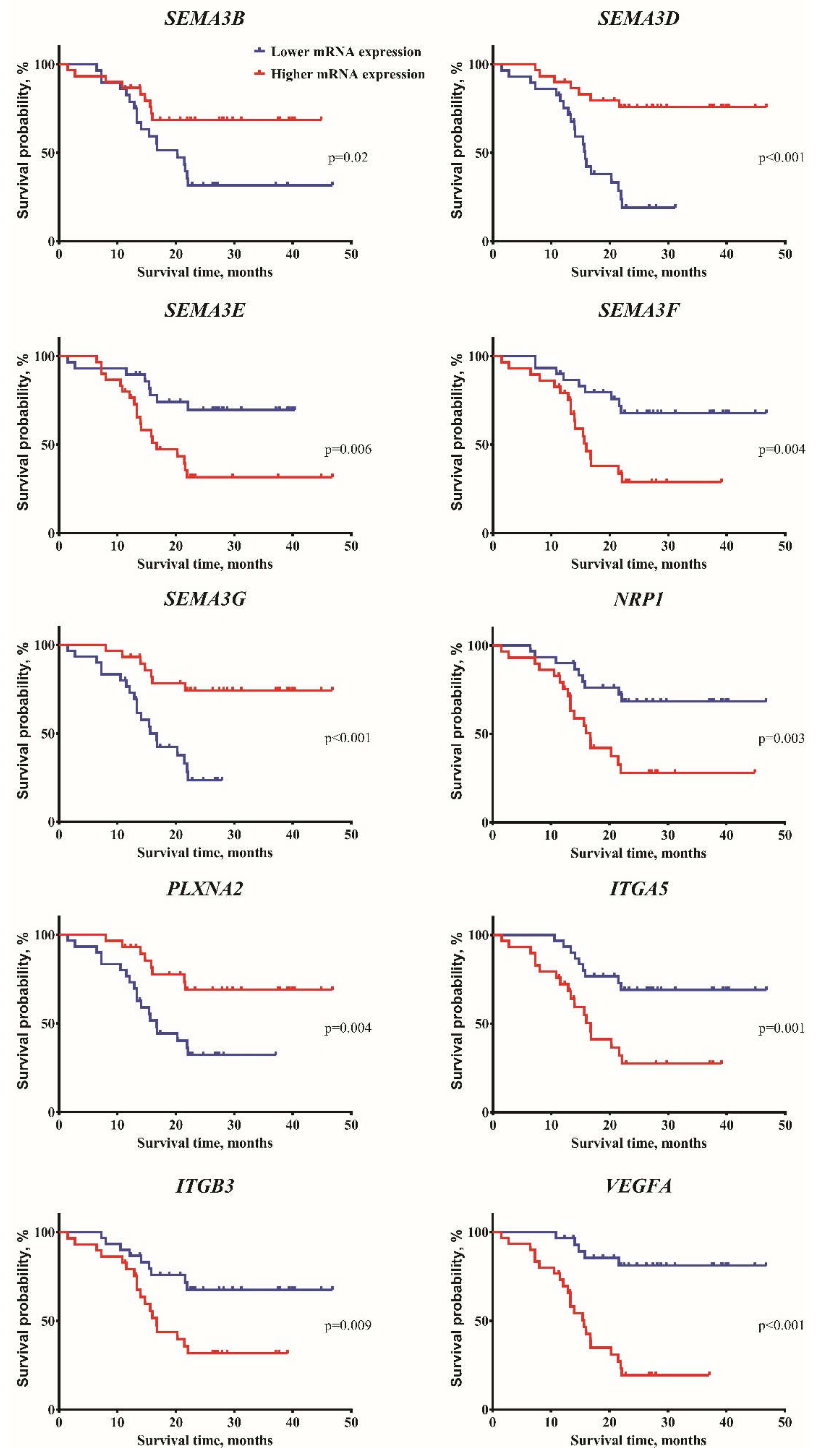
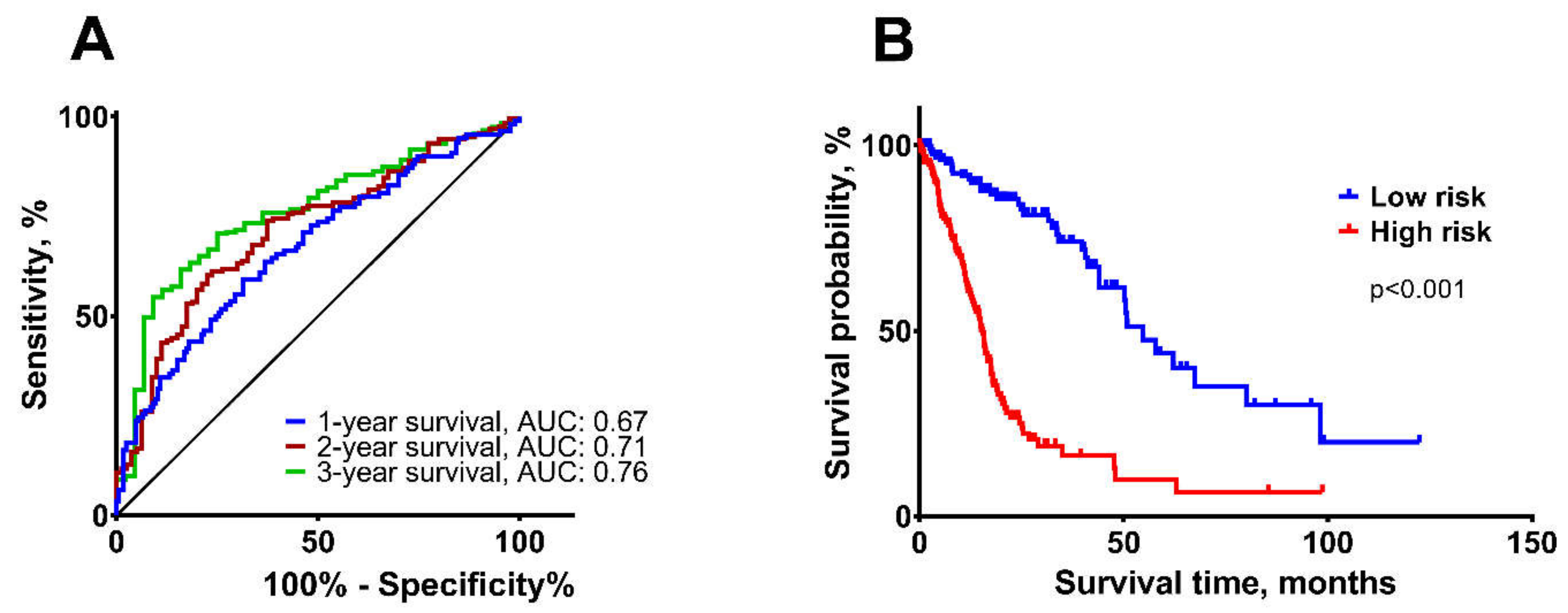
2.2. Construction of the Multi-Biomarker for Better Patient Survival Prediction
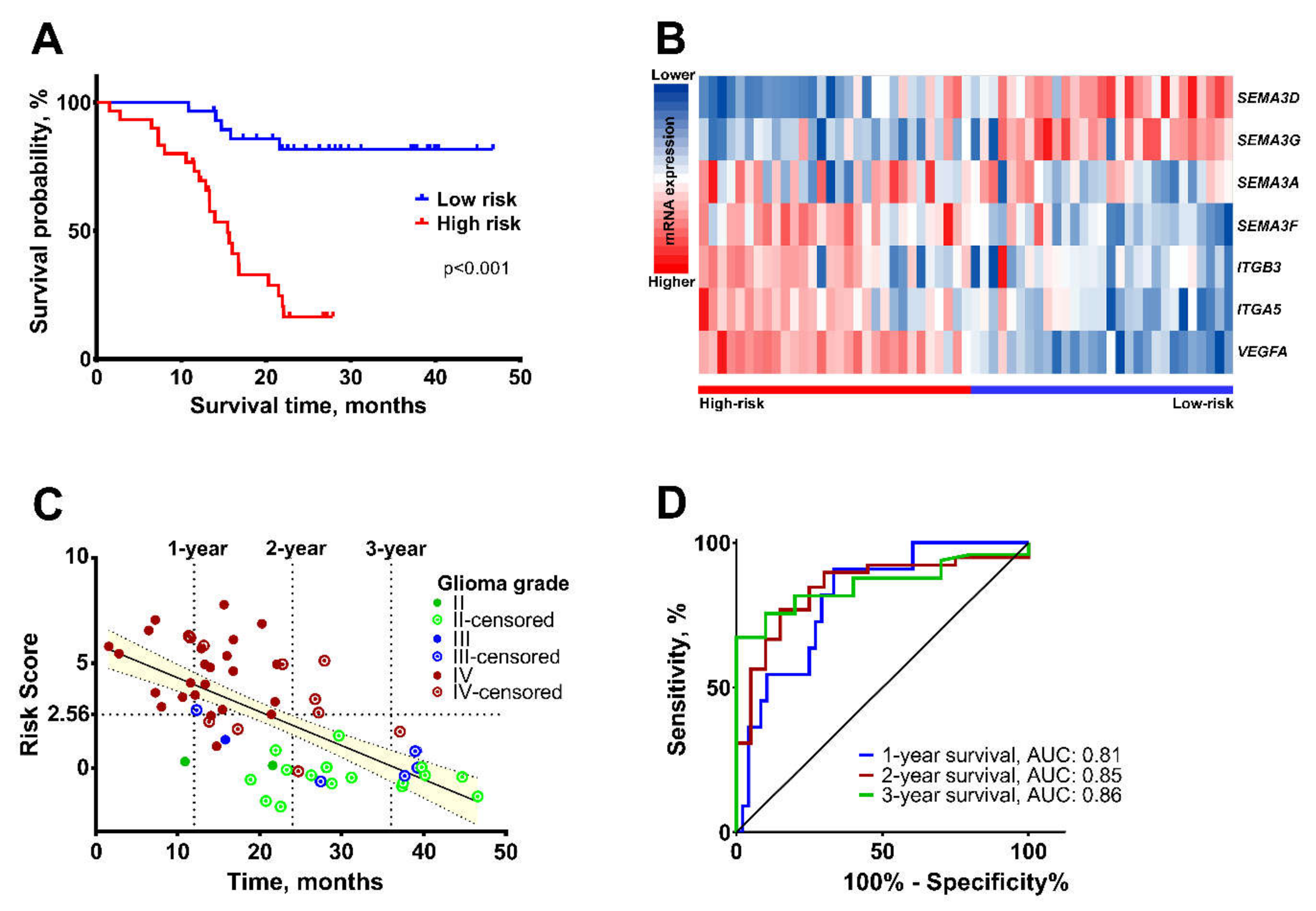
2.3. Validation of the Survival Prediction Model
3. Discussion
4. Materials and Methods
4.1. Data of Patients
4.2. Total RNA Extraction and cDNA Synthesis
4.3. Quantitative Real-Time PCR
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GBM | Glioblastoma |
| CNS | Central Nervous System |
| WHO | World Health Organization |
| TCGA | The Cancer Genomic Atlas |
| SEMA3 | Class 3 Semaphorin |
| NRP | Neuropilin |
| PLXN | Plexin |
| CDH | Cadherin |
| ITG | Integrin |
| VEGF | Vascular Endothelial Growth Factor |
| MGMT | O6-methylguanine DNA Methyltransferase |
| IDH | Isocitrate Dehydrogenase |
| AUC | Area Under the Curve |
References
- Rasmussen, B.K.; Hansen, S.; Laursen, R.J.; Kosteljanetz, M.; Schultz, H.; Norgard, B.M.; Guldberg, R.; Gradel, K.O. Epidemiology of Glioma: Clinical Characteristics, Symptoms, and Predictors of Glioma Patients Grade I–IV in the the Danish Neuro-Oncology Registry. J. Neurooncol. 2017, 135, 571–579. [Google Scholar] [CrossRef]
- Louis, D.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.; Ohgaki, H.; Wiestler, O.; Kleihues, P.; Ellison, D. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A Summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [Green Version]
- Wesseling, P.; Capper, D. WHO 2016 Classification of Gliomas. Neuropathol. Appl. Neurobiol. 2018, 44, 139–150. [Google Scholar] [CrossRef]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO Classification of Tumours of the Central Nervous System. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Furnari, F.B.; Fenton, T.; Bachoo, R.M.; Mukasa, A.; Stommel, J.M.; Stegh, A.; Hahn, W.C.; Ligon, K.L.; Louis, D.N.; Brennan, C.; et al. Malignant Astrocytic Glioma: Genetics, Biology, and Paths to Treatment. Genes Dev. 2007, 21, 2683–2710. [Google Scholar] [CrossRef] [Green Version]
- Grossman, S.A.; Ye, X.; Piantadosi, S.; Desideri, S.; Nabors, L.B.; Rosenfeld, M.; Fisher, J. Survival of Patients with Newly Diagnosed Glioblastoma Treated with Radiation and Temozolomide on Research Studies in the United States. Clin. Cancer Res. 2010, 16, 2443–2449. [Google Scholar] [CrossRef] [Green Version]
- Reifenberger, G.; Wirsching, H.G.; Knobbe-Thomsen, C.B.; Weller, M. Advances in the Molecular Genetics of Gliomas-Implications for Classification and Therapy. Nat. Rev. Clin. Oncol. 2017, 14, 434–452. [Google Scholar] [CrossRef]
- Phillips, H.S.; Kharbanda, S.; Chen, R.; Forrest, W.F.; Soriano, R.H.; Wu, T.D.; Misra, A.; Nigro, J.M.; Colman, H.; Soroceanu, L.; et al. Molecular Subclasses of High-Grade Glioma Predict Prognosis, Delineate a Pattern of Disease Progression, and Resemble Stages in Neurogenesis. Cancer Cell 2006, 9, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. An Integrated Genomic Analysis Identifies Clinically Relevant Subtypes of Glioblastoma Characterized by Abnormalities in PDGFRA, IDH1, EGFR and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Brito, C.; Azevedo, A.; Esteves, S.; Marques, A.R.; Martins, C.; Costa, I.; Mafra, M.; Bravo Marques, J.M.; Roque, L.; Pojo, M. Clinical Insights Gained by Refining the 2016 WHO Classification of Diffuse Gliomas with: EGFR Amplification, TERT Mutations, PTEN Deletion and MGMT Methylation. BMC Cancer 2019, 19, 968. [Google Scholar] [CrossRef] [Green Version]
- Cohen, A.; Holmen, S.; Colman, H. IDH1 and IHD2 Mutations in Gliomas. Curr. Neurol. Neurosci. Rep. 2013, 13, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lang, F.; Chou, F.J.; Zaghloul, K.A.; Yang, C. Isocitrate Dehydrogenase Mutations in Glioma: Genetics, Biochemistry, and Clinical Indications. Biomedicines 2020, 8, 294. [Google Scholar] [CrossRef] [PubMed]
- Hegi, M.E.; Diserens, A.-C.; Gorlia, T.; Hamou, M.-F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT Gene Silencing and Benefit from Temozolomide in Glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledano, S.; Nir-Zvi, I.; Engelman, R.; Kessler, O.; Neufeld, G. Class-3 Semaphorins and Their Receptors: Potent Multifunctional Modulators of Tumor Progression. Int. J. Mol. Sci. 2019, 20, 556–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alto, L.T.; Terman, J.R. Semaphorins and Their Signaling Mechanisms. Methods Mol. Biol. 2017, 1493, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, G.; Mumblat, Y.; Smolkin, T.; Toledano, S.; Nir-Zvi, I.; Ziv, K.; Kessler, O. The Role of the Semaphorins in Cancer. Cell Adh. Migr. 2016, 10, 652–674. [Google Scholar] [CrossRef] [Green Version]
- Tamagnone, L. Emerging Role of Semaphorins as Major Regulatory Signals and Potential Therapeutic Targets in Cancer. Cancer Cell 2012, 22, 145–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaur, P.; Bielenberg, D.R.; Samuel, S.; Bose, D.; Zhou, Y.; Gray, M.J.; Dallas, N.A.; Fan, F.; Xia, L.; Lu, J.; et al. Role of Class 3 Semaphorins and Their Receptors in Tumor Growth and Angiogenesis. Clin. Cancer Res. 2009, 15, 6763–6770. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Gunput, R.A.F.; Pasterkamp, R.J. Semaphorin Signaling: Progress Made and Promises Ahead. Trends Biochem. Sci. 2008, 33, 161–170. [Google Scholar] [CrossRef]
- Nasarre, P.; Gemmill, R.M.; Drabkin, H.A. The Emerging Role of Class-3 Semaphorins and Their Neuropilin Receptors in Oncology. Onco Targets Ther. 2014, 7, 1663–1687. [Google Scholar] [CrossRef] [Green Version]
- Tam, K.J.; Hui, D.H.F.; Lee, W.W.; Dong, M.; Tombe, T.; Jiao, I.Z.F.; Khosravi, S.; Takeuchi, A.; Peacock, J.W.; Ivanova, L.; et al. Semaphorin 3 C Drives Epithelial-to-Mesenchymal Transition, Invasiveness, and Stem-like Characteristics in Prostate Cells. Sci. Rep. 2017, 7, 11501–11513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasarre, P.; Kusy, S.; Constantin, B.; Castellani, V.; Drabkin, H.A.; Bagnard, D.; Roche, J. Semaphorin SEMA3F Has a Repulsing Activity on Breast Cancer Cells and Inhibits E-Cadherin-Mediated Cell Adhesion. Neoplasia 2005, 7, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.T.; Nilchi, L.; Li, X.; Gangaraju, S.; Jiang, S.X.; Aylsworth, A.; Monette, R.; Slinn, J. Semaphorin3A Elevates Vascular Permeability and Contributes to Cerebral Ischemia-Induced Brain Damage. Sci. Rep. 2015, 5, 7890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, W.; Gu, C. The Role and Mechanism-of-Action of Sema3E and Plexin-D1 in Vascular. Semin. Cell Dev. Biol. 2013, 24, 156–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, R.L.; Wang, Q.; Carro, A.; Verhaak, R.G.; Squatrito, M. GlioVis Data Portal for Visualization and Analysis of Brain Tumor Expression Datasets. Neuro Oncol. 2017, 19, 139–141. [Google Scholar] [CrossRef] [Green Version]
- Crocetti, E.; Trama, A.; Stiller, C.; Caldarella, A.; Soffietti, R.; Jaal, J.; Weber, D.C.; Ricardi, U.; Slowinski, J.; Brandes, A.; et al. Epidemiology of Glial and Non-Glial Brain Tumours in Europe. Eur. J. Cancer 2012, 48, 1532–1542. [Google Scholar] [CrossRef]
- Perez-Ibave, D.C.; Burciaga-Flores, C.H.; Elizondo-Riojas, M.A. Prostate-Specific Antigen (PSA) as a Possible Biomarker in Non-Prostatic Cancer: A Review. Cancer Epidemiol. 2018, 54, 48–55. [Google Scholar] [CrossRef]
- James, C.R.; Quinn, J.E.; Mullan, P.B.; Johnston, P.G.; Harkin, D.P. BRCA1, a Potential Predictive Biomarker in the Treatment of Breast Cancer. Oncologist 2007, 12, 142–150. [Google Scholar] [CrossRef]
- Walsh, M.; Nathanson, K.; Couch, F.; Offit, K. Genomic Biomarkers for Breast Cancer Risk. Adv. Exp. Med. Biol. 2016, 882, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.Z.; Guo, L.M.; Xu, T.Q.; Yin, Y.H.; Jia, F. Identification of a Multidimensional Transcriptome Signature for Survival Prediction of Postoperative Glioblastoma Multiforme Patients. J. Transl. Med. 2018, 16, 368. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Tang, G.; Zhou, Q.; Cao, Y.; Li, H.; Fu, X.; Wu, Z.; Jiang, X. Expression Profile Analysis Identifies a Novel Five-Gene Signature to Improve Prognosis Prediction of Glioblastoma. Front. Genet. 2019, 10, 419. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Yang, J.; Wang, H.; Liu, D.; Zhang, H. A Potential Prognostic Gene Signature for Predicting Survival for Glioblastoma Patients. Biomed. Res. Int. 2019, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Lu, J.; Li, H.; Qi, S.; Yu, L. Identification of a Long Noncoding RNA Signature to Predict Outcomes of Glioblastoma. Mol. Med. Rep. 2019, 19, 5406–5416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, J.B.-K.; Chang, T.-H.; Lee, G.A.; Lee, T.-Y.; Chen, C.-Y. Identification of Potential Biomarkers Related to Glioma Survival by Gene Expression Profile Analysis. BMC Med. Genomics 2019, 11, 34. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Guan, G.; Zhao, W.; Zhuang, M. A Risk Classification System with Five-Gene for Survival Prediction of Glioblastoma Patients. Front. Neurol. 2019, 10, 745. [Google Scholar] [CrossRef] [Green Version]
- Vaitkienė, P.; Skiriutė, D.; Steponaitis, G.; Skauminas, K.; Tamašauskas, A. High Level of Sema3C Is Associated with Glioma Malignancy. Diagn. Pathol. 2015, 10, 58. [Google Scholar] [CrossRef] [Green Version]
- Bender, R.J.; Mac Gabhann, F. Expression of VEGF and Semaphorin Genes Define Subgroups of Triple Negative Breast Cancer. PLoS ONE 2013, 8, e61788. [Google Scholar] [CrossRef] [Green Version]
- Bender, R.J.; Mac Gabhann, F. Dysregulation of the Vascular Endothelial Growth Factor and Semaphorin Ligand-Receptor Families in Prostate Cancer Metastasis. BMC Syst. Biol. 2015, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Karayan-Tapon, L.; Wager, M.; Guilhot, J.; Levillain, P.; Marquant, C.; Clarhaut, J.; Potiron, V.; Roche, J. Semaphorin, Neuropilin and VEGF Expression in Glial Tumours: SEMA3G, a Prognostic Marker? Br. J. Cancer 2008, 99, 1153–1160. [Google Scholar] [CrossRef] [Green Version]
- Etxaniz, O.; Carrato, C.; de Aguirre, I.; Queralt, C.; Muñoz, A.; Ramirez, J.L.; Rosell, R.; Villà, S.; Diaz, R.; Estival, A.; et al. IDH mutation status trumps the Pignatti risk score as a prognostic marker in low-grade gliomas. J. Neurooncol. 2017, 135, 273–284. [Google Scholar] [CrossRef]
- Han, S.; Liu, Y.; Cai, S.J.; Qian, M.; Ding, J.; Larion, M.; Gilbert, M.R.; Yang, C. IDH mutation in glioma: Molecular mechanisms and potential therapeutic targets. Br. J. Cancer 2020, 122, 1580–1589. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, C.; Lama, G.; Sica, G. Multifaceted Functional Role of Semaphorins in Glioblastoma. Int. J. Mol. Sci. 2019, 20, 2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabag, A.D.; Bode, J.; Fink, D.; Kigel, B.; Kugler, W.; Neufeld, G. Semaphorin-3D and Semaphorin-3E Inhibit the Development of Tumors from Glioblastoma Cells Implanted in the Cortex of the Brain. PLoS ONE 2012, 7, e42912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Ma, L.; Li, J.; Gu, J.; Shi, Q.; Yu, R. Effects of SEMA3G on Migration and Invasion of Glioma Cells. Oncol. Rep. 2012, 28, 269–275. [Google Scholar] [CrossRef]
- Avraamides, C.; Garmy-Susini, B.; Varner, J. Integrins in Angiogenesis and Lymphangiogenesis. Nat. Rev. Cancer 2008, 8, 604–617. [Google Scholar] [CrossRef] [Green Version]
- Schittenhelm, J.; Schwab, E.I.; Sperveslage, J.; Tatagiba, M.; Meyermann, R.; Fend, F.; Goodman, S.L.; Sipos, B. Longitudinal Expression Analysis of Av Integrins in Human Gliomas Reveals Upregulation of Integrin Avβ3 as a Negative Prognostic Factor. J. Neuropathol. Exp. Neurol. 2013, 72, 194–210. [Google Scholar] [CrossRef] [Green Version]
- Treps, L.; Perret, R.; Edmond, S.; Ricard, D.; Gavard, J. Glioblastoma Stem-like Cells Secrete the pro-Angiogenic VEGF-A Factor in Extracellular Vesicles. J. Extracell. Vesicles 2017, 6, 1359479. [Google Scholar] [CrossRef]
- Mehdi, T.; Ahmadi, B.N. Kernel Smoothing for ROC Curve and Estimation for Thyroid Stimulating Hormone. Int. J. Public Heal. Res. Spec. Issue 2011, 239–242. [Google Scholar]
- Zhu, W.; Zeng, N.; Wang, N. Sensitivity, Specificity, Accuracy, Associated Confidence Interval and ROC Analysis with Practical SAS® Implementations. Heal. Care Life Sci. 2010, 19, 67. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Gender | Age (Years) | Tumor Grade | IDH | MGMT | Survival | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Female, Median | Male, Median | p # | ≤50, Median | >50, Median | p # | II, Median | III, Median | IV, Median | p * | Wt, Median | Mut, Median | p # | M, Median | U, Median | p # | Log-Rank, p | |
| SEMA3A | −0.12 | −0.54 | 0.867 | −0.77 | 0.79 | 0.024 | −0.6 | −1.08 | 0.69 | 0.073 | 0.14 | −0.54 | 0.32 | −0.04 | −0.39 | 0.952 | 0.181 |
| SEMA3B | −1.81 | −0.99 | 0.049 | −0.98 | −2.06 | 0.001 | −0.79 | −0.63 | −2.36 | <0.001 | −2.03 | −0.86 | <0.001 | −1.24 | −1.64 | 0.154 | 0.02 |
| SEMA3C | −2.22 | −1.37 | 0.099 | −1.4 | −2.54 | 0.089 | −0.88 | −1.25 | −2.6 | 0.023 | −2.51 | −0.98 | <0.001 | −1.05 | −2.55 | 0.006 | 0.095 |
| SEMA3D | −0.89 | 1.74 | 0.031 | 2.79 | −1.98 | <0.001 | 2.89 | 2.81 | −1.94 | <0.001 | −1.98 | 2.86 | <0.001 | 1.26 | −0.26 | 0.237 | <0.001 |
| SEMA3E | −3.36 | −2.4 | 0.072 | −3.33 | −2.2 | 0.071 | −2.79 | −3.43 | −2.64 | 0.815 | −2.55 | −3.33 | 0.393 | −3.45 | −2.45 | 0.15 | 0.006 |
| SEMA3F | 0.64 | 0.08 | 0.24 | −0.11 | 0.71 | 0.006 | −0.24 | 0.08 | 0.76 | 0.001 | 0.71 | −0.15 | 0.004 | −0.11 | 0.69 | 0.017 | 0.004 |
| SEMA3G | −3.58 | −1.74 | 0.006 | −1.28 | −3.57 | <0.001 | −0.96 | −0.96 | −3.59 | <0.001 | −3.58 | −1.19 | <0.001 | −1.95 | −3.34 | 0.371 | <0.001 |
| NRP1 | 0.52 | 0.14 | 0.189 | −0.12 | 0.73 | 0.002 | −0.14 | −0.44 | 0.78 | <0.001 | 0.65 | −1.12 | 0.006 | 0.24 | 0.26 | 0.617 | 0.003 |
| NRP2 | 1.69 | 1.29 | 0.436 | 1.74 | 0.98 | 0.011 | 1.79 | 1.79 | 1.14 | 0.086 | 1.23 | 1.66 | 0.043 | 1.66 | 1.14 | 0.262 | 0.300 |
| PLXNA2 | −2.49 | −1.37 | 0.003 | −1.37 | −2.32 | 0.001 | −1.17 | −1.47 | −2.35 | <0.001 | −2.34 | −1.04 | <0.001 | −1.81 | −2.09 | 0.413 | 0.004 |
| PLXND1 | −0.36 | −0.43 | 0.976 | −0.43 | −0.33 | 0.585 | −0.35 | −0.34 | −0.47 | 0.902 | −0.29 | −0.47 | 0.419 | −0.48 | −0.2 | 0.214 | 0.371 |
| CDH1 | −0.14 | −0.63 | 0.541 | −0.03 | −1.23 | 0.044 | 0.39 | −0.98 | −1.1 | 0.029 | −1.14 | 0.06 | 0.017 | −1.08 | −0.05 | 0.628 | 0.065 |
| CDH2 | −0.07 | 0.13 | 0.384 | 0.01 | 0.02 | 0.606 | −0.01 | −0.4 | 0.13 | 0.27 | 0.09 | −0.04 | 0.698 | −0.06 | 0.24 | 0.172 | 0.605 |
| ITGB1 | 1.56 | 1.38 | 0.169 | 1.42 | 1.45 | 0.952 | 1.25 | 0.95 | 1.6 | 0.109 | 1.52 | 1.31 | 0.22 | 1.4 | 1.52 | 0.773 | 0.841 |
| ITGB3 | 2.62 | 2.32 | 0.963 | 2.15 | 3.08 | 0.013 | 1.99 | 2.1 | 3.61 | 0.005 | 2.95 | 2.05 | 0.017 | 2.31 | 2.43 | 0.74 | 0.009 |
| ITGA5 | 1.69 | 1.41 | 0.625 | 1.19 | 2.27 | 0.003 | 0.811 | 1.31 | 2.28 | 0.001 | 2.27 | 1.03 | <0.001 | 1.26 | 1.71 | 0.182 | 0.001 |
| ITGAV | 0.16 | 0.64 | 0.096 | 0.77 | 0.19 | 0.022 | 0.84 | 0.7 | 0.24 | 0.145 | 0.25 | 0.68 | 0.157 | 0.43 | 0.53 | 0.988 | 0.451 |
| VEGFA | 2.22 | −0.09 | 0.063 | −0.68 | 2.95 | <0.001 | −1.07 | −0.64 | 2.94 | <0.001 | 2.83 | −0.78 | <0.001 | −0.06 | 2.61 | 0.008 | <0.001 |
| KDR | 1.12 | 1.39 | 0.123 | 0.92 | 1.37 | 0.66 | 0.86 | 1.14 | 1.36 | 0.769 | 1.34 | 0.89 | 0.975 | 0.91 | 1.68 | 0.012 | 0.574 |
| The signature | 3.68 | 1.36 | 0.067 | 0.01 | 4.34 | <0.001 | −0.39 | 0.41 | 4.62 | <0.001 | 4.34 | −0.33 | <0.001 | 0.96 | 3.38 | 0.027 | <0.001 |
| Univariate | Multivariate | Multivariate * | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HR | 95% CI | p | HR | 95% CI | p | HR | 95% CI | p | ||||
| Lower | Upper | Lower | Upper | Lower | Upper | |||||||
| Sex | ||||||||||||
| Male vs. Female | 1.075 | 0.503 | 2.298 | 0.852 | ||||||||
| Age (years) | 1.072 | 1.041 | 1.105 | <0.001 | - | - | - | - | 1.042 | 1.004 | 1.082 | 0.031 |
| Tumor grade | ||||||||||||
| II | 1 | |||||||||||
| III | 1.723 | 0.156 | 19.010 | 0.657 | ||||||||
| IV (GBM) | 11.825 | 2.769 | 50.495 | 0.001 | - | - | - | - | - | - | - | - |
| MGMT | ||||||||||||
| M vs. U | 0.511 | 0.234 | 1.117 | 0.092 | ||||||||
| IDH | ||||||||||||
| Mut vs. Wt | 0.104 | 0.031 | 0.350 | <0.001 | 5.540 | 1.460 | 21.023 | 0.012 | - | - | - | - |
| Gene expression | ||||||||||||
| SEMA3A | 1.268 | 1.003 | 1.602 | 0.047 | 1.361 | 1.114 | 1.664 | 0.003 | ||||
| SEMA3B | 0.795 | 0.629 | 1.007 | 0.057 | ||||||||
| SEMA3C | 0.781 | 0.591 | 1.033 | 0.083 | ||||||||
| SEMA3D | 0.795 | 0.719 | 0.880 | <0.001 | 0.876 | 0.771 | 0.995 | 0.041 | ||||
| SEMA3E | 1.096 | 0.958 | 1.253 | 0.184 | ||||||||
| SEMA3F | 1.648 | 1.166 | 2.330 | 0.005 | - | - | - | - | ||||
| SEMA3G | 0.796 | 0.676 | 0.938 | 0.006 | - | - | - | - | ||||
| NRP1 | 1.347 | 0.969 | 1.873 | 0.076 | ||||||||
| NRP2 | 0.739 | 0.497 | 1.100 | 0.136 | ||||||||
| PLXNA2 | 0.774 | 0.565 | 1.062 | 0.112 | ||||||||
| PLXND1 | 1.427 | 0.842 | 2.417 | 0.186 | ||||||||
| CDH1 | 0.925 | 0.781 | 1.094 | 0.363 | ||||||||
| CDH2 | 0.741 | 0.460 | 1.192 | 0.217 | ||||||||
| ITGB1 | 0.978 | 0.702 | 1.363 | 0.896 | ||||||||
| ITGB3 | 1.276 | 1.047 | 1.555 | 0.016 | 1.270 | 1.001 | 1.609 | 0.049 | ||||
| ITGA5 | 1.509 | 1.196 | 1.905 | 0.001 | - | - | - | - | ||||
| ITGAV | 0.941 | 0.624 | 1.421 | 0.774 | ||||||||
| VEGFA | 1.354 | 1.168 | 1.571 | <0.001 | - | - | - | - | ||||
| KDR | 1.088 | 0.741 | 1.599 | 0.667 | ||||||||
| The signature | ||||||||||||
| High vs. low risk | 8.308 | 3.109 | 22.206 | <0.001 | 1.274 | 1.055 | 1.539 | 0.012 | ||||
| Variables | Tumor samples | Data from TCGA |
|---|---|---|
| N = 59 (100%) | N = 276 (100%) | |
| Sex | ||
| Female | 26 (44.07) | 116 (42.03) |
| Male | 33 (55.93) | 160 (57.97) |
| Age (years) | ||
| ≤50 | 29 (49.15) | 140 (50.72) |
| >50 | 30 (50.85) | 136 (49.28) |
| Tumor grade | ||
| II | 18 (30.51) | 53 (19.20) |
| III | 6 (10.17) | 111 (40.22) |
| IV (GBM) | 35 (59.32) | 112 (40.58) |
| MGMT | ||
| U | 29 (49.15) | 106 (38.41) |
| M | 30 (50.85) | 170 (61.59) |
| IDH | ||
| Wt | 36 (61.02) | 154 (55.80) |
| Mut | 23 (38.98) | 122 (44.20) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valiulyte, I.; Steponaitis, G.; Kardonaite, D.; Tamasauskas, A.; Kazlauskas, A. A SEMA3 Signaling Pathway-Based Multi-Biomarker for Prediction of Glioma Patient Survival. Int. J. Mol. Sci. 2020, 21, 7396. https://doi.org/10.3390/ijms21197396
Valiulyte I, Steponaitis G, Kardonaite D, Tamasauskas A, Kazlauskas A. A SEMA3 Signaling Pathway-Based Multi-Biomarker for Prediction of Glioma Patient Survival. International Journal of Molecular Sciences. 2020; 21(19):7396. https://doi.org/10.3390/ijms21197396
Chicago/Turabian StyleValiulyte, Indre, Giedrius Steponaitis, Deimante Kardonaite, Arimantas Tamasauskas, and Arunas Kazlauskas. 2020. "A SEMA3 Signaling Pathway-Based Multi-Biomarker for Prediction of Glioma Patient Survival" International Journal of Molecular Sciences 21, no. 19: 7396. https://doi.org/10.3390/ijms21197396
APA StyleValiulyte, I., Steponaitis, G., Kardonaite, D., Tamasauskas, A., & Kazlauskas, A. (2020). A SEMA3 Signaling Pathway-Based Multi-Biomarker for Prediction of Glioma Patient Survival. International Journal of Molecular Sciences, 21(19), 7396. https://doi.org/10.3390/ijms21197396