Riboflavin Plays a Pivotal Role in the UVA-Induced Cytotoxicity of Fibroblasts as a Key Molecule in the Production of H2O2 by UVA Radiation in Collaboration with Amino Acids and Vitamins


Abstract
:
1. Introduction
2. Results
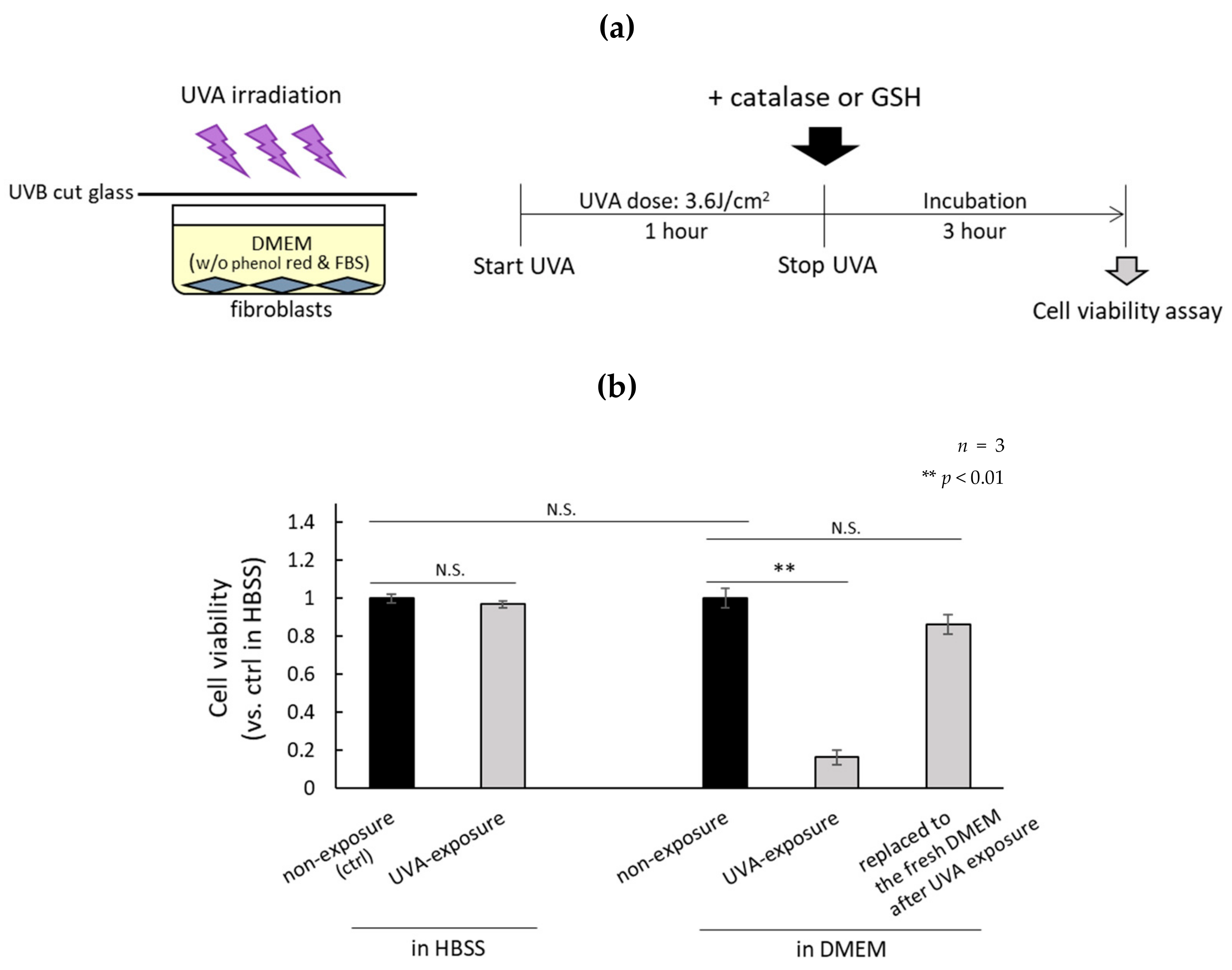
2.1. H2O2 Generated by UVA in DMEM Is Highly Cytotoxic
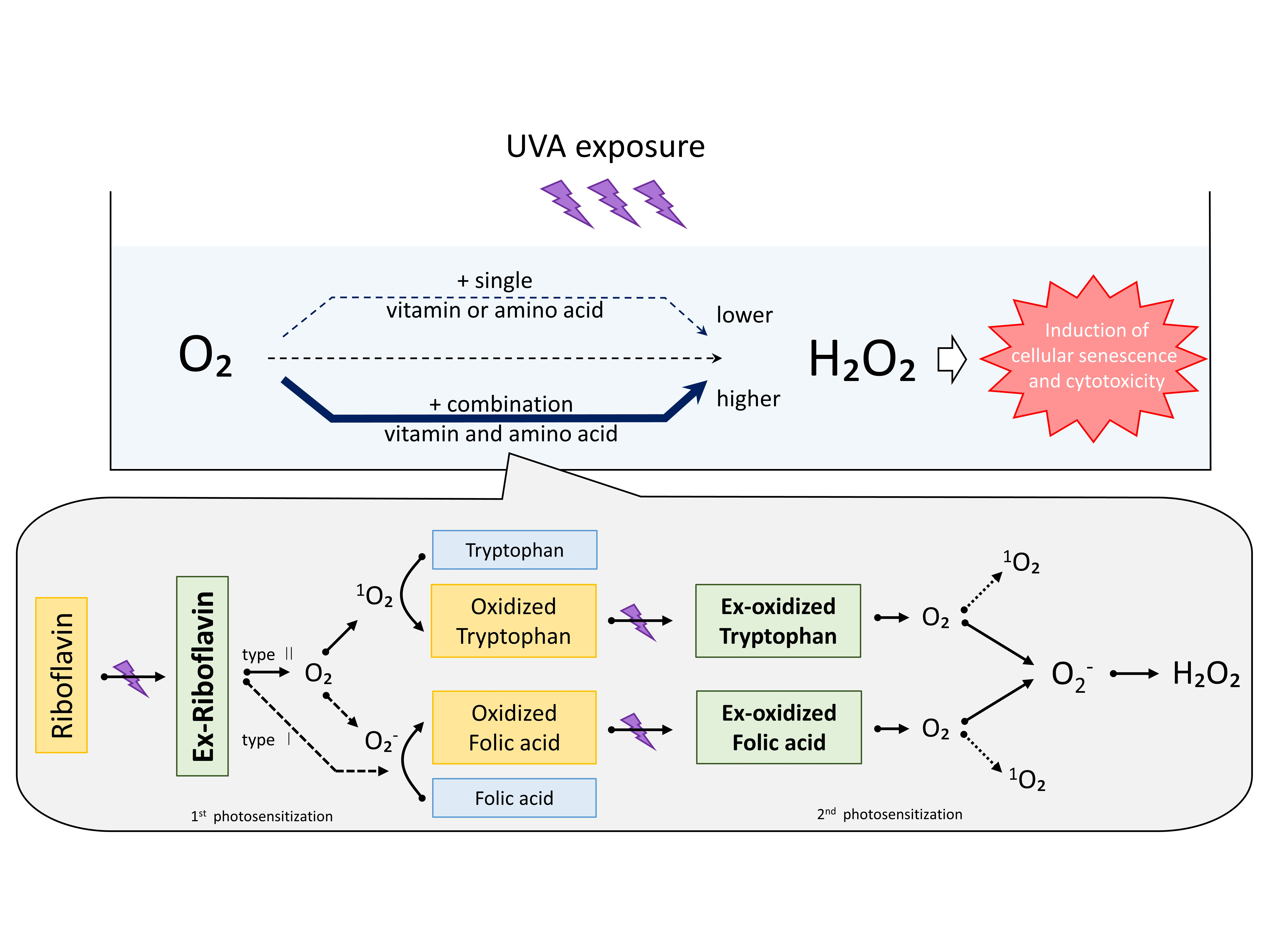
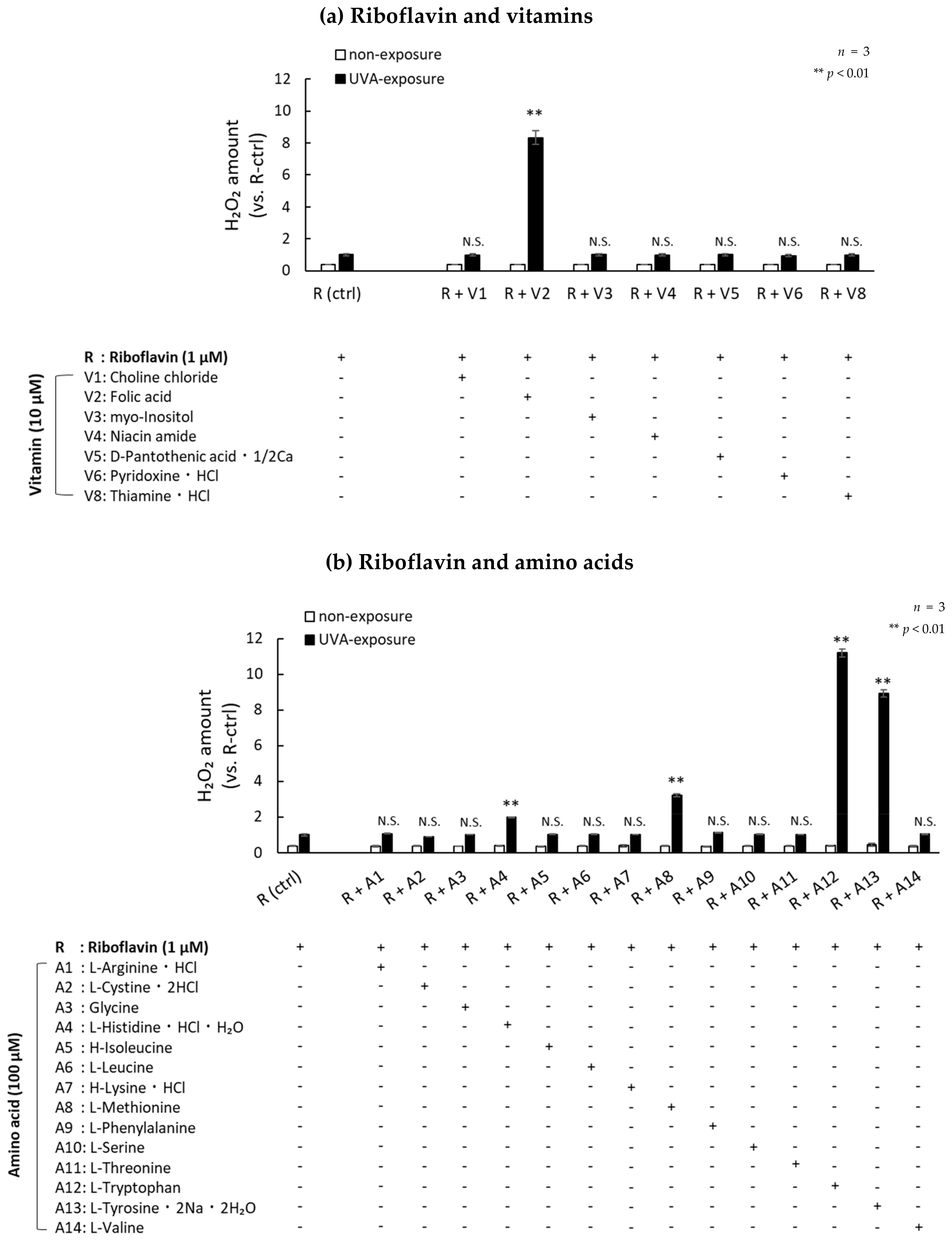
2.2. Photosensitization by UVA Is Amplified in the Presence of Amino Acids and Vitamins
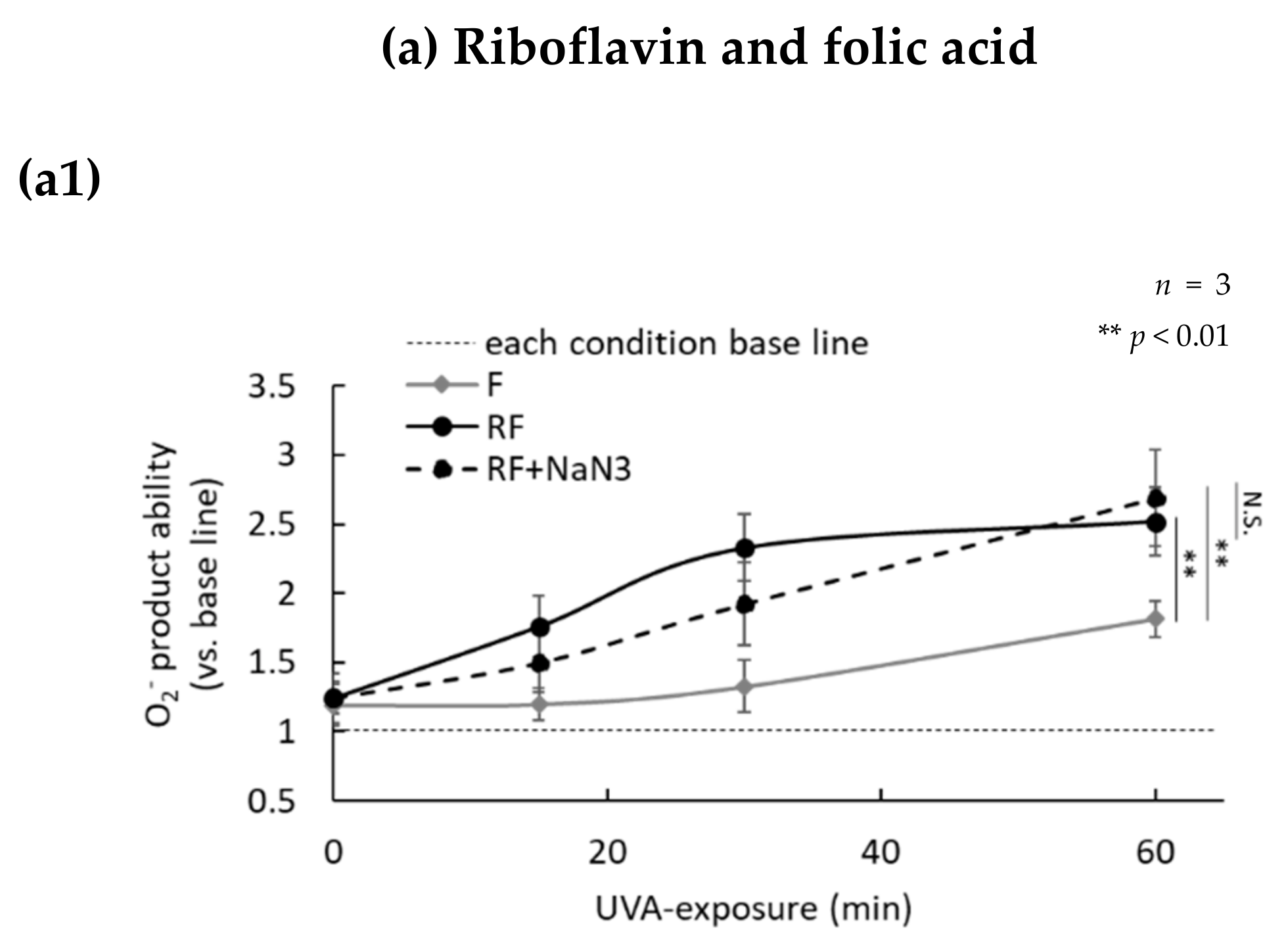
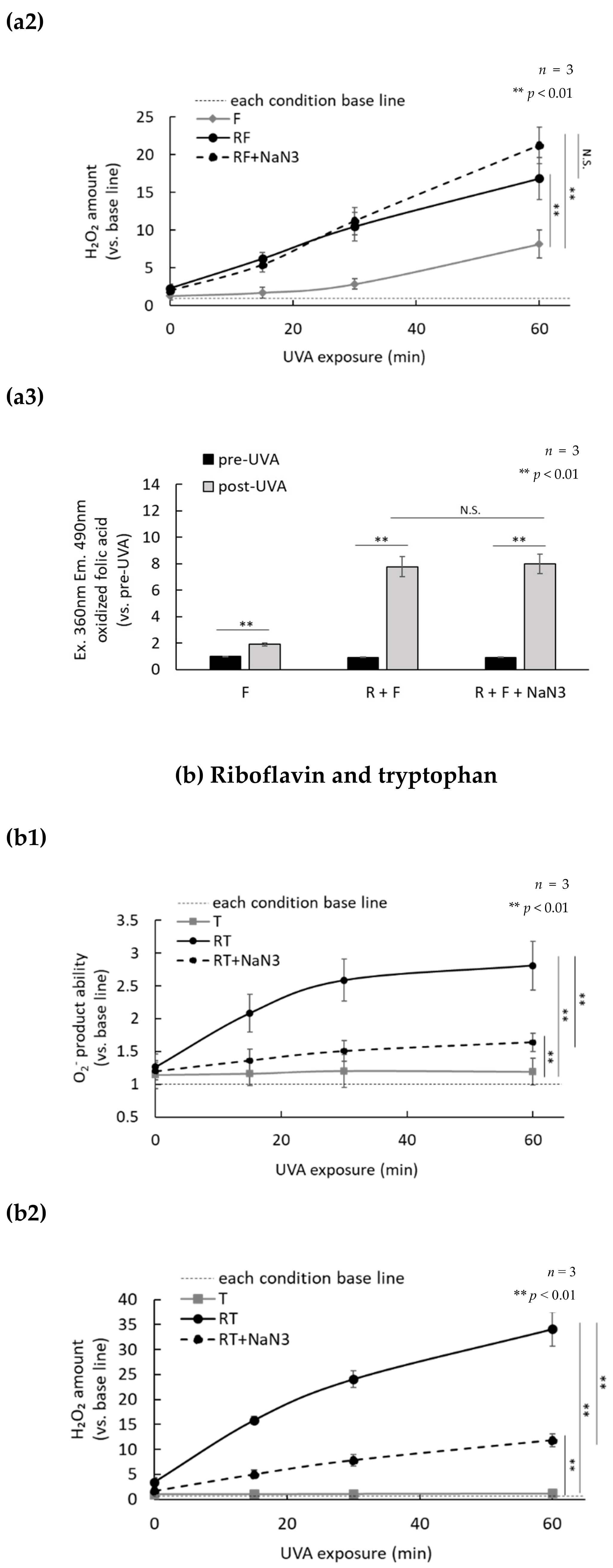
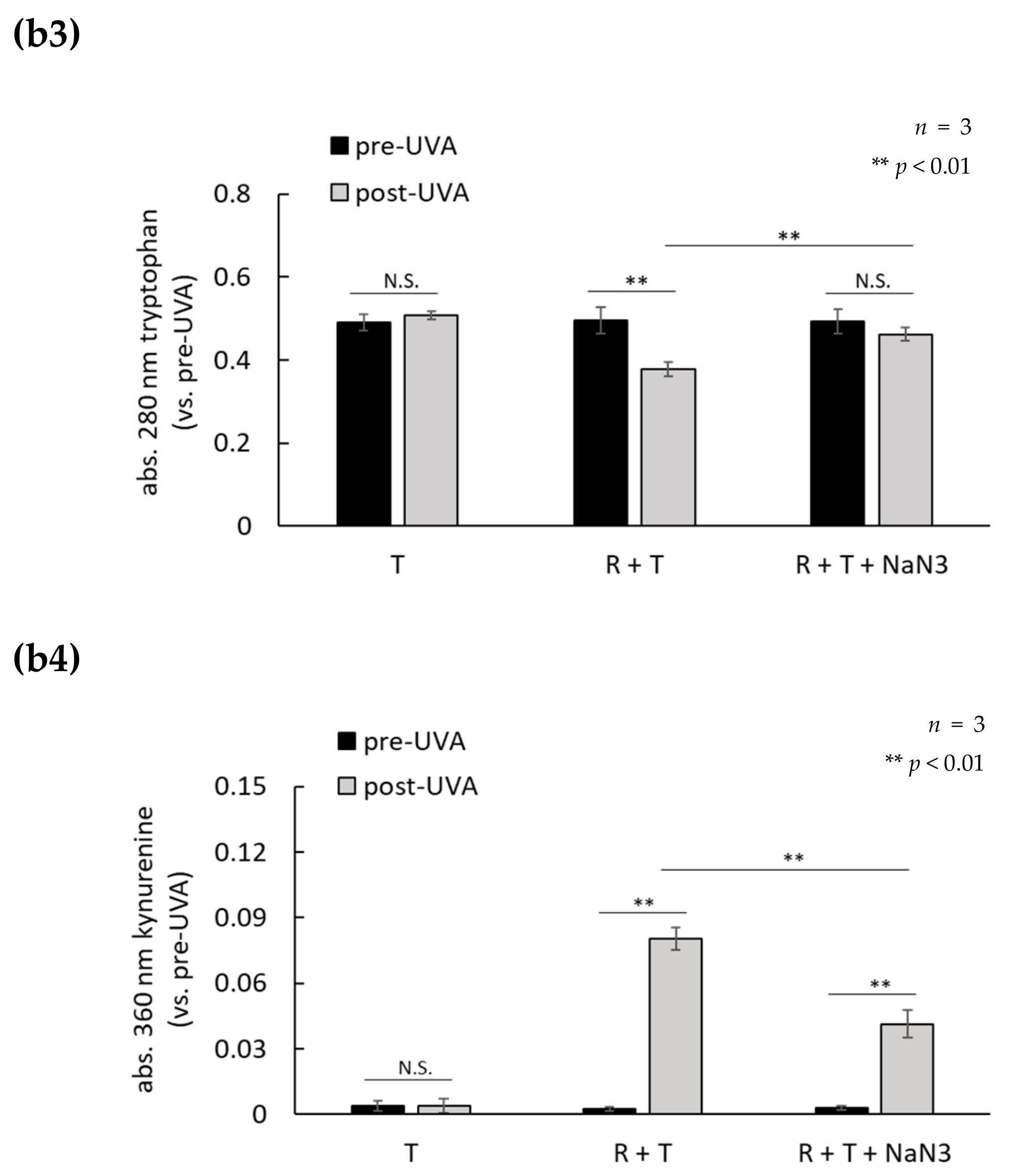
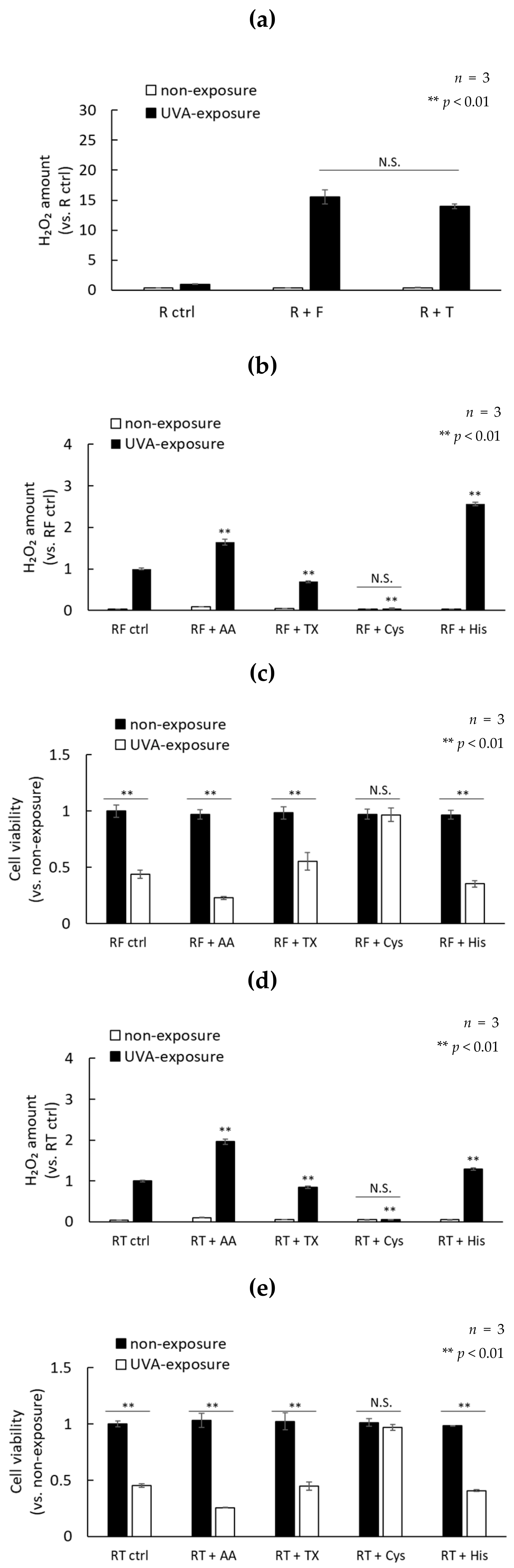
2.3. H2O2 Generated by UVA Exposure Is Amplified by a Photosensitization Reaction between Riboflavin and Amino Acids via a Singlet Oxygen
2.4. UVA Exposure to Antioxidants in the Presence of Riboflavin Increases the Production of H2O2
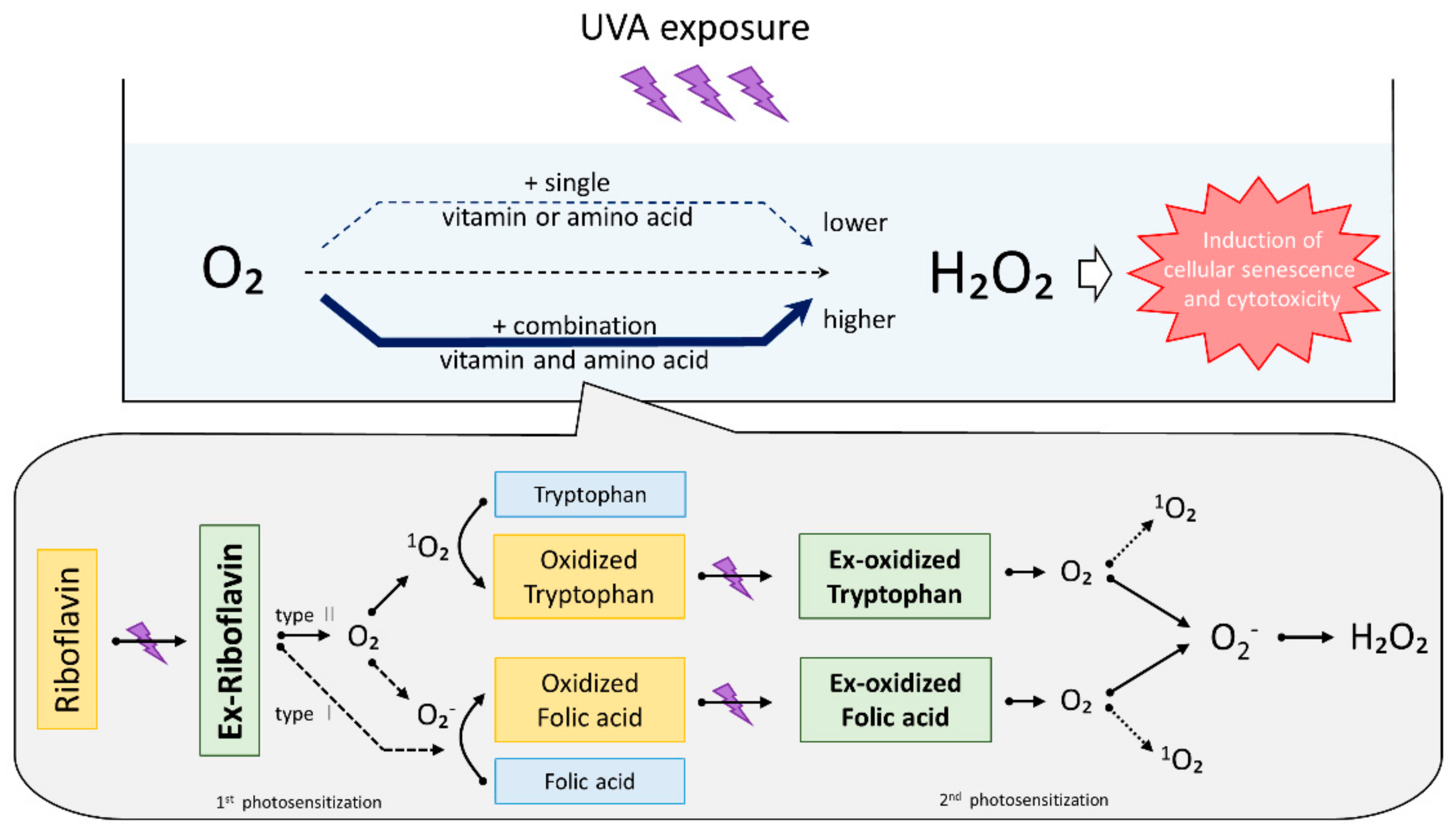
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. UVA Irradiation
4.3. Cell Viability Assay
4.4. Preparation of Media Similar to DMEM
4.5. H2O2 Assay
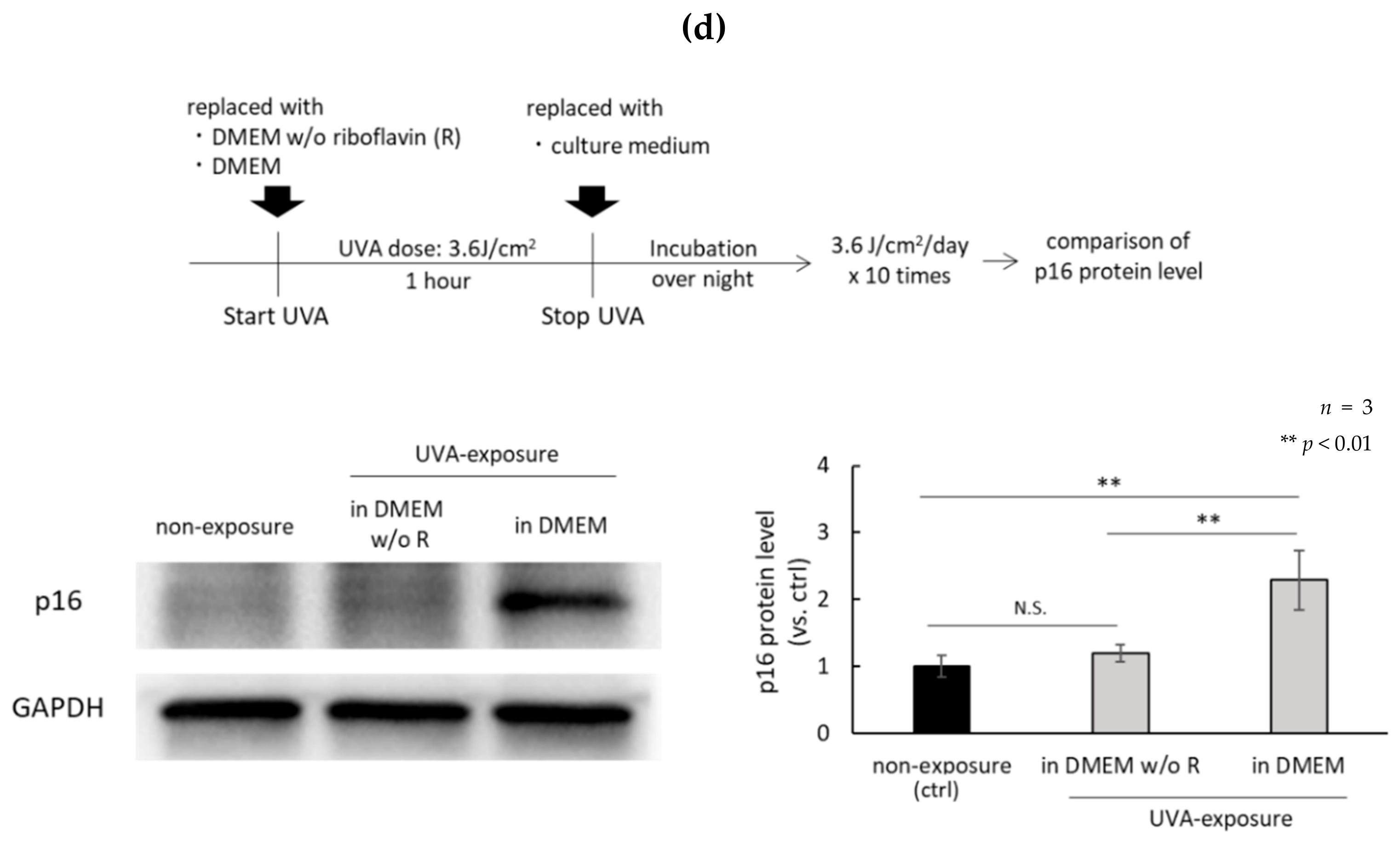
4.6. Induction of Cellular Senescence by Repeated UVA Exposure
4.7. Detection of p16 by Western Blotting
4.8. Superoxide Assay
4.9. Measurement of Auto-Fluorescence of Oxidized Folic Acid
4.10. Measurement of Tryptophan and Kynurenine
4.11. Evaluation of the Effects of Antioxidants on UVA Photosensitization
4.12. H2O2 Scavenging Ability Assay
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DMEM | Dulbecco’s modified Eagle’s medium |
| ROS | Reactive oxygen species |
| PBS | Phosphate Buffered Saline |
| HBSS | Hanks Balanced Salt Solution |
| GSH | Glutathione |
| H2O2 | Hydrogen peroxide |
| UVA | Ultraviolet A |
References
- Ahmed, N.U.; Ueda, M.; Nikaido, O.; Osawa, T.; Ichihashi, M. High levels of 8-hydroxy-2′-deoxyguanosine appear in normal human epidermis after a single dose of ultraviolet radiation. Br. J. Dermatol. 1999, 140, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Rünger, T.M. Longwave UV light induces the aging-associated progerin. J. Investig. Dermatol. 2013, 133, 1857–1862. [Google Scholar] [PubMed] [Green Version]
- Mahns, A.; Melchheier, I.; Suschek, C.V.; Sies, H.; Klotz, L.O. Irradiation of cells with ultraviolet-A (320-400nm) in the presence of cell culture medium elicits biological effects due to extracellular generation of hydrogen peroxide. Free Radic. Res. 2003, 37, 391–397. [Google Scholar] [PubMed]
- Imokawa, G.; Ishida, K. Biological mechanisms underlying the ultraviolet radiation-induced formation of skin wrinkling and sagging I: Reduced skin elasticity, highly associated with enhanced dermal elastase activity, triggers wrinkling and sagging. Int. J. Mol. Sci. 2015, 16, 7753–7775. [Google Scholar] [PubMed] [Green Version]
- Ichihashi, M.; Ando, H. The maximal cumulative solar UVB dose allowed to maintain healthy and young skin and prevent premature photoaging. Exp. Dermatol. 2014, 1, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Mohania, D.; Chandel, S.; Kumar, P.; Verma, V.; Digvijay, K.; Tripathi, D.; Choudhury, K.; Mitten, S.K.; Shah, D. Skin defense-damage mechanism. Ultrav. Radiat. Adv. Exp. Med. Biol. 2017, 996, 71–87. [Google Scholar]
- Edström, D.W.; Porwit, A.; Ros, A.M. Effects on human skin of repetitive ultraviolet-A1 (UVA1) irradiation and visible light. Photodermatol. Photoimmunol. Photomed. 2001, 17, 66–70. [Google Scholar] [CrossRef]
- Mullenders, L.H.F. Solar UV damage to cellular DNA: From mechanisms to biological effects. Photochem. Photobiol. Sci. 2018, 17, 1842–1852. [Google Scholar]
- Ghosh, R.; Amstad, P.; Cerutti, P. UVB-induced DNA breaks interfere with transcriptional induction of c-fos. Mol. Cell. Biol. 1993, 13, 6992–6999. [Google Scholar] [CrossRef] [Green Version]
- El-Mahdy, M.A.; Zhu, Q.; Wang, Q.E.; Wani, G.; Patnaik, S.; Zhao, Q.; Arafa, E.-S.; Barakat, B.; Mir, S.N.; Wani, A.A. Naringenin protects HaCaT human keratinocytes against UVB-induced apoptosis and enhances the removal of cyclobutane pyrimidine dimers from the genome. Photochem. Photobiol. 2008, 84, 307–316. [Google Scholar]
- Babu, V.; Joshi, P.C. Tryptophan as an endogenous photosensitizer to elicit harmful effects of ultraviolet B. Indian J. Biochem. Biophys. 1992, 29, 296–298. [Google Scholar] [PubMed]
- Yamaguchi, Y.; Coelho, S.G.; Zmudzka, B.Z.; Takahashi, K.; Beer, J.Z.; Hearing, V.J.; Miller, S.A. Cyclobutane pyrimidine dimer formation and p53 production in human skin after repeated UV irradiation. Exp. Dermatol. 2008, 17, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.; Jonason, A.S.; Leffell, D.J.; Simon, J.A.; Sharma, H.W.; Kimmelman, J.; Remington, L.; Jacks, T.; Brash, D.E. Sunburn and p53 in the onset of skin cancer. Nature 1994, 372, 773–776. [Google Scholar] [CrossRef] [PubMed]
- Kligman, L.H.; Sayre, R.M. An action spectrum for ultraviolet induced elastosis in hairless mice: Quantification of elastosis by image analysis. Photochem. Photobiol. 1991, 53, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.C. Comparison of the DNA-damaging property of photosensitised riboflavin via singlet oxygen (1O2) and superoxide radical O2-. mechanisms. Toxicol. Lett. 1985, 26, 211–217. [Google Scholar] [CrossRef]
- Park, S.L.; Justiniano, R.; Williams, J.D.; Cabello, C.M.; Qiao, S.; Wondrak, G.T. The tryptophan-derived endogenous aryl hydrocarbon receptor ligand 6-formylindolo[3,2-b]carbazole is a nanomolar UVA photosensitizer in epidermal keratinocytes. J. Investig. Dermatol. 2015, 135, 1649–1658. [Google Scholar] [CrossRef] [Green Version]
- Premi, S.; Wallisch, S.; Mano, C.M.; Weiner, A.B.; Bacchiocchi, A.; Wakamatsu, K.; Bechara, E.J.; Halaban, R.; Douki, T.; Brash, D.E. Photochemistry. Chemiexcitation of melanin derivatives induces DNA photoproducts long after UV exposure. Science 2015, 347, 842–847. [Google Scholar] [CrossRef]
- Kawanishi, S.; Hiraku, Y.; Oikawa, S. Mechanism of guanine-specific DNA damage by oxidative stress and its role in carcinogenesis and aging. Mutat. Res. 2001, 488, 65–76. [Google Scholar] [CrossRef]
- Yoshimoto, S.; Yoshida, M.; Ando, H.; Ichihashi, M. Establishment of photoaging in vitro by repetitive UVA irradiation: Induction of characteristic markers of senescence and its prevention by PAPLAL with potent catalase activity. Photochem. Photobiol. 2018, 94, 438–444. [Google Scholar] [CrossRef]
- Sato, K.; Taguchi, H.; Maeda, T.; Minami, H.; Asada, Y.; Watanabe, Y.; Yoshikawa, K. The primary cytotoxicity in ultraviolet-a-irradiated riboflavin solution is derived from hydrogen peroxide. J. Investig. Dermatol. 1995, 105, 608–612. [Google Scholar] [CrossRef] [Green Version]
- Klotz, L.O.; Briviba, K.; Sies, H. Singlet oxygen mediates the activation of JNK by UVA radiation in human skin fibroblasts. FEBS Lett. 1997, 408, 289–291. [Google Scholar] [CrossRef] [Green Version]
- Chou, P.T.; Khan, A.U. L-ascorbic acid quenching of singlet delta molecular oxygen in aqueous media: Generalized antioxidant property of vitamin C. Biochem. Biophys. Res. Commun. 1983, 115, 932–937. [Google Scholar] [CrossRef]
- Ohtsu, I.; Wiriyathanawudhiwong, N.; Morigasaki, S.; Nakatani, T.; Kadokura, H.; Takagi, H. The L-cysteine/L-cystine shuttle system provides reducing equivalents to the periplasm in Escherichia coli. J. Biol. Chem. 2010, 23, 17479–17487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hearse, D.J.; Kusama, Y.; Bernier, M. Rapid electrophysiological changes leading to arrhythmias in the aerobic rat heart. Photosensitization studies with rose bengal-derived reactive oxygen intermediates. Circ. Res. 1989, 1, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotoh, N.; Niki, E. Rates of interactions of superoxide with vitamin E, vitamin C and related compounds as measured by chemiluminescence. Biochim. Biophys. Acta 1992, 1115, 201–207. [Google Scholar] [CrossRef]
- Dántola, M.L.; Denofrio, M.P.; Zurbano, B.; Gimenez, C.S.; Ogilby, P.R.; Lorente, C.; Thomas, A.H. Mechanism of photooxidation of folic acid sensitized by unconjugated pterins. Photochem. Photobiol. Sci. 2010, 9, 1604–1612. [Google Scholar] [CrossRef]
- Kitagishi, K.; Hiromi, K.; Tokushige, M. Binding between thermolysin and talopeptin (MKI) in which the tryptophan residue was converted into kynurenine. J. Biochem. 1983, 93, 1045–1054. [Google Scholar] [CrossRef]
- Shimizu, R.; Yagi, M.; Kikuchi, A. Suppression of riboflavin-sensitized singlet oxygen generation by l-ascorbic acid, 3-O-ethyl-l-ascorbic acid and Trolox. J. Photochem. Photobiol. B 2019, 191, 116–122. [Google Scholar] [CrossRef]
- Yi, Y.; Xie, H.; Xiao, X.; Wang, B.; Du, R.; Liu, Y.; Li, Z.; Wang, J.; Sun, L.; Deng, Z.; et al. Ultraviolet A irradiation induces senescence in dermal fibroblasts by down-regulating OXMT1 via ZEO1. Aging 2018, 10, 212–228. [Google Scholar] [CrossRef] [Green Version]
- Schuch, A.P.; Moreno, N.C.; Schuch, N.J.; Menck, C.F.M.; Garcia, C.C.M. Sunlight damage to cellular DNA: Focus on oxidatively generated lesions. Free Radic. Biol. 2017, 107, 110–112. [Google Scholar] [CrossRef]
- Tyrrell, R. Redox regulation and oxidant activation of heme oxygenase-1. Free Radic. Res. 1999, 31, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Juzeniene, A.; Thu Tam, T.T.; Iani, V.; Moan, J. The action spectrum for folic acid photodegradation in aqueous solutions. J. Photochem. Photobiol. B 2013, 126, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.H.; Serrano, M.P.; Rahal, V.; Vicendo, P.; Claparols, C.; Oliveros, E.; Lorente, C. Tryptophan oxidation photosensitized by pterin. Free Radic. Biol. Med. 2013, 63, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Rokos, H.; Beazley, W.D.; Schallreuter, K.U. Oxidative stress in vitiligo: Photo-oxidation of pterins produces H(2)O(2) and pterin-6-carboxylic acid. Biochem. Biophys. Res. Commun. 2002, 292, 805–811. [Google Scholar] [CrossRef]
- Castaño, C.; Dántola, M.L.; Oliveros, E.; Thomas, A.H.; Lorente, C. Oxidation of tyrosine photoinduced by pterin in aqueous solution. Photochem. Photobiol. 2013, 89, 1448–1455. [Google Scholar] [CrossRef]
- Plowman, J.E.; Deb-Choudhury, S.; Grosvenor, A.J.; Dyer, J.M. Protein oxidation: Identification and utilisation of molecular markers to differentiate singlet oxygen and hydroxyl radical-mediated oxidative pathways. Photochem. Photobiol. Sci. 2013, 12, 1960–1967. [Google Scholar] [CrossRef]
- Walrant, P.; Santus, R. N-formyl-kynurenine, a tryptophan photooxidation product, as a photodynamic sensitizer. Photochem. Photobiol. 1974, 19, 411–417. [Google Scholar] [CrossRef]
- Brem, R.; Macpherson, P.; Guven, M.; Karran, P. Oxidative stress induced by UVA photoactivation of the tryptophan UVB photoproduct 6-formylindolo[3,2-b]carbazole (FICZ) inhibits nucleotide excision repair in human cells. Sci. Rep. 2017, 7, 4310. [Google Scholar] [CrossRef]
- Kramarenko, G.G.; Hummel, S.G.; Martin, S.M.; Buettner, G.R. Ascorbate reacts with singlet oxygen to produce hydrogen peroxide. Photochem. Photobiol. 2007, 82, 1634–1637. [Google Scholar] [CrossRef]
- Bissett, D.L.; Chatterjee, R.; Hannon, D.P. Photoprotective effect of superoxide-scavenging antioxidants against ultraviolet radiation-induced chronic skin damage in the hairless mouse. Photodermatol. Photoimmunol. Photomed. 1990, 7, 56–62. [Google Scholar]
- Eli, M.; Li, D.-S.; Zhang, W.-W.; Kong, B.; Du, C.-S.; Wumar, M.; Mamtimin, B.; Sheyhidin, I.; Hasim, A. Decreased blood riboflavin levels are correlated with defective expression of RFT2 gene in gastric cancer. World J. Gastroenterol. 2012, 18, 3112–3118. [Google Scholar] [CrossRef] [PubMed]
- Aili, A.; Hasim, A.; Kelimu, A.; Guo, X.; Mamtimin, B.; Abudula, A.; Upur, H. Association of the plasma and tissue riboflavin levels with C20orf54 expression in cervical lesions and its relationship to HPV16 infection. PLoS ONE 2014, 7, e103377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiciner, E.R.; Haliman, R.S. Effect of alcohol on serum folate level. J. Clin. Investig. 1973, 3, 584–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Peng, D.; Liu, C.; Huang, C.; Luo, J. Serum high concentrations of homocysteine and low levels of folic acid and vitamin B12 are significantly correlated with the categories of coronary artery diseases. BMC Cardiovasc. Disord. 2017, 1, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichler, R.; Fritz, J.; Heidegger, I.; Steiner, E.; Culig, Z.; Klocker, H.; Fuchs, D. Predictive and prognostic role of serum neopterin and tryptophan breakdown in prostate cancer. Cancer Sci. 2017, 4, 663–670. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids (in DMEM) | (g/L) | Vitamins (in DMEM) | (g/L) |
|---|---|---|---|
| A1: L-Arginine·HCl | 0.084 | V1: Choline chloride | 0.004 |
| A2: L-Cystine·2HCl | 0.0626 | V2: Folic acid | 0.004 |
| A3: Glycine | 0.03 | V3: myo-Inositol | 0.0072 |
| A4: L-Histidine·HCl·H2O | 0.042 | V4: Niacin amide | 0.004 |
| A5: H-Isoleucine | 0.105 | V5: D-Pantothenic acid·1/2Ca | 0.004 |
| A6: L-Leucine | 0.105 | V6: Pyridoxine·HCl | 0.00404 |
| A7: H-Lysine·HCl | 0.146 | V7: Riboflavin | 0.0004 |
| A8: L-Methionine | 0.03 | V8: Thiamine·HCl | 0.004 |
| A9: L-Phenylalanine | 0.066 | ||
| A10: L-Serine | 0.042 | ||
| A11: L-Threonine | 0.095 | ||
| A12: L-Tryptophan | 0.016 | ||
| A13: L-Tyrosine·2Na·2H2O | 0.12037 | ||
| A14: L-Valine | 0.094 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshimoto, S.; Kohara, N.; Sato, N.; Ando, H.; Ichihashi, M. Riboflavin Plays a Pivotal Role in the UVA-Induced Cytotoxicity of Fibroblasts as a Key Molecule in the Production of H2O2 by UVA Radiation in Collaboration with Amino Acids and Vitamins. Int. J. Mol. Sci. 2020, 21, 554. https://doi.org/10.3390/ijms21020554
Yoshimoto S, Kohara N, Sato N, Ando H, Ichihashi M. Riboflavin Plays a Pivotal Role in the UVA-Induced Cytotoxicity of Fibroblasts as a Key Molecule in the Production of H2O2 by UVA Radiation in Collaboration with Amino Acids and Vitamins. International Journal of Molecular Sciences. 2020; 21(2):554. https://doi.org/10.3390/ijms21020554
Chicago/Turabian StyleYoshimoto, Satoshi, Nana Kohara, Natsu Sato, Hideya Ando, and Masamitsu Ichihashi. 2020. "Riboflavin Plays a Pivotal Role in the UVA-Induced Cytotoxicity of Fibroblasts as a Key Molecule in the Production of H2O2 by UVA Radiation in Collaboration with Amino Acids and Vitamins" International Journal of Molecular Sciences 21, no. 2: 554. https://doi.org/10.3390/ijms21020554
APA StyleYoshimoto, S., Kohara, N., Sato, N., Ando, H., & Ichihashi, M. (2020). Riboflavin Plays a Pivotal Role in the UVA-Induced Cytotoxicity of Fibroblasts as a Key Molecule in the Production of H2O2 by UVA Radiation in Collaboration with Amino Acids and Vitamins. International Journal of Molecular Sciences, 21(2), 554. https://doi.org/10.3390/ijms21020554