Eicosanoids in Skin Wound Healing


Abstract
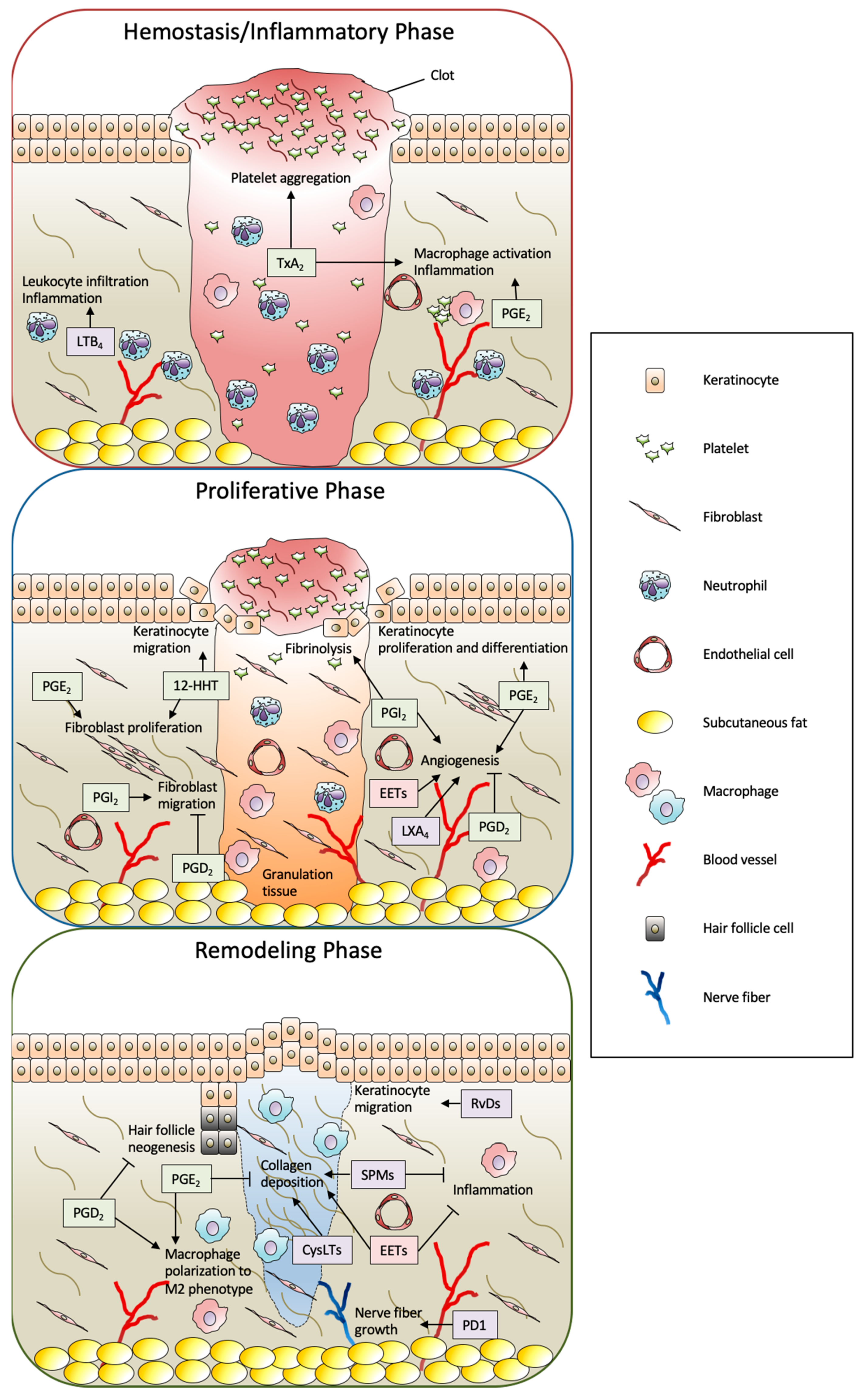
:1. Introduction
2. Biosynthetic Pathway of Eicosanoids and SPMs
3. Functions of Eicosanoids and SPMs in Skin Wound Healing
3.1. COX Metabolites
3.1.1. TxA2
3.1.2. 12(S)-Hydroxyheptadeca-5Z,8E,10E-Trienoic Acid (12-HHT)
3.1.3. PGE2
3.1.4. PGD2
3.1.5. PGF2α
3.1.6. PGI2
3.2. LOX Metabolites
3.2.1. Leukotrienes (LTB4 and CysLTs; LTC4, LTD4, and LTE4)
3.2.2. HETEs
3.3. CYP Metabolites (EETs)
3.4. SPMs (Lipoxins, Resolvins, Protectins, and Maresins)
4. Summary and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PUFA | polyunsaturated fatty acid |
| PLA2 | phospholipase A2 |
| PG | Prostaglandin |
| TxA2 | thromboxane A2 |
| AA | arachidonic acid |
| COX | Cyclooxygenase |
| LT | Leukotriene |
| HETE | hydroxyeicosatetraenoic acid |
| LX | Lipoxin |
| LOX | lipoxygenase |
| EET | epoxyeicosatrienoic acid |
| CYP | cytochrome P450 |
| SPM | specialized pro-resolving mediator |
| EPA | eicosapentaenoic acid |
| DHA | docosahexaenoic acid |
| NSAID | nonsteroidal anti-inflammatory drug |
| HpETE | hydroperoxyeicosatetraenoic acid |
| CysLT | cysteinyl leukotriene |
| LTA4H | LTA4 hydrolase |
| LTC4S | LTC4 synthase |
| MaR1 | maresin 1 |
| TxAS | thromboxane A synthase |
| IL-6, 1β | interleukin-6, 1β |
| 12-HHT | 12(S)-hydroxyheptadeca-5Z,8E,10E-trienoic acid |
| TNFα | tumor necrosis factor α |
| MMP | matrix metalloproteinase |
| TGF-β1 | transforming growth factor-β1 |
| bFGF | basic fibroblast growth factor |
| cPGES | cytosolic prostaglandin E synthase |
| mPGES | membrane-associated prostaglandin E synthase |
| GPCR | G-protein coupled receptor |
| PGT | prostaglandin transporter |
| OSM | oncostatin M |
| CREB | cyclic AMP-responsive element binding |
| KLF4 | Krupple-like factor 4 |
| TIMP | tissue inhibitor of matrix metalloproteinase |
| H-PGDS | hematopoietic-type prostaglandin D synthase |
| L-PGDS | lipocalin-type prostaglandin D synthase |
| PPARγ, δ | peroxisome proliferator-activated receptor γ, δ |
| PGIS | prostacyclin synthase |
| uPA | urokinase-type plasminogen activator |
| VEGF | vascular endothelial growth factor |
| ROS | reactive oxygen species |
| HO-1 | heme oxygenase-1 |
| GM-CSF | granulocyte macrophage colony-stimulating factor |
| sEH | soluble epoxide hydrolase |
| DHET | dihydroxyeicosatrienoic acids |
| SDF-1α | stromal cell-derived factor 1α |
References
- Jean, K. Anatomy, histology and immunohistochemistry of normal human skin. Eur. J. Dermatol. 2002, 12, 390–401. [Google Scholar]
- Ludriksone, L.; Garcia Bartels, N.; Kanti, V.; Blume-Peytavi, U.; Kottner, J. Skin barrier function in infancy: A systematic review. Arch. Dermatol. Res 2014, 306, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Canedo-Dorantes, L.; Canedo-Ayala, M. Skin Acute Wound Healing: A Comprehensive Review. Int. J. Inflamm. 2019, 2019, 3706315. [Google Scholar] [CrossRef] [PubMed]
- Shaw, T.J.; Martin, P. Wound repair at a glance. J. Cell Sci. 2009, 122, 3209–3213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, G.; Ceilley, R. Chronic Wound Healing: A Review of Current Management and Treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falanga, V.; Grinnell, F.; Gilchrest, B.; Maddox, Y.T.; Moshell, A. Workshop on the Pathogenesis of Chronic Wounds. J. Investig. Dermatol. 1994, 102, 125–127. [Google Scholar] [CrossRef] [Green Version]
- Singer, A.J.; Clark, R.A.F. Cutaneous Wound Healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Rosique, R.G.; Rosique, M.J.; Farina Junior, J.A. Curbing Inflammation in Skin Wound Healing: A Review. Int. J. Inflamm. 2015, 2015, 316235. [Google Scholar] [CrossRef] [Green Version]
- Moulin, V. Growth factors in skin wound healing. Eur. J. Cell Biol. 1995, 68, 1–7. [Google Scholar]
- Steed, D.L. The Role of Growth Factors in Wound Healing. Surg. Clin. 1997, 77, 575–586. [Google Scholar] [CrossRef]
- Ridiandries, A.; Tan, J.T.M.; Bursill, C.A. The Role of Chemokines in Wound Healing. Int. J. Mol. Sci. 2018, 19, 3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esser-von Bieren, J. Eicosanoids in tissue repair. Immunol. Cell Biol. 2019, 97, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.-P.; Yin, J.; Yu, F.-S.X. Lysophosphatidic Acid Promoting Corneal Epithelial Wound Healing by Transactivation of Epidermal Growth Factor Receptor. Investig. Ophthalmol. Vis. Sci. 2007, 48, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Aoki, H.; Mukhopadhyay, P.; Tsuge, T.; Yamamoto, H.; Matsumoto, N.M.; Toyohara, E.; Okubo, Y.; Ogawa, R.; Takabe, K. Sphingosine-1-Phosphate Facilitates Skin Wound Healing by Increasing Angiogenesis and Inflammatory Cell Recruitment with Less Scar Formation. Int. J. Mol. Sci. 2019, 20, 3381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.M.; Han, H.J. Autotaxin-LPA axis regulates hMSC migration by adherent junction disruption and cytoskeletal rearrangement via LPAR1/3-dependent PKC/GSK3beta/beta-catenin and PKC/Rho GTPase pathways. Stem Cells 2015, 33, 819–832. [Google Scholar] [CrossRef]
- Aoki, M.; Aoki, H.; Ramanathan, R.; Hait, N.C.; Takabe, K. Sphingosine-1-Phosphate Signaling in Immune Cells and Inflammation: Roles and Therapeutic Potential. Mediat. Inflamm. 2016, 2016, 8606878. [Google Scholar] [CrossRef] [Green Version]
- Dennis, E.A. Phospholipase A2 in eicosanoid generation. Am. J. Respir. Crit. Care Med. 2000, 161, S32–S35. [Google Scholar] [CrossRef]
- Shimizu, T. Lipid mediators in health and disease: Enzymes and receptors as therapeutic targets for the regulation of immunity and inflammation. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 123–150. [Google Scholar] [CrossRef] [Green Version]
- Esser-von Bieren, J. Immune-regulation and -functions of eicosanoid lipid mediators. Biol. Chem. 2017, 398, 1177–1191. [Google Scholar] [CrossRef] [Green Version]
- López, D.E.; Ballaz, S.J. The Role of Brain Cyclooxygenase-2 (Cox-2) Beyond Neuroinflammation: Neuronal Homeostasis in Memory and Anxiety. Mol. Neurobiol. 2020, 57, 5167–5176. [Google Scholar] [CrossRef]
- Ferreira, S.H.; Vane, J.R. Prostaglandins: Their Disappearance from and Release into the Circulation. Nature 1967, 216, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Kirkby, N.S. Eicosanoids, prostacyclin and cyclooxygenase in the cardiovascular system. Br. J. Pharmacol. 2019, 176, 1038–1050. [Google Scholar] [CrossRef] [PubMed]
- Samuelsson, B. Leukotrienes: Mediators of immediate hypersensitivity reactions and inflammation. Science 1983, 220, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.S.; Rokach, J. Biosynthesis, biological effects, and receptors of hydroxyeicosatetraenoic acids (HETEs) and oxoeicosatetraenoic acids (oxo-ETEs) derived from arachidonic acid. Biochim. Biophys. Acta 2015, 1851, 340–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capdevila, J.; Chacos, N.; Werringloer, J.; Prough, R.A.; Estabrook, R.W. Liver microsomal cytochrome P-450 and the oxidative metabolism of arachidonic acid. Proc. Natl. Acad. Sci. USA 1981, 78, 5362–5366. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, K.L.; Endo, T.; Darwesh, A.M.; Samokhvalov, V.; Seubert, J.M. Cytochrome P450-derived eicosanoids and heart function. Pharmacol. Ther. 2017, 179, 47–83. [Google Scholar] [CrossRef]
- Hellmann, J.; Sansbury, B.E.; Wong, B.; Li, X.; Singh, M.; Nuutila, K.; Chiang, N.; Eriksson, E.; Serhan, C.N.; Spite, M. Biosynthesis of D-Series Resolvins in Skin Provides Insights into their Role in Tissue Repair. J. Investig. Dermatol. 2018, 138, 2051–2060. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Levy, B.D. Resolvins in inflammation: Emergence of the pro-resolving superfamily of mediators. J. Clin. Investig. 2018, 128, 2657–2669. [Google Scholar] [CrossRef]
- Dennis, E.A. Diversity of group types, regulation, and function of phospholipase A2. J. Biol. Chem. 1994, 269, 13057–13060. [Google Scholar]
- Funk, C.D. Prostaglandins and leukotrienes: Advances in eicosanoid biology. Science 2001, 294, 1871–1875. [Google Scholar] [CrossRef] [Green Version]
- Hirata, T.; Narumiya, S. Prostanoids as regulators of innate and adaptive immunity. Adv. Immunol. 2012, 116, 143–174. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R.; Bakhle, Y.S.; Botting, R.M. Cyclooxygenases 1 AND 2. Annu. Rev. Pharmacol. Toxicol. 1998, 38, 97–120. [Google Scholar] [CrossRef] [PubMed]
- Penning, T.D.; Talley, J.J.; Bertenshaw, S.R.; Carter, J.S.; Collins, P.W.; Docter, S.; Graneto, M.J.; Lee, L.F.; Malecha, J.W.; Miyashiro, J.M.; et al. Synthesis and Biological Evaluation of the 1,5-Diarylpyrazole Class of Cyclooxygenase-2 Inhibitors: Identification of 4-[5-(4-Methylphenyl)-3-(trifluoromethyl)-1H-pyrazol-1-yl]benzenesulfonamide (SC-58635, Celecoxib). J. Med. Chem. 1997, 40, 1347–1365. [Google Scholar] [CrossRef] [PubMed]
- Futagami, A.; Ishizaki, M.; Fukuda, Y.; Kawana, S.; Yamanaka, N. Wound healing involves induction of cyclooxygenase-2 expression in rat skin. Lab. Investig. 2002, 82, 1503–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, M.; Arai, I.; Futaki, N.; Hashimoto, Y.; Honma, Y.; Nakaike, S. Role of COX-1 and COX-2 on skin PGs biosynthesis by mechanical scratching in mice. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 1–8. [Google Scholar] [CrossRef]
- Buckman, S.Y.; Gresham, A.; Hale, P.; Hruza, G.; Anast, J.; Masferrer, J.; Pentland, A.P. COX-2 expression is induced by UVB exposure in human skin: Implications for the development of skin cancer. Carcinogenesis 1998, 19, 723–729. [Google Scholar] [CrossRef] [Green Version]
- Higashi, Y.; Kanekura, T.; Kanzaki, T. Enhanced expression of cyclooxygenase (COX)-2 in human skin epidermal cancer cells: Evidence for growth suppression by inhibiting COX-2 expression. Int. J. Cancer 2000, 86, 667–671. [Google Scholar] [CrossRef]
- Noverr, M.C.; Erb-Downward, J.R.; Huffnagle, G.B. Production of eicosanoids and other oxylipins by pathogenic eukaryotic microbes. Clin. Microbiol. Rev. 2003, 16, 517–533. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T.; Willoughby, D.A.; Gilroy, D.W. Anti-inflammatory lipid mediators and insights into the resolution of inflammation. Nat. Rev. Immunol. 2002, 2, 787–795. [Google Scholar] [CrossRef]
- Yokomizo, T.; Nakamura, M.; Shimizu, T. Leukotriene receptors as potential therapeutic targets. J. Clin. Investig. 2018, 128, 2691–2701. [Google Scholar] [CrossRef]
- Kuhn, H.; Banthiya, S.; van Leyen, K. Mammalian lipoxygenases and their biological relevance. Biochim. Biophys. Acta 2015, 1851, 308–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rådmark, O.; Werz, O.; Steinhilber, D.; Samuelsson, B. 5-Lipoxygenase, a key enzyme for leukotriene biosynthesis in health and disease. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Sheppard, K.A. Lipoxin formation during human neutrophil-platelet interactions. Evidence for the transformation of leukotriene A4 by platelet 12-lipoxygenase in vitro. J. Clin. Investig. 1990, 85, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, N.; Serhan, C.N.; Dahlen, S.E.; Drazen, J.M.; Hay, D.W.; Rovati, G.E.; Shimizu, T.; Yokomizo, T.; Brink, C. The lipoxin receptor ALX: Potent ligand-specific and stereoselective actions in vivo. Pharmacol. Rev. 2006, 58, 463–487. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.A. Arachidonic acid cytochrome P450 epoxygenase pathway. J. Lipid Res. 2009, 50, S52–S56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svensson, J.; Hamberg, M.; Samuelsson, B. On the formation and effects of thromboxane A2 in human platelets. Acta Physiol. Scand. 1976, 98, 285–294. [Google Scholar] [CrossRef]
- Ogletree, M.L. Overview of physiological and pathophysiological effects of thromboxane A2. Fed. Proc 1987, 46, 133–138. [Google Scholar]
- Thomas, D.W.; Mannon, R.B.; Mannon, P.J.; Latour, A.; Oliver, J.A.; Hoffman, M.; Smithies, O.; Koller, B.H.; Coffman, T.M. Coagulation defects and altered hemodynamic responses in mice lacking receptors for thromboxane A2. J. Clin. Investig. 1998, 102, 1994–2001. [Google Scholar] [CrossRef] [Green Version]
- Lau, C.S.; Khan, F.; McLaren, M.; Bancroft, A.; Walker, M.; Belch, J.J.F. The effects of thromboxane receptor blockade on platelet aggregation and digital skin blood flow in patients with secondary Raynaud’s syndrome. Rheumatol. Int. 1991, 11, 163–168. [Google Scholar] [CrossRef]
- Leslie, M. Beyond Clotting: The Powers of Platelets. Science 2010, 328, 562–564. [Google Scholar] [CrossRef] [PubMed]
- Mekaj, Y.H. The roles of platelets in inflammation, immunity, wound healing and malignancy. Int. J. Clin. Exp. Med. 2016, 9, 5347–5358. [Google Scholar]
- Pierre, S.; Linke, B.; Suo, J.; Tarighi, N.; Del Turco, D.; Thomas, D.; Ferreiros, N.; Stegner, D.; Frölich, S.; Sisignano, M.; et al. GPVI and Thromboxane Receptor on Platelets Promote Proinflammatory Macrophage Phenotypes during Cutaneous Inflammation. J. Investig. Dermatol. 2017, 137, 686–695. [Google Scholar] [CrossRef] [Green Version]
- Daniel, T.O.; Liu, H.; Morrow, J.D.; Crews, B.C.; Marnett, L.J. Thromboxane A2 is a mediator of cyclooxygenase-2-dependent endothelial migration and angiogenesis. Cancer Res. 1999, 59, 4574–4577. [Google Scholar] [PubMed]
- Nie, D.; Lamberti, M.; Zacharek, A.; Li, L.; Szekeres, K.; Tang, K.; Chen, Y.; Honn, K.V. Thromboxane A2 Regulation of Endothelial Cell Migration, Angiogenesis, and Tumor Metastasis. Biochem. Biophys. Res. Commun. 2000, 267, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Okuno, T.; Iizuka, Y.; Okazaki, H.; Yokomizo, T.; Taguchi, R.; Shimizu, T. 12(S)-Hydroxyheptadeca-5Z, 8E, 10E-trienoic acid is a natural ligand for leukotriene B4 receptor 2. J. Exp. Med. 2008, 205, 759–766. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Saeki, K.; Matsunobu, T.; Okuno, T.; Koga, T.; Sugimoto, Y.; Yokoyama, C.; Nakamizo, S.; Kabashima, K.; Narumiya, S.; et al. 12-Hydroxyheptadecatrienoic acid promotes epidermal wound healing by accelerating keratinocyte migration via the BLT2 receptor. J. Exp. Med. 2014, 211, 1063–1078. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Tanaka, R.; Kanazawa, S.; Lu, F.; Hayashi, A.; Yokomizo, T.; Mizuno, H. A synthetic leukotriene B4 receptor type 2 agonist accelerates the cutaneous wound healing process in diabetic rats by indirect stimulation of fibroblasts and direct stimulation of keratinocytes. J. Diabetes Its Complicat. 2017, 31, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Pollack, S.V. Systemic drugs and nutritional aspects of wound healing. Clin. Dermatol. 1984, 2, 68–80. [Google Scholar] [CrossRef]
- Kaushal, M.; Gopalan Kutty, N.; Mallikarjuna Rao, C. Wound healing activity of NOE-aspirin: A pre-clinical study. Nitric Oxide 2007, 16, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Tanioka, T.; Nakatani, Y.; Semmyo, N.; Murakami, M.; Kudo, I. Molecular identification of cytosolic prostaglandin E2 synthase that is functionally coupled with cyclooxygenase-1 in immediate prostaglandin E2 biosynthesis. J. Biol. Chem. 2000, 275, 32775–32782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, M.; Naraba, H.; Tanioka, T.; Semmyo, N.; Nakatani, Y.; Kojima, F.; Ikeda, T.; Fueki, M.; Ueno, A.; Oh, S.; et al. Regulation of prostaglandin E2 biosynthesis by inducible membrane-associated prostaglandin E2 synthase that acts in concert with cyclooxygenase-2. J. Biol. Chem. 2000, 275, 32783–32792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuge, K.; Inazumi, T.; Shimamoto, A.; Sugimoto, Y. Molecular mechanisms underlying prostaglandin E2-exacerbated inflammation and immune diseases. Int. Immunol. 2019, 31, 597–606. [Google Scholar] [CrossRef]
- Sakai, Y.; Fujita, K.; Sakai, H.; Mizuno, K. Prostaglandin E2 regulates the expression of basic fibroblast growth factor messenger RNA in normal human fibroblasts. Kobe J. Med. Sci. 2001, 47, 35–45. [Google Scholar]
- Fairweather, M.; Heit, Y.I.; Buie, J.; Rosenberg, L.M.; Briggs, A.; Orgill, D.P.; Bertagnolli, M.M. Celecoxib inhibits early cutaneous wound healing. J. Surg. Res. 2015, 194, 717–724. [Google Scholar] [CrossRef]
- Kämpfer, H.; Schmidt, R.; Geisslinger, G.; Pfeilschifter, J.; Frank, S. Wound inflammation in diabetic ob/ob mice: Functional coupling of prostaglandin biosynthesis to cyclooxygenase-1 activity in diabetes-impaired wound healing. Diabetes 2005, 54, 1543–1551. [Google Scholar] [CrossRef] [Green Version]
- Syeda, M.M.; Jing, X.; Mirza, R.H.; Yu, H.; Sellers, R.S.; Chi, Y. Prostaglandin transporter modulates wound healing in diabetes by regulating prostaglandin-induced angiogenesis. Am. J. Pathol. 2012, 181, 334–346. [Google Scholar] [CrossRef]
- Ganesh, K.; Das, A.; Dickerson, R.; Khanna, S.; Parinandi, N.L.; Gordillo, G.M.; Sen, C.K.; Roy, S. Prostaglandin E(2) induces oncostatin M expression in human chronic wound macrophages through Axl receptor tyrosine kinase pathway. J. Immunol. 2012, 189, 2563–2573. [Google Scholar] [CrossRef] [Green Version]
- Lowe, N.J.; Stoughton, R.B. Effects of topical prostaglandin E analogue on normal hairless mouse epidermal DNA synthesis. J. Investig. Dermatol. 1977, 68, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Pentland, A.P.; Needleman, P. Modulation of keratinocyte proliferation in vitro by endogenous prostaglandin synthesis. J. Clin. Investig. 1986, 77, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, C.B.; Pillai, S.; Goldyne, M.E. Endogenous prostaglandin E2 modulates calcium-induced differentiation in human skin keratinocytes. Prostaglandins Leukot. Essent. Fat. Acids 1993, 49, 777–781. [Google Scholar] [CrossRef]
- Konger, R.L.; Malaviya, R.; Pentland, A.P. Growth regulation of primary human keratinocytes by prostaglandin E receptor EP2 and EP3 subtypes. Biochim. Biophys. Acta 1998, 1401, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.H.; Huang, B.S.; Horng, H.C.; Yeh, C.C.; Chen, Y.J. Wound healing. J. Chin. Med. Assoc. 2018, 81, 94–101. [Google Scholar] [CrossRef]
- Kotwal, G.J.; Chien, S. Macrophage Differentiation in Normal and Accelerated Wound Healing. Results Probl. Cell Differ. 2017, 62, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Luan, B.; Yoon, Y.-S.; Le Lay, J.; Kaestner, K.H.; Hedrick, S.; Montminy, M. CREB pathway links PGE2 signaling with macrophage polarization. Proc. Natl. Acad. Sci. USA 2015, 112, 15642–15647. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Liu, Y.; Zhang, X.; Zhu, D.; Qi, X.; Cao, X.; Fang, Y.; Che, Y.; Han, Z.C.; He, Z.X.; et al. Prostaglandin E2 hydrogel improves cutaneous wound healing via M2 macrophages polarization. Theranostics 2018, 8, 5348–5361. [Google Scholar] [CrossRef]
- Zhao, J.; Shu, B.; Chen, L.; Tang, J.; Zhang, L.; Xie, J.; Liu, X.; Xu, Y.; Qi, S. Prostaglandin E2 inhibits collagen synthesis in dermal fibroblasts and prevents hypertrophic scar formation in vivo. Exp. Dermatol. 2016, 25, 604–610. [Google Scholar] [CrossRef]
- Helliwell, R.J.; Adams, L.F.; Mitchell, M.D. Prostaglandin synthases: Recent developments and a novel hypothesis. Prostaglandins Leukot. Essent. Fat. Acids 2004, 70, 101–113. [Google Scholar] [CrossRef]
- Joo, M.; Sadikot, R.T. PGD synthase and PGD2 in immune resposne. Mediat. Inflamm. 2012, 2012, 503128. [Google Scholar] [CrossRef] [Green Version]
- Arai, I.; Takano, N.; Hashimoto, Y.; Futaki, N.; Sugimoto, M.; Takahashi, N.; Inoue, T.; Nakaike, S. Prostanoid DP1 receptor agonist inhibits the pruritic activity in NC/Nga mice with atopic dermatitis. Eur. J. Pharmacol. 2004, 505, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Honma, Y.; Arai, I.; Hashimoto, Y.; Futaki, N.; Sugimoto, M.; Tanaka, M.; Nakaike, S. Prostaglandin D2 and prostaglandin E2 accelerate the recovery of cutaneous barrier disruption induced by mechanical scratching in mice. Eur. J. Pharmacol. 2005, 518, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Honma, Y.; Arai, I.; Sakurai, T.; Futaki, N.; Hashimoto, Y.; Sugimoto, M.; Nakanishi, Y.; Nakaike, S. Effects of indomethacin and dexamethasone on mechanical scratching-induced cutaneous barrier disruption in mice. Exp. Dermatol. 2006, 15, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Kliewer, S.A.; Lenhard, J.M.; Willson, T.M.; Patel, I.; Morris, D.C.; Lehmann, J.M. A prostaglandin J2 metabolite binds peroxisome proliferator-activated receptor gamma and promotes adipocyte differentiation. Cell 1995, 83, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Forman, B.M.; Tontonoz, P.; Chen, J.; Brun, R.P.; Spiegelman, B.M.; Evans, R.M. 15-Deoxy-Δ12,14-Prostaglandin J2 is a ligand for the adipocyte determination factor PPARγ. Cell 1995, 83, 803–812. [Google Scholar] [CrossRef] [Green Version]
- Bouhlel, M.A.; Derudas, B.; Rigamonti, E.; Dièvart, R.; Brozek, J.; Haulon, S.; Zawadzki, C.; Jude, B.; Torpier, G.; Marx, N.; et al. PPARgamma activation primes human monocytes into alternative M2 macrophages with anti-inflammatory properties. Cell Metab. 2007, 6, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Shi, R.; Luo, B.; Yang, X.; Qiu, L.; Xiong, J.; Jiang, M.; Liu, Y.; Zhang, Z.; Wu, Y. Macrophage peroxisome proliferator-activated receptor B deficiency delays skin wound healing through impairing apoptotic cell clearance in mice. Cell Death Dis. 2015, 6, e1597. [Google Scholar] [CrossRef] [Green Version]
- Mirza, R.E.; Fang, M.M.; Novak, M.L.; Urao, N.; Sui, A.; Ennis, W.J.; Koh, T.J. Macrophage PPARgamma and impaired wound healing in type 2 diabetes. J. Pathol. 2015, 236, 433–444. [Google Scholar] [CrossRef]
- Kohyama, T.; Liu, X.D.; Wen, F.Q.; Kim, H.J.; Takizawa, H.; Rennard, S.I. Prostaglandin D2 inhibits fibroblast migration. Eur. Respir. J. 2002, 19, 684–689. [Google Scholar] [CrossRef] [Green Version]
- Murata, T.; Lin, M.I.; Aritake, K.; Matsumoto, S.; Narumiya, S.; Ozaki, H.; Urade, Y.; Hori, M.; Sessa, W.C. Role of prostaglandin D2 receptor DP as a suppressor of tumor hyperpermeability and angiogenesis in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 20009–20014. [Google Scholar] [CrossRef] [Green Version]
- Nelson, A.M.; Loy, D.E.; Lawson, J.A.; Katseff, A.S.; Fitzgerald, G.A.; Garza, L.A. Prostaglandin D2 inhibits wound-induced hair follicle neogenesis through the receptor, Gpr44. J. Investig. Dermatol. 2013, 133, 881–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garza, L.A.; Liu, Y.; Yang, Z.; Alagesan, B.; Lawson, J.A.; Norberg, S.M.; Loy, D.E.; Zhao, T.; Blatt, H.B.; Stanton, D.C.; et al. Prostaglandin D2 inhibits hair growth and is elevated in bald scalp of men with androgenetic alopecia. Sci. Transl. Med. 2012, 4, 126ra134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, G.G. Induction of term labor with intravenous PGF2α: A review. Prostaglandins 1973, 4, 765–774. [Google Scholar] [CrossRef]
- Lee, P.Y.; Shao, H.; Xu, L.A.; Qu, C.K. The effect of prostaglandin F2 alpha on intraocular pressure in normotensive human subjects. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1474–1477. [Google Scholar]
- Hernández-Cueto, C.; Vieira, D.N.; Girela, E.; Marques, E.; Calvo, M.D.; Villalobos, M.; Oliveira de Sà, F.; Villanueva, E. Prostaglandin F2a (PGF2a): An inadequate marker of the vitality of wounds? Int. J. Legal Med. 1994, 106, 312–314. [Google Scholar] [CrossRef]
- Muller, K.; Krieg, P.; Marks, F.; Furstenberger, G. Expression of PGF2α receptor mRNA in normal, hyperplastic and neoplastic skin. Carcinogenesis 2000, 21, 1063–1066. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-C.; Tominaga, M.; Yasukawa, K.; Ohba, M.; Takahashi, N.; Honda, K.; Okuno, T.; Takamori, K.; Yokomizo, T. Dietary supplementation of omega-3 fatty acid eicosapentaenoic acid does not ameliorate pruritus in murine models of atopic dermatitis and psoriasis. J. Dermatol. Sci. 2019, 95, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Stjernschantz, J.W. From PGF2α-Isopropyl Ester to Latanoprost: A Review of the Development of Xalatan The Proctor Lecture. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1134–1145. [Google Scholar]
- Vane, J.; Corin, R.E. Prostacyclin: A Vascular Mediator. Eur. J. Vasc. Endovasc. Surgery 2003, 26, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Jackson, W.F.; König, A.; Dambacher, T.; Busse, R. Prostacyclin-induced vasodilation in rabbit heart is mediated by ATP-sensitive potassium channels. Am. J. Physiol. 1993, 264, H238–H243. [Google Scholar] [CrossRef]
- Hatane, T.; Yoshida, E.; Kawano, J.; Sugiki, M.; Onitsuka, T.; Maruyama, M. Prostaglandin I2 analog enhances the expression of urokinase-type plasminogen activator and wound healing in cultured human fibroblast. Biochim. Biophys. Acta 1998, 1403, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Pola, R.; Gaetani, E.; Flex, A.; Aprahamian, T.R.; Bosch-MarcÈ, M.; Losordo, D.; Smith, R.C.; Pola, P. Comparative analysis of the in vivo angiogenic properties of stable prostacyclin analogs: A possible role for peroxisome proliferator-activated receptors. J. Mol. Cell. Cardiol. 2004, 36, 363–370. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Lu, T.; d’Uscio, L.V.; Lam, C.F.; Lee, H.C.; Katusic, Z.S. Angiogenic function of prostacyclin biosynthesis in human endothelial progenitor cells. Circ. Res. 2008, 103, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokomizo, T.; Izumi, T.; Chang, K.; Takuwa, Y.; Shimizu, T. A G-protein-coupled receptor for leukotriene B4 that mediates chemotaxis. Nature 1997, 387, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Brandt, S.L.; Wang, S.; Dejani, N.N.; Klopfenstein, N.; Winfree, S.; Filgueiras, L.; McCarthy, B.P.; Territo, P.R.; Serezani, C.H. Excessive localized leukotriene B4 levels dictate poor skin host defense in diabetic mice. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Yokomizo, T.; Kato, K.; Terawaki, K.; Izumi, T.; Shimizu, T. A second leukotriene B(4) receptor, BLT2. A new therapeutic target in inflammation and immunological disorders. J. Exp. Med. 2000, 192, 421–432. [Google Scholar] [CrossRef]
- Kanaoka, Y.; Boyce, J.A. Cysteinyl leukotrienes and their receptors: Cellular distribution and function in immune and inflammatory responses. J. Immunol. 2004, 173, 1503–1510. [Google Scholar] [CrossRef] [Green Version]
- Maekawa, A.; Kanaoka, Y.; Xing, W.; Austen, K.F. Functional recognition of a distinct receptor preferential for leukotriene E4 in mice lacking the cysteinyl leukotriene 1 and 2 receptors. Proc. Natl. Acad. Sci. USA 2008, 105, 16695–16700. [Google Scholar] [CrossRef] [Green Version]
- Kanaoka, Y.; Maekawa, A.; Austen, K.F. Identification of GPR99 protein as a potential third cysteinyl leukotriene receptor with a preference for leukotriene E4 ligand. J. Biol. Chem. 2013, 288, 10967–10972. [Google Scholar] [CrossRef] [Green Version]
- Brogliato, A.R.; Moor, A.N.; Kesl, S.L.; Guilherme, R.F.; Georgii, J.L.; Peters-Golden, M.; Canetti, C.; Gould, L.J.; Benjamim, C.F. Critical role of 5-lipoxygenase and heme oxygenase-1 in wound healing. J. Invest. Dermatol. 2014, 134, 1436–1445. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, F.R.; Sales-Campos, H.; Nardini, V.; da Costa, T.A.; Fonseca, M.T.C.; Junior, V.R.; Sorgi, C.A.; da Silva, J.S.; Chica, J.E.L.; Faccioli, L.H.; et al. The inhibition of 5-Lipoxygenase (5-LO) products leukotriene B4 (LTB4) and cysteinyl leukotrienes (cysLTs) modulates the inflammatory response and improves cutaneous wound healing. Clin. Immunol. 2018, 190, 74–83. [Google Scholar] [CrossRef]
- Ramalho, T.; Filgueiras, L.; Silva-Jr, I.A.; Pessoa, A.F.M.; Jancar, S. Impaired wound healing in type 1 diabetes is dependent on 5-lipoxygenase products. Sci. Rep. 2018, 8, 14164. [Google Scholar] [CrossRef]
- Oyoshi, M.K.; He, R.; Kanaoka, Y.; ElKhal, A.; Kawamoto, S.; Lewis, C.N.; Austen, K.F.; Geha, R.S. Eosinophil-derived leukotriene C4 signals via type 2 cysteinyl leukotriene receptor to promote skin fibrosis in a mouse model of atopic dermatitis. Proc. Natl. Acad. Sci. USA 2012, 109, 4992–4997. [Google Scholar] [CrossRef] [Green Version]
- Raja, S.K.; Sivamani, K.; Garcia, M.S.; Isseroff, R.R. Wound re-epithelialization: Modulating keratinocyte migration in wound healing. Front. Biosci. 2007, 12, 2849–2868. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.C.; Duhamel, L.; Ford-Hutchison, A. Leukotriene B4 and 12-hydroxyeicosatetraenoic acid stimulate epidermal proliferation in vivo in the guinea pig. J. Investig. Dermatol. 1985, 85, 333–334. [Google Scholar] [CrossRef] [Green Version]
- Ruzicka, T. The role of the epidermal 12-hydroxyeicosatetraenoic acid receptor in the skin. Eicosanoids 1992, 5, S63–S65. [Google Scholar]
- Yokomizo, T.; Kato, K.; Hagiya, H.; Izumi, T.; Shimizu, T. Hydroxyeicosanoids bind to and activate the low affinity leukotriene B4 receptor, BLT2. J. Biol. Chem. 2001, 276, 12454–12459. [Google Scholar] [CrossRef] [Green Version]
- Panigrahy, D.; Kalish, B.T.; Huang, S.; Bielenberg, D.R.; Le, H.D.; Yang, J.; Edin, M.L.; Lee, C.R.; Benny, O.; Mudge, D.K.; et al. Epoxyeicosanoids promote organ and tissue regeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 13528–13533. [Google Scholar] [CrossRef] [Green Version]
- Sander, A.L.; Sommer, K.; Neumayer, T.; Fleming, I.; Marzi, I.; Barker, J.H.; Frank, J.; Jakob, H. Soluble epoxide hydrolase disruption as therapeutic target for wound healing. J. Surg. Res. 2013, 182, 362–367. [Google Scholar] [CrossRef]
- Sommer, K.; Jakob, H.; Badjlan, F.; Henrich, D.; Frank, J.; Marzi, I.; Sander, A.L. 11,12 and 14,15 epoxyeicosatrienoic acid rescue deteriorated wound healing in ischemia. PLoS ONE 2019, 14, e0209158. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, J.; Chai, J.; Zhang, Y.; Yu, C.; Pan, Z.; Gao, P.; Zong, C.; Guan, Q.; Fu, Y.; et al. Cytochrome P450 (CYP) epoxygenases as potential targets in the management of impaired diabetic wound healing. Lab. Investig. 2017, 97, 782–791. [Google Scholar] [CrossRef]
- Supp, D.M.; Hahn, J.M.; McFarland, K.L.; Combs, K.A.; Lee, K.S.; Inceoglu, B.; Wan, D.; Boyce, S.T.; Hammock, B.D. Soluble Epoxide Hydrolase Inhibition and Epoxyeicosatrienoic Acid Treatment Improve Vascularization of Engineered Skin Substitutes. Plast Reconstr. Surg. Glob. Open 2016, 4, e1151. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Hellmann, J.; Tang, Y.; Spite, M. Proresolving lipid mediators and diabetic wound healing. Curr. Opin. Endocrinol. Diabetes Obes. 2012, 19, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Reis, M.B.; Pereira, P.A.T.; Caetano, G.F.; Leite, M.N.; Galvao, A.F.; Paula-Silva, F.W.G.; Frade, M.A.C.; Faccioli, L.H. Lipoxin A4 encapsulated in PLGA microparticles accelerates wound healing of skin ulcers. PLoS ONE 2017, 12, e0182381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, B.S.; Kantarci, A.; Zarrough, A.; Hasturk, H.; Leung, K.P.; Van Dyke, T.E. LXA4 actions direct fibroblast function and wound closure. Biochem. Biophys. Res. Commun. 2015, 464, 1072–1077. [Google Scholar] [CrossRef] [Green Version]
- Arita, M.; Bianchini, F.; Aliberti, J.; Sher, A.; Chiang, N.; Hong, S.; Yang, R.; Petasis, N.A.; Serhan, C.N. Stereochemical assignment, antiinflammatory properties, and receptor for the omega-3 lipid mediator resolvin E1. J. Exp. Med. 2005, 201, 713–722. [Google Scholar] [CrossRef]
- Arita, M.; Ohira, T.; Sun, Y.P.; Elangovan, S.; Chiang, N.; Serhan, C.N. Resolvin E1 selectively interacts with leukotriene B4 receptor BLT1 and ChemR23 to regulate inflammation. J. Immunol. 2007, 178, 3912–3917. [Google Scholar] [CrossRef] [Green Version]
- Menon, R.; Krzyszczyk, P.; Berthiaume, F. Pro-Resolution Potency of Resolvins D1, D2 and E1 on Neutrophil Migration and in Dermal Wound Healing. Nano Life 2017, 07, 1750002. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.; Tian, H.; Lu, Y.; Laborde, J.M.; Muhale, F.A.; Wang, Q.; Alapure, B.V.; Serhan, C.N.; Bazan, N.G. Neuroprotectin/protectin D1: Endogenous biosynthesis and actions on diabetic macrophages in promoting wound healing and innervation impaired by diabetes. Am. J. Physiol Cell Physiol 2014, 307, C1058–C1067. [Google Scholar] [CrossRef] [Green Version]
- Cezar, T.L.C.; Martinez, R.M.; Rocha, C.D.; Melo, C.P.B.; Vale, D.L.; Borghi, S.M.; Fattori, V.; Vignoli, J.A.; Camilios-Neto, D.; Baracat, M.M.; et al. Treatment with maresin 1, a docosahexaenoic acid-derived pro-resolution lipid, protects skin from inflammation and oxidative stress caused by UVB irradiation. Sci. Rep. 2019, 9, 3062. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.W.; Yu, S.H.; Fretwurst, T.; Larsson, L.; Sugai, J.V.; Oh, J.; Lehner, K.; Jin, Q.; Giannobile, W.V. Maresin 1 Promotes Wound Healing and Socket Bone Regeneration for Alveolar Ridge Preservation. J. Dent. Res. 2020, 99, 930–937. [Google Scholar] [CrossRef]
- Ito, K.; Ito, S.; Sekine, M.; Abe, M. Reconstruction of the soft tissue of a deep diabetic foot wound with artificial dermis and recombinant basic fibroblast growth factor. Plast. Reconstr. Surg. 2005, 115, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Tai, H.H.; Ensor, C.M.; Tong, M.; Zhou, H.; Yan, F. Prostaglandin catabolizing enzymes. Prostaglandins Other Lipid Mediat. 2002, 68–69, 483–493. [Google Scholar] [CrossRef]
- Clish, C.B.; Levy, B.D.; Chiang, N.; Tai, H.H.; Serhan, C.N. Oxidoreductases in lipoxin A4 metabolic inactivation: A novel role for 15-onoprostaglandin 13-reductase/leukotriene B4 12-hydroxydehydrogenase in inflammation. J. Biol. Chem. 2000, 275, 25372–25380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolete, R.; Lima, K.d.M.; Júnior, J.M.R.; Baruffi, M.D.; de Medeiros, A.I.; Bentley, M.V.L.B.; Silva, C.L.; Faccioli, L.H. In vitro and in vivo activities of leukotriene B4-loaded biodegradable microspheres. Prostaglandins Other Lipid Mediat. 2007, 83, 121–129. [Google Scholar] [CrossRef]
- Park, S.K.; Herrnreiter, A.; Pfister, S.L.; Gauthier, K.M.; Falck, B.A.; Falck, J.R.; Campbell, W.B. GPR40 is a low-affinity epoxyeicosatrienoic acid receptor in vascular cells. J. Biol. Chem. 2018, 293, 10675–10691. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.V.; Vik, A.; Serhan, C.N. The Protectin Family of Specialized Pro-resolving Mediators: Potent Immunoresolvents Enabling Innovative Approaches to Target Obesity and Diabetes. Front. Pharmacol. 2018, 9, 1582. [Google Scholar] [CrossRef]
- Bang, S.; Xie, Y.K.; Zhang, Z.J.; Wang, Z.; Xu, Z.Z.; Ji, R.R. GPR37 regulates macrophage phagocytosis and resolution of inflammatory pain. J. Clin. Investig. 2018, 128, 3568–3582. [Google Scholar] [CrossRef] [Green Version]
- Chiang, N.; Libreros, S.; Norris, P.C.; de la Rosa, X.; Serhan, C.N. Maresin 1 activates LGR6 receptor promoting phagocyte immunoresolvent functions. J. Clin. Investig. 2019, 129, 5294–5311. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Eicosanoid and SPM | Source | Biosynthetic Pathway | Function Related to Skin Wound Healing | Reference |
|---|---|---|---|---|
| TxA2 | AA | COX-1/2 TxAS | Platelet aggregation (Hemostasis) ↑ Inflammation ↑ Endothelial migration and angiogenesis ↑ (in vitro) | [50,51] [54] [55,56] |
| 12-HHT | AA | COX-1/2 TxAS | Keratinocyte migration ↑ Fibroblast proliferation ↑ | [58] [59] |
| PGE2 | AA | COX-1/2 cPGES or mPGES | Angiogenesis ↑ Fibroblast proliferation ↑ Inflammation ↓ Keratinocyte proliferation and differentiation ↑ Macrophage polarization to M2 phenotype ↑ Collagen synthesis/Fibrosis ↓ | [65,66] [65,66] [69] [70,71,72,73,74] [76,77] [78] |
| PGD2 | AA | COX-1/2 H-PGDS or L-PGDS | Cutaneous barrier function ↑ Macrophage polarization to M2 phenotype ↑ Fibroblast migration ↓ (in vitro) Angiogenesis ↓ Hair follicle neogenesis ↓ | [82,83] [84,85,86,87,88] [89] [90] [91,92] |
| PGF2α | AA | COX-1/2 PGFS | Unknown | - |
| PGI2 | AA | COX-1/2 PGIS | Fibrinolysis ↑ Fibroblast migration ↑ (in vitro) Angiogenesis ↑ | [101] [101] [102,103] |
| LTB4 | AA | 5-LOX LTA4H | ROS production ↑ Inflammation ↑ Macrophage polarization to M2 phenotype ↓ | [110] [111] [112] |
| CysLTs (LTC4, LTD4, and LTE4) | AA | 5-LOX LTC4S | ROS production ↑ Inflammation ↑ Macrophage polarization to M2 phenotype ↓ Collagen deposition ↑ | [110] [111] [112] [113] |
| 12-HETE | AA | 12/15-LOX | Keratinocyte proliferation and migration ↑ (in vitro) | [115,116] |
| EETs (5,6-, 8,9-, 11,12-, and 14,15-EET) | AA | CYP2C or CYP2J | Angiogenesis ↑ Inflammation ↓ Collagen deposition ↑ | [118,121,122] [119,120,121] [121] |
| LXA4 | AA | 5-LOX 12- or 15-LOX | Inflammation ↓ Angiogenesis ↑ Collagen deposition ↑ Fibroblast proliferation and migration ↓ (in vitro) | [125] [125] [125] [126] |
| RvE1 | EPA | CYP LOX | Inflammation ↓ Collagen deposition ↑ | [127,128] [129] |
| RvD1, RvD2, and RvD4 | DHA | LOX | Collagen deposition ↑ Keratinocyte migration ↑ Fibroblast proliferation and migration ↓ (in vitro) | [129] [27] [126] |
| PD1 | DHA | LOX | Inflammation ↓ Nerve fiber growth ↑ Re-epithelialization ↑ Collagen deposition ↑ | [130] [130] [130] [130] |
| MaR1 | DHA | LOX | Inflammation ↓ Re-epithelialization ↑ Macrophage polarization to M2 phenotype ↑ | [131,132] [132] [132] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasukawa, K.; Okuno, T.; Yokomizo, T. Eicosanoids in Skin Wound Healing. Int. J. Mol. Sci. 2020, 21, 8435. https://doi.org/10.3390/ijms21228435
Yasukawa K, Okuno T, Yokomizo T. Eicosanoids in Skin Wound Healing. International Journal of Molecular Sciences. 2020; 21(22):8435. https://doi.org/10.3390/ijms21228435
Chicago/Turabian StyleYasukawa, Ken, Toshiaki Okuno, and Takehiko Yokomizo. 2020. "Eicosanoids in Skin Wound Healing" International Journal of Molecular Sciences 21, no. 22: 8435. https://doi.org/10.3390/ijms21228435
APA StyleYasukawa, K., Okuno, T., & Yokomizo, T. (2020). Eicosanoids in Skin Wound Healing. International Journal of Molecular Sciences, 21(22), 8435. https://doi.org/10.3390/ijms21228435