Metalloporphyrin Pd(T4) Exhibits Oncolytic Activity and Cumulative Effects with 5-ALA Photodynamic Treatment against C918 Cells




Abstract
:1. Introduction
2. Results
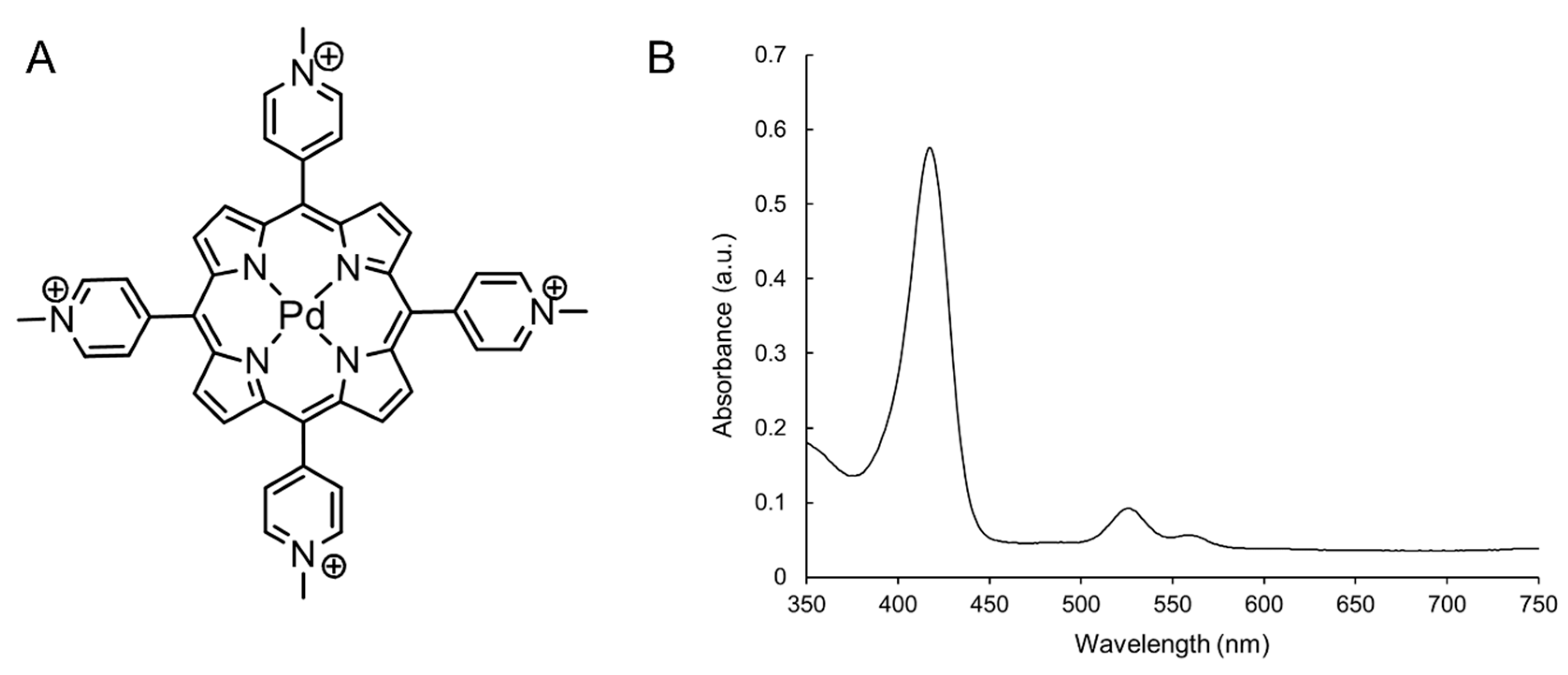
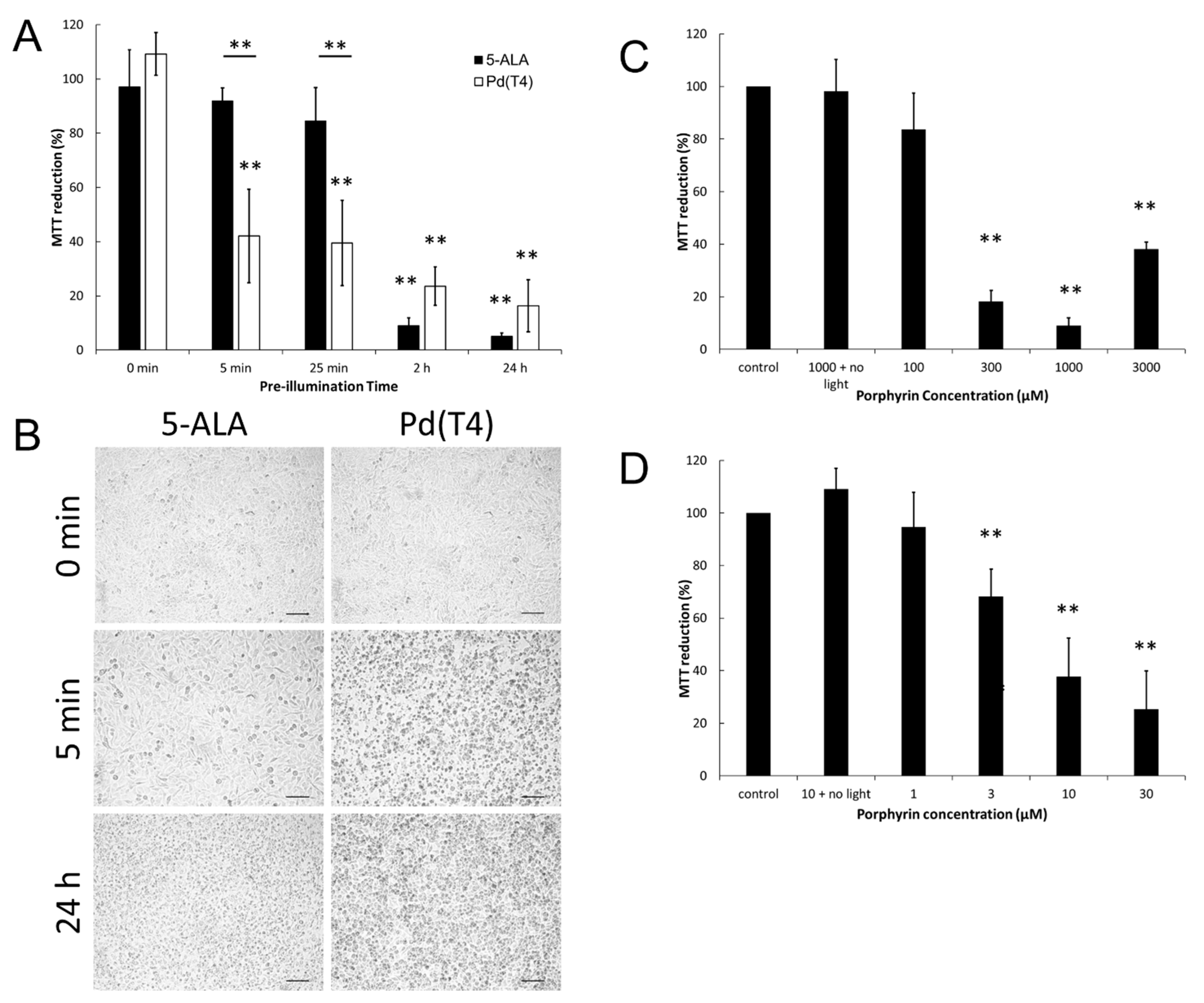
2.1. Oncolytic Effects of Photosensitizer Pd(T4) Compared to 5-ALA during PDT
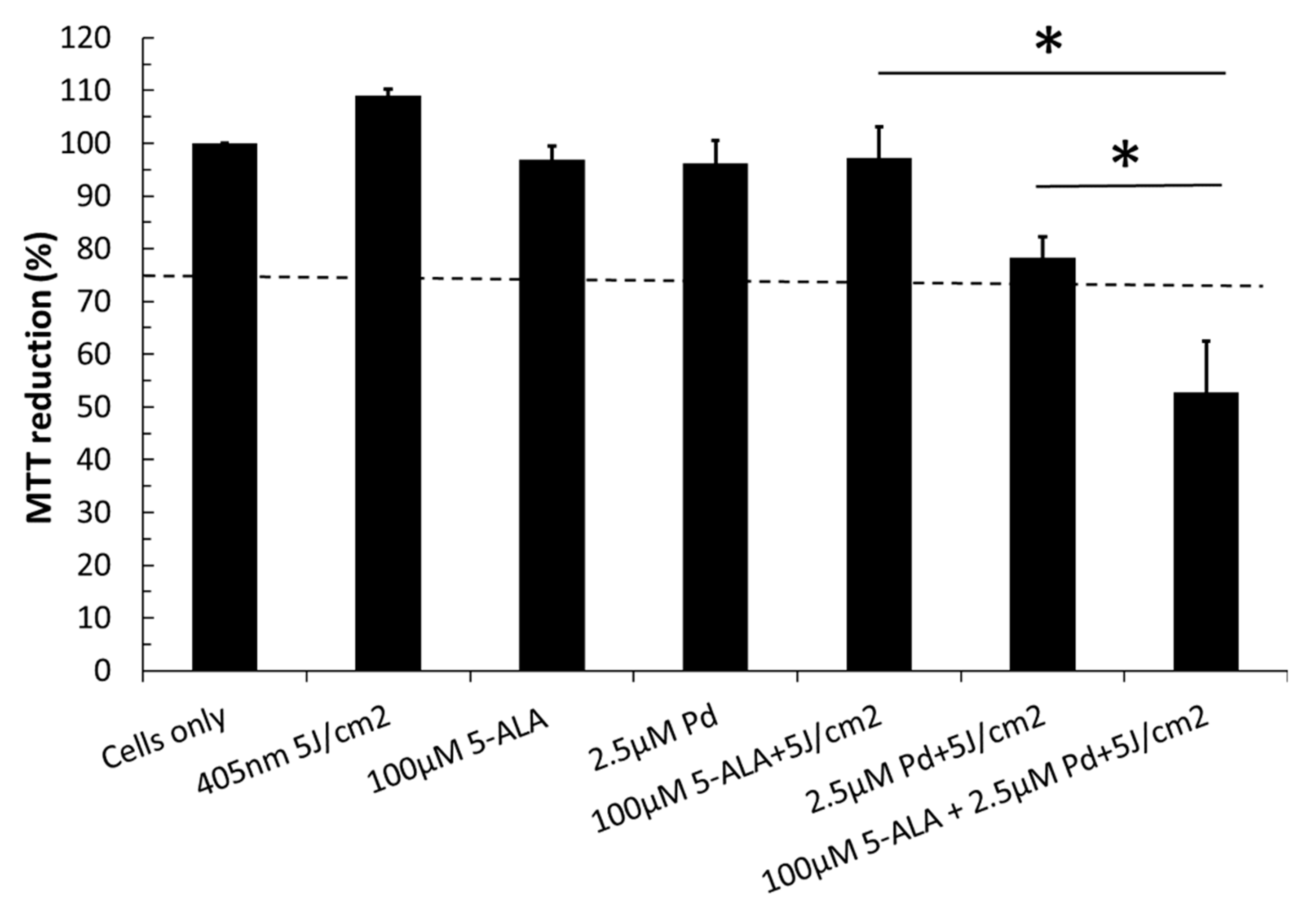
2.2. Combinatorial Effects of Two Different Photosensitizers
2.3. PDT on Three-Dimensional Cultures with Vasculogenic Mimicry-Forming Cells
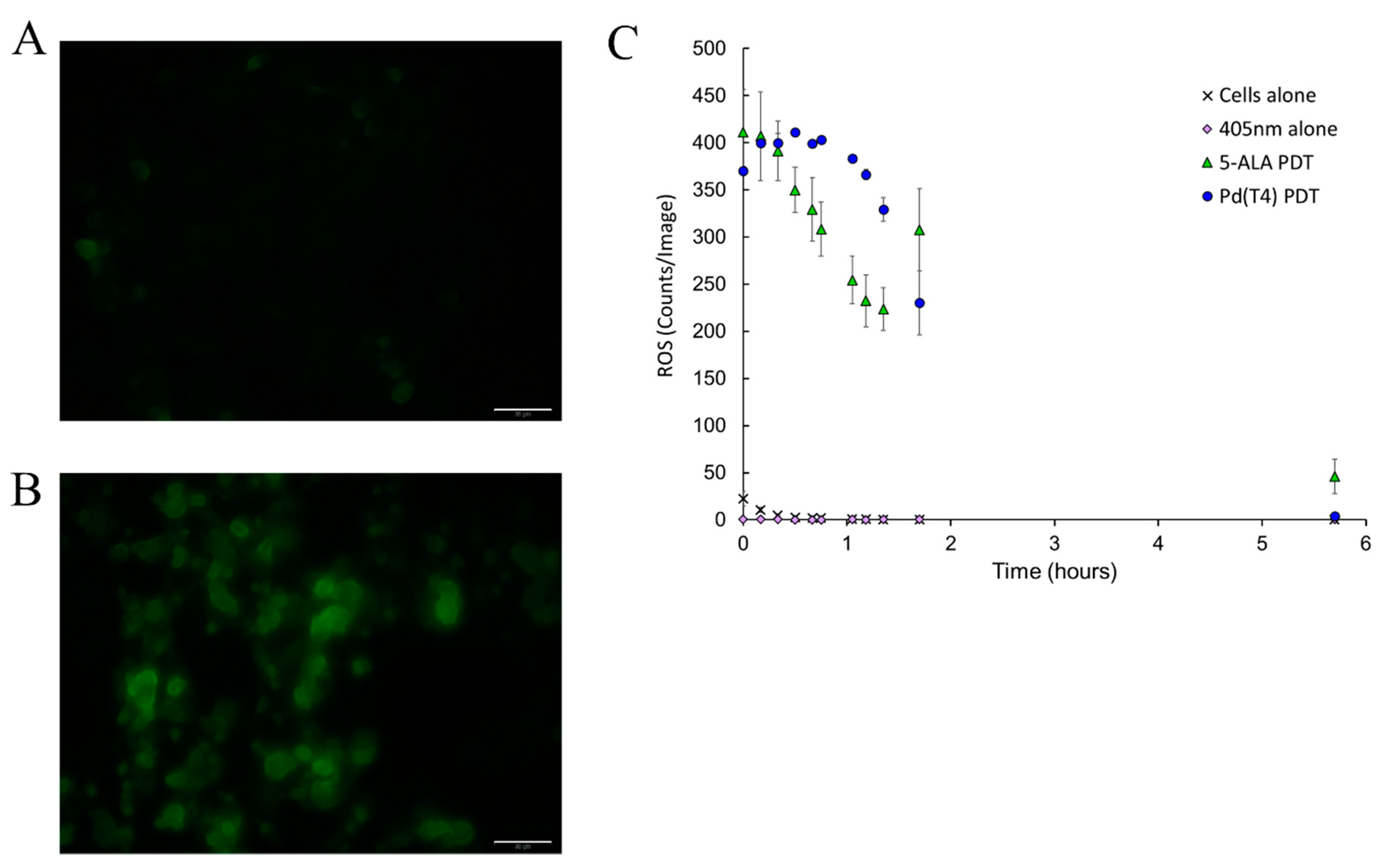
2.4. Oncolytic Activity Mediated through Reactive Oxygen Species
2.5. Macromolecular Interactions with Pd(T4)
2.6. Pd(T4)-PDT Induces Oncotic Necrosis
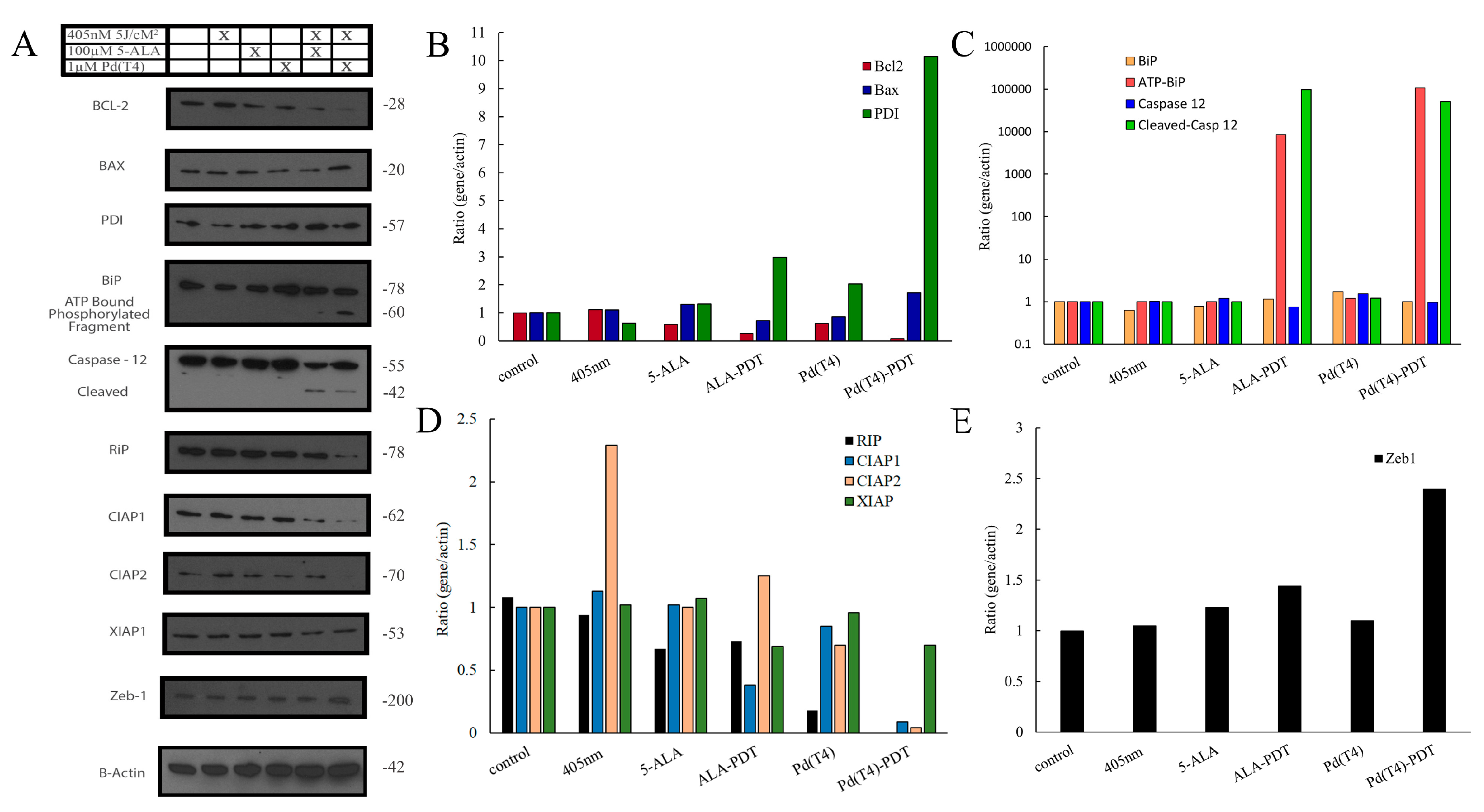
2.7. Anticancer Signaling Pathways Modulated by Pd(T4)-PDT Compared to ALA-PDT
3. Discussion
4. Materials and Methods
4.1. Cells and Media
4.2. Synthesis of Pd(T4) Photosensitizer
4.3. Cell Culture and Photodynamic Therapy
4.4. Assessment of Cell Viability Using MTT Reduction
Avg. OD of control cells
4.5. ROS Detection Assay
4.6. Propidium Iodide Staining
4.7. Molecular Interactions with Pd(T4)
4.8. SDS-PAGE and Western Blotting
4.9. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Raigani, S.; Cohen, S.; Boland, G.M. The Role of Surgery for Melanoma in an Era of Effective Systemic Therapy. Curr. Oncol. Rep. 2017, 19, 17. [Google Scholar] [CrossRef]
- Reinhardt, J.; Landsberg, J.; Schmid-Burgk, J.L.; Ramis, B.B.; Bald, T.; Glodde, N.; Lopez-Ramos, D.; Young, A.; Ngiow, S.F.; Nettersheim, D.; et al. MAPK Signaling and Inflammation Link Melanoma Phenotype Switching to Induction of CD73 during Immunotherapy. Cancer Res. 2017, 77, 4697–4709. [Google Scholar] [CrossRef] [Green Version]
- Ahn, A.; Chatterjee, A.; Eccles, M.R. The Slow Cycling Phenotype: A Growing Problem for Treatment Resistance in Melanoma. Mol. Cancer Ther. 2017, 16, 1002–1009. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.Y.; Menzies, A.M.; Rizos, H. Mechanisms and strategies to overcome resistance to molecularly targeted therapy for melanoma. Cancer 2017, 123, 2118–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, A.; Larkin, J. The safety of nivolumab for the treatment of metastatic melanoma. Expert Opin. Drug Saf. 2017, 16, 955–961. [Google Scholar] [CrossRef]
- Hao, C.; Tian, J.; Liu, H.; Li, F.; Niu, H.; Zhu, B. Efficacy and safety of anti-PD-1 and anti-PD-1 combined with anti-CTLA-4 immunotherapy to advanced melanoma: A systematic review and meta-analysis of randomized controlled trials. Medicine (Baltimore) 2017, 96, e7325. [Google Scholar] [CrossRef] [PubMed]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment A Review. Jama-J. Am. Med. Assoc. 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Rombouts, A.J.M.; Hugen, N.; Elferink, M.A.G.; Poortmans, P.M.P.; Nagtegaal, I.D.; de Wilt, J.H.W. Increased risk for second primary rectal cancer after pelvic radiation therapy. Eur. J. Cancer 2020, 124, 142–151. [Google Scholar] [CrossRef] [Green Version]
- Thunshelle, C.; Yin, R.; Chen, Q.Q.; Hamblin, M.R. Current Advances in 5-Aminolevulinic Acid Mediated Photodynamic Therapy. Curr. Dermatol. Rep. 2016, 5, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.; Li, Y.; Hamblin, M.R. Photodynamic therapy in dermatology beyond non-melanoma cancer: An update. Photodiagnosis Photodyn. Ther. 2017, 19, 140–152. [Google Scholar] [CrossRef]
- Holzer, G.; Pinkowicz, A.; Radakovic, S.; Schmidt, J.B.; Tanew, A. Randomized controlled trial comparing 35% trichloroacetic acid peel and 5-aminolaevulinic acid photodynamic therapy for treating multiple actinic keratosis. Br. J. Derm. Dermatol. 2017, 176, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Buinauskaite, E.; Maciulaitis, R.; Buinauskiene, J.; Valiukeviciene, S. Topical photodynamic therapy of actinic keratoses with 5-aminolevulinic acid: Randomized controlled trial with six months follow-up. J. Dermatol. Treat. 2014, 25, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.S.; Felicio, L.B.A.; Ferreira, J.; Kurachi, C.; Bentley, M.V.B.; Tedesco, A.C.; Bagnato, V.S. Long-term follow-up of topical 5-aminolaevulinic acid photodynamic therapy diode laser single session for non-melanoma skin cancer. Photodiagnosis Photodyn. Ther. 2009, 6, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Michael, A.P.; Watson, V.L.; Ryan, D.; Delfino, K.R.; Bekker, S.V.; Cozzens, J.W. Effects of 5-ALA dose on resection of glioblastoma. J. Neuro-Oncol. 2019, 141, 523–531. [Google Scholar] [CrossRef]
- Cordova, J.S.; Gurbani, S.S.; Holder, C.A.; Olson, J.J.; Schreibmann, E.; Shi, R.; Guo, Y.; Shu, H.K.G.; Shim, H.; Hadjipanayis, C.G. Semi-Automated Volumetric and Morphological Assessment of Glioblastoma Resection with Fluorescence-Guided Surgery. Mol. Imaging Biol. 2016, 18, 454–462. [Google Scholar] [CrossRef] [Green Version]
- Stummer, W.; Pichlmeier, U.; Meinel, T.; Wiestler, O.D.; Zanella, F.; Hans-Jurgen, R.; Ala-Glioma Study, G. Fluorescence-guided surgery with 5-aminolevulinic acid for resection of malignant glioma: A randomised controlled multicentre phase III trial. Lancet Oncol. 2006, 7, 392–401. [Google Scholar] [CrossRef]
- Kennedy, J.C.; Pottier, R.H.; Pross, D.C. Photodynamic therapy with endogenous protoporphyrin IX: Basic principles and present clinical experience. J. Photochem. Photobiol. B 1990, 6, 143–148. [Google Scholar] [CrossRef]
- de Souza, A.L.; Marra, K.; Gunn, J.; Samkoe, K.S.; Kanick, S.C.; Davis, S.C.; Chapman, M.S.; Maytin, E.V.; Hasan, T.; Pogue, B.W. Comparing desferrioxamine and light fractionation enhancement of ALA-PpIX photodynamic therapy in skin cancer. Br. J. Cancer 2016, 115, 805–813. [Google Scholar] [CrossRef] [Green Version]
- Krestyn, E.; Kolarova, H.; Bajgar, R.; Tomankova, K. Photodynamic properties of ZnTPPSClAlPcS(2) and ALA in human melanoma G361 cells. Toxicol In Vitro 2010, 24, 286–291. [Google Scholar] [CrossRef]
- Iinuma, S.; Farshi, S.S.; Ortel, B.; Hasan, T. A mechanistic study of cellular photodestruction with 5-aminolaevulinic acid-induced porphyrin. Br. J. Cancer 1994, 70, 21–28. [Google Scholar] [CrossRef]
- Hempstead, J.; Jones, D.P.; Ziouche, A.; Cramer, G.M.; Rizvi, I.; Arnason, S.; Hasan, T.; Celli, J.P. Low-cost photodynamic therapy devices for global health settings: Characterization of battery-powered LED performance and smartphone imaging in 3D tumor models. Sci. Rep. 2015, 5, 10093. [Google Scholar] [CrossRef] [Green Version]
- Cairnduff, F.; Stringer, M.R.; Hudson, E.J.; Ash, D.V.; Brown, S.B. Superficial photodynamic therapy with topical 5-aminolaevulinic acid for superficial primary and secondary skin cancer. Br. J. Cancer 1994, 69, 605–608. [Google Scholar] [CrossRef] [Green Version]
- Apalla, Z.; Sotiriou, E.; Chovarda, E.; Lefaki, I.; Devliotou-Panagiotidou, D.; Ioannides, D. Skin cancer: Preventive photodynamic therapy in patients with face and scalp cancerization. A randomized placebo-controlled study. Br. J. Derm. 2010, 162, 171–175. [Google Scholar] [CrossRef]
- Warren, C.B.; Karai, L.J.; Vidimos, A.; Maytin, E.V. Pain associated with aminolevulinic acid-photodynamic therapy of skin disease. J. Am. Acad. Derm. 2009, 61, 1033–1043. [Google Scholar] [CrossRef] [Green Version]
- Ericson, M.B.; Grapengiesser, S.; Gudmundson, F.; Wennberg, A.M.; Larko, O.; Moan, J.; Rosen, A. A spectroscopic study of the photobleaching of protoporphyrin IX in solution. Lasers Med. Sci. 2003, 18, 56–62. [Google Scholar] [CrossRef]
- Markwardt, N.A.; Haj-Hosseini, N.; Hollnburger, B.; Stepp, H.; Zelenkov, P.; Ruhm, A. 405 nm versus 633 nm for protoporphyrin IX excitation in fluorescence-guided stereotactic biopsy of brain tumors. J. Biophotonics 2016, 9, 901–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skwor, T.A.; Klemm, S.; Zhang, H.; Schardt, B.; Blaszczyk, S.; Bork, M.A. Photodynamic inactivation of methicillin-resistant Staphylococcus aureus and Escherichia coli: A metalloporphyrin comparison. J. Photochem. Photobiol. B 2016, 165, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Lebedeva, N.S.; Yurina, E.S.; Gubarev, Y.A.; Syrbu, S.A. Interactions of tetracationic porphyrins with DNA and their effects on DNA cleavage. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2018, 199, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Ruan, T.L.; Davis, S.J.; Powell, B.M.; Harbeck, C.P.; Habdas, J.; Habdas, P.; Yatsunyk, L.A. Lowering the overall charge on TMPyP4 improves its selectivity for G-quadruplex DNA. Biochimie 2017, 132, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Sabharwal, N.C.; Mendoza, O.; Nicoludis, J.M.; Ruan, T.; Mergny, J.L.; Yatsunyk, L.A. Investigation of the interactions between Pt(II) and Pd(II) derivatives of 5,10,15,20-tetrakis (N-methyl-4-pyridyl) porphyrin and G-quadruplex DNA. J. Biol. Inorg Chem. 2016, 21, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Cenklova, V. Photodynamic therapy with TMPyP-Porphyrine induces mitotic catastrophe and microtubule disorganization in HeLa and G361 cells, a comprehensive view of the action of the photosensitizer. J. Photochem. Photobiol. B 2017, 173, 522–537. [Google Scholar] [CrossRef]
- Liu, H.; Lv, C.; Ding, B.; Wang, J.; Li, S.; Zhang, Y. Antitumor activity of G-quadruplex-interactive agent TMPyP4 with photodynamic therapy in ovarian carcinoma cells. Oncol Lett. 2014, 8, 409–413. [Google Scholar] [CrossRef] [Green Version]
- Cohen, D.K.; Lee, P.K. Photodynamic Therapy for Non-Melanoma Skin Cancers. Cancers (Basel) 2016, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Maytin, E.V.; Kaw, U.; Ilyas, M.; Mack, J.A.; Hu, B. Blue light versus red light for photodynamic therapy of basal cell carcinoma in patients with Gorlin syndrome: A bilaterally controlled comparison study. Photodiagnosis Photodyn. Ther. 2018, 22, 7–13. [Google Scholar] [CrossRef]
- Valyi-Nagy, K.; Kormos, B.; Ali, M.; Shukla, D.; Valyi-Nagy, T. Stem cell marker CD271 is expressed by vasculogenic mimicry-forming uveal melanoma cells in three-dimensional cultures. Mol. Vis. 2012, 18, 588–592. [Google Scholar]
- Oberst, A. Death in the fast lane: What’s next for necroptosis? Febs J. 2016, 283, 2616–2625. [Google Scholar] [CrossRef]
- Krebs, A.M.; Mitschke, J.; Lasierra Losada, M.; Schmalhofer, O.; Boerries, M.; Busch, H.; Boettcher, M.; Mougiakakos, D.; Reichardt, W.; Bronsert, P.; et al. The EMT-activator Zeb1 is a key factor for cell plasticity and promotes metastasis in pancreatic cancer. Nat. Cell Biol. 2017, 19, 518–529. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; He, S.F.; Liu, L.L.; Sun, X.X.; Yang, F.; Ge, Q.; Wong, W.K.; Meng, J.Y. Potential role of ZEB1 as a DNA repair regulator in colorectal cancer cells revealed by cancer-associated promoter profiling. Oncol. Rep. 2017, 38, 1941–1948. [Google Scholar] [CrossRef]
- Pizova, K.; Bajgar, R.; Fillerova, R.; Kriegova, E.; Cenklova, V.; Langova, K.; Konecny, P.; Kolarova, H. C-MYC and C-FOS expression changes and cellular aspects of the photodynamic reaction with photosensitizers TMPyP and ClAlPcS2. J. Photochem. Photobiol. B 2015, 142, 186–196. [Google Scholar] [CrossRef]
- Nakayama, T.; Otsuka, S.; Kobayashi, T.; Okajima, H.; Matsumoto, K.; Hagiya, Y.; Inoue, K.; Shuin, T.; Nakajima, M.; Tanaka, T.; et al. Dormant cancer cells accumulate high protoporphyrin IX levels and are sensitive to 5-aminolevulinic acid-based photodynamic therapy. Sci. Rep. 2016, 6, 36478. [Google Scholar] [CrossRef] [Green Version]
- Rapozzi, V.; Zorzet, S.; Zacchigna, M.; Della Pietra, E.; Cogoi, S.; Xodo, L.E. Anticancer activity of cationic porphyrins in melanoma tumour-bearing mice and mechanistic in vitro studies. Mol. Cancer 2014, 13, 75. [Google Scholar] [CrossRef] [Green Version]
- Patito, I.A.; Rothmann, C.; Malik, Z. Nuclear transport of photosensitizers during photosensitization and oxidative stress. Biol. Cell 2001, 93, 285–291. [Google Scholar] [CrossRef]
- Ricchelli, F.; Franchi, L.; Miotto, G.; Borsetto, L.; Gobbo, S.; Nikolov, P.; Bommer, J.C.; Reddi, E. Meso-substituted tetra-cationic porphyrins photosensitize the death of human fibrosarcoma cells via lysosomal targeting. Int. J. Biochem. Cell B 2005, 37, 306–319. [Google Scholar] [CrossRef]
- Kawai, C.; Araujo-Chaves, J.C.; Magrini, T.; Sanches, C.; Pinto, S.M.S.; Martinho, H.; Daghastanli, N.; Nantes, I.L. Photodamage in a Mitochondrial Membrane Model Modulated by the Topology of Cationic and Anionic Meso-Tetrakis Porphyrin Free Bases. Photochem. Photobiol. 2014, 90, 596–608. [Google Scholar] [CrossRef]
- Khan, A.Q.; Kuttikrishnan, S.; Siveen, K.S.; Prabhu, K.S.; Shanmugakonar, M.; Al-Naemi, H.A.; Haris, M.; Dermime, S.; Uddin, S. RAS-mediated oncogenic signaling pathways in human malignancies. Semin. Cancer Biol. 2019, 54, 1–13. [Google Scholar] [CrossRef]
- Lei, W.H.; Jiang, G.Y.; Zhou, Q.X.; Zhang, B.W.; Wang, X.S. Greatly enhanced binding of a cationic porphyrin towards bovine serum albumin by cucurbit 8 uril. Phys. Chem. Chem. Phys. 2010, 12, 13255–13260. [Google Scholar] [CrossRef]
- Acedo, P.; Stockert, J.C.; Canete, M.; Villanueva, A. Two combined photosensitizers: A goal for more effective photodynamic therapy of cancer. Cell Death Dis. 2014, 5, e1122. [Google Scholar] [CrossRef] [Green Version]
- Zamiri, B.; Reddy, K.; Macgregor, R.B., Jr.; Pearson, C.E. TMPyP4 porphyrin distorts RNA G-quadruplex structures of the disease-associated r(GGGGCC)n repeat of the C9orf72 gene and blocks interaction of RNA-binding proteins. J. Biol. Chem. 2014, 289, 4653–4659. [Google Scholar] [CrossRef] [Green Version]
- Mikami-Terao, Y.; Akiyama, M.; Yuza, Y.; Yanagisawa, T.; Yamada, O.; Kawano, T.; Agawa, M.; Ida, H.; Yamada, H. Antitumor activity of TMPyP4 interacting G-quadruplex in retinoblastoma cell lines. Exp. Eye Res. 2009, 89, 200–208. [Google Scholar] [CrossRef]
- Tahmasebi, H.; Khoshgard, K.; Sazgarnia, A.; Mostafaie, A.; Eivazi, M.T. Enhancing the efficiency of 5-aminolevulinic acid-mediated photodynamic therapy using 5-fluorouracil on human melanoma cells. Photodiagnosis Photodyn. Ther. 2016, 13, 297–302. [Google Scholar] [CrossRef]
- Vanden Berghe, T.; Vanlangenakker, N.; Parthoens, E.; Deckers, W.; Devos, M.; Festjens, N.; Guerin, C.J.; Brunk, U.T.; Declercq, W.; Vandenabeele, P. Necroptosis, necrosis and secondary necrosis converge on similar cellular disintegration features. Cell Death Differ. 2010, 17, 922–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, J.M.; Cory, S. The BCL-2 arbiters of apoptosis and their growing role as cancer targets. Cell Death Differ. 2018, 25, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyna, D.E.; Gavathiotis, E. Pulling the BAX trigger for tumor cell death. Oncotarget 2018, 9, 8204–8205. [Google Scholar] [CrossRef] [PubMed]
- Raisova, M.; Hossini, A.M.; Eberle, J.; Riebeling, C.; Wieder, T.; Sturm, I.; Daniel, P.T.; Orfanos, C.E.; Geilen, C.C. The Bax/Bcl-2 ratio determines the susceptibility of human melanoma cells to CD95/Fas-mediated apoptosis. J. Investig. Derm. 2001, 117, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.P.; Lu, H.Y.; Li, C. Proapoptotic Activities of Protein Disulfide Isomerase (PDI) and PDIA3 Protein, a Role of the Bcl-2 Protein Bak. J. Biol. Chem. 2015, 290, 8949–8963. [Google Scholar] [CrossRef] [Green Version]
- Lovat, P.E.; Corazzari, M.; Armstrong, J.L.; Martin, S.; Pagliarini, V.; Hill, D.; Brown, A.M.; Piacentini, M.; Birch-Machin, M.A.; Redfern, C.P. Increasing melanoma cell death using inhibitors of protein disulfide isomerases to abrogate survival responses to endoplasmic reticulum stress. Cancer Res. 2008, 68, 5363–5369. [Google Scholar] [CrossRef] [Green Version]
- Ellgaard, L.; Ruddock, L.W. The human protein disulphide isomerase family: Substrate interactions and functional properties. Embo. Rep. 2005, 6, 28–32. [Google Scholar] [CrossRef]
- Ko, H.S.; Uehara, T.; Nomura, Y. Role of ubiquilin associated with protein-disulfide isomerase in the endoplasmic reticulum in stress-induced apoptotic cell death. J. Biol. Chem. 2002, 277, 35386–35392. [Google Scholar] [CrossRef] [Green Version]
- N’Diaye, E.N.; Kajihara, K.K.; Hsieh, I.; Morisaki, H.; Debnath, J.; Brown, E.J. PLIC proteins or ubiquilins regulate autophagy-dependent cell survival during nutrient starvation. Embo Rep. 2009, 10, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Morales, N.; Quiroga-Roger, D.; Alfaro-Valdes, H.M.; Alavi, Z.; Lagos-Espinoza, M.I.A.; Zocchi, G.; Wilson, C.A.M. Mechanical properties of BiP protein determined by nano-rheology. Protein Sci. 2018, 27, 1418–1426. [Google Scholar] [CrossRef] [Green Version]
- McClellan, A.J.; Endres, J.B.; Vogel, J.P.; Palazzi, D.; Rose, M.D.; Brodsky, J.L. Specific molecular chaperone interactions and an ATP-dependent conformational change are required during posttranslational protein translocation into the yeast ER. Mol. Biol. Cell 1998, 9, 3533–3545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muaddi, H.; Majumder, M.; Peidis, P.; Papadakis, A.I.; Holcik, M.; Scheuner, D.; Kaufman, R.J.; Hatzoglou, M.; Koromilas, A.E. Phosphorylation of eIF2alpha at serine 51 is an important determinant of cell survival and adaptation to glucose deficiency. Mol. Biol. Cell 2010, 21, 3220–3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Liu, C.; Zhang, D.; Yao, X.; Zhang, K.; Li, H.; Chen, G. Role for RIP1 in mediating necroptosis in experimental intracerebral hemorrhage model both in vivo and in vitro. Cell Death Dis. 2017, 8, e2641. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chen, W.; Bai, L.; Chen, W.; Padilla, M.T.; Lin, A.S.; Shi, S.; Wang, X.; Lin, Y. Receptor-interacting protein 1 increases chemoresistance by maintaining inhibitor of apoptosis protein levels and reducing reactive oxygen species through a microRNA-146a-mediated catalase pathway. J. Biol. Chem. 2014, 289, 5654–5663. [Google Scholar] [CrossRef] [Green Version]
- Varfolomeev, E.; Goncharov, T.; Fedorova, A.V.; Dynek, J.N.; Zobel, K.; Deshayes, K.; Fairbrother, W.J.; Vucic, D. c-IAP1 and c-IAP2 are critical mediators of tumor necrosis factor alpha (TNFalpha)-induced NF-kappaB activation. J. Biol. Chem. 2008, 283, 24295–24299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.Q.; Webster, J.D.; Dugger, D.L.; Goncharov, T.; Roose-Girma, M.; Hung, J.; Kwon, Y.C.; Vucic, D.; Newton, K.; Dixit, V.M. Ubiquitin Ligases cIAP1 and cIAP2 Limit Cell Death to Prevent Inflammation. Cell Rep. 2019, 27, 2679–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vucic, D.; Deshayes, K.; Ackerly, H.; Pisabarro, M.T.; Kadkhodayan, S.; Fairbrother, W.J.; Dixit, V.M. SMAC negatively regulates the anti-apoptotic activity of melanoma inhibitor of apoptosis (ML-IAP). J. Biol. Chem. 2002, 277, 12275–12279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Lu, X.; Montoya-Durango, D.E.; Liu, Y.H.; Dean, K.C.; Darling, D.S.; Kaplan, H.J.; Dean, D.C.; Gao, L.; Liu, Y. ZEB1 Regulates Multiple Oncogenic Components Involved in Uveal Melanoma Progression. Sci. Rep. 2017, 7, 45. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wei, Y.; Wang, L.; Debeb, B.G.; Yuan, Y.; Zhang, J.S.; Yuan, J.S.; Wang, M.; Chen, D.H.; Sun, Y.T.; et al. ATM-mediated stabilization of ZEB1 promotes DNA damage response and radioresistance through CH K1. Nat. Cell Biol. 2014, 16, 864–875. [Google Scholar] [CrossRef]
- Li, M.D.; Wong, N.K.; Xiao, J.; Zhu, R.X.; Wu, L.L.; Dai, S.Y.; Chen, F.; Huang, G.H.; Xu, L.; Bai, X.Y.; et al. Dynamics of Oxygen-Independent Photocleavage of Blebbistatin as a One-Photon Blue or Two-Photon Near-Infrared Light-Gated Hydroxyl Radical Photocage. J. Am. Chem. Soc. 2018, 140, 15957–15968. [Google Scholar] [CrossRef] [Green Version]
- Hamblin, M.R. Upconversion in photodynamic therapy: Plumbing the depths. Dalton Trans. 2018, 47, 8571–8580. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Z.H.; Zha, S.; Zhu, Y.Y.; Wu, P.Y.; Ehrenberg, B.; Chen, J.Y. Carbon nanodots featuring efficient FRET for two-photon photodynamic cancer therapy with a low fs laser power density. Biomaterials 2014, 35, 9372–9381. [Google Scholar] [CrossRef] [PubMed]
- Shieh, Y.A.; Yang, S.J.; Wei, M.F.; Shieh, M.J. Aptamer-Based Tumor-Targeted Drug Delivery for Photodynamic Therapy. Acs Nano 2010, 4, 1433–1442. [Google Scholar] [CrossRef]
- Folberg, R.; Kadkol, S.S.; Frenkel, S.; Valyi-Nagy, K.; Jager, M.J.; Pe’er, J.; Maniotis, A.J. Authenticating cell lines in ophthalmic research laboratories. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4697–4701. [Google Scholar] [CrossRef]
- Bork, M.A.; Gianopoulos, C.G.; Zhang, H.; Fanwick, P.E.; Choi, J.H.; McMillin, D.R. Accessibility and External versus Intercalative Binding to DNA As Assessed by Oxygen-Induced Quenching of the Palladium(II)-Containing Cationic Porphyrins Pd(T4) and Pd(tD4). Biochemistry 2014, 53, 714–724. [Google Scholar] [CrossRef]
- Lange, C.; Bednarski, P.J. Evaluation for Synergistic Effects by Combinations of Photodynamic Therapy (PDT) with Temoporfin (mTHPC) and Pt(II) Complexes Carboplatin, Cisplatin or Oxaliplatin in a Set of Five Human Cancer Cell Lines. Int. J. Mol. Sci. 2018, 19, 3183. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Dosage (J/cm2) | % death in 3D culture | ||
|---|---|---|---|---|
| Top of Matrigel | VM | Bottom of Plate | ||
| No PDT | 0 | <3% | <3% | <3% |
| 100 M Pd(T4) | 0 | <3% | <3% | <3% |
| 30 M Pd(T4) | 10 | 5% | <3% | <3% |
| 30mM Pd(T4) | 15 | 5% | <3% | 40% |
| 100 M Pd(T4) | 10 | 20% | <3% | 50% |
| 100 M Pd(T4) | 15 | 60% | 50% | 90% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leviskas, B.; Valyi-Nagy, T.; Munirathinam, G.; Bork, M.; Valyi-Nagy, K.; Skwor, T. Metalloporphyrin Pd(T4) Exhibits Oncolytic Activity and Cumulative Effects with 5-ALA Photodynamic Treatment against C918 Cells. Int. J. Mol. Sci. 2020, 21, 669. https://doi.org/10.3390/ijms21020669
Leviskas B, Valyi-Nagy T, Munirathinam G, Bork M, Valyi-Nagy K, Skwor T. Metalloporphyrin Pd(T4) Exhibits Oncolytic Activity and Cumulative Effects with 5-ALA Photodynamic Treatment against C918 Cells. International Journal of Molecular Sciences. 2020; 21(2):669. https://doi.org/10.3390/ijms21020669
Chicago/Turabian StyleLeviskas, Brandon, Tibor Valyi-Nagy, Gnanasekar Munirathinam, Matthew Bork, Klara Valyi-Nagy, and Troy Skwor. 2020. "Metalloporphyrin Pd(T4) Exhibits Oncolytic Activity and Cumulative Effects with 5-ALA Photodynamic Treatment against C918 Cells" International Journal of Molecular Sciences 21, no. 2: 669. https://doi.org/10.3390/ijms21020669
APA StyleLeviskas, B., Valyi-Nagy, T., Munirathinam, G., Bork, M., Valyi-Nagy, K., & Skwor, T. (2020). Metalloporphyrin Pd(T4) Exhibits Oncolytic Activity and Cumulative Effects with 5-ALA Photodynamic Treatment against C918 Cells. International Journal of Molecular Sciences, 21(2), 669. https://doi.org/10.3390/ijms21020669