Comprehensive Analysis of Antibodies Induced by Vaccination with 4 Kinds of Avian Influenza H5N1 Pre-Pandemic Vaccines

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Isolation, from Volunteers Vaccinated with H5N1 Vaccine, of Abs That Bound to H5N1 Virus Particles
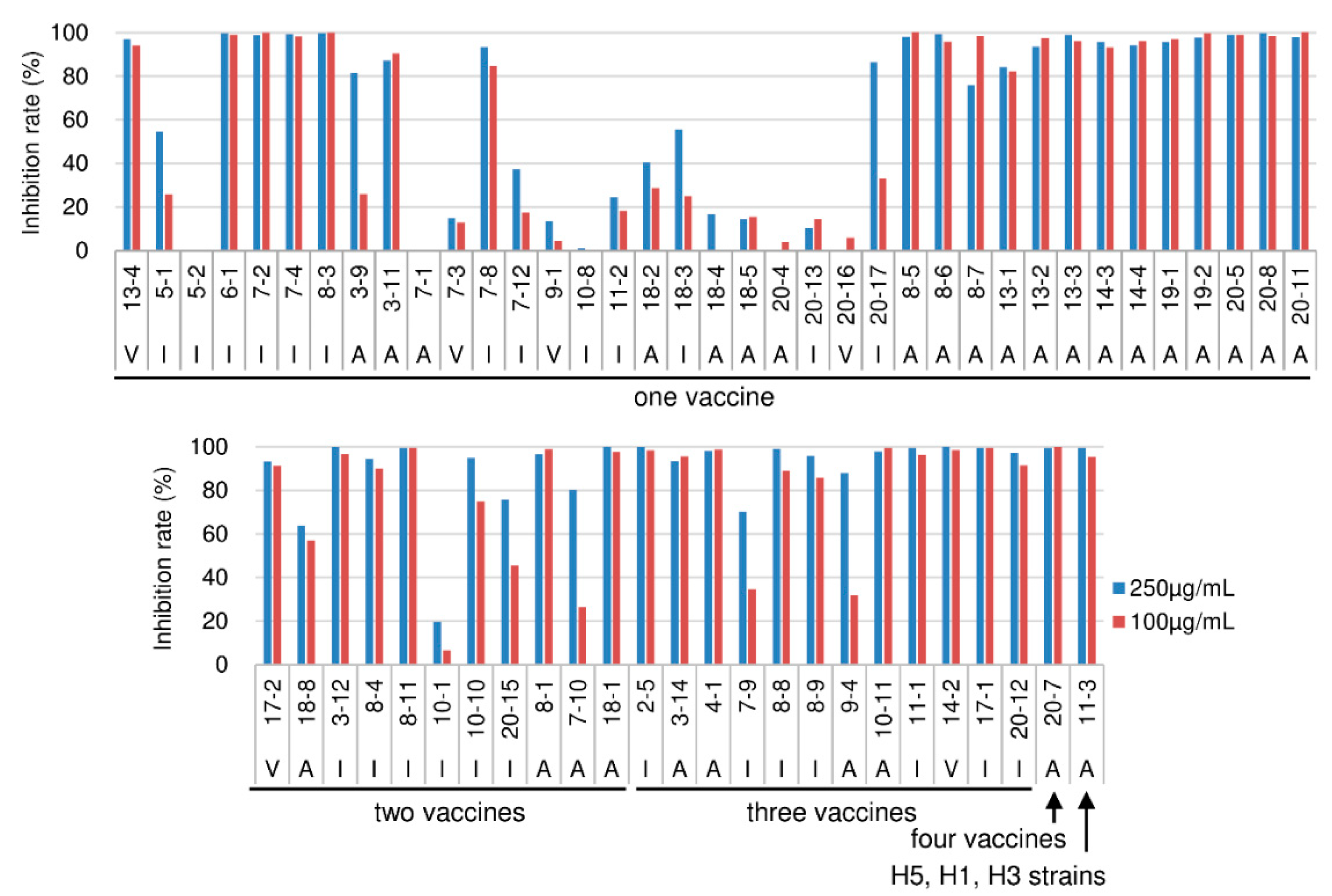
2.2. HI Activities
2.3. Analyses of HI-Positive Clones
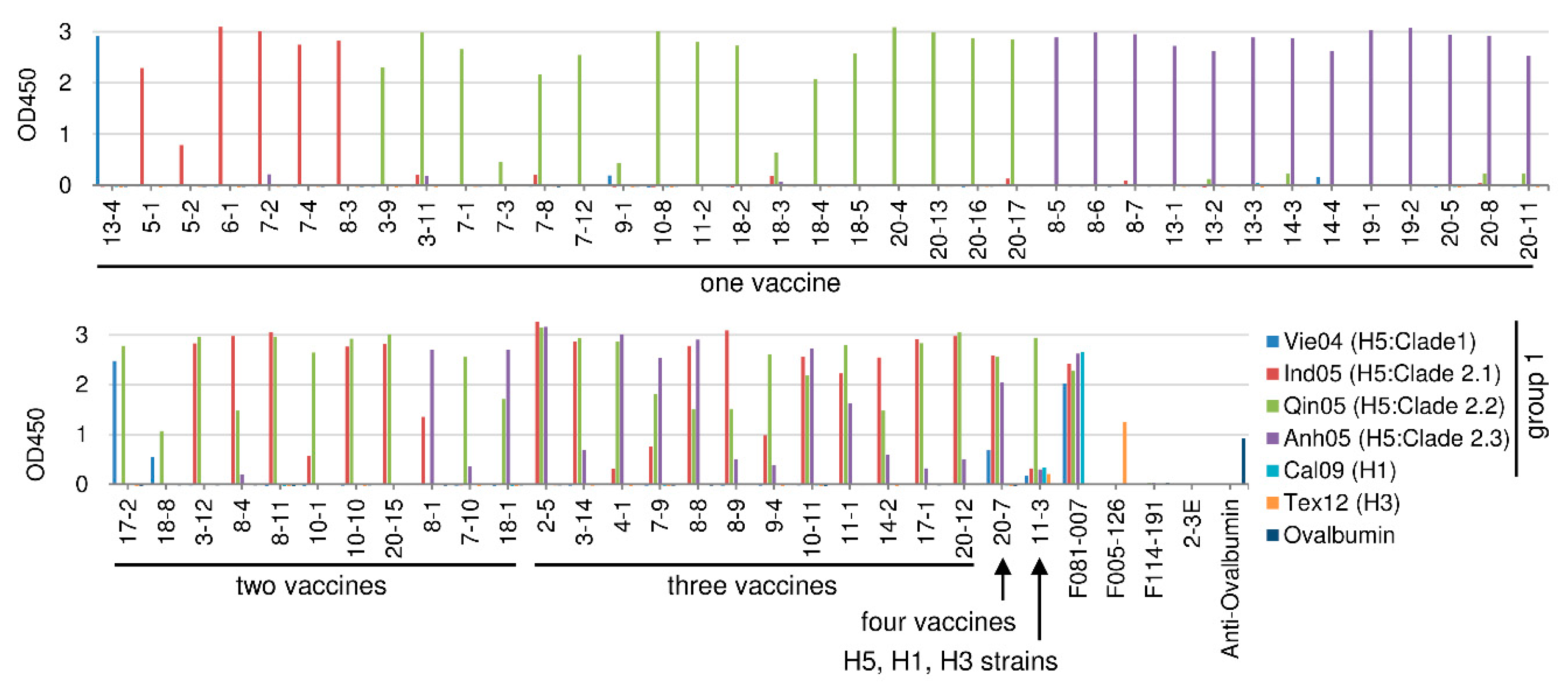
2.3.1. H5N1 Vaccine Strain Binding by the HI-Positive Clones
2.3.2. H5N1 Virus-Neutralizing Activity among the HI-Positive Clones
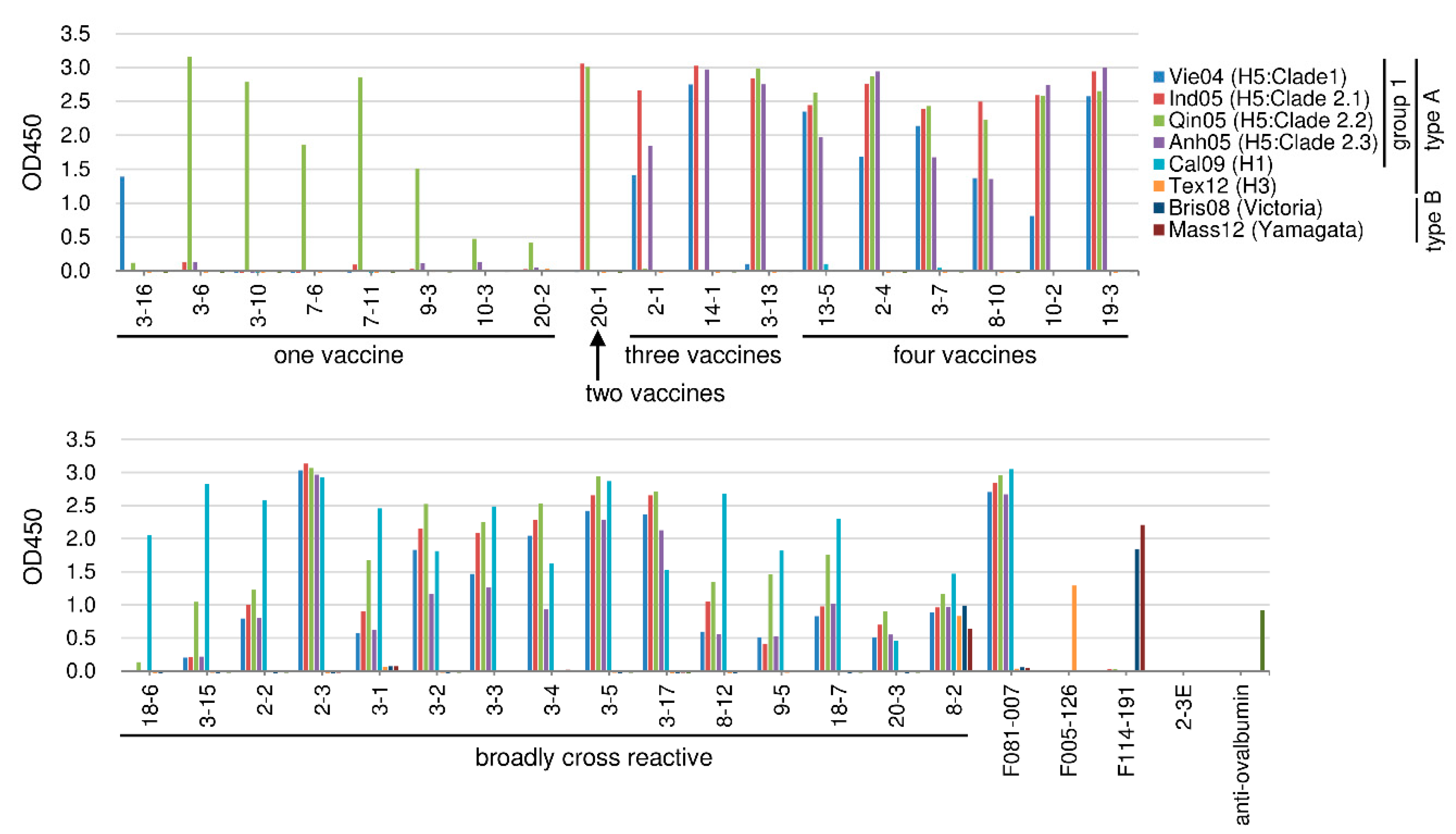
2.3.3. Cross-Reactivity against Recombinant HA Molecule among the HI-Positive Clones
2.4. Analyses of HI-Negative Clones
2.4.1. Binding Activity to H5N1 Vaccine Strains by the HI-Negative Clones
2.4.2. H5N1 Virus-Neutralizing Activity among the HI-Negative Clones
2.4.3. Cross-Reactivity against Recombinant HA Molecule among the HI-Negative Clones
2.5. Summary of Cross-Reactivity of Isolated Clones
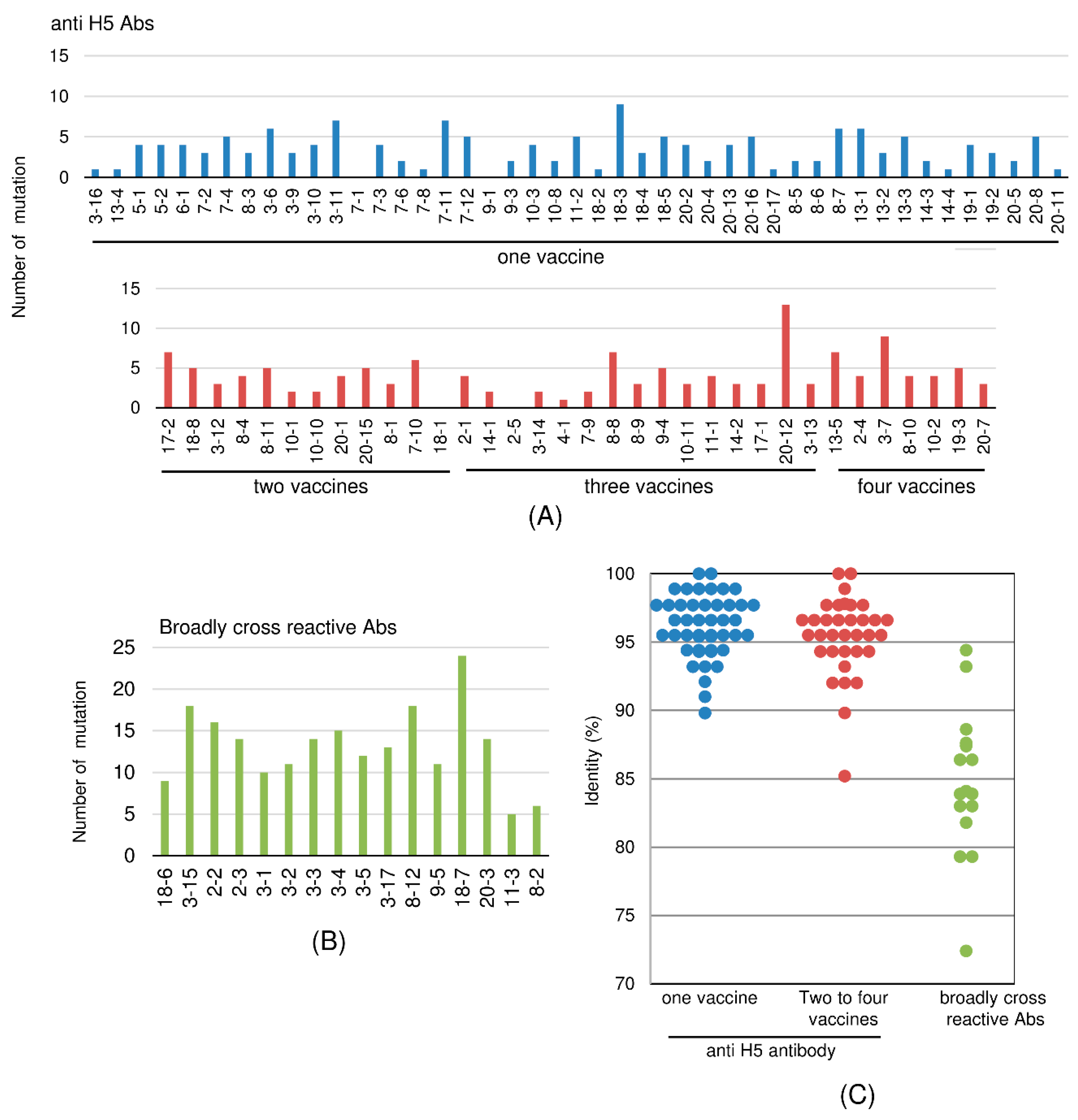
2.6. Sequence-Based Analysis of Frequency of Mutation in VH-Encoding Gene
2.7. Mutation Ratio-Based Evaluation of Cross-Reactive Clones
3. Discussion
4. Materials and Methods
4.1. Viruses
4.2. Ethics Statement
4.3. Construction of Ab-Encoding Libraries
4.4. Screening of the Libraries
4.5. Sequence Analysis
4.6. Preparation of Fab-PP
4.7. HI Assay
4.8. ELISA
4.9. Virus-Neutralizing Activity
4.10. Expression and Purification of Soluble HAs Using Baculovirus Expression Systems
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Abs | antibodies |
| HI | hemagglutination inhibition |
| HPAI | highly pathogenic avian influenza |
| HA | hemagglutinin |
| Vie04 | A/Vietnam/1194/2004 (Clade 1) (H5N1) |
| Ind05 | A/Indonesia/5/2005 (Clade 2.1) (H5N1) |
| Qin05 | A/Qinghai/1A/2005 (Clade 2.2) (H5N1) |
| Anh05 | A/Anhui/1/2005 (Clade 2.3) (H5N1) |
| ELISA | enzyme-linked immunosorbent assay |
| Cal09 | A/California/7/2009 (H1N1) |
| Tex12 | A/Texas/50/2012 (H3N2) |
| Bris08 | B/Brisbane/60/2008 |
| Mass12 | B/Massachusetts/2/2012 |
| LB | Luria–Bertani |
| 2× YT | yeast extract tryptone |
| Fab | antigen-binding fragment |
| IPTG | isopropyl-β-d-thiogalactopyranoside |
| PBS | phosphate-buffered saline |
| HRP | horse radish peroxidase |
| TMB | 3,3’,5,5’-tetramethylbenzidine |
| FFU | focus-forming units |
| MDCK | Madin–Darby canine kidney |
| MEM | modified Eagle’s medium |
| BSA | bovine serum albumin |
| PAP | peroxidase and anti-peroxidase |
| SOC | super optimal broth with catabolites repression |
| X-Gal | 5-bromo-4-chloro-3-indolyl-β-d-galactoside |
| CDR3 | complementarity-determining region 3 |
| Egy13 | A/turkey/Egypt/137/2013 (Clade 2.2.1.2) (H5N1) |
| Vie14 | A/muscovy duck/Vietnam/LBM635/2014 (Clade 2.3.4.4) (H5N1) |
References
- Subbarao, K.; Klimov, A.; Katz, J.; Regnery, H.; Lim, W.; Hall, H.; Perdue, M.; Swayne, D.; Bender, C.; Huang, J.; et al. Characterization of an avian influenza A (H5N1) virus isolated from a child with a fatal respiratory illness. Science 1998, 279, 393–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claas, E.C.; Osterhaus, A.D.; van Beek, R.; De Jong, J.C.; Rimmelzwaan, G.F.; Senne, D.A.; Krauss, S.; Shortridge, K.F.; Webster, R.G. Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet 1998, 351, 472–477. [Google Scholar] [CrossRef]
- Writing Committee of the World Health Organization (WHO). Consultation on Clinical Aspects of Human Infection with Avian Influenza A (H5N1) Virus. Update on avian influenza A (H5N1) virus infection in humans. N. Engl. J. Med. 2008, 358, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization Home Page. Cumulative Number of Confirmed Human Cases of Avian Influenza A/(H5N1) Reported to WHO. Available online: https://www.who.int/influenza/human_animal_interface/H5N1_cumulative_table_archives/en/ (accessed on 25 August 2020).
- Treanor, J.J.; Campbell, J.D.; Kenneth, M.; Zangwill, K.M.; Rowe, T.; Wolff, M. Safety and immunogenicity of an inactivated subvirion influenza A (H5N1) vaccine. N. Engl. J. Med. 2006, 354, 1343–1351. [Google Scholar] [CrossRef]
- Masuda, M.; Sugita, S.; Kuroda, K.; Nishimura, H. H5N1 influenza vaccination policy in Japan. Lancet Infect. Dis. 2009, 9, 266–267. [Google Scholar] [CrossRef]
- Whittle, J.R.R.; Wheatley, A.K.; Wu, L.; Lingwood, D.; Kanekiyo, M.; Ma, S.S.; Narpala, S.R.; Yassine, H.M.; Frank, G.M.; Yewdell, J.W.; et al. Flow cytometry reveals that H5N1 vaccination elicits cross-reactive stem-directed antibodies from multiple Ig heavy-chain lineages. J. Virol. 2014, 88, 4047–4057. [Google Scholar] [CrossRef] [Green Version]
- Nachbagauer, R.; Wohlbold, T.J.; Hirsh, A.; Hai, R.; Sjursen, H.; Palese, P.; Cox, R.J.; Krammer, F. Induction of broadly reactive anti-hemagglutinin stalk antibodies by an H5N1 vaccine in humans. J. Virol. 2014, 88, 13260–13268. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Nachbagauer, R.; Zhu, L.; Huang, Y.; Xie, X.; Jin, S.; Zhang, A.; Wan, Y.; Hirsh, A.; Tian, D.; et al. Induction of broadly cross-reactive stalk-specific antibody responses to influenza group 1 and group 2 hemagglutinins by natural H7N9 virus infection in humans. J. Infect. Dis. 2017, 215, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.G.; Therkelsen, M.D.; Stewart, S.; Kepler, T.B.; Liao, H.X.; Moody, M.A.; Haynes, B.F.; Harrison, S.C. Viral receptor-binding site antibodies with diverse germline origins. Cell 2015, 161, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Dunand, C.J.H.; Leon, P.E.; Kaur, K.; Tan, G.S.; Zheng, N.Y.; Andrews, S.; Huang, M.; Qu, X.; Huang, Y.; Salgado-Ferrer, M.; et al. Preexisting human antibodies neutralize recently emerged H7N9 influenza strains. J. Clin. Invest. 2015, 125, 1255–1268. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.Z.; Holiday, C.; Liu, F.; Jefferson, S.; Gillis, E.; Bellamy, A.R.; Tumpey, T.; Katz, J.M. Cross-reactive antibody responses to novel H5Nx influenza viruses following homologous and heterologous prime-boost vaccination with a prepandemic stockpiled A(H5N1) vaccine in humans. J. Infect. Dis. 2017, 216, S555–S559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marks, J.D.; Hoogenboom, H.R.; Bonnert, T.P.; McCafferty, J.; Griffith, A.D.; Winter, G. By-passing immunization. Human antibodies from V-gene libraries displayed on phage. J. Mol. Biol. 1991, 222, 581–597. [Google Scholar] [CrossRef]
- Stadbauer, D.; Rajabhathor, A.; Amanat, F.; Kaplan, D.; Masud, A.; Treanor, J.J.; Izikson, R.; Cox, M.M.; Nachbagauer, R.; Krammer, F. Vaccination with a recombinant H7 hemagglutinin-based influenza virus vaccine induces broadly reactive antibodies in humans. mSphere 2017, 2, e502–e517. [Google Scholar] [CrossRef] [Green Version]
- Okada, J.; Ohshima, N.; Kubota-Koketsu, R.; Ota, S.; Takase, W.; Azuma, M.; Iba, Y.; Nakagawa, N.; Yoshikawa, T.; Nakajima, Y.; et al. Monoclonal antibodies in man that neutralized H3N2 influenza viruses were classified into three groups with distinct strain specificity: 1968–1973, 1977–1993 and 1997–2003. Virology 2010, 397, 322–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Boutz, D.R.; Chromikova, V.; Joyce, M.G.; Vollmers, C.; Leung, K.; Horton, A.P.; DeKosky, B.J.; Lee, C.H.; Lavinder, J.J.; et al. Molecular-level analysis of the serum antibody repertoire in young adults before and after seasonal influenza vaccination. Nat. Med. 2016, 22, 1456–1464. [Google Scholar] [CrossRef]
- Morino, K.; Katsumi, H.; Akahori, Y.; Iba, Y.; Shinohara, M.; Ukai, Y.; Kohara, Y.; Kurosawa, Y. Antibody fusions with fluorescent proteins: A versatile reagent for profiling protein expression. J. Immunol. Methods 2001, 257, 175–184. [Google Scholar] [CrossRef]
- Sumitomo-Kondo, M.; Ukai, Y.; Iba, Y.; Ohshima, N.; Miura, K.; Takasaki, A.; Kurosawa, Y.; Kurosawa, G. Isolation of human monoclonal antibodies that bind to two different antigens and are encoded by germline VH and VL genes. Biochem. Biophys. Res. Commun. 2018, 503, 1141–1147. [Google Scholar] [CrossRef]
- Iba, Y.; Ito, W.; Kurosawa, Y. Expression vectors for the introduction of highly diverged sequences into the six complementarity-determining regions of an antibody. Gene 1997, 194, 35–46. [Google Scholar] [CrossRef]
- Ohshima, N.; Iba, Y.; Kubota-Koketsu, R.; Asano, Y.; Okuno, Y.; Kurosawa, Y. Naturally occurring antibodies in humans can neutralize a variety of influenza virus strains, including H3, H1, H2 and H5. J. Virol. 2011, 85, 11048–11057. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, N.; Kubota-Koketsu, R.; Iba, Y.; Okuno, Y.; Kurosawa, Y. Two types of antibodies are induced by vaccination with A/California/2009pdm virus: Binding near the sialic acid-binding pocket and neutralizing both H1N1 and H5N1 viruses. PLoS ONE 2014, 9, e87305. [Google Scholar] [CrossRef] [Green Version]
- Hirano, D.; Ohshima, N.; Kubota-Koketsu, R.; Yamasaki, A.; Kurosawa, G.; Okuno, Y.; Yoshida, S.; Kurosawa, Y. Three types of broadly reacting antibodies against influenza B viruses induced by vaccination with seasonal influenza viruses. J. Immunol. Res. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuno, Y.; Tanaka, K.; Baba, K.; Maeda, A.; Kunita, N.; Ueda, S. Rapid focus reduction neutralization test of influenza A and B viruses in microtiter system. J. Clin. Microbiol. 1990, 28, 1308–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laursen, N.S.; Wilson, I.A. Broadly neutralizing antibodies against influenza viruses. Antivir. Res. 2013, 98, 476–483. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO); World Organization for Animal Health (OIE); Food and Agriculture Organization (FAO). Revised and updated nomenclature for highly pathogenic avian influenza A (H5N1) viruses. Influenza Other Respir. Viruses 2014, 8, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.; Brown, J.H.; Cusack, S.; Paulson, J.C.; Skehel, J.J.; Wiley, D.C. Structure of the influenza virus haemagglutinin complexed with its receptor, sialic acid. Nature 1988, 333, 426–431. [Google Scholar] [CrossRef]
- Wu, N.C.; Wilson, I.A. A perspective on the structural and functional constraints for immune evasion: Insights from influenza virus. J. Mol. Biol. 2017, 429, 2694–2709. [Google Scholar] [CrossRef]
- Burke, D.F.; Smith, D.J. A recommended numbering scheme for influenza A subtypes. PLoS ONE 2014, 9, e112302. [Google Scholar] [CrossRef] [Green Version]
- Caton, A.J.; Brownlee, G.G.; Yewdell, J.W.; Gerhard, W. The antigenic structure of the influenza virus A/PR/8/34 hemagglutinin (H1 subtype). Cell 1982, 31, 417–427. [Google Scholar] [CrossRef]
- Gerhard, W.; Yewdell, J.; Frankel, M.E.; Webster, R. Antigenic structure of influenza virus haemagglutinin defined by hybridoma antibodies. Nature 1981, 290, 713–717. [Google Scholar] [CrossRef]
- Throsby, M.; van den Brink, E.; Jongeneelen, M.; Poon, L.L.M.; Alard, P.; Cornelissen, L.; Bakker, A.; Cox, F.; van Deventer, E.; Guan, Y.; et al. Heterosubtypic neutralizing monoclonal antibodies cross-protective against H5N1 and H1N1 recovered from human IgM+ memory B cells. PLoS ONE 2008, 3, e3942. [Google Scholar] [CrossRef] [Green Version]
- Ekiert, D.C.; Bhabha, G.; Elsliger, M.A.; Friesen, R.H.E.; Jongeneelen, M.; Throsby, M.; Goudsmit, J.; Wilson, I.A. Antibody recognition of a highly conserved influenza virus epitope. Science 2009, 324, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, C.J.; Boyington, J.C.; McTamney, P.M.; Kong, W.P.; Pearce, M.B.; Xu, L.; Andersen, H.; Rao, S.; Tumpey, T.M.; Yang, Z.-Y.; et al. Induction of broadly neutralizing H1N1 influenza antibodies by vaccination. Science 2010, 329, 1060–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrammert, J.; Koutsonanos, D.; Li, G.M.; Edupuganti, S.; Sui, J.; Morrissey, M.; McCausland, M.; Skountzou, I.; Hornig, M.; Lipkin, W.I.; et al. Broadly cross-reactive antibodies dominate the human B cell response against 2009 pandemicH1N1 influenza virus infection. J. Exp. Med. 2011, 208, 181–193. [Google Scholar] [CrossRef]
- Sui, J.; Sheehan, J.; Hwang, W.C.; Bankston, L.A.; Burchett, S.K.; Huang, C.-Y.; Liddington, R.C.; Beigel, J.H.; Marasco, W.A. Wide prevalence of heterosubtypic broadly neutralizing human anti-influenza A antibodies. Clin. Infect. Dis. 2011, 52, 1003–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corti, D.; Voss, J.; Gamblin, S.J.; Codoni, G.; Macagno, A.; Jarrossay, D.; Vachieri, S.G.; Pinna, D.; Minola, A.; Vanzetta, F.; et al. A neutralizing antibody selected from plasma cells that binds to group 1 and group 2 influenza A hemagglutinins. Science 2011, 333, 850–856. [Google Scholar] [CrossRef]
- Lee, P.S.; Wilson, I.A. Structural characterization of viral epitopes recognized by broadly cross-reactive antibodies. Curr. Top Microbiol. Immunol. 2015, 386, 323–341. [Google Scholar] [CrossRef] [Green Version]
- Joyce, M.G.; Wheatley, A.K.; Thomas, P.V.; Chuang, G.Y.; Soto, C.; Bailer, R.T.; Druz, A.; Georgiev, I.S.; Gillespie, R.A.; Kanekiyo, M.; et al. Vaccine-induced antibodies that neutralize group 1 and group 2 influenza A viruses. Cell 2016, 166, 609–623. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor | Gender | Age | Vaccination | Number of Mononuclear Cells | Clone Size of Ab Library | Number of Isolated Clones | |
|---|---|---|---|---|---|---|---|
| First | Second | ||||||
| 1 | M | 67 | Vie04 (1) a | Vie04 (1) | 1.86 × 108 | 2.89 × 109 | 72 |
| 2 | M | 72 | Vie04 (1) | Ind05 (2.1) | 1.23 × 108 | 1.73 × 109 | 160 |
| 3 | F | 67 | Vie04 (1) | Qin05 (2.2) | 9.10 × 107 | 1.12 × 109 | 176 |
| 4 | M | 40 | Vie04 (1) | Anh05 (2.3) | 1.23 × 108 | 1.96 × 109 | 133 |
| 5 | M | 54 | Ind05 (2.1) | Vie04 (1) | 1.61 × 108 | 1.30 × 109 | 149 |
| 6 | F | 44 | Ind05 (2.1) | Ind05 (2.1) | 3.30 × 108 | 1.00 × 109 | 65 |
| 7 | M | 29 | Ind05 (2.1) | Qin05 (2.2) | 1.50 × 108 | 5.48 × 109 | 167 |
| 8 | F | 28 | Ind05 (2.1) | Anh05 (2.3) | 9.94 × 107 | 1.03 × 109 | 156 |
| 9 | M | 56 | Qin05 (2.2) | Vie04 (1) | 1.75 × 108 | 2.22 × 109 | 159 |
| 10 | M | 49 | Qin05 (2.2) | Ind05 (2.1) | 8.19 × 107 | 4.76 × 109 | 174 |
| 11 | M | 51 | Qin05 (2.2) | Qin05 (2.2) | 1.73 × 108 | 2.02 × 109 | 61 |
| 13 | M | 31 | Anh05 (2.3) | Vie04 (1) | 1.26 × 108 | 2.88 × 109 | 160 |
| 14 | F | 60 | Anh05 (2.3) | Ind05 (2.1) | 8.67 × 107 | 4.33 × 109 | 137 |
| 16 | M | 33 | Anh05 (2.3) | Anh05 (2.3) | 1.37 × 108 | 1.66 × 109 | 3 |
| 17 | M | 40 | Vie04 (1) | Ind05 (2.1) | 1.44 × 108 | 2.01 × 109 | 144 |
| 18 | M | 41 | Vie04 (1) | Qin05 (2.2) | 6.42 × 107 | 6.33 × 109 | 115 |
| 19 | M | 49 | Ind05 (2.1) | Anh05 (2.3) | 8.94 × 107 | 6.72 × 109 | 200 |
| 20 | M | 40 | Qin05 (2.2) | Anh05 (2.3) | 2.12 × 108 | 3.84 × 109 | 230 |
| Clone | Strain | HI Activity (µg/mL) |
|---|---|---|
| 2-5 | Ind05 (2.1) a | 6.25 |
| 3-9 | Qin05 (2.2) | 100 |
| 3-11 | Qin05 (2.2) | 3.13 |
| 3-12 | Qin05 (2.2) | 12.5 |
| 3-14 | Qin05 (2.2) | 25 |
| 4-1 | Anh05 (2.3) | 0.78 |
| 5-1 | Ind05 (2.1) | 6.25 |
| 5-2 | Ind05 (2.1) | 12.5 |
| 6-1 | Ind05 (2.1) | 12.5 |
| 7-1 | Qin05 (2.2) | 6.25 |
| 7-2 | Ind05 (2.1) | 12.5 |
| 7-3 | Qin05 (2.2) | 50 |
| 7-4 | Ind05 (2.1) | 50 |
| 7-8 | Ind05 (2.1) | 50 |
| 7-9 | Ind05 (2.1) | 50 |
| 7-10 | Qin05 (2.2) | 100 |
| 7-12 | Qin05 (2.2) | 6.25 |
| 8-1 | Anh05 (2.3) | 25 |
| 8-3 | Ind05 (2.1) | 6.25 |
| 8-4 | Ind05 (2.1) | 12.5 |
| 8-5 | Anh05 (2.3) | 12.5 |
| 8-6 | Anh05 (2.3) | 12.5 |
| 8-7 | Anh05 (2.3) | 12.5 |
| 8-8 | Ind05 (2.1) | 6.25 |
| 8-9 | Ind05 (2.1) | 3.13 |
| 8-11 | Ind05 (2.1) | 12.5 |
| 9-1 | Qin05 (2.2) | 50 |
| 9-4 | Qin05 (2.2) | 6.25 |
| 10-1 | Qin05 (2.2) | 25 |
| 10-8 | Qin05 (2.2) | 50 |
| 10-10 | Qin05 (2.2) | 25 |
| 10-11 | Qin05 (2.2) | 12.5 |
| 11-1 | Qin05 (2.2) | 6.25 |
| 11-2 | Qin05 (2.2) | 12.5 |
| 11-3 | Qin05 (2.2) | 6.25 |
| 13-1 | Anh05 (2.3) | 50 |
| 13-2 | Anh05 (2.3) | 3.13 |
| 13-3 | Anh05 (2.3) | 6.25 |
| 13-4 | Vie04 (1) | 12.5 |
| 14-2 | Ind05 (2.1) | 3.13 |
| 14-3 | Anh05 (2.3) | 25 |
| 14-4 | Anh05 (2.3) | 12.5 |
| 17-1 | Ind05 (2.1) | 1.56 |
| 17-2 | Vie04 (1) | 6.25 |
| 18-1 | Qin05 (2.2) | 25 |
| 18-2 | Qin05 (2.2) | 25 |
| 18-3 | Qin05 (2.2) | 25 |
| 18-4 | Qin05 (2.2) | 100 |
| 18-5 | Qin05 (2.2) | 25 |
| 18-8 | Qin05 (2.2) | 12.5 |
| 19-1 | Anh05 (2.3) | 6.25 |
| 19-2 | Anh05 (2.3) | 12.5 |
| 20-4 | Qin05 (2.2) | 12.5 |
| 20-5 | Anh05 (2.3) | 12.5 |
| 20-7 | Qin05 (2.2) | 100 |
| 20-8 | Anh05 (2.3) | 6.25 |
| 20-11 | Anh05 (2.3) | 6.25 |
| 20-12 | Qin05 (2.2) | 3.13 |
| 20-13 | Qin05 (2.2) | 25 |
| 20-15 | Qin05 (2.2) | 50 |
| 20-16 | Qin05 (2.2) | 12.5 |
| 20-17 | Qin05 (2.2) | 50 |
| Clone | Germline | Identity (%) |
|---|---|---|
| 2-5 | 4-34*01 | 100.0 |
| 3-9 | 4-4*02 | 96.6 |
| 3-11 | 4-31*03 | 91.0 |
| 3-12 | 4-34*01 | 96.6 |
| 3-14 | 4-34*01 | 97.7 |
| 4-1 | 3-9*01 | 98.9 |
| 5-1 | 3-9*01 | 95.5 |
| 5-2 | 3-9*01 | 95.5 |
| 6-1 | 4-34*01 | 95.4 |
| 7-1 | 3-9*01 | 100.0 |
| 7-2 | 3-20*01 | 96.6 |
| 7-3 | 3-21*01 | 95.5 |
| 7-4 | 3-23*01 | 94.3 |
| 7-8 | 4-34*01 | 98.9 |
| 7-9 | 4-34*01 | 97.7 |
| 7-10 | 4-39*01 | 93.2 |
| 7-12 | 4-39*01 | 94.4 |
| 8-1 | 1-18*01 | 96.6 |
| 8-3 | 3-9*01 | 96.6 |
| 8-4 | 3-9*01 | 95.5 |
| 8-5 | 3-9*01 | 97.7 |
| 8-6 | 3-9*01 | 97.7 |
| 8-7 | 3-21*01 | 93.2 |
| 8-8 | 3-33*01 | 92.0 |
| 8-9 | 3-43*01 | 96.6 |
| 8-11 | 4-34*01 | 94.3 |
| 9-1 | 3-9*01 | 100.0 |
| 9-4 | 3-66*01 | 94.3 |
| 10-1 | 1-8*01 | 97.7 |
| 10-8 | 3-33*01 | 97.7 |
| 10-10 | 4-39*01 | 97.8 |
| 10-11 | 4-34*01 | 96.6 |
| 11-1 | 1-18*04 | 95.5 |
| 11-2 | 4-30-2*01 | 94.4 |
| 11-3 | 4-31*03 | 94.4 |
| 13-1 | 1-8*01 | 93.2 |
| 13-2 | 1-18*01 | 96.6 |
| 13-3 | 3-9*01 | 94.3 |
| 13-4 | 3-33*06 | 98.9 |
| 14-2 | 3-30*04 | 96.6 |
| 14-3 | 4-34*01 | 97.7 |
| 14-4 | 4-4*02 | 98.9 |
| 17-1 | 4-34*01 | 96.6 |
| 17-2 | 4-59*01 | 92.0 |
| 18-1 | 3-9*01 | 100.0 |
| 18-2 | 3-11*01 | 98.9 |
| 18-3 | 3-33*06 | 89.8 |
| 18-4 | 4-31*03 | 96.6 |
| 18-5 | 4-31*03 | 94.4 |
| 18-8 | 5-51*01 | 94.3 |
| 19-1 | 3-7*01 | 95.5 |
| 19-2 | 3-20*01 | 96.6 |
| 20-4 | 3-9*01 | 97.7 |
| 20-5 | 3-9*01 | 97.7 |
| 20-7 | 3-9*01 | 96.6 |
| 20-8 | 3-9*01 | 95.5 |
| 20-11 | 3-20*01 | 97.7 |
| 20-12 | 3-33*01 | 85.2 |
| 20-13 | 3-53*01 | 95.4 |
| 20-15 | 4-34*01 | 94.3 |
| 20-16 | 4-39*01 | 94.4 |
| 20-17 | 4-59*01 | 98.9 |
| Clone | Germline | Identity (%) |
|---|---|---|
| 2-1 | 1-69*01 | 95.5 |
| 2-2 | 1-69*01 | 81.8 |
| 2-3 | 1-69*01 | 83.9 |
| 2-4 | 3-30-3*01 | 95.5 |
| 3-1 | 1-2*01 | 88.6 |
| 3-2 | 1-69*01 | 86.4 |
| 3-3 | 1-69*01 | 84.1 |
| 3-4 | 1-69*01 | 83.0 |
| 3-5 | 1-69*01 | 86.4 |
| 3-6 | 1-69*04 | 93.2 |
| 3-7 | 1-69*06 | 85.2 |
| 3-10 | 4-4*02 | 95.5 |
| 3-13 | 4-34*01 | 96.6 |
| 3-15 | 4-59*01 | 79.3 |
| 3-16 | 5-51*01 | 98.9 |
| 3-17 | 5-51*01 | 83.0 |
| 7-6 | 4-4*02 | 97.7 |
| 7-11 | 4-39*01 | 92.1 |
| 8-2 | 3-7*02 | 93.2 |
| 8-10 | 4-4*02 | 95.5 |
| 8-12 | 4-59*01 | 79.3 |
| 9-3 | 3-30*01 | 97.7 |
| 9-5 | 4-4*08 | 87.4 |
| 10-2 | 1-46*03 | 95.5 |
| 10-3 | 3-9*01 | 95.5 |
| 13-5 | 4-34*01 | 92.0 |
| 14-1 | 3-21*01 | 97.7 |
| 18-6 | 4-39*01 | 87.6 |
| 18-7 | 4-59*01 | 72.4 |
| 19-3 | 3-21*01 | 94.3 |
| 20-1 | 1-2*02 | 95.5 |
| 20-2 | 1-18*01 | 95.5 |
| 20-3 | 1-69*01 | 83.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohshima, N.; Iba, Y.; Kubota-Koketsu, R.; Yamasaki, A.; Majima, K.; Kurosawa, G.; Hirano, D.; Yoshida, S.; Sugiura, M.; Asano, Y.; et al. Comprehensive Analysis of Antibodies Induced by Vaccination with 4 Kinds of Avian Influenza H5N1 Pre-Pandemic Vaccines. Int. J. Mol. Sci. 2020, 21, 7422. https://doi.org/10.3390/ijms21197422
Ohshima N, Iba Y, Kubota-Koketsu R, Yamasaki A, Majima K, Kurosawa G, Hirano D, Yoshida S, Sugiura M, Asano Y, et al. Comprehensive Analysis of Antibodies Induced by Vaccination with 4 Kinds of Avian Influenza H5N1 Pre-Pandemic Vaccines. International Journal of Molecular Sciences. 2020; 21(19):7422. https://doi.org/10.3390/ijms21197422
Chicago/Turabian StyleOhshima, Nobuko, Yoshitaka Iba, Ritsuko Kubota-Koketsu, Ayami Yamasaki, Keiko Majima, Gene Kurosawa, Daisuke Hirano, Shunji Yoshida, Mototaka Sugiura, Yoshizo Asano, and et al. 2020. "Comprehensive Analysis of Antibodies Induced by Vaccination with 4 Kinds of Avian Influenza H5N1 Pre-Pandemic Vaccines" International Journal of Molecular Sciences 21, no. 19: 7422. https://doi.org/10.3390/ijms21197422
APA StyleOhshima, N., Iba, Y., Kubota-Koketsu, R., Yamasaki, A., Majima, K., Kurosawa, G., Hirano, D., Yoshida, S., Sugiura, M., Asano, Y., Okuno, Y., & Kurosawa, Y. (2020). Comprehensive Analysis of Antibodies Induced by Vaccination with 4 Kinds of Avian Influenza H5N1 Pre-Pandemic Vaccines. International Journal of Molecular Sciences, 21(19), 7422. https://doi.org/10.3390/ijms21197422