rad21 Is Involved in Corneal Stroma Development by Regulating Neural Crest Migration

 , ,
, , 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Expression of Cell Adhesion Pathway Genes Is Altered in LCLs Isolated from Family Members Affected by Peripheral Sclerocornea
2.2. The Genes Involved in Chromosome Organization of Cell Adhesion Pathways Are Altered in LCLs Derived from Family Members Affected by Peripheral Sclerocornea
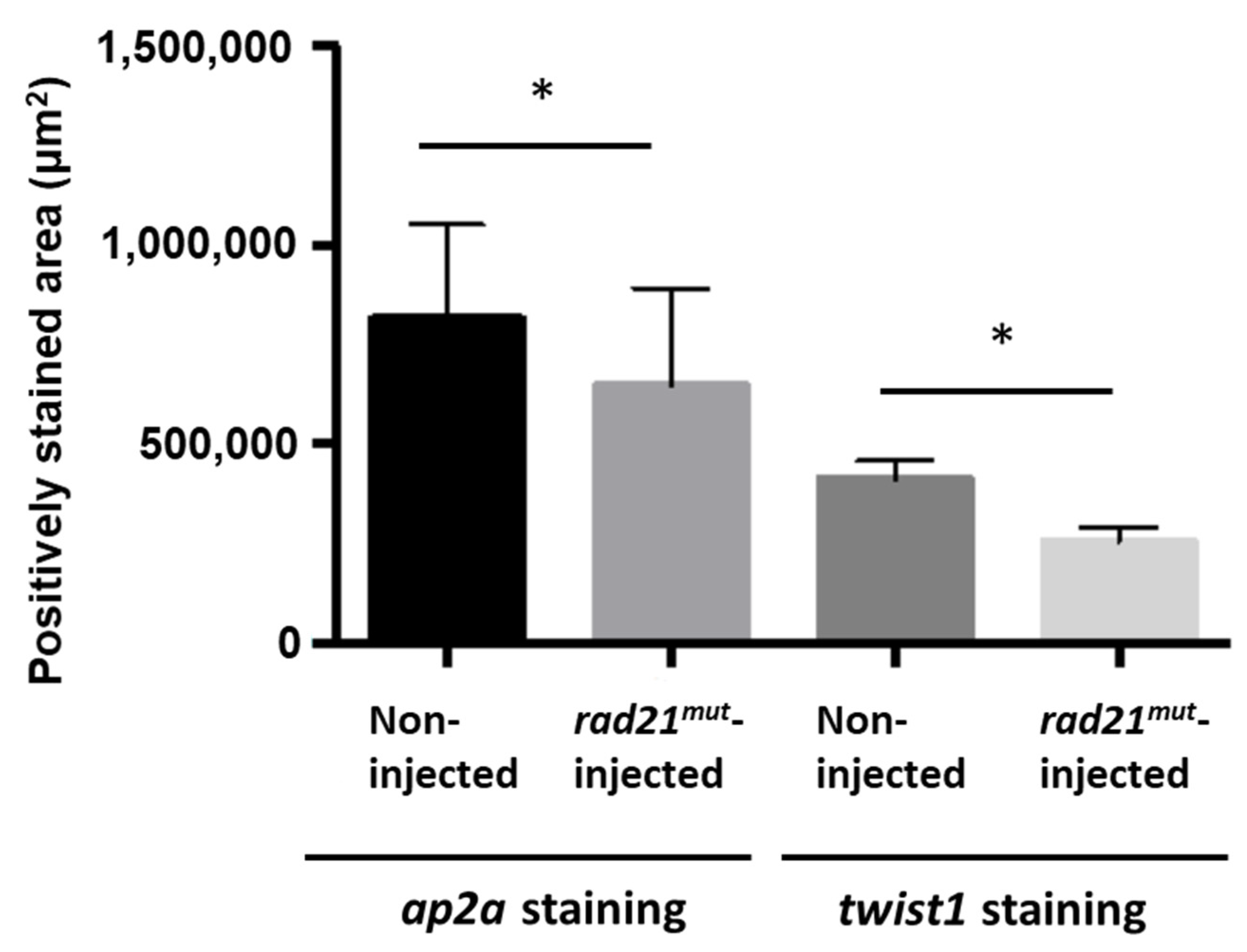
2.3. Disrupted Neural Crest Migration in rad21mut-Injected X. laevis Embryos
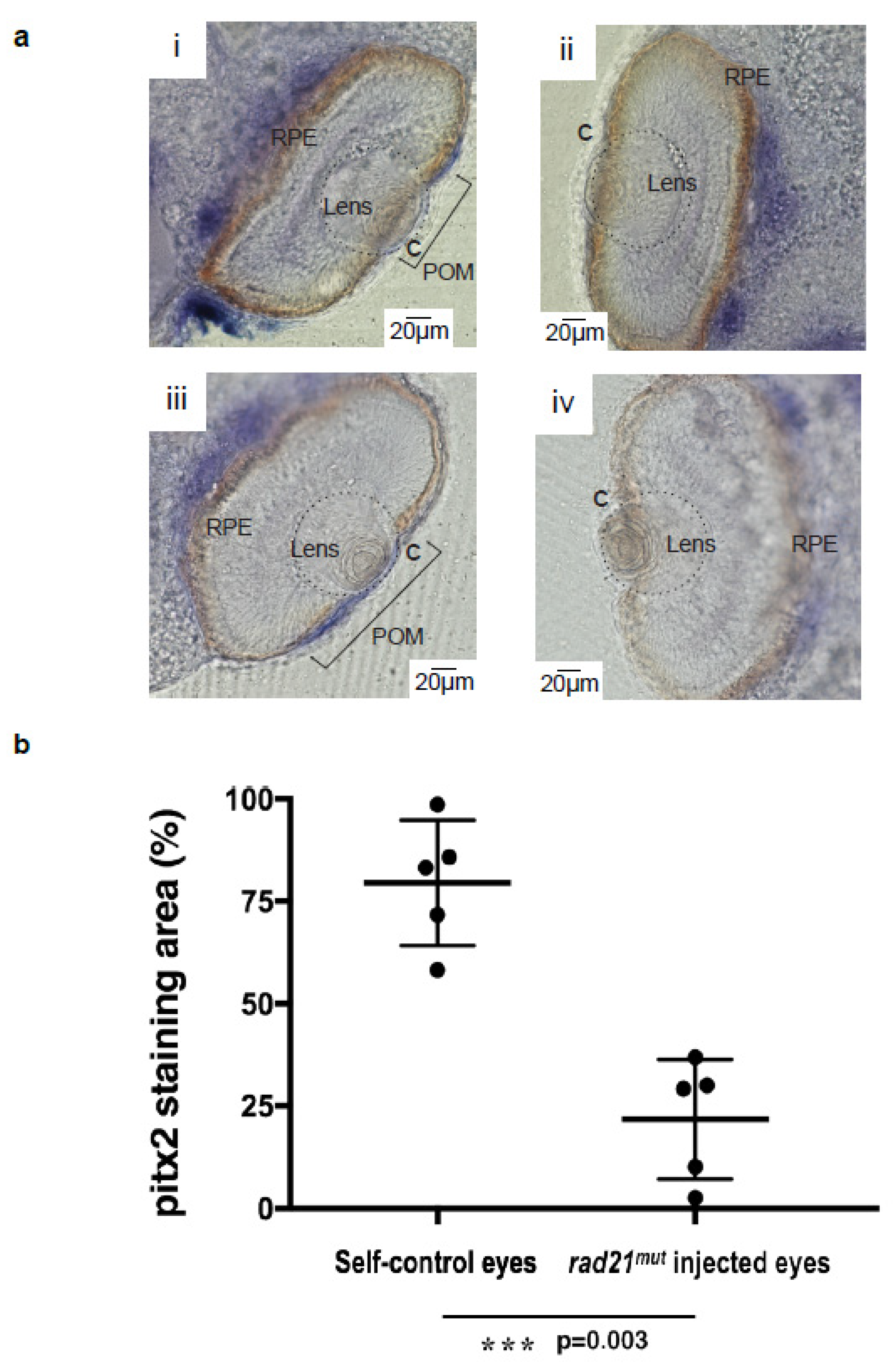
2.4. Decreased Periocular Mesenchymes Invading into the Cornea in rad21mut-Injected X. laevis Embryos
3. Discussion
4. Materials and Methods
4.1. Ethics
4.2. Lymphoblastoid Cell Lines
4.3. RNA Sequencing
4.4. Quantitative Real-Time PCR (qPCR)
4.5. Hi-C
4.6. Wild-Type and Mutant X. laevis Embryos
4.7. Whole-Mount In Situ Hybridization (WISH)
4.8. Vibratome Section
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, B.N.; Chan, T.C.Y.; Tam, P.O.S.; Liu, Y.; Pang, C.P.; Jhanji, V.; Chen, L.J.; Chu, W.K. A Cohesin Subunit Variant Identified from a Peripheral Sclerocornea Pedigree. Dis. Markers 2019, 2019, 8781524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.N.; Wong, T.C.B.; Yip, Y.W.Y.; Liu, Z.; Wang, C.; Wong, J.S.C.; He, J.N.; Chan, T.C.Y.; Jhanji, V.; Pang, C.P.; et al. A sclerocornea-associated RAD21 variant induces corneal stroma disorganization. Exp. Eye Res. 2019, 185, 107687. [Google Scholar] [CrossRef] [PubMed]
- Kanai, A.; Wood, T.C.; Polack, F.M.; Kaufman, H.E. The fine structure of sclerocornea. Investig. Ophthalmol. 1971, 10, 687–694. [Google Scholar]
- Meek, K.M. Corneal collagen-its role in maintaining corneal shape and transparency. Biophys. Rev. 2009, 1, 83–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.N.; Chan, T.C.Y.; Tam, P.O.S.; Liu, Y.; Pang, C.P.; Jhanji, V.; Chen, L.J.; Chu, W.K. Clinical investigation and genetic analysis of a peripheral sclerocornea pedigree. bioRxiv 2019. [Google Scholar] [CrossRef]
- Fan, B.J.; Chen, X.; Sondhi, N.; Sharmila, P.F.; Soumittra, N.; Sripriya, S.; Sacikala, S.; Asokan, R.; Friedman, D.S.; Pasquale, L.R.; et al. Family-Based Genome-Wide Association Study of South Indian Pedigrees Supports WNT7B as a Central Corneal Thickness Locus. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2495–2502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lwigale, P.Y. Corneal Development: Different Cells from a Common Progenitor. Prog. Mol. Biol. Transl. Sci. 2015, 134, 43–59. [Google Scholar] [CrossRef]
- Hay, E.D.; Revel, J.P. Fine structure of the developing avian cornea. Monogr. Dev. Biol. 1969, 1, 1–144. [Google Scholar] [PubMed]
- Sadaghiani, B.; Thiebaud, C.H. Neural crest development in the Xenopus laevis embryo, studied by interspecific transplantation and scanning electron microscopy. Dev. Biol. 1987, 124, 91–110. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Hu, W.; Haamedi, N.; Lee, J.; Kinoshita, T.; Ohnuma, S. The structure and development of Xenopus laevis cornea. Exp. Eye Res. 2013, 116, 109–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West-Mays, J.A.; Dwivedi, D.J. The keratocyte: Corneal stromal cell with variable repair phenotypes. Int. J. Biochem. Cell Biol. 2006, 38, 1625–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasmyth, K. Segregating sister genomes: The molecular biology of chromosome separation. Science 2002, 297, 559–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkenbihl, R.P.; Subramani, S. Cloning and characterization of rad21 an essential gene of Schizosaccharomyces pombe involved in DNA double-strand-break repair. Nucleic Acids Res. 1992, 20, 6605–6611. [Google Scholar] [CrossRef] [Green Version]
- Nagao, K.; Adachi, Y.; Yanagida, M. Separase-mediated cleavage of cohesin at interphase is required for DNA repair. Nature 2004, 430, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Watrin, E.; Peters, J.M. The cohesin complex is required for the DNA damage-induced G2/M checkpoint in mammalian cells. EMBO J. 2009, 28, 2625–2635. [Google Scholar] [CrossRef]
- Kagey, M.H.; Newman, J.J.; Bilodeau, S.; Zhan, Y.; Orlando, D.A.; van Berkum, N.L.; Ebmeier, C.C.; Goossens, J.; Rahl, P.B.; Levine, S.S.; et al. Mediator and cohesin connect gene expression and chromatin architecture. Nature 2010, 467, 430–435. [Google Scholar] [CrossRef] [Green Version]
- Fudenberg, G.; Imakaev, M.; Lu, C.; Goloborodko, A.; Abdennur, N.; Mirny, L.A. Formation of Chromosomal Domains by Loop Extrusion. Cell Rep. 2016, 15, 2038–2049. [Google Scholar] [CrossRef] [Green Version]
- Aquila, L.; Ohm, J.; Woloszynska-Read, A. The role of STAG2 in bladder cancer. Pharmacol. Res. 2018, 131, 143–149. [Google Scholar] [CrossRef]
- Kon, A.; Shih, L.Y.; Minamino, M.; Sanada, M.; Shiraishi, Y.; Nagata, Y.; Yoshida, K.; Okuno, Y.; Bando, M.; Nakato, R.; et al. Recurrent mutations in multiple components of the cohesin complex in myeloid neoplasms. Nat. Genet. 2013, 45, 1232–1237. [Google Scholar] [CrossRef]
- Deardorff, M.A.; Wilde, J.J.; Albrecht, M.; Dickinson, E.; Tennstedt, S.; Braunholz, D.; Monnich, M.; Yan, Y.; Xu, W.; Gil-Rodriguez, M.C.; et al. RAD21 mutations cause a human cohesinopathy. Am. J. Hum. Genet. 2012, 90, 1014–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieberman-Aiden, E.; van Berkum, N.L.; Williams, L.; Imakaev, M.; Ragoczy, T.; Telling, A.; Amit, I.; Lajoie, B.R.; Sabo, P.J.; Dorschner, M.O.; et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 2009, 326, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, J.R.; Selvaraj, S.; Yue, F.; Kim, A.; Li, Y.; Shen, Y.; Hu, M.; Liu, J.S.; Ren, B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 2012, 485, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekker, J.; Heard, E. Structural and functional diversity of Topologically Associating Domains. FEBS Lett. 2015, 589, 2877–2884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pope, J.H.; Horne, M.K.; Scott, W. Transformation of foetal human keukocytes in vitro by filtrates of a human leukaemic cell line containing herpes-like virus. Int. J. Cancer 1968, 3, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.S.P.; Huang, S.C.; Glenn St Hilaire, B.; Engreitz, J.M.; Perez, E.M.; Kieffer-Kwon, K.R.; Sanborn, A.L.; Johnstone, S.E.; Bascom, G.D.; Bochkov, I.D.; et al. Cohesin Loss Eliminates All Loop Domains. Cell 2017, 171, 305–320.e24. [Google Scholar] [CrossRef] [Green Version]
- Schwarzer, W.; Abdennur, N.; Goloborodko, A.; Pekowska, A.; Fudenberg, G.; Loe-Mie, Y.; A Fonseca, N.; Huber, W.; Haering, C.H.; Mirny, L.; et al. Two independent modes of chromatin organization revealed by cohesin removal. Nature 2017, 551, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Crane, E.; Bian, Q.; McCord, R.P.; Lajoie, B.R.; Wheeler, B.S.; Ralston, E.J.; Uzawa, S.; Dekker, J.; Meyer, B.J. Condensin-driven remodelling of X chromosome topology during dosage compensation. Nature 2015, 523, 240–244. [Google Scholar] [CrossRef]
- Mizuguchi, T.; Fudenberg, G.; Mehta, S.; Belton, J.M.; Taneja, N.; Folco, H.D.; FitzGerald, P.; Dekker, J.; Mirny, L.; Barrowman, J.; et al. Cohesin-dependent globules and heterochromatin shape 3D genome architecture in S. pombe. Nature 2014, 516, 432–435. [Google Scholar] [CrossRef] [Green Version]
- Gerstein, M.B.; Kundaje, A.; Hariharan, M.; Landt, S.G.; Yan, K.K.; Cheng, C.; Mu, X.J.; Khurana, E.; Rozowsky, J.; Alexander, R.; et al. Architecture of the human regulatory network derived from ENCODE data. Nature 2012, 489, 91–100. [Google Scholar] [CrossRef]
- Dixon, J.R.; Jung, I.; Selvaraj, S.; Shen, Y.; Antosiewicz-Bourget, J.E.; Lee, A.Y.; Ye, Z.; Kim, A.; Rajagopal, N.; Xie, W.; et al. Chromatin architecture reorganization during stem cell differentiation. Nature 2015, 518, 331–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hnisz, D.; Weintraub, A.S.; Day, D.S.; Valton, A.L.; Bak, R.O.; Li, C.H.; Goldmann, J.; Lajoie, B.R.; Fan, Z.P.; Sigova, A.A.; et al. Activation of proto-oncogenes by disruption of chromosome neighborhoods. Science 2016, 351, 1454–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfandari, D.; Cousin, H.; Marsden, M. Mechanism of Xenopus cranial neural crest cell migration. Cell Adh. Migr. 2010, 4, 553–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Kratzer, M.C.; Wedlich, D.; Kashef, J. E-cadherin is required for cranial neural crest migration in Xenopus laevis. Dev. Biol. 2016, 411, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Cvekl, A.; Tamm, E.R. Anterior eye development and ocular mesenchyme: New insights from mouse models and human diseases. Bioessays 2004, 26, 374–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenyon, K.R. Mesenchymal dysgenesis in Peter’s anomaly, sclerocornea and congenital endothelial dystrophy. Exp. Eye Res. 1975, 21, 125–142. [Google Scholar] [CrossRef]
- Beauchamp, G.R.; Knepper, P.A. Role of the neural crest in anterior segment development and disease. J. Pediatr. Ophthalmol. Strabismus. 1984, 21, 209–214. [Google Scholar]
- Williams, A.L.; Bohnsack, B.L. Neural crest derivatives in ocular development: Discerning the eye of the storm. Birth Defects Res. C Embryo Today 2015, 105, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.F.; Pressman, C.; Dyer, R.; Johnson, R.L.; Martin, J.F. Function of Rieger syndrome gene in left-right asymmetry and craniofacial development. Nature 1999, 401, 276–278. [Google Scholar] [CrossRef]
- Matt, N.; Dupe, V.; Garnier, J.M.; Dennefeld, C.; Chambon, P.; Mark, M.; Ghyselinck, N.B. Retinoic acid-dependent eye morphogenesis is orchestrated by neural crest cells. Development 2005, 132, 4789–4800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasim, A.; Smith, B.P. Genetic control of radiation sensitivity in Schizosaccharomyces pombe. Genetics 1975, 79, 573–582. [Google Scholar] [PubMed]
- Phipps, J.; Nasim, A.; Miller, D.R. Recovery, repair, and mutagenesis in Schizosaccharomyces pombe. Adv. Genet. 1985, 23, 1–72. [Google Scholar] [PubMed]
- Tanaka, T.; Cosma, M.P.; Wirth, K.; Nasmyth, K. Identification of cohesin association sites at centromeres and along chromosome arms. Cell 1999, 98, 847–858. [Google Scholar] [CrossRef] [Green Version]
- Hauf, S.; Waizenegger, I.C.; Peters, J.M. Cohesin cleavage by separase required for anaphase and cytokinesis in human cells. Science 2001, 293, 1320–1323. [Google Scholar] [CrossRef]
- Ying, S.; Minocherhomji, S.; Chan, K.L.; Palmai-Pallag, T.; Chu, W.K.; Wass, T.; Mankouri, H.W.; Liu, Y.; Hickson, I.D. MUS81 promotes common fragile site expression. Nat. Cell Biol. 2013, 15, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Johnston, M.C.; Noden, D.M.; Hazelton, R.D.; Coulombre, J.L.; Coulombre, A.J. Origins of avian ocular and periocular tissues. Exp. Eye Res. 1979, 29, 27–43. [Google Scholar] [CrossRef]
- Carmona-Fontaine, C.; Matthews, H.K.; Kuriyama, S.; Moreno, M.; Dunn, G.A.; Parsons, M.; Stern, C.D.; Mayor, R. Contact inhibition of locomotion in vivo controls neural crest directional migration. Nature 2008, 456, 957–961. [Google Scholar] [CrossRef] [Green Version]
- Theveneau, E.; Steventon, B.; Scarpa, E.; Garcia, S.; Trepat, X.; Streit, A.; Mayor, R. Chase-and-run between adjacent cell populations promotes directional collective migration. Nat. Cell Biol. 2013, 15, 763–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpa, E.; Szabo, A.; Bibonne, A.; Theveneau, E.; Parsons, M.; Mayor, R. Cadherin Switch during EMT in Neural Crest Cells Leads to Contact Inhibition of Locomotion via Repolarization of Forces. Dev. Cell 2015, 34, 421–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astin, J.W.; Batson, J.; Kadir, S.; Charlet, J.; Persad, R.A.; Gillatt, D.; Oxley, J.D.; Nobes, C.D. Competition amongst Eph receptors regulates contact inhibition of locomotion and invasiveness in prostate cancer cells. Nat. Cell Biol. 2010, 12, 1194–1204. [Google Scholar] [CrossRef]
- Roycroft, A.; Mayor, R. Molecular basis of contact inhibition of locomotion. Cell Mol. Life Sci. 2016, 73, 1119–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akaneya, Y.; Sohya, K.; Kitamura, A.; Kimura, F.; Washburn, C.; Zhou, R.; Ninan, I.; Tsumoto, T.; Ziff, E.B. Ephrin-A5 and EphA5 interaction induces synaptogenesis during early hippocampal development. PLoS ONE 2010, 5, e12486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, F.M.; Vearing, C.; Lackmann, M.; Treutlein, H.; Himanen, J.; Chen, K.; Saul, A.; Nikolov, D.; Boyd, A.W. Dissecting the EphA3/Ephrin-A5 interactions using a novel functional mutagenesis screen. J. Biol. Chem. 2004, 279, 9522–9531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, T.; Chung, T.Y.; Chang, J.H.; Sayegh, R.; Casanova, F.H.; Azar, D.T. Comparison of EphA receptor tyrosine kinases and ephrinA ligand expression to EphB-ephrinB in vascularized corneas. Cornea 2007, 26, 569–578. [Google Scholar] [CrossRef]
- Pasquale, E.B. Eph receptors and ephrins in cancer: Bidirectional signalling and beyond. Nat. Rev. Cancer 2010, 10, 165–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquale, E.B. Eph receptor signalling casts a wide net on cell behaviour. Nat. Rev. Mol. Cell Biol. 2005, 6, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Robinson, V.; Smith, A.; Flenniken, A.M.; Wilkinson, D.G. Roles of Eph receptors and ephrins in neural crest pathfinding. Cell Tissue Res. 1997, 290, 265–274. [Google Scholar] [CrossRef]
- Liu, Y.; Noon, A.P.; Aguiar Cabeza, E.; Shen, J.; Kuk, C.; Ilczynski, C.; Ni, R.; Sukhu, B.; Chan, K.; Barbosa-Morais, N.L.; et al. Next-generation RNA sequencing of archival formalin-fixed paraffin-embedded urothelial bladder cancer. Eur. Urol. 2014, 66, 982–986. [Google Scholar] [CrossRef]
- Belaghzal, H.; Dekker, J.; Gibcus, J.H. Hi-C 2.0: An optimized Hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods 2017, 123, 56–65. [Google Scholar] [CrossRef]
- Belton, J.M.; McCord, R.P.; Gibcus, J.H.; Naumova, N.; Zhan, Y.; Dekker, J. Hi-C: A comprehensive technique to capture the conformation of genomes. Methods 2012, 58, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Lajoie, B.R.; Dekker, J.; Kaplan, N. The Hitchhiker’s guide to Hi-C analysis: Practical guidelines. Methods 2015, 72, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, S.S.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S.; et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.N.; Liu, Y.; Yang, Q.; Leung, P.Y.; Wang, C.; Wong, T.C.B.; Tham, C.C.; Chan, S.O.; Pang, C.P.; Chen, L.J.; et al. rad21 Is Involved in Corneal Stroma Development by Regulating Neural Crest Migration. Int. J. Mol. Sci. 2020, 21, 7807. https://doi.org/10.3390/ijms21207807
Zhang BN, Liu Y, Yang Q, Leung PY, Wang C, Wong TCB, Tham CC, Chan SO, Pang CP, Chen LJ, et al. rad21 Is Involved in Corneal Stroma Development by Regulating Neural Crest Migration. International Journal of Molecular Sciences. 2020; 21(20):7807. https://doi.org/10.3390/ijms21207807
Chicago/Turabian StyleZhang, Bi Ning, Yu Liu, Qichen Yang, Pui Ying Leung, Chengdong Wang, Thomas Chi Bun Wong, Clement C. Tham, Sun On Chan, Chi Pui Pang, Li Jia Chen, and et al. 2020. "rad21 Is Involved in Corneal Stroma Development by Regulating Neural Crest Migration" International Journal of Molecular Sciences 21, no. 20: 7807. https://doi.org/10.3390/ijms21207807
APA StyleZhang, B. N., Liu, Y., Yang, Q., Leung, P. Y., Wang, C., Wong, T. C. B., Tham, C. C., Chan, S. O., Pang, C. P., Chen, L. J., Dekker, J., Zhao, H., & Chu, W. K. (2020). rad21 Is Involved in Corneal Stroma Development by Regulating Neural Crest Migration. International Journal of Molecular Sciences, 21(20), 7807. https://doi.org/10.3390/ijms21207807