New Insights into the Structure-Function Relationship of the Endosomal-Type Na+, K+/H+ Antiporter NHX6 from Mulberry (Morus notabilis)

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Halotolerance Phenotypes of MnNHX6
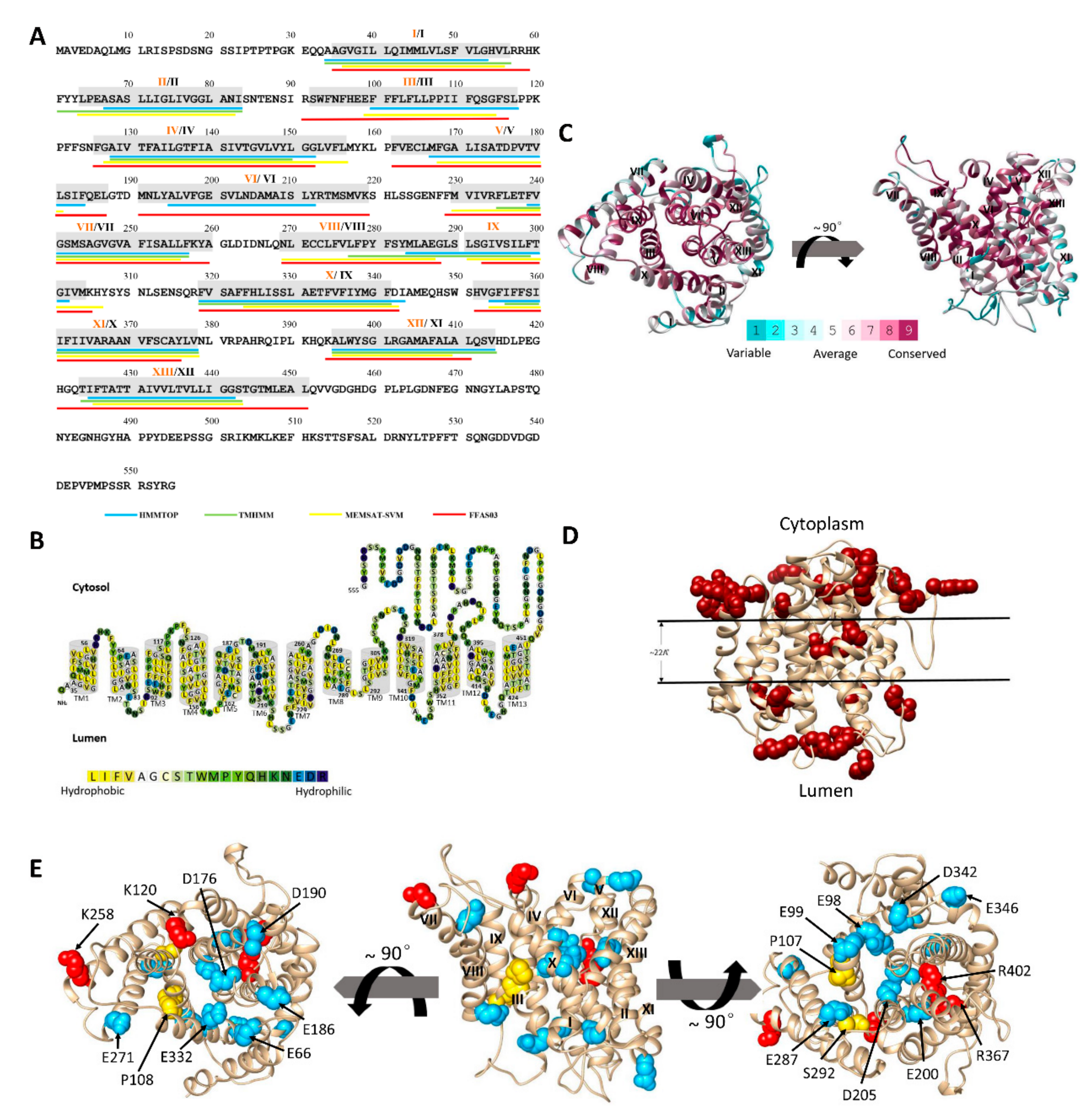
2.2. The Predicted Topology of MnNHX6 Contains 13 Transmembrane Segments
2.3. The 3D Model of MnNHX6 Shares a Typical ‘Funnel’ Fold
2.4. Evolutionary Conservation Supports the MnNHX6 Model Structure
2.5. The MnNHX6 Model Structure Is Consistent with Hydrophobic Characteristics and the ‘Positive-Inside’ Rule
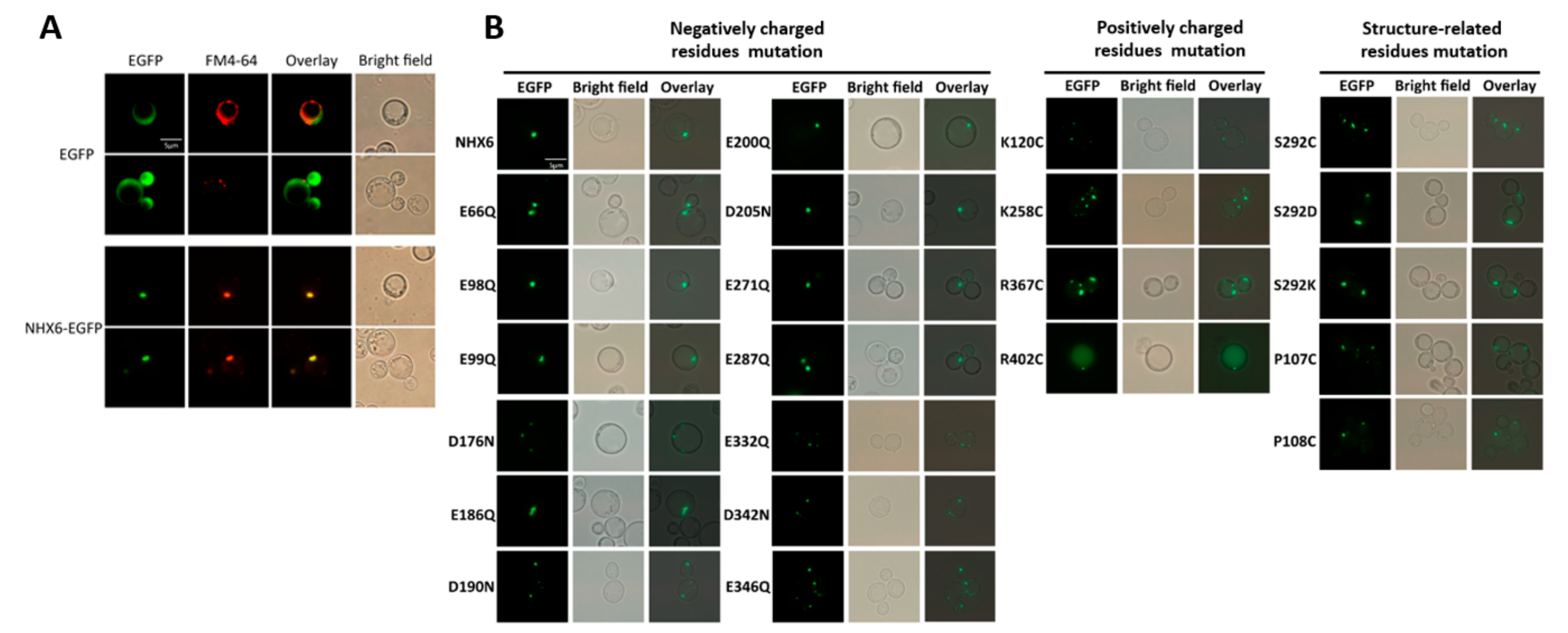
2.6. Experimental Validation of the MnNHX6 Model by Structure-Guided Mutants
3. Discussion
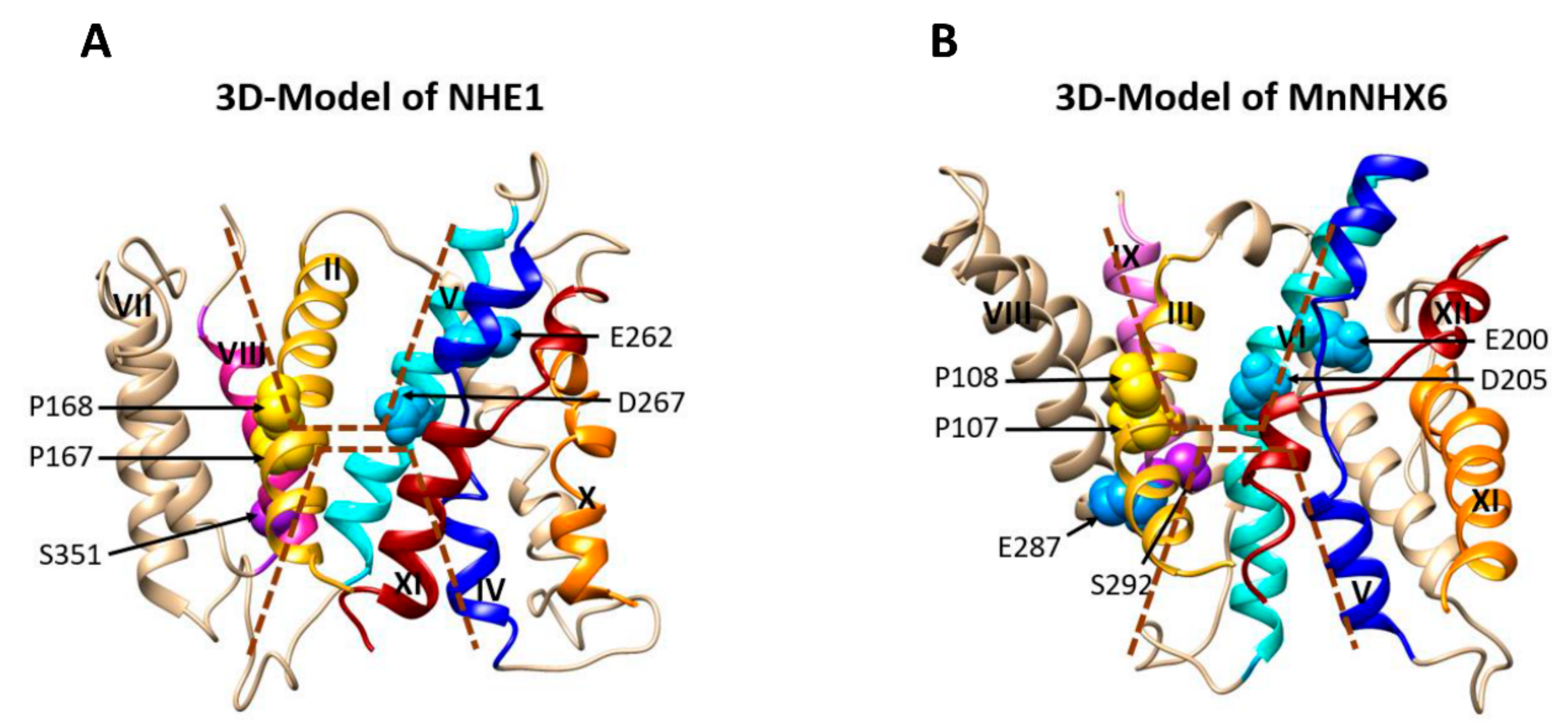
3.1. MnNHX6 Contains a Conserved Transmembrane Assembly and Shares a Similar Charge-Compensated Pattern with NHE1
3.2. Unique Features of MnNHX6′s TM5–TM12 Assembly
3.3. The Endosomal-Type MnNHX6 Shares Similar Transport Mechanisms with NHE1, but Differ from the Vacuolar-Type NHXs
4. Materials and Methods
4.1. Plant Transformation and Stress-Tolerance Analysis of Transgenic Plants
4.2. Yeast Strain, Medium, and Plasmids
4.3. Preparation of Yeast Microsomal Membranes and Western Blotting
4.4. Protein Structure Prediction and Evolutionary Conservation Analysis
4.5. Site-Directed Mutagenesis and Microscopy
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NHX | Na+, K+/H+ antiporters |
| TM | transmembrane |
| 3D | three-dimensional |
| WT | wild-type |
| OE | overexpressing |
| MDA | malondialdehyde |
| qRT-PCR | reverse transcription quantitative PCR |
| TGN | trans-Golgi network |
| PVC | prevacuolar compartment |
| HYG | hygromycin B |
| EGFP | enhanced green fluorescent protein |
References
- Leigh, R.A.; Jones, R.G.W. A hypothesis relating critical potassium concentrations for growth to the distribution and functions of this ion in the plant cell. New Phytol. 2010, 97, 1–13. [Google Scholar] [CrossRef]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Bassil, E.; Blumwald, E. The ins and outs of intracellular ion homeostasis: NHX-type cation/H+ transporters. Curr. Opin. Plant Biol. 2014, 22, 1–6. [Google Scholar] [CrossRef]
- Blumwald, E. Tonoplast vesicles as a tool in the study of ion transport at the plant vacuole. Physiol. Plant. 1987, 69, 731–734. [Google Scholar] [CrossRef]
- Chanroj, S.; Wang, G.; Venema, K.; Zhang, M.W.; Delwiche, C.F.; Sze, H. Conserved and diversified gene families of monovalent cation/H+ antiporters from algae to flowering plants. Front. Plant Sci. 2012, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Bassil, E.; Coku, A.; Blumwald, E. Cellular ion homeostasis: Emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 2012, 63, 5727–5740. [Google Scholar] [CrossRef] [Green Version]
- Bassil, E.; Ohto, M.A.; Esumi, T.; Tajima, H.; Zhu, Z.; Blumwald, E. The Arabidopsis intracellular Na+/H+ antiporters NHX5 and NHX6 are endosome associated and necessary for plant growth and development. Plant Cell 2011, 23, 224–239. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Apse, M.P.; Shi, H.; Blumwald, E. Topological analysis of a plant vacuolar Na+/H+ antiporter reveals a luminal C terminus that regulates antiporter cation selectivity. Proc. Natl. Acad. Sci. USA 2003, 100, 12510–12515. [Google Scholar] [CrossRef] [Green Version]
- Wakabayashi, S.; Pang, T.; Su, X.; Shigekawa, M. A novel topology model of the human Na+/H+ exchanger isoform 1. J. Biol. Chem. 2000, 275, 7942–7949. [Google Scholar] [CrossRef] [Green Version]
- Landau, M.; Herz, K.; Padan, E.; Ben-Tal, N. Model structure of the Na+/H+ exchanger 1 (NHE1) functional and clinical implications. J. Biol. Chem. 2007, 282, 37854–37863. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.A. Three-dimensional structure of the ion-coupled transport protein NhaA. Nature 2000, 403, 112–115. [Google Scholar] [CrossRef]
- Schushan, M.; Xiang, M.; Bogomiakov, P.; Padan, E.; Rao, R.; Ben-Tal, N. Model-guided mutagenesis drives functional studies of human NHA2, implicated in hypertension. J. Mol. Biol. 2010, 396, 1181–1196. [Google Scholar] [CrossRef] [Green Version]
- Kondapalli, K.C.; Hack, A.; Schushan, M.; Landau, M.; Ben-Tal, N.; Rao, R. Functional evaluation of autism-associated mutations in NHE9. Nat. Commun. 2013, 4, 2510. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Feng, X.; Zhao, H.; Wang, L.; An, L.; Qiu, Q.S. Functional analysis of the Na+, K+/H+ antiporter PeNHX3 from the tree halophyte Populus euphratica in yeast by model-guided mutagenesis. PLoS ONE 2014, 9, e104147. [Google Scholar] [CrossRef] [Green Version]
- Paulino, C.; Wöhlert, D.; Kapotova, E.; Yildiz, Ö.; Kühlbrandt, W. Structure and transport mechanism of the sodium/proton antiporter MjNhaP1. Elife 2014, 3, e03583. [Google Scholar] [CrossRef]
- Wöhlert, D.; Kühlbrandt, W.; Yildiz, Ö. Structure and substrate ion binding in the sodium/proton antiporter PaNhaP. Elife 2014, 3, e03579. [Google Scholar] [CrossRef]
- Ashnest, J.R.; Huynh, D.L.; Dragwidge, J.M.; Ford, B.A.; Gendall, A.R. Arabidopsis intracellular NHX-type sodium-proton antiporters are required for seed storage protein processing. Plant Cell Physiol. 2015, 56, 2220–2233. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rosales, M.P.; Gálvez, F.J.; Huertas, R.; Aranda, M.N.; Baghour, M.; Cagnac, O.; Venema, K. Plant NHX cation/proton antiporters. Plant Signal. Behav. 2009, 4, 265–276. [Google Scholar] [CrossRef] [Green Version]
- He, N.J.; Zhang, C.; Qi, X.W.; Zhao, S.C.; Tao, Y.; Yang, G.J.; Lee, T.H.; Wang, X.Y.; Cai, Q.L.; Li, D.; et al. Draft genome sequence of the mulberry tree Morus notabilis. Nat. Commun. 2013, 4, 2445. [Google Scholar] [CrossRef] [Green Version]
- Checker, V.G.; Khurana, P. Molecular and functional characterization of mulberry EST encoding remorin (MiREM) involved in abiotic stress. Plant Cell Rep. 2013, 32, 1729–1741. [Google Scholar] [CrossRef]
- Cao, B.; Long, D.; Zhang, M.; Liu, C.; Xiang, Z.; Zhao, A. Molecular characterization and expression analysis of the mulberry Na+/H+ exchanger gene family. Plant Physiol. Biochem. 2016, 99, 49–58. [Google Scholar] [CrossRef]
- Hernández, A.; Jiang, X.; Cubero, B.; Nieto, P.M.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. Mutants of the Arabidopsis thaliana Cation/H+ antiporter AtNHX1 conferring increased salt tolerance in Yeast the endosome/prevacuolar compartment is a target for salt toxicity. J. Biol. Chem. 2009, 284, 14276–14285. [Google Scholar] [CrossRef] [Green Version]
- Jaroszewski, L.; Rychlewski, L.; Li, Z.; Li, W.; Godzik, A. FFAS03: A server for profile-profile sequence alignments. Nucleic Acids Res. 2005, 33, 284–288. [Google Scholar] [CrossRef] [Green Version]
- Tusnady, G.E.; Simon, I. The HMMTOP transmembrane topology prediction server. Bioinformatics 2001, 17, 849–850. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Nugent, T.; Jones, D.T. Detecting pore-lining regions in transmembrane protein sequences. BMC Bioinform. 2012, 13, 169. [Google Scholar] [CrossRef] [Green Version]
- Kessel, A.; Ben, T.N. Free energy determinants of peptide association with lipid bilayers. Curr. Top. Membr. 2002, 52, 205–253. [Google Scholar]
- Heijne, G. The distribution of positively charged residues in bacterial inner membrane proteins correlates with the transmembrane topology. EMBO J. 1986, 5, 3021–3027. [Google Scholar] [CrossRef]
- Wallin, E.; Heijne, G.V. Genome-wide analysis of integral membrane proteins from eubacterial, archaean, and eukaryotic organisms. Protein Sci. 1998, 7, 1029–1038. [Google Scholar] [CrossRef]
- Ali, R.; Brett, C.L.; Mukherjee, S.; Rao, R. Inhibition of sodium/proton exchange by a Rab-GTPase-activating protein regulates endosomal traffic in yeast. J. Biol. Chem. 2004, 279, 4498–4506. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, I.; Rubio, F.; Rodriguez-Navarro, A.; Pardo, J.M. The protein phosphatase calcineurin is essential for NaCl tolerance of Saccharomyces cerevisiae. J. Biol. Chem. 1994, 269, 8792–8796. [Google Scholar]
- Pires, I.S.; Negrão, S.; Pentony, M.M.; Abreu, I.A.; Oliveira, M.M.; Purugganan, M.D. Different evolutionary histories of two cation/proton exchanger gene families in plants. BMC Plant Biol. 2013, 13, 97. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rosales, M.P.; Jiang, X.; Gálvez, F.J.; Aranda, M.N.; Cubero, B.; Venema, K. Overexpression of the tomato K+/H+ antiporter LeNHX2 confers salt tolerance by improving potassium compartmentalization. New Phytol. 2008, 179, 366–377. [Google Scholar] [CrossRef]
- Sun, M.H.; Ma, Q.J.; Hu, D.G.; Zhu, X.P.; You, C.X.; Shu, H.R. The glucose sensor mdhxk1 phosphorylates a tonoplast Na+/H+ exchanger to improve salt tolerance. Plant Physiol. 2018, 176, 2977–2990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, R.J.; Murtha-Riel, P.; Pittman, D.D.; Davies, M.V. Characterization of wild-type and ser53 mutant eukaryotic initiation factor 4e overexpression in mammalian cells. J. Biol. Chem. 1993, 268, 11902–11909. [Google Scholar] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant J. 1949, 24, 1. [Google Scholar] [CrossRef] [Green Version]
- Mayrose, I.; Mitchell, A.; Pupko, T. Site-specific evolutionary rate inference: Taking phylogenetic uncertainty into account. J. Mol. Evol. 2005, 60, 345–353. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Biol. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Zhang, H.; Blumwald, E.; Xia, T. A novel plant vacuolar Na+/H+ antiporter gene evolved by DNA shuffling confers improved salt tolerance in yeast. J. Biol. Chem. 2010, 285, 22999–23006. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Damage | Subcellular | Conservation | Essential | Putative | |||||
|---|---|---|---|---|---|---|---|---|---|
| Mutant | NaCl | KCl | HYG | Localization | Residue | Location | Score | Residue (Y/N) | Role |
| E66Q | Mild | Significant | Significant | PVC | Glu66 | Core | 8 | Y | Charge compensated |
| E98Q | Mild | Significant | Subtle | PVC | Glu98 | Extramembrane loop | 6 | Y | Unclear |
| E99Q | Mild | Significant | Subtle | PVC | Glu99 | Extramembrane loop | 6 | Y | Unclear |
| D176N | Abolish | Abolish | Abolish | PVC | Asp176 | Core | 9 | Y | Charge compensated |
| E186Q | Mild | Significant | Subtle | PVC | Glu186 | Extramembrane loop | 7 | Y | Unclear |
| D190N | Subtle | Mild | Subtle | PVC | Asp190 | Extramembrane loop | 8 | N | - |
| E200Q | Abolish | Abolish | Abolish | PVC | Glu200 | Core | 9 | Y | Ion translocation |
| D205N | Abolish | Abolish | Abolish | PVC | Asp205 | Core | 9 | Y | Ion translocation |
| E271Q | Mild | Significant | Abolish | PVC | Glu271 | Extramembrane loop | 9 | Y | pH regulation |
| E287Q | Mild | Significant | Abolish | PVC | Glu287 | Core | 8 | Y | Ion translocation |
| E332Q | Mild | Significant | Abolish | PVC | Glu332 | Core | 9 | Y | Charge compensated |
| D342N | Subtle | Mild | Subtle | PVC | Asp342 | Extramembrane loop | 7 | N | - |
| D346N | Mild | Significant | Abolish | PVC | Glu346 | Extramembrane loop | 6 | Y | pH regulation |
| K120C | Mild | Significant | Abolish | PVC | Lys120 | Extramembrane loop | 5 | Y | pH regulation |
| K258C | Mild | Significant | Abolish | PVC | Lys258 | Extramembrane loop | 9 | Y | pH regulation |
| R367C | Abolish | Abolish | Abolish | PVC | Arg367 | Core | 9 | Y | Charge compensated |
| R402C | Mild | Significant | Abolish | Partial missorting | Arg402 | Core | 9 | Y | Charge compensated |
| P107C | Mild | Significant | Abolish | PVC | Pro107 | Core | 9 | Y | Ion translocation |
| P108C | Mild | Significant | Abolish | PVC | Pro108 | Core | 9 | Y | Ion translocation |
| S292C/D/K | Mild | Significant | Abolish | PVC | Ser292 | Core | 9 | Y | Ion translocation |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, B.; Xia, Z.; Liu, C.; Fan, W.; Zhang, S.; Liu, Q.; Xiang, Z.; Zhao, A. New Insights into the Structure-Function Relationship of the Endosomal-Type Na+, K+/H+ Antiporter NHX6 from Mulberry (Morus notabilis). Int. J. Mol. Sci. 2020, 21, 428. https://doi.org/10.3390/ijms21020428
Cao B, Xia Z, Liu C, Fan W, Zhang S, Liu Q, Xiang Z, Zhao A. New Insights into the Structure-Function Relationship of the Endosomal-Type Na+, K+/H+ Antiporter NHX6 from Mulberry (Morus notabilis). International Journal of Molecular Sciences. 2020; 21(2):428. https://doi.org/10.3390/ijms21020428
Chicago/Turabian StyleCao, Boning, Zhongqiang Xia, Changying Liu, Wei Fan, Shuai Zhang, Qiao Liu, Zhonghuai Xiang, and Aichun Zhao. 2020. "New Insights into the Structure-Function Relationship of the Endosomal-Type Na+, K+/H+ Antiporter NHX6 from Mulberry (Morus notabilis)" International Journal of Molecular Sciences 21, no. 2: 428. https://doi.org/10.3390/ijms21020428
APA StyleCao, B., Xia, Z., Liu, C., Fan, W., Zhang, S., Liu, Q., Xiang, Z., & Zhao, A. (2020). New Insights into the Structure-Function Relationship of the Endosomal-Type Na+, K+/H+ Antiporter NHX6 from Mulberry (Morus notabilis). International Journal of Molecular Sciences, 21(2), 428. https://doi.org/10.3390/ijms21020428