Overexpression of CA1 mRNA and the CA I Protein in Tumor Cells Does Not Change the Gene Expression of the ECM Proteins


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
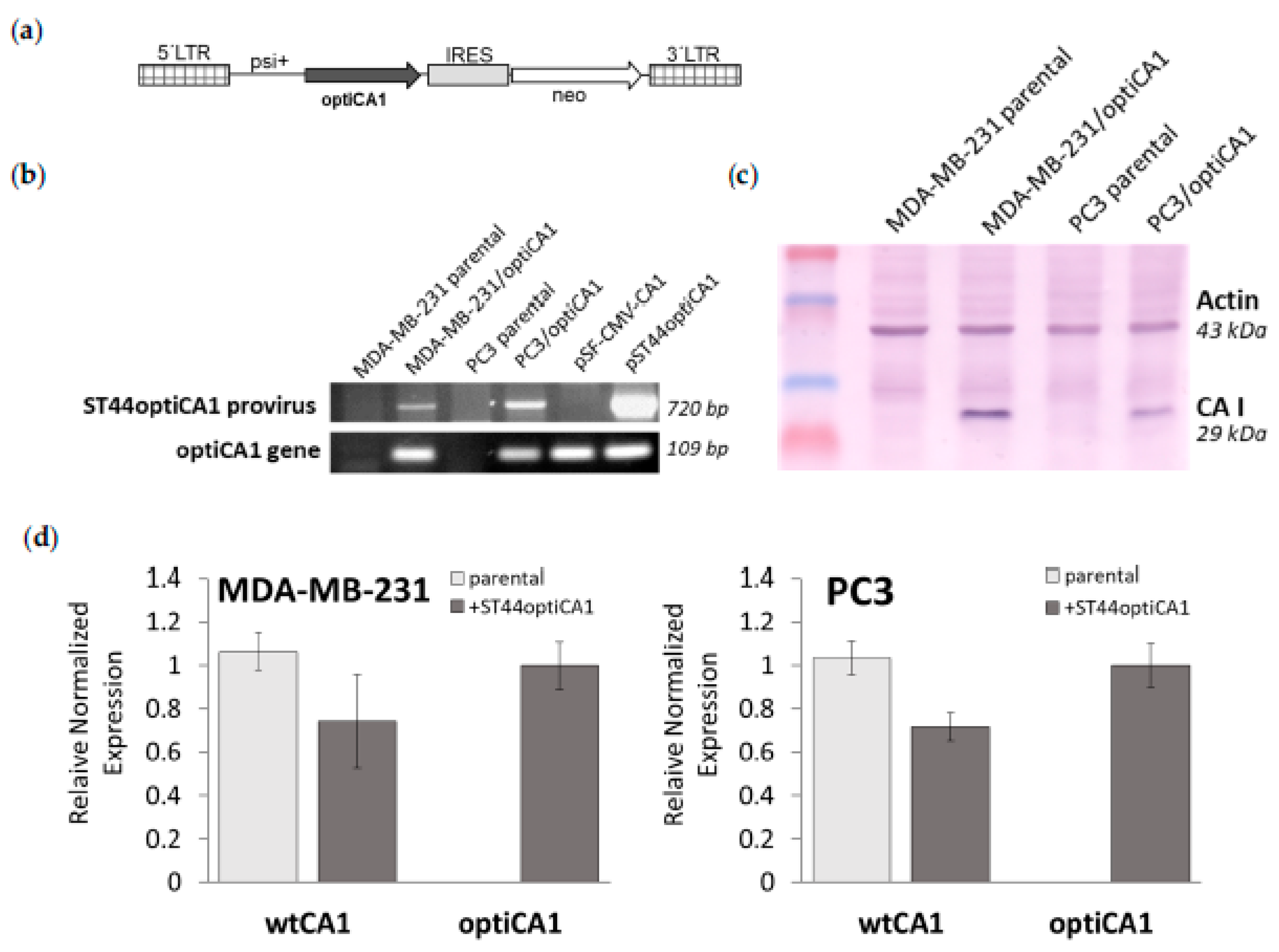
2.1. Overexpression of Codon Optimised Human CA1 Gene Reduces Endogenous CA1 Gene Expression but Does Not Change Expression of Genes Encoding Extracellular Matrix Proteins
2.2. Overexpression of Codon Optimised Human CA1 Gene Doesn’t Reduce Short Term Cell Proliferation
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Chemicals
4.2. Construction of Recombinant Retroviral Vector Containing Human CA1 Gene and Retrovirus Production
4.3. Western Blotting
4.4. Gene Expression Analysis
4.5. Kinetic Measurement of Cell Proliferation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Supuran, C.T. Carbonic Anhydrases and Metabolism. Metabolites 2018, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supuran, C.T. Structure-based drug discovery of carbonic anhydrase inhibitors. J. Enzyme Inhib. Med. Chem. 2012, 27, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Sly, W.S. Human Carbonic Anhydrases and Carbonic Anhydrase Deficiencies. Annu. Rev. Biochem. 1995, 64, 375–401. [Google Scholar] [CrossRef]
- Kendall, A.G.; Tashian, R.E. Erythrocyte carbonic anhydrase I: Inherited deficiency in humans. Science 1977, 197, 471–472. [Google Scholar] [CrossRef]
- Lakota, J.; Skultety, L.; Dubrovcakova, M.; Altaner, C. Presence of serum carbonic anhydrase autoantibodies in patients relapsed after autologous stem cell transplantation indicates an improved prognosis. Neoplasma 2008, 55, 488–492. [Google Scholar] [PubMed]
- Lakota, J.; Vranovsky, A.; Ladicka, M. Spontaneous regression of tumors relapsed following high dose chemotherapy and autologous stem cell transplantation: A relationship with an aplastic anemia type syndrome? Exp. Oncol. 2003, 25, 285–288. [Google Scholar]
- Lakota, J.; Vulic, R.; Dubrovcakova, M.; Tyciakova, S. Sera of patients with spontaneous tumour regression and elevated anti-CA I autoantibodies change the gene expression of ECM proteins. J. Cell Mol. Med. 2017, 21, 543–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyciakova, S.; Matuskova, M.; Bohovic, R.; Polakova, K.; Toro, L.; Skolekova, S.; Kucerova, L. Genetically engineered mesenchymal stromal cells producing TNFα have tumour suppressing effect on human melanoma xenograft. J. Gene Med. 2015, 17, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Vulic, R.; Tyciakova, S.; Dubrovcakova, M.; Skultety, L.; Lakota, J. Silencing of CA1 mRNA in tumour cells does not change the gene expression of the extracellular matrix proteins. J. Cell Mol. Med. 2018, 22, 695–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lakota, J.; Dubrovčáková, M. Overexpression of CA1 mRNA and the CA I Protein in Tumor Cells Does Not Change the Gene Expression of the ECM Proteins. Int. J. Mol. Sci. 2020, 21, 639. https://doi.org/10.3390/ijms21020639
Lakota J, Dubrovčáková M. Overexpression of CA1 mRNA and the CA I Protein in Tumor Cells Does Not Change the Gene Expression of the ECM Proteins. International Journal of Molecular Sciences. 2020; 21(2):639. https://doi.org/10.3390/ijms21020639
Chicago/Turabian StyleLakota, Ján, and Mária Dubrovčáková. 2020. "Overexpression of CA1 mRNA and the CA I Protein in Tumor Cells Does Not Change the Gene Expression of the ECM Proteins" International Journal of Molecular Sciences 21, no. 2: 639. https://doi.org/10.3390/ijms21020639
APA StyleLakota, J., & Dubrovčáková, M. (2020). Overexpression of CA1 mRNA and the CA I Protein in Tumor Cells Does Not Change the Gene Expression of the ECM Proteins. International Journal of Molecular Sciences, 21(2), 639. https://doi.org/10.3390/ijms21020639