Low-Cost Cellulase-Hemicellulase Mixture Secreted by Trichoderma harzianum EM0925 with Complete Saccharification Efficacy of Lignocellulose

Abstract
:1. Introduction
2. Results
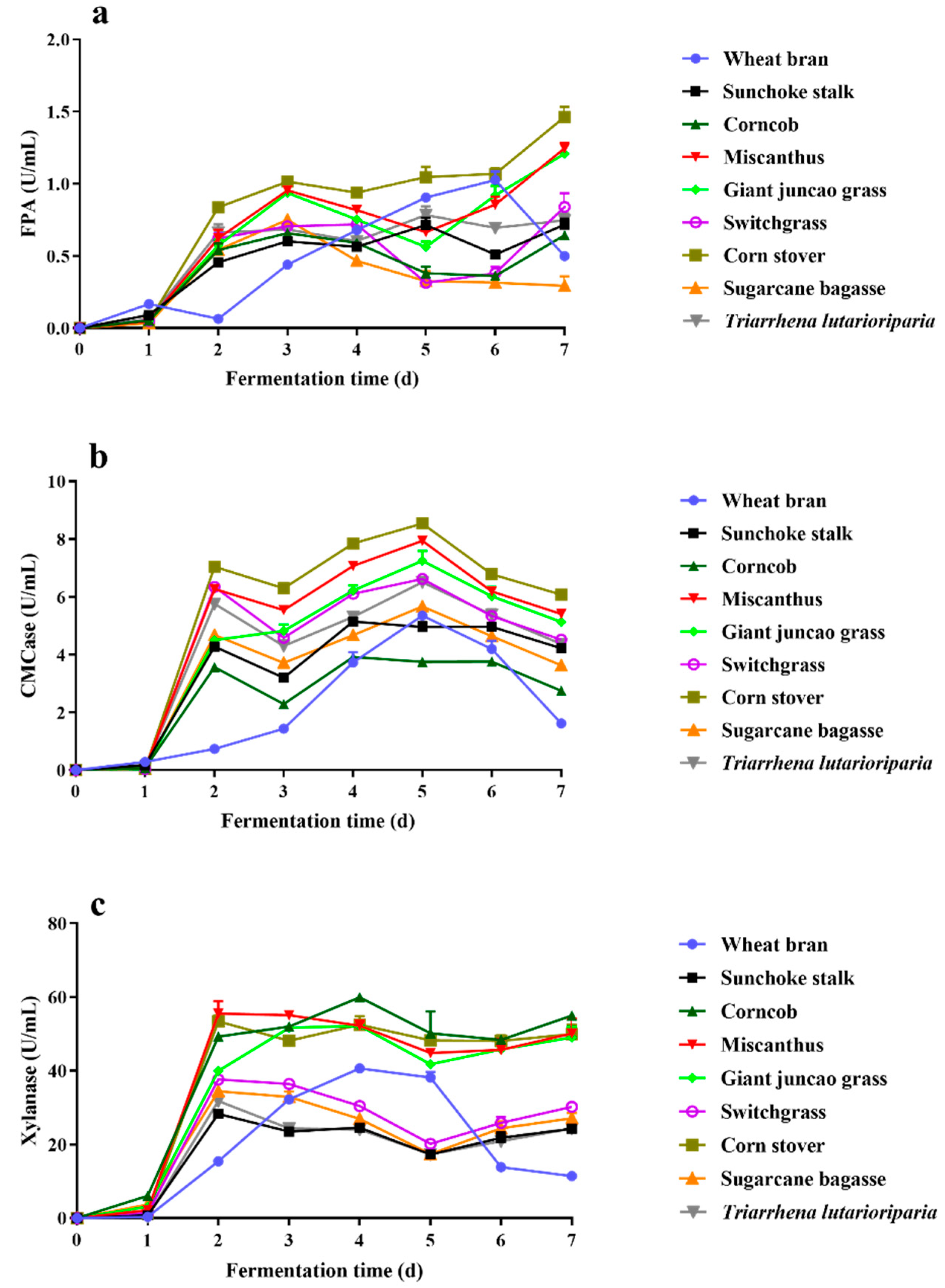
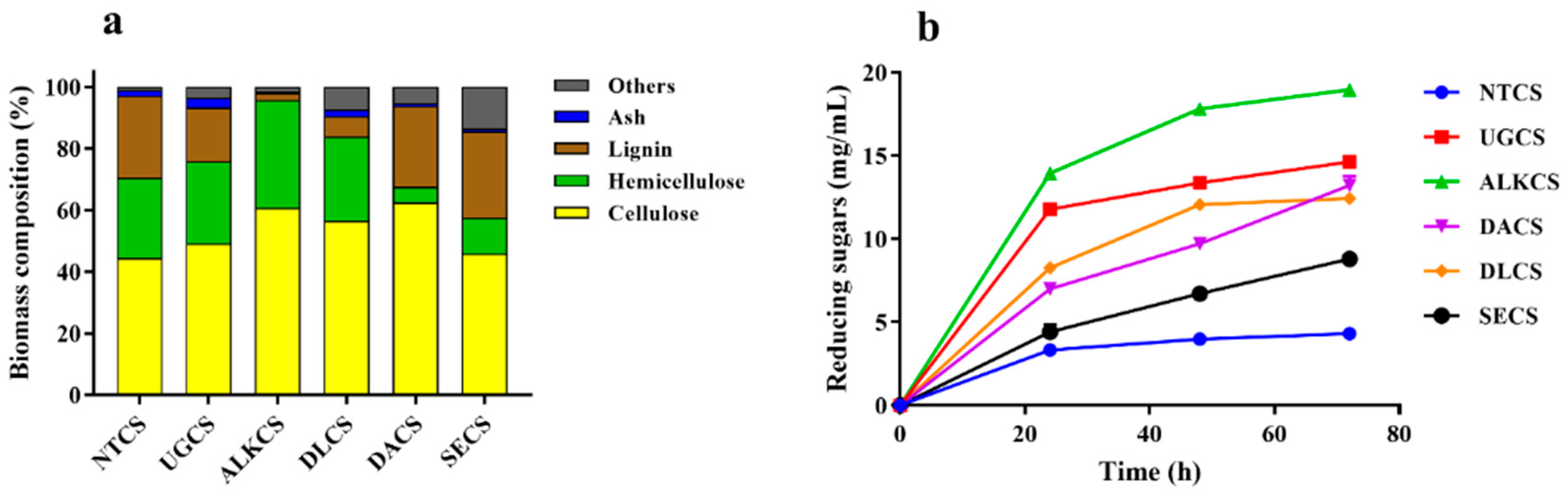
2.1. Substrate Selection for Lignocellulosic Enzyme Production by T. Harzianum EM0925
2.2. Enzyme Activities of Enzyme Cocktail EM0925
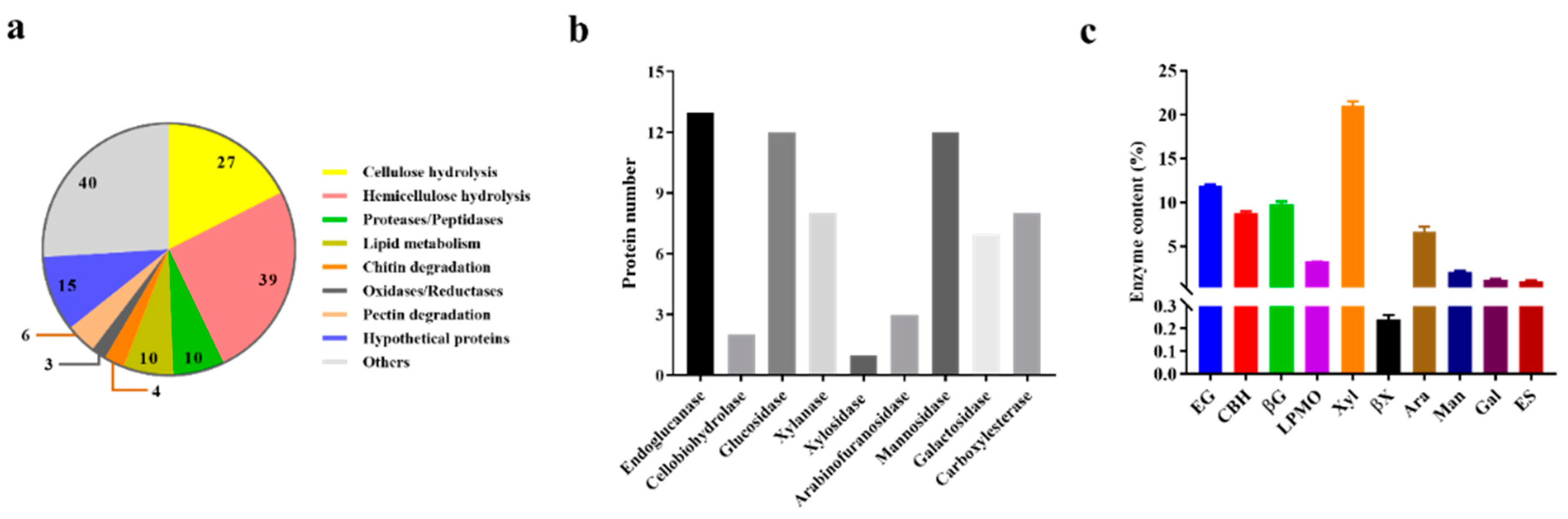
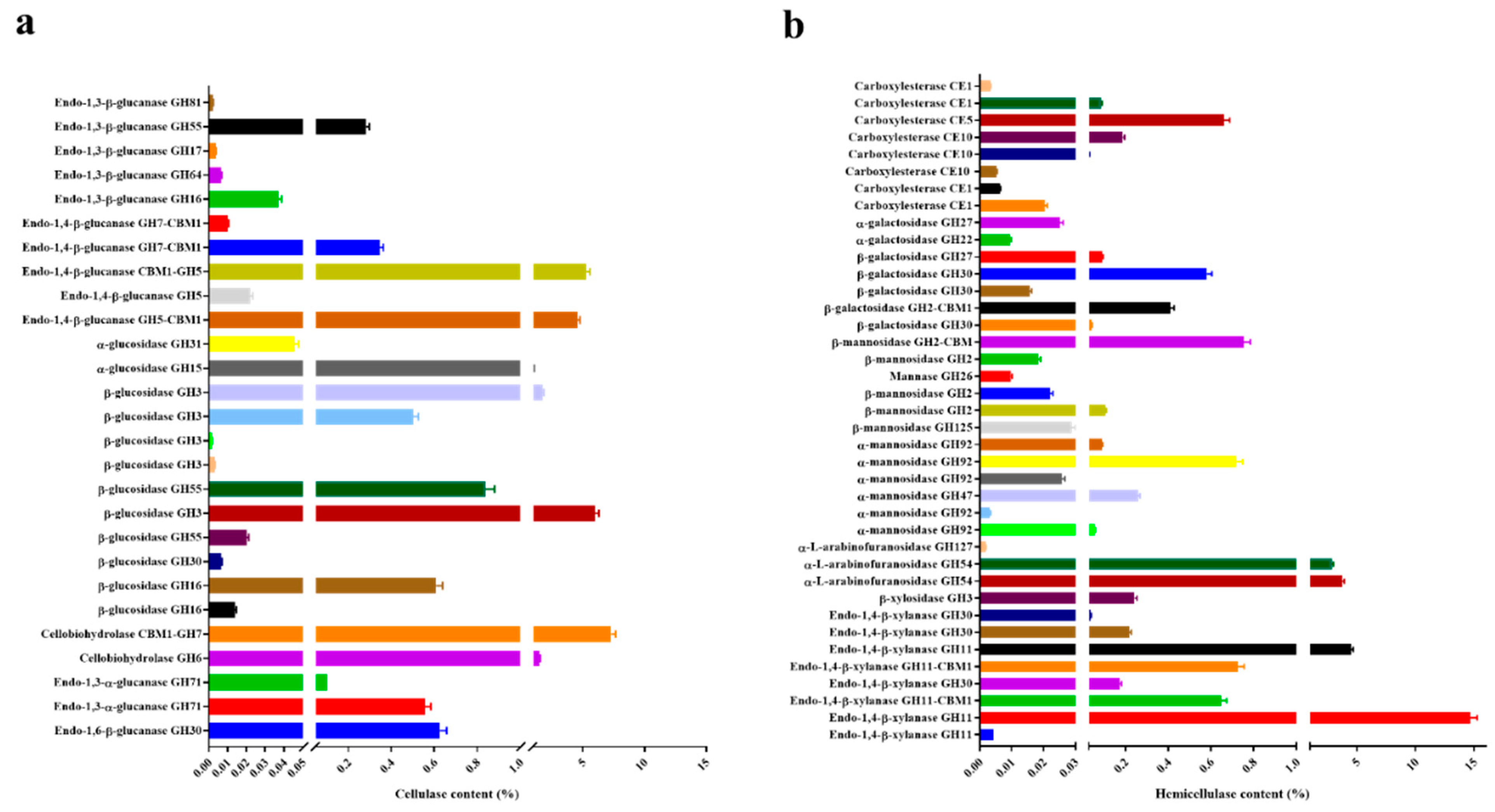
2.3. Extracellular Enzymes in the Proteome of T. harzianum EM0925
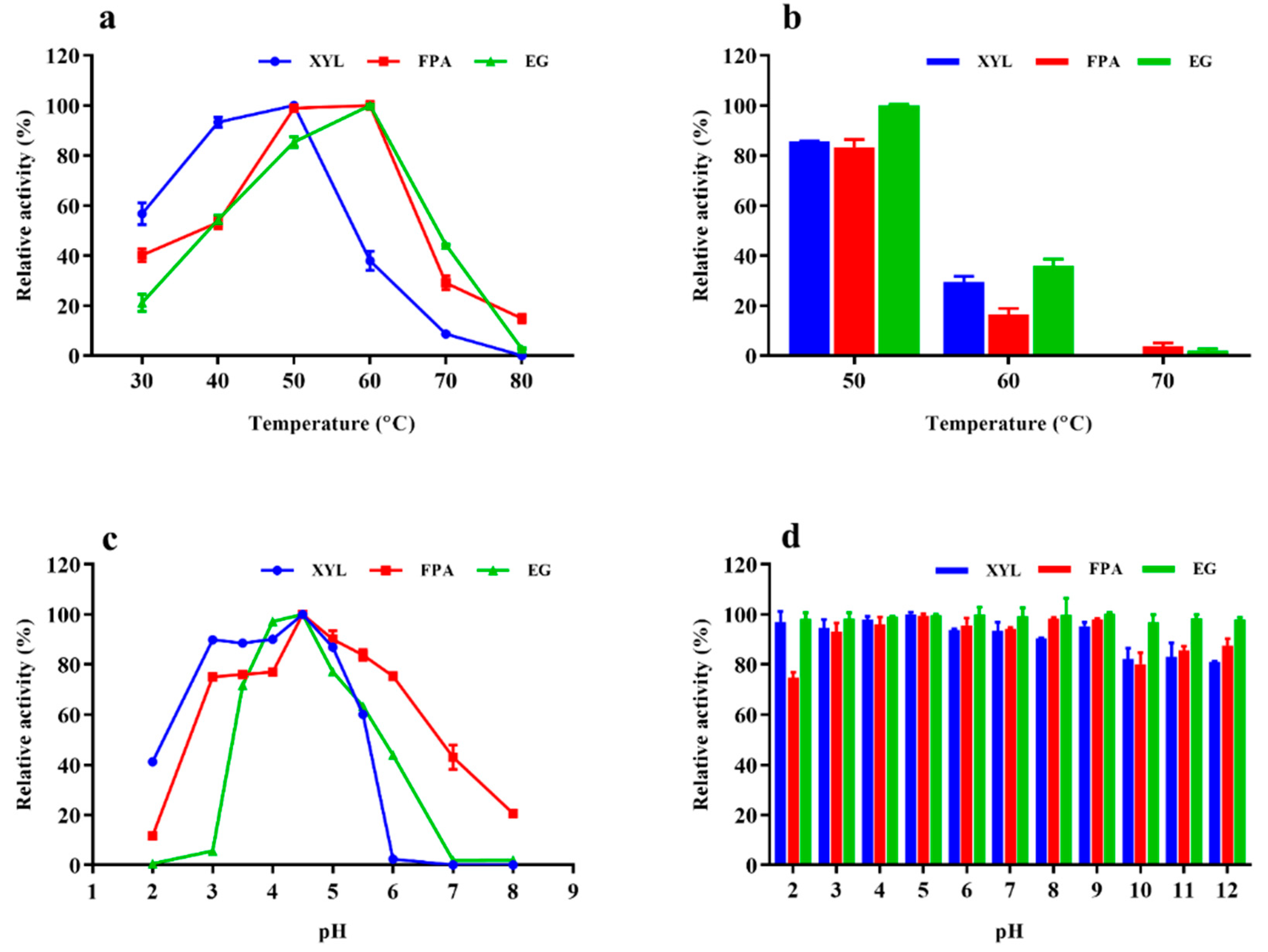
2.4. Properties of Enzyme Cocktail EM0925
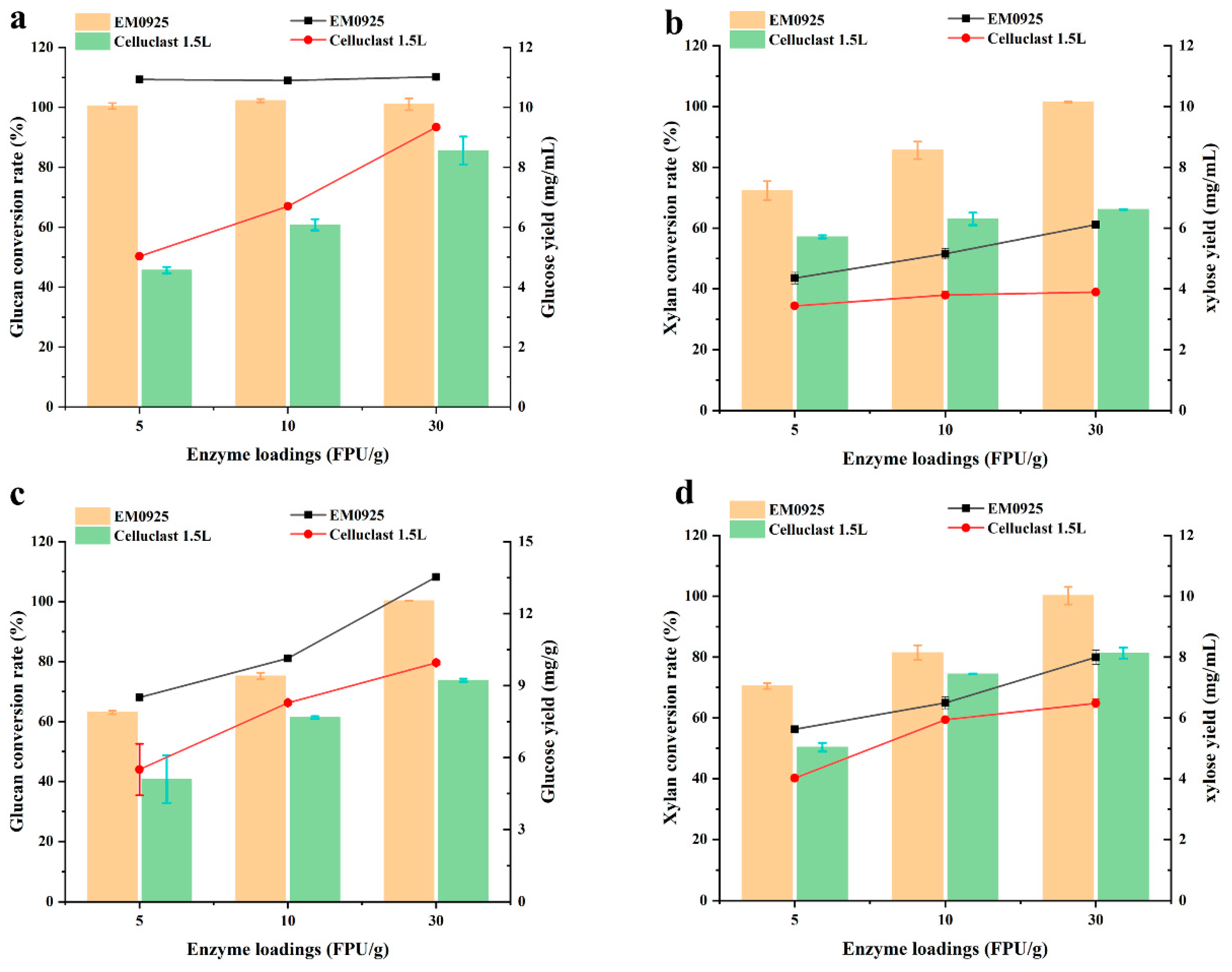
2.5. Hydrolysis of Lignocellulosic Biomass by Enzyme Cocktail EM0925
3. Discussion
4. Materials and Methods
4.1. Strain and Culture Conditions
4.2. Preparation of Enzyme Cocktail EM0925
4.3. Enzymatic Assays
4.4. Biomass Pretreatment and Component Analysis
4.5. Enzymatic Hydrolysis
4.6. Extracellular Quantitative Proteome Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CAZy | Carbohydrate active enzymes database |
| CBH | Cellobiohydrolase |
| CBM | Carbohydrate-binding module |
| EG | Endoglucanase |
| GH | Glycoside hydrolase |
| LPMO | Lytic polysaccharide monooxygenase |
| HPLC | High-performance liquid chromatography |
| FPA | Filter paper activity |
| CMC | Carboxymethylcellulose |
| βG | β-glucosidase |
| Xyl | Xylanase |
| βX | β-xylosidase |
| Ara | Arabinofuranosidase |
| Man | Mannase |
| Amy | Amylase |
| LC-MS/MS | Liquid chromatography-tandem mass spectrometry |
| NTCS | Non-pretreated corn stover |
| UGCS | Ultrafine grinding pretreated corn stover |
| ALKCS | Alkali pretreated corn stover |
| DACS | Dilute acid pretreated corn stover |
| DLCS | Sodium chlorite/acetic acid delignificated corn stover |
| SECS | Steam explosion pretreated corn stover |
| pNPC | p-nitrophenyl-β-d-cellobioside |
| pNPG | p-nitrophenyl-β-d-glucoside |
| pNPX | p-nitrophenyl-β-d-xyloside |
| pNPAF | p-nitrophenyl-α-l-arabinofuranoside |
| iBAQ | Intensity-based absolute quantification |
| LFQ | Label free quantification |
References
- Wang, J.; Chen, X.; Chio, C.; Yang, C.; Su, E.; Jin, Y.; Cao, F.; Qin, W. Delignification overmatches hemicellulose removal for improving hydrolysis of wheat straw using the enzyme cocktail from Aspergillus niger. Bioresour. Technol. 2019, 274, 459–467. [Google Scholar] [CrossRef]
- Paz, A.; Outeirino, D.; Perez Guerra, N.; Dominguez, J.M. Enzymatic hydrolysis of brewer’s spent grain to obtain fermentable sugars. Bioresour. Technol. 2018, 275, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Xin, F.; Dong, W.; Zhang, W.; Ma, J.; Jiang, M. Biobutanol production from crystalline cellulose through consolidated bioprocessing. Trends Biotechnol. 2019, 37, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Kun, R.S.; Gomes, A.C.S.; Hilden, K.S.; Cerezo, S.S.; Makela, M.R.; de Vries, R.P. Developments and opportunities in fungal strain engineering for the production of novel enzymes and enzyme cocktails for plant biomass degradation. Biotechnol. Adv. 2019, 37, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Castro, A.M.; Pedro, K.C.; da Cruz, J.C.; Ferreira, M.C.; Leite, S.G.; Pereira, N., Jr. Trichoderma harzianum IOC-4038: A promising strain for the production of a cellulolytic complex with significant beta-glucosidase activity from sugarcane bagasse cellulignin. Appl. Biochem. Biotechnol. 2010, 162, 2111–2122. [Google Scholar] [CrossRef]
- Gama, R.; Van Dyk, J.S.; Pletschke, B.I. Optimisation of enzymatic hydrolysis of apple pomace for production of biofuel and biorefinery chemicals using commercial enzymes. 3 Biotech 2015, 5, 1075–1087. [Google Scholar] [CrossRef] [Green Version]
- De Paula, R.G.; Antonieto, A.C.C.; Nogueira, K.M.V.; Ribeiro, L.F.C.; Rocha, M.C.; Malavazi, I.; Almeida, F.; Silva, R.N. Extracellular vesicles carry cellulases in the industrial fungus Trichoderma reesei. Biotechnol. Biofuels 2019, 12, 146. [Google Scholar] [CrossRef]
- Jiang, Y.; Wu, R.; Zhou, J.; He, A.; Xu, J.; Xin, F.; Zhang, W.; Ma, J.; Jiang, M.; Dong, W. Recent advances of biofuels and biochemicals production from sustainable resources using co-cultivation systems. Biotechnol. Biofuels 2019, 12, 155. [Google Scholar] [CrossRef] [Green Version]
- Sanhueza, C.; Carvajal, G.; Soto-Aguilar, J.; Lienqueo, M.E.; Salazar, O. The effect of a lytic polysaccharide monooxygenase and a xylanase from Gloeophyllum trabeum on the enzymatic hydrolysis of lignocellulosic residues using a commercial cellulase. Enzym. Microb. Technol. 2018, 113, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Goldbeck, R.; Gonçalves, T.A.; Damásio, A.R.L.; Brenelli, L.B.; Wolf, L.D.; Paixão, D.A.A.; Rocha, G.J.M.; Squina, F.M. Effect of hemicellulolytic enzymes to improve sugarcane bagasse saccharification and xylooligosaccharides production. J. Mol. Catal. B Enzym. 2016, 131, 36–46. [Google Scholar] [CrossRef]
- Sun, F.F.; Hong, J.; Hu, J.; Saddler, J.N.; Fang, X.; Zhang, Z.; Shen, S. Accessory enzymes influence cellulase hydrolysis of the model substrate and the realistic lignocellulosic biomass. Enzym. Microb. Technol. 2015, 79–80, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, F.; Li, J.; Zhang, Z.; Bai, F.; Chen, J.; Zhao, X. Rapid production of lignocellulolytic enzymes by Trichoderma harzianum LZ117 isolated from Tibet for biomass degradation. Bioresour. Technol. 2019, 292, 122063. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Qin, Y.; Li, Z.; Qu, Y. Improving lignocellulolytic enzyme production with Penicillium: From strain screening to systems biology. Biofuels 2014, 4, 523–534. [Google Scholar] [CrossRef] [Green Version]
- Gusakov, A.V.; Sinitsyn, A.P. Cellulases from Penicillium species for producing fuels from biomass. Biofuels 2014, 3, 463–477. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, J.; Liu, J.; Wang, R.; Liu, L.; Wang, F.; Yuan, H. The composition of accessory enzymes of Penicillium chrysogenum P33 revealed by secretome and synergistic effects with commercial cellulase on lignocellulose hydrolysis. Bioresour. Technol. 2018, 257, 54–61. [Google Scholar] [CrossRef]
- Berlin, A.; Maximenko, V.; Gilkes, N.; Saddler, J. Optimization of enzyme complexes for lignocellulose hydrolysis. Biotechnol. Bioeng. 2007, 97, 287–296. [Google Scholar] [CrossRef]
- Berrin, J.G.; Herpoel-Gimbert, I.; Ferreira, N.L.; Margeot, A.; Heiss-Blanquet, S. Use of cellulases from Trichoderma reesei in the Twenty-First Century-Part II: Optimization of cellulolytic cocktails for saccharification of lignocellulosic feedstocks. In Biotechnology and Biology of Trichoderma; Eisevier: Amsterdam, The Netherlands, 2014; pp. 263–280. [Google Scholar]
- Chundawat, S.P.; Lipton, M.S.; Purvine, S.O.; Uppugundla, N.; Gao, D.; Balan, V.; Dale, B.E. Proteomics-based compositional analysis of complex cellulase-hemicellulase mixtures. J. Proteome Res. 2011, 10, 4365–4372. [Google Scholar] [CrossRef]
- De Castro, A.M.; Ferreira, M.C.; da Cruz, J.C.; Pedro, K.C.; Carvalho, D.F.; Leite, S.G.; Pereira, N. High-Yield Endoglucanase Production by Trichoderma harzianum IOC-3844 cultivated in pretreated sugarcane mill byproduct. Enzym. Res. 2010, 2010, 854526. [Google Scholar] [CrossRef] [Green Version]
- Ferreira Filho, J.A.; Horta, M.A.C.; Beloti, L.L.; Dos Santos, C.A.; de Souza, A.P. Carbohydrate-active enzymes in Trichoderma harzianum: A bioinformatic analysis bioprospecting for key enzymes for the biofuels industry. BMC Genom. 2017, 18, 779. [Google Scholar] [CrossRef] [Green Version]
- Benoliel, B.; Torres, F.A.G.; de Moraes, L.M.P. A novel promising Trichoderma harzianum strain for the production of a cellulolytic complex using sugarcane bagasse in natura. Springer Plus 2013, 2, 656. [Google Scholar] [CrossRef] [Green Version]
- Delabona Pda, S.; Farinas, C.S.; da Silva, M.R.; Azzoni, S.F.; Pradella, J.G. Use of a new Trichoderma harzianum strain isolated from the Amazon rainforest with pretreated sugar cane bagasse for on-site cellulase production. Bioresour. Technol. 2012, 107, 517–521. [Google Scholar] [CrossRef]
- Zhao, X.; Xiong, L.; Zhang, M.; Bai, F. Towards efficient bioethanol production from agricultural and forestry residues: Exploration of unique natural microorganisms in combination with advanced strain engineering. Bioresour. Technol. 2016, 215, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xie, X.; Liu, J.; Wu, D.; Cai, G.; Lu, J. Characterization of a putative glycoside hydrolase family 43 arabinofuranosidase from Aspergillus niger and its potential use in beer production. Food Chem. 2020, 305, 125382. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Arantes, V.; Pribowo, A.; Gourlay, K.; Saddler, J.N. Substrate factors that influence the synergistic interaction of AA9 and cellulases during the enzymatic hydrolysis of biomass. Energy Environ. Sci. 2014, 7, 2308–2315. [Google Scholar] [CrossRef]
- Gusakov, A.V. Alternatives to Trichoderma reesei in biofuel production. Trends Biotechnol. 2011, 29, 419–425. [Google Scholar] [CrossRef]
- De Souza, M.F.; da Silva, A.S.A.; Bon, E.P.S. A novel Trichoderma harzianum strain from the Amazon Forest with high cellulolytic capacity. Biocatal. Agric. Biotechnol. 2018, 14, 183–188. [Google Scholar] [CrossRef]
- Li, C.; Pang, A.P.; Yang, H.; Lv, R.; Zhou, Z.; Wu, F.G.; Lin, F. Tracking localization and secretion of cellulase spatiotemporally and directly in living Trichoderma reesei. Biotechnol. Biofuels 2019, 12, 200. [Google Scholar] [CrossRef] [PubMed]
- Boussaid, A.; Robinson, J.; Cai, Y.; Gregg, D.J.; Saddler, J.N. Fermentability of the hemicellulose-derived sugars from steam-exploded softwood (douglas fir). Biotechnol. Bioeng. 1999, 64, 284–289. [Google Scholar] [CrossRef]
- Li, C.; Knierim, B.; Manisseri, C.; Arora, R.; Scheller, H.V.; Auer, M.; Vogel, K.P.; Simmons, B.A.; Singh, S. Comparison of dilute acid and ionic liquid pretreatment of switchgrass: Biomass recalcitrance, delignification and enzymatic saccharification. Bioresour. Technol. 2010, 101, 4900–4906. [Google Scholar] [CrossRef] [Green Version]
- Herbaut, M.; Zoghlami, A.; Habrant, A.; Falourd, X.; Foucat, L.; Chabbert, B.; Paes, G. Multimodal analysis of pretreated biomass species highlights generic markers of lignocellulose recalcitrance. Biotechnol. Biofuels 2018, 11, 52. [Google Scholar] [CrossRef] [Green Version]
- Chylenski, P.; Forsberg, Z.; Stahlberg, J.; Varnai, A.; Lersch, M.; Bengtsson, O.; Saebo, S.; Horn, S.J.; Eijsink, V.G.H. Development of minimal enzyme cocktails for hydrolysis of sulfite-pulped lignocellulosic biomass. J. Biotechnol. 2017, 246, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, H.; Lu, M.; Han, L. Comparison and intrinsic correlation analysis based on composition, microstructure and enzymatic hydrolysis of corn stover after different types of pretreatments. Bioresour. Technol. 2019, 293, 122016. [Google Scholar] [CrossRef]
- Sills, D.L.; Gossett, J.M. Assessment of commercial hemicellulases for saccharification of alkaline pretreated perennial biomass. Bioresour. Technol. 2011, 102, 1389–1398. [Google Scholar] [CrossRef]
- Liu, G.; Qu, Y. Engineering of filamentous fungi for efficient conversion of lignocellulose: Tools, recent advances and prospects. Biotechnol. Adv. 2018, 37, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Liu, C.; Zhao, X.; Bai, F. Engineering Trichoderma reesei Rut-C30 with the overexpression of egl1 at the ace1 locus to relieve repression on cellulase production and to adjust the ratio of cellulolytic enzymes for more efficient hydrolysis of lignocellulosic biomass. J. Biotechnol. 2018, 285, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Ajijolakewu, K.A.; Leh, C.P.; Lee, C.K.; Wan Nadiah, W.A. Characterization of novel Trichoderma hemicellulase and its use to enhance downstream processing of lignocellulosic biomass to simple fermentable sugars. Biocatal. Biotransfor. 2017, 11, 166–175. [Google Scholar] [CrossRef]
- Li, Y.; Yi, P.; Liu, J.; Yan, Q.; Jiang, Z. High-level expression of an engineered beta-mannanase (mRmMan5A) in Pichia pastoris for manno-oligosaccharide production using steam explosion pretreated palm kernel cake. Bioresour. Technol. 2018, 256, 30–37. [Google Scholar] [CrossRef]
- Lyu, Y.; Zeng, W.; Du, G.; Chen, J.; Zhou, J. Efficient bioconversion of epimedin C to icariin by a glycosidase from Aspergillus nidulans. Bioresour. Technol. 2019, 289, 121612. [Google Scholar] [CrossRef]
- Oliveira, D.M.; Mota, T.R.; Oliva, B.; Segato, F.; Marchiosi, R.; Ferrarese-Filho, O.; Faulds, C.B.; Dos Santos, W.D. Feruloyl esterases: Biocatalysts to overcome biomass recalcitrance and for the production of bioactive compounds. Bioresour. Technol. 2019, 278, 408–423. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.J.; Lee, H.J.; Kim, K.H. Pure enzyme cocktails tailored for the saccharification of sugarcane bagasse pretreated by using different methods. Process Biochem. 2017, 57, 167–174. [Google Scholar] [CrossRef]
- Hwangbo, M.; Tran, J.L.; Chu, K.H. Effective one-step saccharification of lignocellulosic biomass using magnetite-biocatalysts containing saccharifying enzymes. Sci. Total Environ. 2019, 647, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Vanessa, A.L.R.; Roberto, N.M.; Pereira, N.; Marcelo, F.K.; Elias, L.; Rachael, S.; Clare, S.K.; Leonardo, D.G.; Simon, J.M.M. Characterization of the cellulolytic secretome of Trichoderma harzianum during growth on sugarcane bagasse and analysis of the activity boosting effects of swollenin. Biotechnol. Prog. 2016, 32, 327–336. [Google Scholar]
- Hu, J.; Chandra, R.; Arantes, V.; Gourlay, K.; Van Dyk, J.S.; Saddler, J.N. The addition of accessory enzymes enhances the hydrolytic performance of cellulase enzymes at high solid loadings. Bioresour. Technol. 2015, 186, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhu, N.; Yang, J.; Lin, Y.; Liu, J.; Wang, R.; Wang, F.; Yuan, H. A novel bifunctional acetyl xylan esterase/arabinofuranosidase from Penicillium chrysogenum P33 enhances enzymatic hydrolysis of lignocellulose. Microb. Cell Factories 2017, 16, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, N.; Yang, J.; Ji, L.; Liu, J.; Yang, Y.; Yuan, H. Metagenomic and metaproteomic analyses of a corn stover-adapted microbial consortium EMSD5 reveal its taxonomic and enzymatic basis for degrading lignocellulose. Biotechnol. Biofuels 2016, 9, 243. [Google Scholar] [CrossRef] [Green Version]
- Agger, J.W.; Isaksen, T.; Varnai, A.; Vidal-Melgosa, S.; Willats, W.G.T.; Ludwig, R.; Horn, S.J.; Eijsink, V.G.H.; Westereng, B. Discovery of LPMO activity on hemicelluloses shows the importance of oxidative processes in plant cell wall degradation. Proc. Natl. Acad. Sci. USA 2014, 111, 6287–6292. [Google Scholar] [CrossRef] [Green Version]
- Šuchová, K.; Puchart, V.; Spodsberg, N.; Mørkeberg Krogh, K.B.R.; Biely, P. A novel GH30 xylobiohydrolase from Acremonium alcalophilum releasing xylobiose from the non-reducing end. Enzym. Microb. Technol. 2020, 134, 109484. [Google Scholar] [CrossRef]
- Von Freiesleben, P.; Spodsberg, N.; Blicher, T.H.; Anderson, L.; Jorgensen, H.; Stalbrand, H.; Meyer, A.S.; Krogh, K.B. An Aspergillus nidulans GH26 endo-beta-mannanase with a novel degradation pattern on highly substituted galactomannans. Enzym. Microb. Technol. 2016, 83, 68–77. [Google Scholar] [CrossRef]
- Aulitto, M.; Fusco, F.A.; Fiorentino, G.; Bartolucci, S.; Contursi, P.; Limauro, D. A thermophilic enzymatic cocktail for galactomannans degradation. Enzym. Microb. Technol. 2018, 111, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Liao, Y.; Fu, X.; Zaporski, J.; Peters, S.; Jamison, M.; Liu, Y.; Wullschleger, S.D.; Graham, D.E.; Gu, B. Temperature sensitivity of mineral-enzyme interactions on the hydrolysis of cellobiose and indican by beta-glucosidase. Sci. Total Environ. 2019, 686, 1194–1201. [Google Scholar] [CrossRef]
- Gatt, E.; Rigal, L.; Vandenbossche, V. Biomass pretreatment with reactive extrusion using enzymes: A review. Ind. Crop. Prod. 2018, 122, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Braga, C.M.P.; Delabona, P.D.S.; Lima, D.; Paixao, D.A.A.; Pradella, J.; Farinas, C.S. Addition of feruloyl esterase and xylanase produced on-site improves sugarcane bagasse hydrolysis. Bioresour. Technol. 2014, 170, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Valadares, F.; Goncalves, T.A.; Goncalves, D.S.; Segato, F.; Romanel, E.; Milagres, A.M.; Squina, F.M.; Ferraz, A. Exploring glycoside hydrolases and accessory proteins from wood decay fungi to enhance sugarcane bagasse saccharification. Biotechnol. Biofuels 2016, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Mendoza, D.P.; Junqueira, M.; do Vale, L.H.; Domont, G.B.; Ferreira Filho, E.X.; Sousa, M.V.; Ricart, C.A. Secretomic survey of Trichoderma harzianum grown on plant biomass substrates. J. Proteome Res. 2014, 13, 1810–1822. [Google Scholar] [CrossRef]
- Urbánszki, K.; Szakács, G.; Tengerdy, R.P. Standardization of the filter paper activity assay for solid substrate fermentation. Biotechnol. Lett. 2000, 22, 65–69. [Google Scholar] [CrossRef]
- Liu, L.; Yang, J.; Yang, Y.; Luo, L.; Wang, R.; Zhang, Y.; Yuan, H. Consolidated bioprocessing performance of bacterial consortium EMSD5 on hemicellulose for isopropanol production. Bioresour. Technol. 2019, 292, 121965. [Google Scholar] [CrossRef]
- He, L.; Mao, Y.; Zhang, L.; Wang, H.; Alias, S.A.; Gao, B.; Wei, D. Functional expression of a novel alpha-amylase from Antarctic psychrotolerant fungus for baking industry and its magnetic immobilization. BMC Biotechnol. 2017, 17, 22. [Google Scholar] [CrossRef] [Green Version]
- Ji, G.; Han, L.; Gao, C.; Xiao, W.; Zhang, Y.; Cao, Y. Quantitative approaches for illustrating correlations among the mechanical fragmentation scales, crystallinity and enzymatic hydrolysis glucose yield of rice straw. Bioresour. Technol. 2017, 241, 262–268. [Google Scholar] [CrossRef]
- Hussin, M.H.; Rahim, A.A.; Mohamad Ibrahim, M.N.; Yemloul, M.; Perrin, D.; Brosse, N. Investigation on the structure and antioxidant properties of modified lignin obtained by different combinative processes of oil palm fronds (OPF) biomass. Ind. Crop. Prod. 2014, 52, 544–551. [Google Scholar] [CrossRef]
- Zhu, N.; Liu, J.; Yang, J.; Lin, Y.; Yang, Y.; Ji, L.; Li, M.; Yuan, H. Comparative analysis of the secretomes of Schizophyllum commune and other wood-decay basidiomycetes during solid-state fermentation reveals its unique lignocellulose-degrading enzyme system. Biotechnol. Biofuels 2016, 9, 42. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cocktail | FPA | EG | CBH | βG | Xyl | βX | Ara | Man | Amy |
|---|---|---|---|---|---|---|---|---|---|
| C 9748 | 1.60 ± 0.20 a | 19.70 ± 2.90 a | 52.20 ± 1.90 a | 29.0 ± 2.70 a | 26.60 ± 3.20 a | 34.80 ± 1.40 a | 0.03 ± 0.00 a | 0.71 ± 0.00 a | 0.30 ± 1.00 a |
| Celluclast 1.5 L | 1.90 ± 0.02 a | 8.33 ± 0.18 b | 32.70 ± 1.80 b | 0.27 ± 0.002 b | 10.80 ± 0.14 b | 0.61 ± 0.02 b | 0.31 ± 0.00 b | 3.90 ± 0.11 b | ND |
| EM0925 | 2.77 ± 0.08 b | 11.20 ± 0.13 c | 33.76 ± 2.40 b | 84.97 ± 0.62 c | 314.70 ± 3.23 c | 203.60 ± 18.60 c | 11.72 ± 0.33 c | 6.70 ± 0.12 c | 3.92 ± 0.02 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Yang, J.; Luo, L.; Wang, E.; Wang, R.; Liu, L.; Liu, J.; Yuan, H. Low-Cost Cellulase-Hemicellulase Mixture Secreted by Trichoderma harzianum EM0925 with Complete Saccharification Efficacy of Lignocellulose. Int. J. Mol. Sci. 2020, 21, 371. https://doi.org/10.3390/ijms21020371
Zhang Y, Yang J, Luo L, Wang E, Wang R, Liu L, Liu J, Yuan H. Low-Cost Cellulase-Hemicellulase Mixture Secreted by Trichoderma harzianum EM0925 with Complete Saccharification Efficacy of Lignocellulose. International Journal of Molecular Sciences. 2020; 21(2):371. https://doi.org/10.3390/ijms21020371
Chicago/Turabian StyleZhang, Yu, Jinshui Yang, Lijin Luo, Entao Wang, Ruonan Wang, Liang Liu, Jiawen Liu, and Hongli Yuan. 2020. "Low-Cost Cellulase-Hemicellulase Mixture Secreted by Trichoderma harzianum EM0925 with Complete Saccharification Efficacy of Lignocellulose" International Journal of Molecular Sciences 21, no. 2: 371. https://doi.org/10.3390/ijms21020371
APA StyleZhang, Y., Yang, J., Luo, L., Wang, E., Wang, R., Liu, L., Liu, J., & Yuan, H. (2020). Low-Cost Cellulase-Hemicellulase Mixture Secreted by Trichoderma harzianum EM0925 with Complete Saccharification Efficacy of Lignocellulose. International Journal of Molecular Sciences, 21(2), 371. https://doi.org/10.3390/ijms21020371