The Effect of Allopregnanolone on Enzymatic Activity of the DNA Base Excision Repair Pathway in the Sheep Hippocampus and Amygdala under Natural and Stressful Conditions



Abstract
:1. Introduction
2. Results
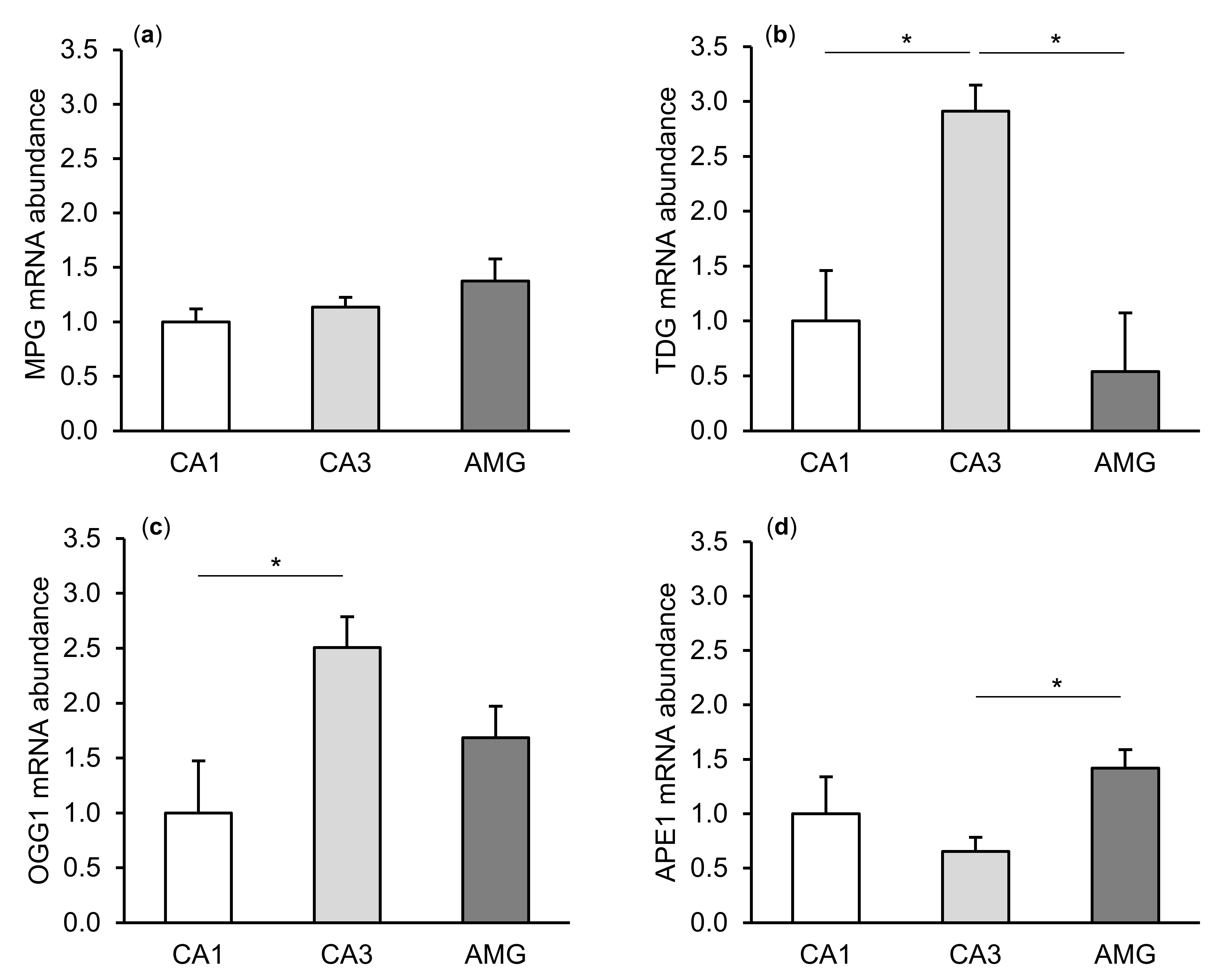
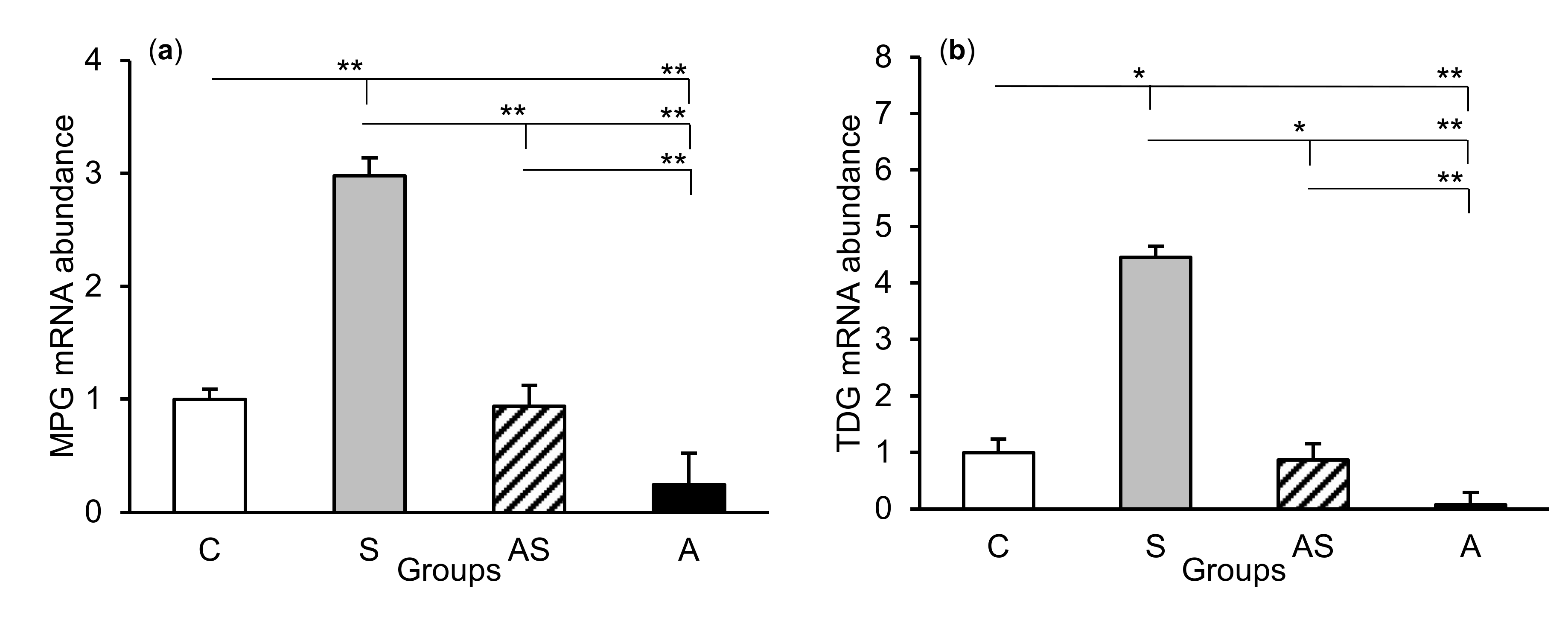
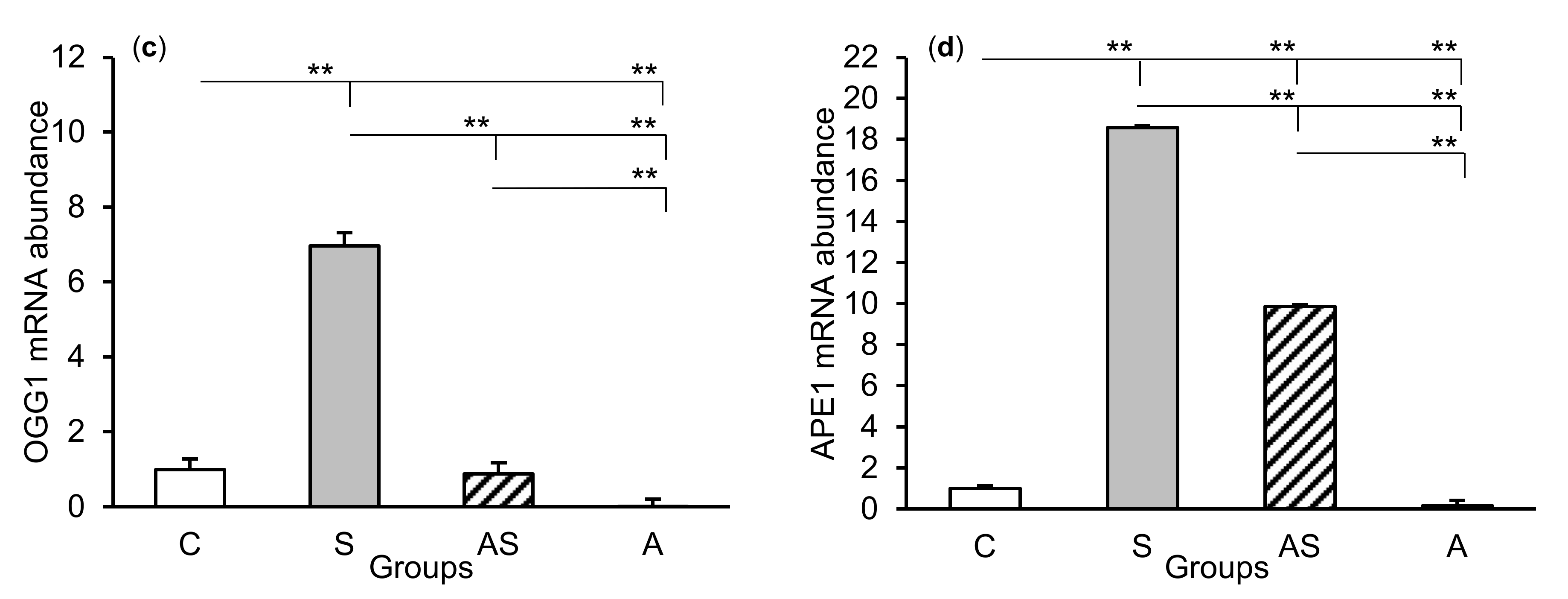
2.1. mRNA Abundance of DNA Repair Enzymes
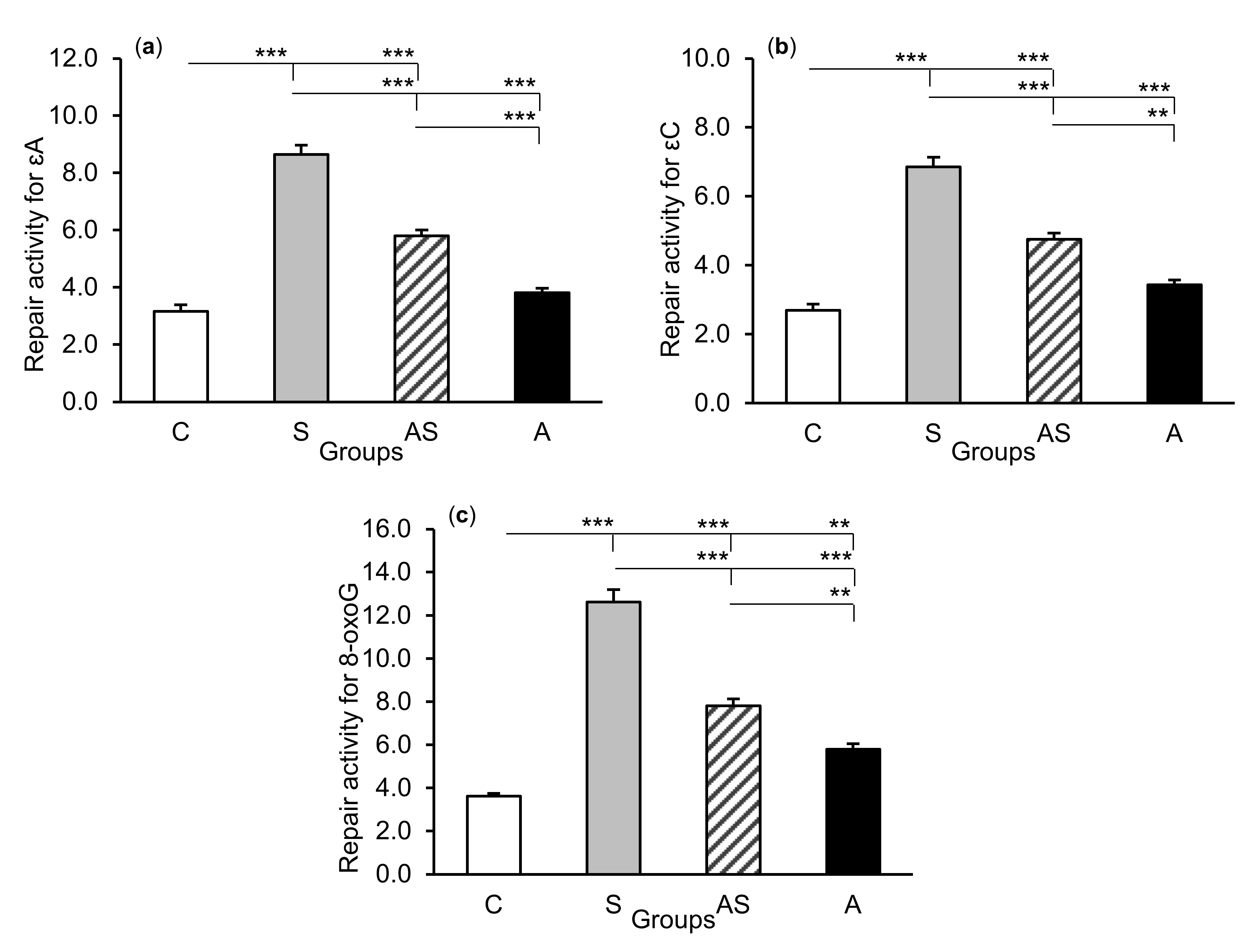
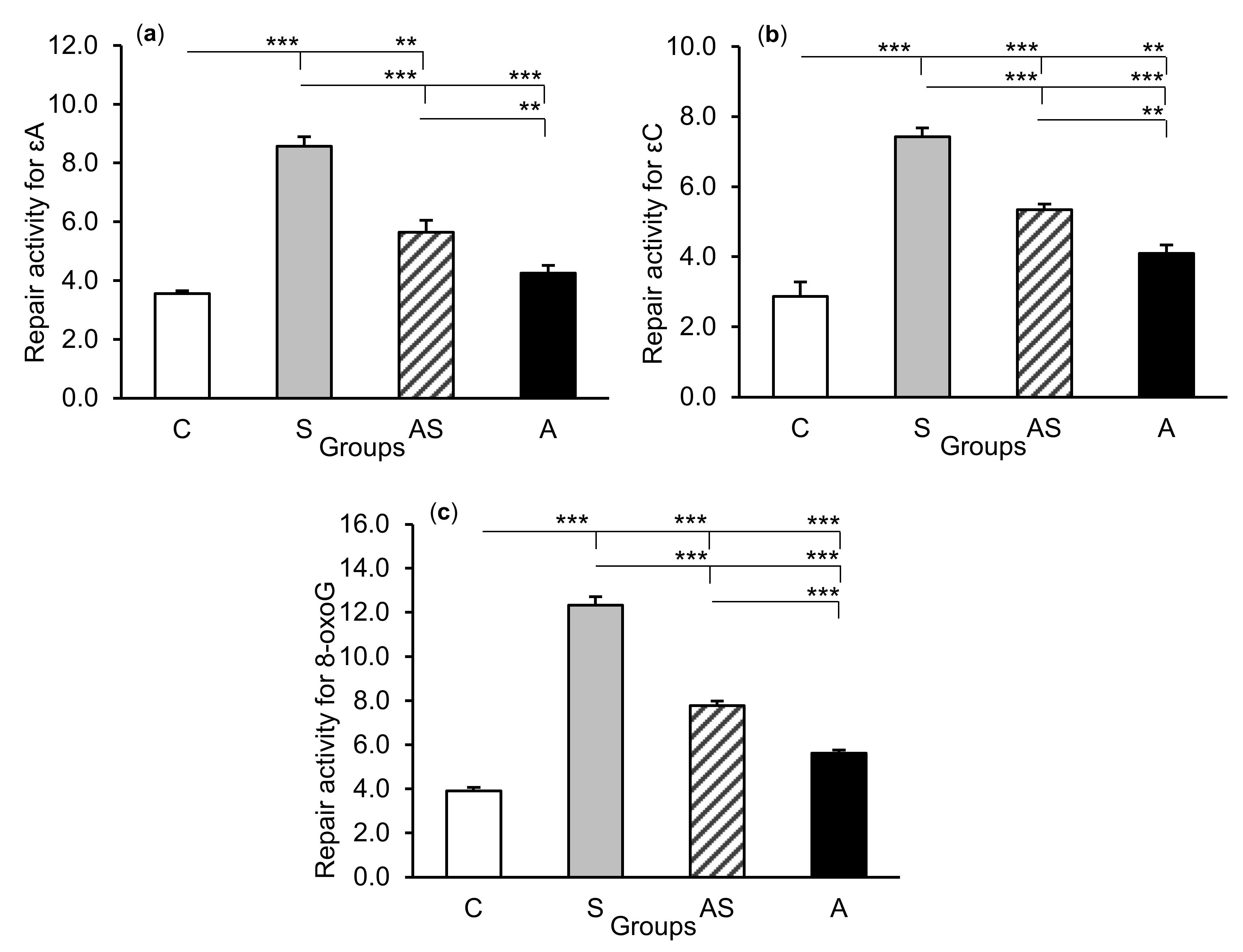
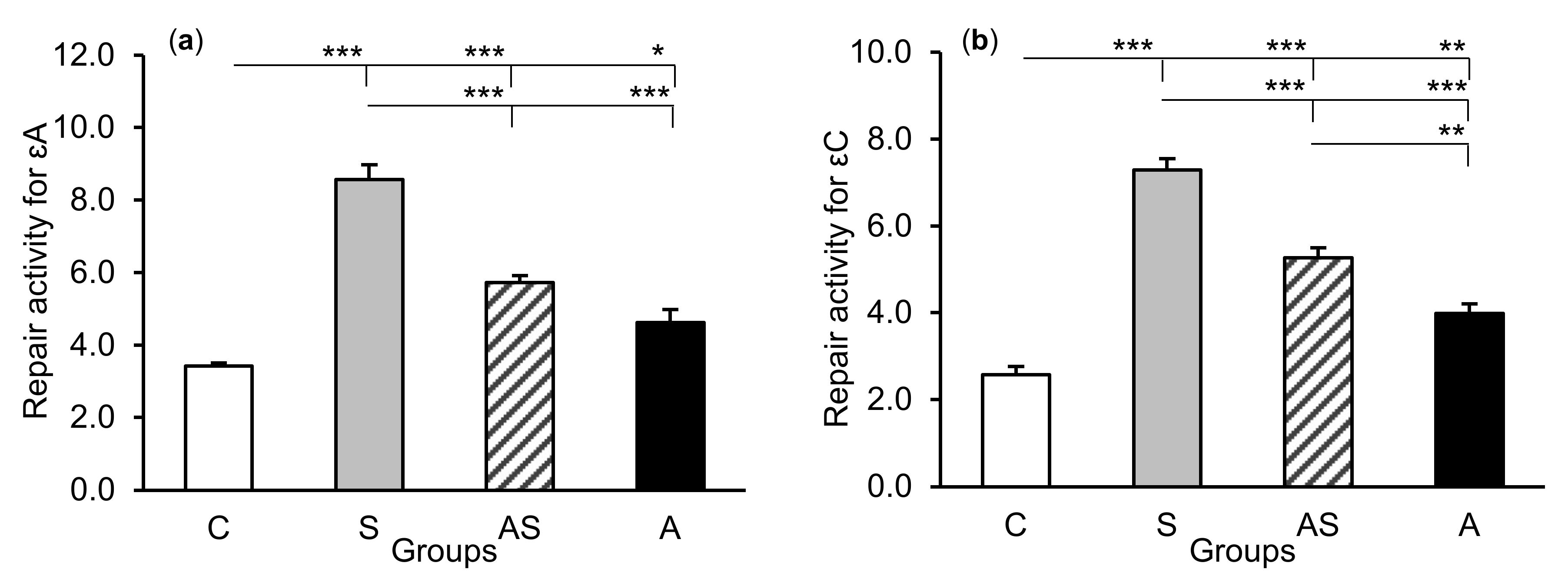
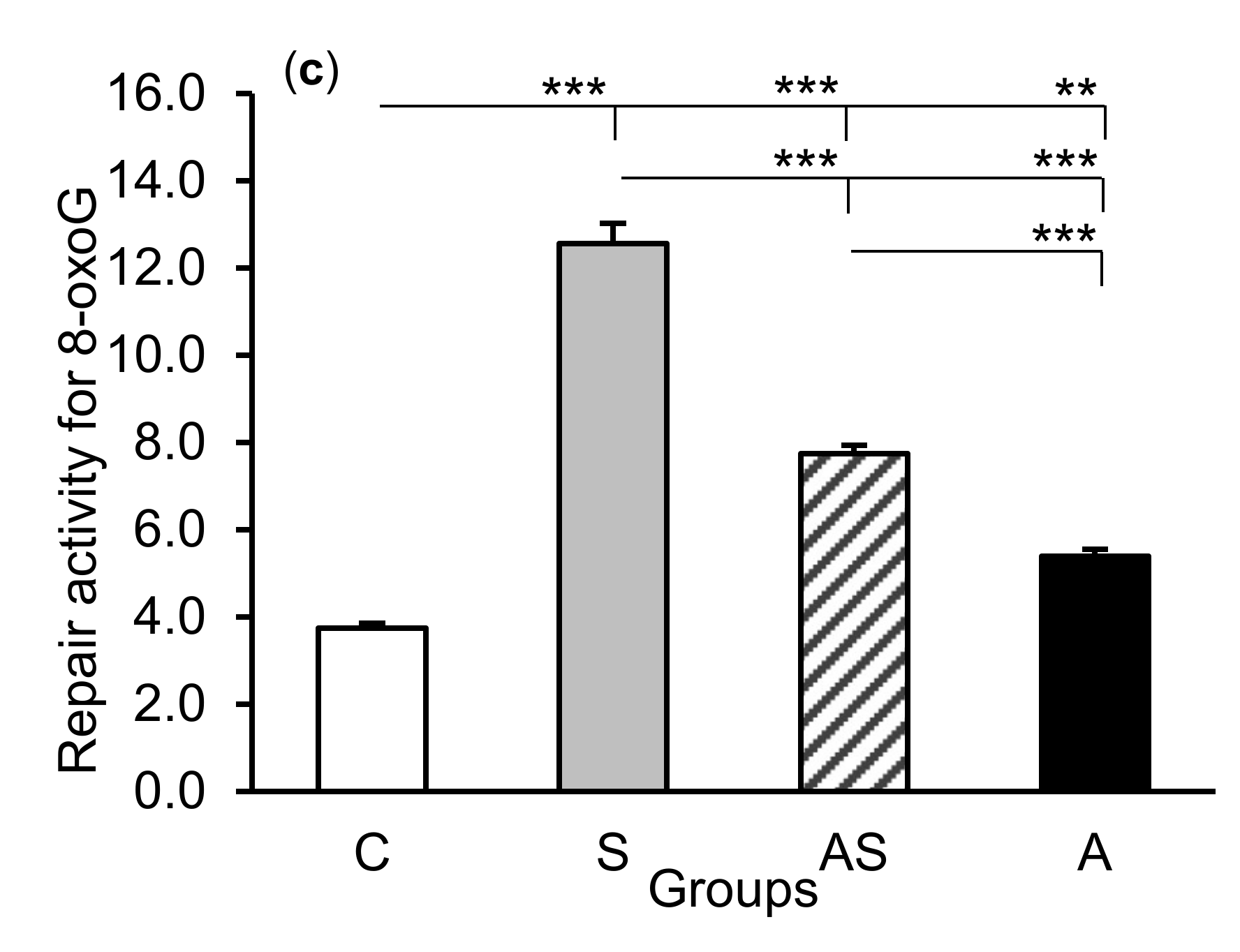
2.2. Enzyme Repair Activity
3. Discussion
4. Materials and Methods
4.1. Animal Management
4.2. Experimental Design and Brain Tissue Collection
4.3. Drug Preparation
4.4. Relative Abundance of mRNA Analysis
4.5. Enzyme Repair Activity
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| εA | 1,N6-ethenoadenine |
| εC | 3,N4-ethenocytosine |
| 8-oxoG | 8-oxoguanine |
| IIIv | Third brain ventricle |
| ACTH | Adrenocorticotropic hormone |
| AL | Allopregnanolone |
| AMG | Amygdaloid nucleus |
| ANOVA | Analysis of variance |
| AP | Apurinic/apyrimidinic site |
| APE1 | AP-endonuclease 1 |
| BDNF | Brain-derived neurotrophic factor |
| BER | Base excision repair |
| CA | Cornu ammonis |
| CNS | Central nervous system |
| CRH | Corticotropin-releasing hormone |
| CSF | Cerebrospinal fluid |
| DMSO | Dimethyl sulfoxide |
| GABA | γ-aminobutyric acid |
| HPA | Hypothalamic–pituitary–adrenal axis |
| icv | Intracerebroventricular |
| MPG | N-methylpurine DNA glycosylase |
| NER | Nucleotide excision repair |
| TDG | Thymine DNA glycosylase |
| OGG1 | 8-oxoguanine glycosylase |
| PVN | Paraventricular nucleus |
| RL | Ringer-Locke solution |
| ROS | Reactive oxygen species |
| SEM | Standard error of the mean |
| SOD | Superoxide dismutase |
References
- Kehrer, J.P. Free radicals as mediators of tissue injury and disease. Critic. Rev. Toxicol. 1993, 23, 21–48. [Google Scholar] [CrossRef] [PubMed]
- Mc Cord, J.M. The evolution of free radicals and oxidative stress. Am. J. Med. 2000, 108, 652–659. [Google Scholar] [CrossRef]
- Marnett, L.J.; Plastaras, J.P. Endogenous DNA damage and mutation. Trends Genet. 2001, 17, 214–221. [Google Scholar] [CrossRef]
- Bartsch, H.; Barbin, A.; Marion, M.J.; Nair, J.; Guichard, Y. Formation, detection, and role in carcinogenesis of ethenobases in DNA. Drug Metab. Rev. 1994, 26, 349–371. [Google Scholar] [CrossRef] [PubMed]
- El Ghissassi, F.; Barbin, A.; Nair, J.; Bartsch, H. Formation of 1,N6-ethenoadenine and 3,N4-ethenocytosine by lipid peroxidation products and nucleic acid bases. Chem. Res. Toxicol. 1995, 8, 278–283. [Google Scholar] [CrossRef]
- Winczura, A.; Zdzalik, D.; Tudek, B. Damage of DNA and proteins by major lipid peroxidation products in genome stability. Free Radic. Res. 2012, 46, 442–459. [Google Scholar] [CrossRef]
- Olinski, R.; Gackowski, D.; Rozalski, R.; Foksinski, M.; Bialkowski, K. Oxidative DNA damage in cancer patients: A cause or a consequence of the disease development? Mutat. Res. 2003, 531, 177–190. [Google Scholar] [CrossRef]
- Bartsch, H.; Nair, J. Oxidative stress and lipid peroxidation derived DNA-lesions in inflammation driven carcinogenesis. Cancer Detect. Prev. 2004, 28, 385–391. [Google Scholar] [CrossRef]
- Krokan, H.E.; Bjørås, M. Base excision repair. Cold Spring Harb. Perspect. Biol. 2013, 5, a012583. [Google Scholar] [CrossRef]
- Krokan, H.E.; Nilsen, H.; Skorpen, F.; Otterlei, M.; Slupphaug, G. Base excision repair of DNA in mammalian cells. Febs Lett. 2000, 476, 73–77. [Google Scholar] [CrossRef]
- Memisoglu, A.; Samson, L. Base-excision repair in yeast and mammals. Mutat. Res. 2000, 451, 39–51. [Google Scholar] [CrossRef]
- Kasai, H. Analysis of a form of oxidative DNA damage, 8-hydroxy-2’deoxyguanosine, as a marker of cellular oxidative stress during carcinogenesis. Mutat. Res. 1997, 387, 147–163. [Google Scholar] [CrossRef]
- Salim, S. Oxidative stress and the central nervous system. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Purdy, R.H.; Morrow, A.L.; Moore, P.H.J.; Paul, S.M. Stress-induced elevations of gamma-aminobutyric acid type A receptor-active steroids in the rat brain. Proc. Natl. Acad. Sci. USA 1991, 88, 4553–4557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valle´e, M.; Rivera, J.D.; Koob, G.F.; Purdy, R.H.; Fitzgerald, R.L. Quantification of neurosteroids in rat plasma and brain following swim stress and allopregnanolone administration using negative chemical ionization gas chromatography/mass spectrometry. Anal. Biochem. 2000, 287, 153–166. [Google Scholar] [CrossRef]
- Brunton, P.J. Neuroactive steroids and stress axis regulation: Pregnancy and beyond. J. Steroid Biochem. Mol. Biol. 2016, 160, 160–168. [Google Scholar] [CrossRef]
- Gunn, B.G.; Cunningham, L.; Mitchell, S.G.; Swinny, J.D.; Lambert, J.J.; Belelli, D. GABAA receptor–acting neurosteroids: A role in the development and regulation of the stress response. Front. Neuroendocrinol. 2015, 36, 28–48. [Google Scholar] [CrossRef]
- Pibiri, F.; Nelson, M.; Guidotti, A.; Costa, E.; Pinna, G. Decreased corticolimbic allopregnanolone expression during social isolation enhances contextual fear: A model relevant for posttraumatic stress disorder. Proc. Natl. Acad. Sci. USA 2008, 105, 5567–5572. [Google Scholar] [CrossRef] [Green Version]
- Biggio, G.; Pisu, M.G.; Biggio, F.; Serra, M. Allopregnanolone modulation of HPA axis function in the adult rat. Psychopharmacology 2014, 231, 3437–3444. [Google Scholar] [CrossRef]
- Locci, A.; Pinna, G. Neurosteroid biosynthesis down-regulation and changes in GABAA receptor subunit composition: A biomarker axis in stress-induced cognitive and emotional impairment. Br. J. Pharmacol. 2017, 174, 3226–3241. [Google Scholar] [CrossRef] [Green Version]
- Pinna, G.; Rasmusson, A.M. Up-regulation of neurosteroid biosynthesis as a pharmacological strategy to improve behavioural deficits in a putative mouse model of post-traumatic stress disorder. J. Neuroendocrinol. 2011, 24, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Boero, G.; Porcu, P.; Morrow, A.L. Pleiotropic actions of allopregnanolone underlie therapeutic benefits in stress-related disease. Neurobiol. Stress 2020, 12, 100203. [Google Scholar] [CrossRef]
- Herman, J.P.; Cullinan, W.E. Neurocircuitry of stress: Central control of the hypothalamo-pituitary adrenocortical axis. Trends Neurosci. 1997, 20, 78–84. [Google Scholar] [CrossRef]
- Sigurdsson, T.; Doyere, V.; Cain, C.K.; LeDoux, J.E. Long-term potentiation in the amygdala: A cellular mechanism of fear learning and memory. Neuropharmacology 2007, 52, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Misztal, T.; Młotkowska, P.; Marciniak, E.; Misztal, A. Allopregnanolone reduces neuroendocrine response to acute stressful stimuli in sheep. J. Endocrinol. 2020, 244, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Sousa, N.; Lukoyanov, N.V.; Madeira, M.D.; Almeida, O.F.; Paula-Barbosa, M.M. Reorganization of the morphology of hippocampal neurites and synapses after stress-induced damage correlates with behavioral improvement. Neuroscience 2000, 97, 253–266. [Google Scholar] [CrossRef]
- Wang, X.; Michaelis, E.K. Selective neuronal vulnerability to oxidative stress in the brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef]
- Wilde, G.J.; Pringle, A.K.; Wright, P.; Iannotti, F. Differential vulnerability of the CA1 and CA3 subfields of the hippocampus to superoxide and hydroxyl radicals in vitro. J. Neurochem. 1997, 69, 883–886. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Pal, R.; Chen, X.W.; Limpeanchob, N.; Kumar, K.N.; Michaelis, E.K. High intrinsic oxidative stress may underlie selective vulnerability of the hippocampal CA1 region. Brain Res. 2005, 140, 120–126. [Google Scholar] [CrossRef]
- Moghaddam, B. Stress activation of glutamate neurotransmission in the prefrontal cortex: Implications for dopamine-associated psychiatric disorders. Biol. Psych. 2002, 51, 775–787. [Google Scholar] [CrossRef]
- Carrasco, G.A.; Van de Kar, L.D. Neuroendocrine pharmacology of stress. Eur. J. Pharmacol. 2003, 463, 235–272. [Google Scholar] [CrossRef]
- Gray, T.S.; Carney, M.E.; Magnuson, D.J. Direct projections from the central amygdaloid nucleus to the hypothalamic paraventricular nucleus: Possible role in stress-induced adrenocorticotropin release. Neuroendocrinology 1989, 50, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Sawchenko, P.E.; Swanson, L.W. The organization of forebrain afferents to the paraventricular and supraoptic nuclei of the rat. J. Comp. Neurol. 1983, 218, 121–144. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Umegaki, H.; Suzuki, Y.; Miura, H.; Iguchi, A. Involvement of the bed nucleus of the stria terminalis in hippocampal cholinergic system-mediated activation of the hypothalamo—pituitary—adrenocortical axis in rats. Brain Res. 2001, 916, 101–106. [Google Scholar] [CrossRef]
- Hill, J.W.; Hazra, T.K.; Izumi, T.; Mitra, S. Stimulation of human 8-oxoguanine-DNA glycosylase by AP-endonuclease: Potential coordination of the initial steps in base excision repair. Nucleic Acids Res. 2001, 29, 430–438. [Google Scholar] [CrossRef]
- Reardon, J.T.; Bessho, T.; Kung, H.C.; Bolton, P.H.; Sancar, A. In vitro repair of oxidative DNA damage by human nucleotide excision repair system: Possible explanation for neurodegeneration in xeroderma pigmentosum patients. Proc. Natl. Acad. Sci. USA 1997, 94, 9463–9468. [Google Scholar] [CrossRef] [Green Version]
- Swanson, R.L.; Morey, N.J.; Doetsch, P.W.; Jinks-Robertson, S. Overlapping specificities of base excision repair, nucleotide excision repair, recombination, and trans lesion synthesis path-ways for DNA base damage in Saccharomyces cerevisiae. Mol. Cell. Biol. 1999, 19, 2929–2935. [Google Scholar] [CrossRef] [Green Version]
- Maki, H.; Sekiguchi, M. MutT protein specifically hydrolyses a potent mutagenic substrate for DNA synthesis. Nature 1992, 355, 273–275. [Google Scholar] [CrossRef]
- Gredilla, R. DNA damage and base excision repair in mitochondria and their role in aging. J. Aging Res. 2011, 2011, 257093. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, P.; Jaworek, J.; Kot, M.; Sokołowska, B.; Bieleń, A.; Janowska, B.; Cieśla, J.M.; Szparecki, G.; Sadoś, B.; Tudek, B. Inflammation increases oxidative DNA damage repair and stimulates preneoplastic changes in colons of newborn rats. J. Physiol. Pharmacol. 2016, 67, 277–286. [Google Scholar]
- Olinski, R.; Zastawny, T.; Budzbon, J.; Skokowski, J.; Zegarski, W.; Dizdaroglu, M. DNA base modifications in chromatin of human cancerous tissues. FEBS Lett. 1992, 309, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and oxidative stress in human diseases: From molecular mechanisms to novel treatments. Int. J. Mol. Sci. 2019, 20, 4472. [Google Scholar] [CrossRef] [Green Version]
- Shigenaga, M.K.; Gimeno, C.J.; Ames, B.N. Urinary 8-hydroxy-2-deoxyguanosine as a biological marker of in vivo oxidative DNA damage. Proc. Nat. Acad. Sci. USA 1989, 86, 9697–9701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B. Can oxidative DNA damage be used as a biomarker of cancer risk in humans? Problems, resolutions, and preliminary results from nutritional supplementation studies. Free Radic. Res. 1998, 29, 469–486. [Google Scholar] [CrossRef] [PubMed]
- Marnett, L.J. Oxyradicals and DNA damage. Carcinogenesis 2000, 21, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraga, C.G.; Shigenaga, M.K.; Park, J.W.; Degan, P.; Ames, B.N. Oxidative damage to DNA during aging: 8-hydroxy-2-deoxyguanosine in rat organ DNA and urine. Proc. Nat. Acad. Sci. USA 1990, 87, 4533–4537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayre, L.M.; Smith, M.A.; Perry, G. Chemistry and biochemistry of oxidative stress in neurodegenerative disease. Cur. Med. Chem 2001, 8, 721–738. [Google Scholar] [CrossRef]
- Kasai, H.; Iwamoto-Tanaka, N.; Miyamoto, T.; Kawanami, K.; Kawanami, S.; Kido, R.; Ikeda, M. Life style and urinary 8-hydroxydeoxygua-nosine, a marker of oxidative DNA damage: Effects of exercise, working conditions, meat intake, body mass index, and smoking. Japan. J. Cancer Res. 2001, 92, 9–15. [Google Scholar] [CrossRef]
- Halliwell, B. Effect of diet on cancer development: Is oxidative DNA damage a biomarker? Free Rad. Biol. Med. 2002, 32, 968–974. [Google Scholar]
- Chaouloff, F. Physiopharmacological interactions between stress hormones and central serotonergic systems. Brain Res. Rev. 1993, 18, 1–32. [Google Scholar] [CrossRef]
- Landfield, P.W. Increased hippocampal Ca2+ channel activity in brain aging and dementia. Hormonal and pharmacologic modulation. Ann. N. Y. Acad. Sci. 1994, 747, 35–364. [Google Scholar]
- Magarinos, A.M.; Deslandes, A.; McEwen, B.S. Effects of antidepressants and benzodiazepine treatments on the dendritic structure of CA3 pyramidal neurons after chronic stress. Eur. J. Pharmacol. 1999, 371, 113–122. [Google Scholar] [CrossRef]
- Ray, S.D.; Lam, L.S.; Rotollo, J.A.; Phadke, S.; Patel, C.; Dontabhaktuni, A.; Mohammad, S.; Lee, H.; Strika, S.; Dobrogowska, A.; et al. Oxidative stress is the master operator of drug and chemically-induced programmed and unprogrammed cell death: Implications of natural antioxidants in vivo. BioFactors 2004, 21, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; Patel, P.D.; Akil, H.; Watson, S.J. Localization and regulation of glucocorticoid and mineralocorticoid receptor messenger RNAs in the hippocampal formation of the rat. Mol. Endocrinol. 1989, 3, 1886–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.J.; Song, E.Y.; Kosten, T.A. Stress effects in the hippocampus: Synaptic plasticity and memory. Stress 2006, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Joëls, M.; de Kloet, E.R. 30 years of the mineralocorticoid receptor: The brain mineralocorticoid receptor: A saga in three episodes. J. Endocrinol. 2017, 234, T49–T66. [Google Scholar] [CrossRef]
- Watanabe, Y.; Gould, E.; McEwen, B.S. Stress induces atrophy of apical dendrites of hippocampal CA3 pyramidal neurons. Brain Res. 1992, 588, 341–345. [Google Scholar] [CrossRef]
- Gould, E.; Tanapat, P.; McEwen, B.S.; Flugge, G.; Fuchs, E. Proliferation of granule cell precursors in the dentate gyrus of adult monkeys is diminished by stress. Proc. Natl. Acad. Sci. USA 1998, 95, 3168–3171. [Google Scholar] [CrossRef] [Green Version]
- Sapolsky, R.M. Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Arch. Gen. Psychiatry 2000, 57, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.M.; Purdy, R.H. Neuroactive steroids. Faseb J. 1992, 6, 2311–2322. [Google Scholar] [CrossRef]
- Agis-Balboa, R.C.; Pinna, G.; Zhubi, A.; Maloku, E.; Veldic, M.; Costa, E.; Guidotti, A. Characterization of brain neurons that express enzymes mediating neurosteroid biosynthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 14602–14607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbaccia, M.L.; Roscetti, G.; Trabucchi, M.; Cuccheddu, T.; Concas, A.; Biggio, G. Neurosteroids in the brain of handling-habituated and naive rats: Effect of CO2 inhalation. Europ. J. Pharmacol. 1994, 261, 317–320. [Google Scholar] [CrossRef]
- Billiards, S.S.; Walker, D.W.; Canny, B.J.; Hirst, J.J. Endotoxin increases sleep and brain allopregnanolone concentrations in newborn lambs. Pediat. Res. 2002, 52, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Sze, Y.; Brunton, P.J. Sex, stress and steroids. Eur. J. Neurosci. 2020, 52, 2487–2515. [Google Scholar] [CrossRef]
- Concas, A.; Mostallino, M.C.; Porcu, P.; Follesa, P.; Barbaccia, M.L.; Trabucchi, M.; Purdy, R.H.; Grisenti, P.; Biggio, G. Role of brain allopregnanolone in the plasticity of γ–aminobutyric acid type A receptor in rat brain during pregnancy and after delivery. Proc. Natl. Acad. Sci. USA 1998, 95, 13284–13289. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, J.; Wakefield, S.; MacKenzie, G.; Moss, S.J.; Maguire, J. Neurosteroidogenesis is required for the physiological response to stress: Role of neurosteroid-sensitive GABAA receptors. J. Neurosci. 2011, 31, 18198–18210. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.L.; Lin, Y.T.; Chuang, P.C.; Bohr, V.A.; Mattson, M.P. BDNF and exercise enhance neuronal DNA repair by stimulating CREB-mediated production of apurinic/apyrimidinic endonuclease 1. Neuromolecular Med. 2014, 16, 161–174. [Google Scholar] [CrossRef]
- Almeida, F.B.; Nin, M.S.; Barros, H.M.T. The role of allopregnanolone in depressive-like behaviors: Focus on neurotrophic proteins. Neurobiol. Stress 2020, 12, 100218. [Google Scholar] [CrossRef]
- Cho, I.; Kim, W.-J.; Kim, H.-W.; Heo, K.; Lee, B.I.; Cho, Y.-J. Increased superoxide dismutase 2 by allopregnanolone ameliorates ROS-mediated neuronal death in mice with pilocarpine-induced status epilepticus. Neurochem. Res. 2018, 43, 1464–1475. [Google Scholar] [CrossRef]
- Zampieri, S.; Mellon, S.H.; Butters, T.D.; Nevyjel, M.; Covey, D.F.; Bembi, B.; Dardis, A. Oxidative stress in NPC1 deficient cells: Protective effect of allopregnanolone. J. Cell. Mol. Med. 2009, 13, 3786–3796. [Google Scholar] [CrossRef]
- Qian, X.; Cao, H.; Ma, Q.; Wang, Q.; He, W.; Qin, P.; Ji, B.; Yuan, K.; Yang, F.; Liu, X.; et al. Allopregnanolone attenuates Abeta25-35-induced neurotoxicity in PC12 cells by reducing oxidative stress. Int. J. Clin. Exp. Med. 2015, 8, 13610–13615. [Google Scholar] [PubMed]
- Strzetelski, J. IZ PIB–INRA Nutrient requirements for ruminants. IZ PIB Krakow 2009. (In Polish) [Google Scholar]
- Welento, J.; Szteyn, S.; Milart, Z. Observations on the stereotaxic configuration of the hypothalamus nuclei in the sheep. Anat. Anz. 1969, 124, 1–27. [Google Scholar] [PubMed]
- Traczyk, W.; Przekop, F. Methods of investigation of the function of the hypothalamus and hypophysis in chronic experiments in sheep. Acta Physiol. Polon. 1963, 14, 217–226. [Google Scholar]
- Johnson, J.I.; Sudheimer, K.D.; Davis, K.K.; Kerndt, G.M.; Winn, B.M. The Sheep Brain Atlas. Michigan State University, Brain Biodiversity Bank. Available online: http://brains.anatomy.msu.edu/brains/sheep/index.html (accessed on 30 August 2020).
- Lanteaume, L.; Khalfa, S.; Régis, J.; Marquis, P.; Chauvel, P.; Bartolomei, F. Emotion induction after direct intracerebral stimulations of human amygdala. Cer. Cortex. 2007, 17, 1307–1313. [Google Scholar] [CrossRef]
- Van de Kar, L.D.; Blair, M.L. Forebrain pathways mediating stress-induced hormone secretion. Front. Neuroendocrinol. 1999, 20, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)). Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers (5′–3′) | GenBank Accession Number | Amplicon Size |
|---|---|---|---|
| MPG | F: GCTGAGGGCCAGCCAACACCTGC R: CGCCCCTTTACCCACGGAGCCCA | NC_040275.1/XM027962018.1 | 140 |
| TDG | F: ACACAGGATGCTGTGGGGCT R: TCCCTCGGCCTAGAATTTTC | NC_040254.1 | 120 |
| OGG1 | F: CAGTCATAATAACAGTA R: AACCTCCTCTAAGCACTCAT | NC_040270.1/XM004018285 | 140 |
| APE1 | F: TTAGACATTTGGTTGCC R: GGCACCAACAGGGCTAGCA | NC_040272.1 | 140 |
| 18S rRNA | F: GCAATTATTCCCCCATGAACG R: GGGACTTAATCAACGCAAGC | NR_003286 | 115 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Misztal, T.; Kowalczyk, P.; Młotkowska, P.; Marciniak, E. The Effect of Allopregnanolone on Enzymatic Activity of the DNA Base Excision Repair Pathway in the Sheep Hippocampus and Amygdala under Natural and Stressful Conditions. Int. J. Mol. Sci. 2020, 21, 7762. https://doi.org/10.3390/ijms21207762
Misztal T, Kowalczyk P, Młotkowska P, Marciniak E. The Effect of Allopregnanolone on Enzymatic Activity of the DNA Base Excision Repair Pathway in the Sheep Hippocampus and Amygdala under Natural and Stressful Conditions. International Journal of Molecular Sciences. 2020; 21(20):7762. https://doi.org/10.3390/ijms21207762
Chicago/Turabian StyleMisztal, Tomasz, Paweł Kowalczyk, Patrycja Młotkowska, and Elżbieta Marciniak. 2020. "The Effect of Allopregnanolone on Enzymatic Activity of the DNA Base Excision Repair Pathway in the Sheep Hippocampus and Amygdala under Natural and Stressful Conditions" International Journal of Molecular Sciences 21, no. 20: 7762. https://doi.org/10.3390/ijms21207762
APA StyleMisztal, T., Kowalczyk, P., Młotkowska, P., & Marciniak, E. (2020). The Effect of Allopregnanolone on Enzymatic Activity of the DNA Base Excision Repair Pathway in the Sheep Hippocampus and Amygdala under Natural and Stressful Conditions. International Journal of Molecular Sciences, 21(20), 7762. https://doi.org/10.3390/ijms21207762