Aux/IAA14 Regulates microRNA-Mediated Cold Stress Response in Arabidopsis Roots




Abstract
:1. Introduction
2. Results
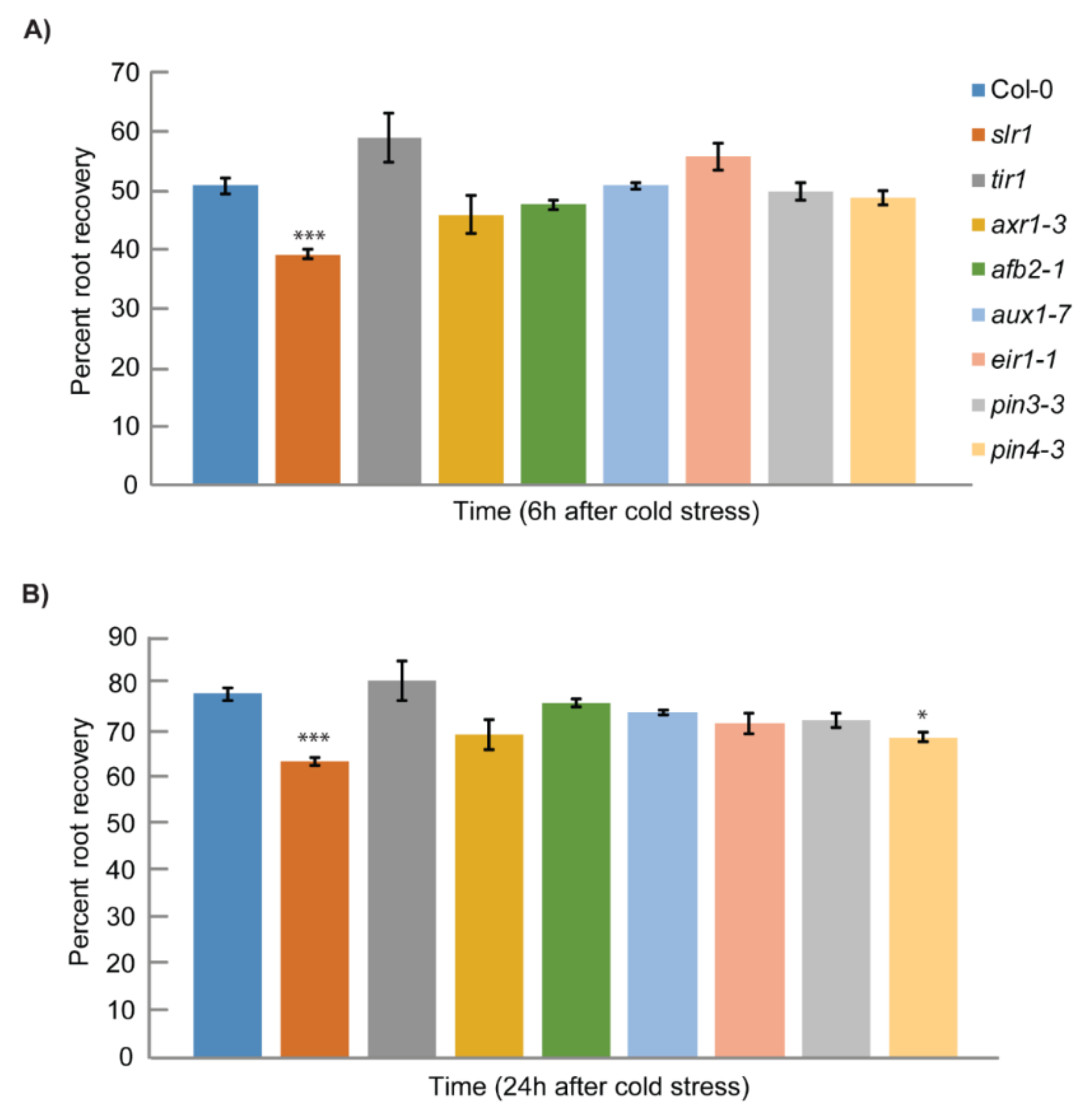
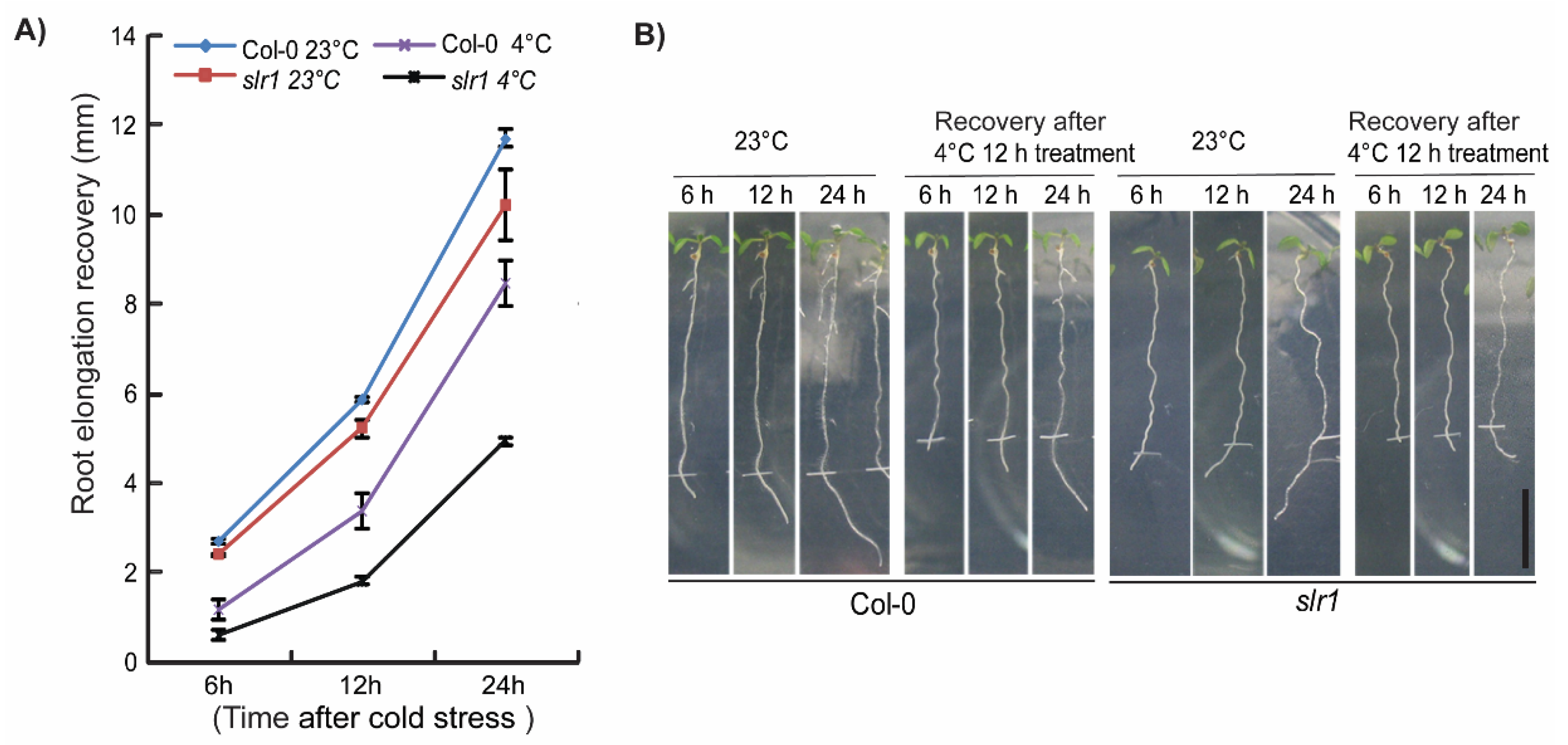
2.1. Auxin Signaling Mutant slr1 Is Susceptible to Low Temperature
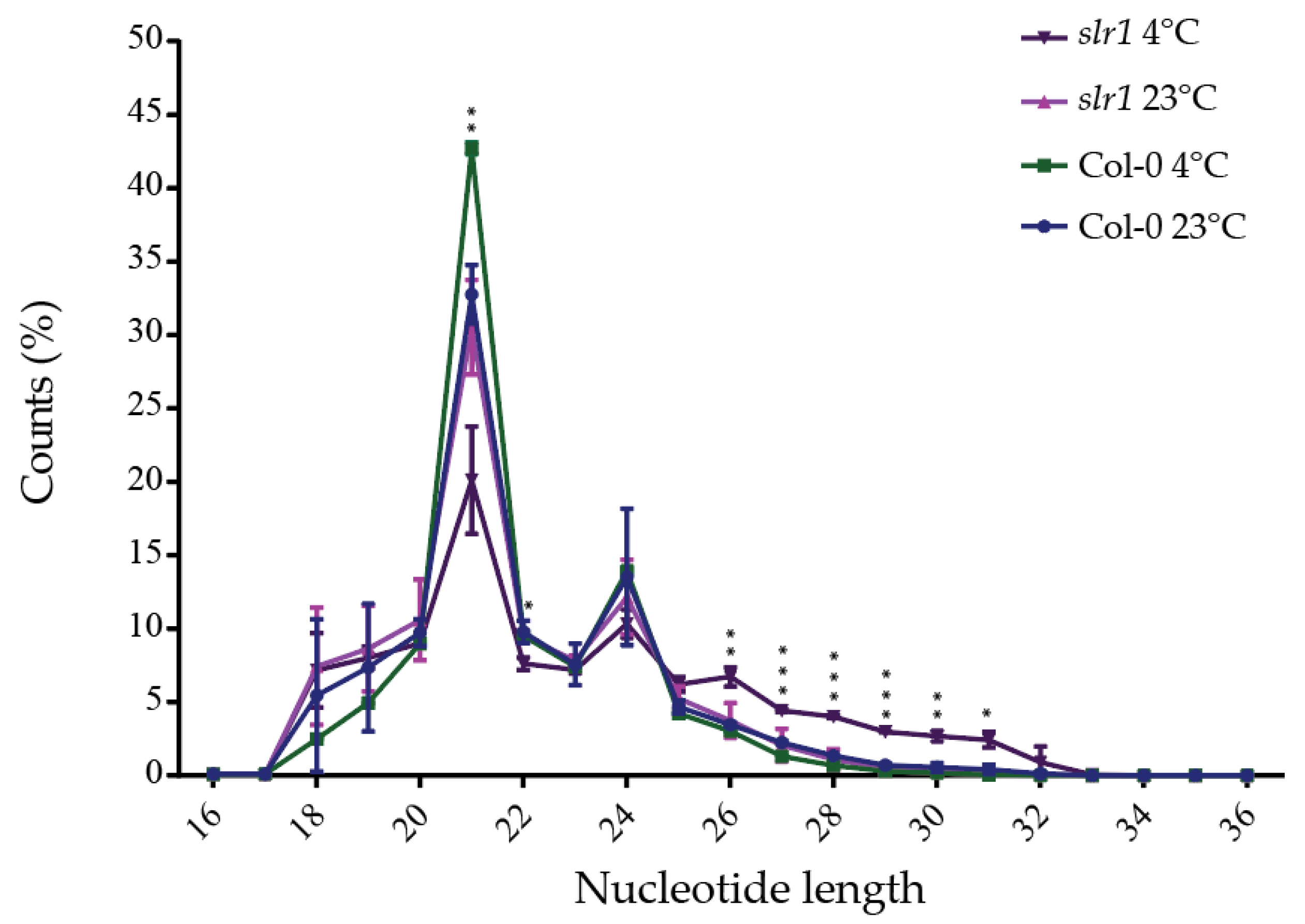
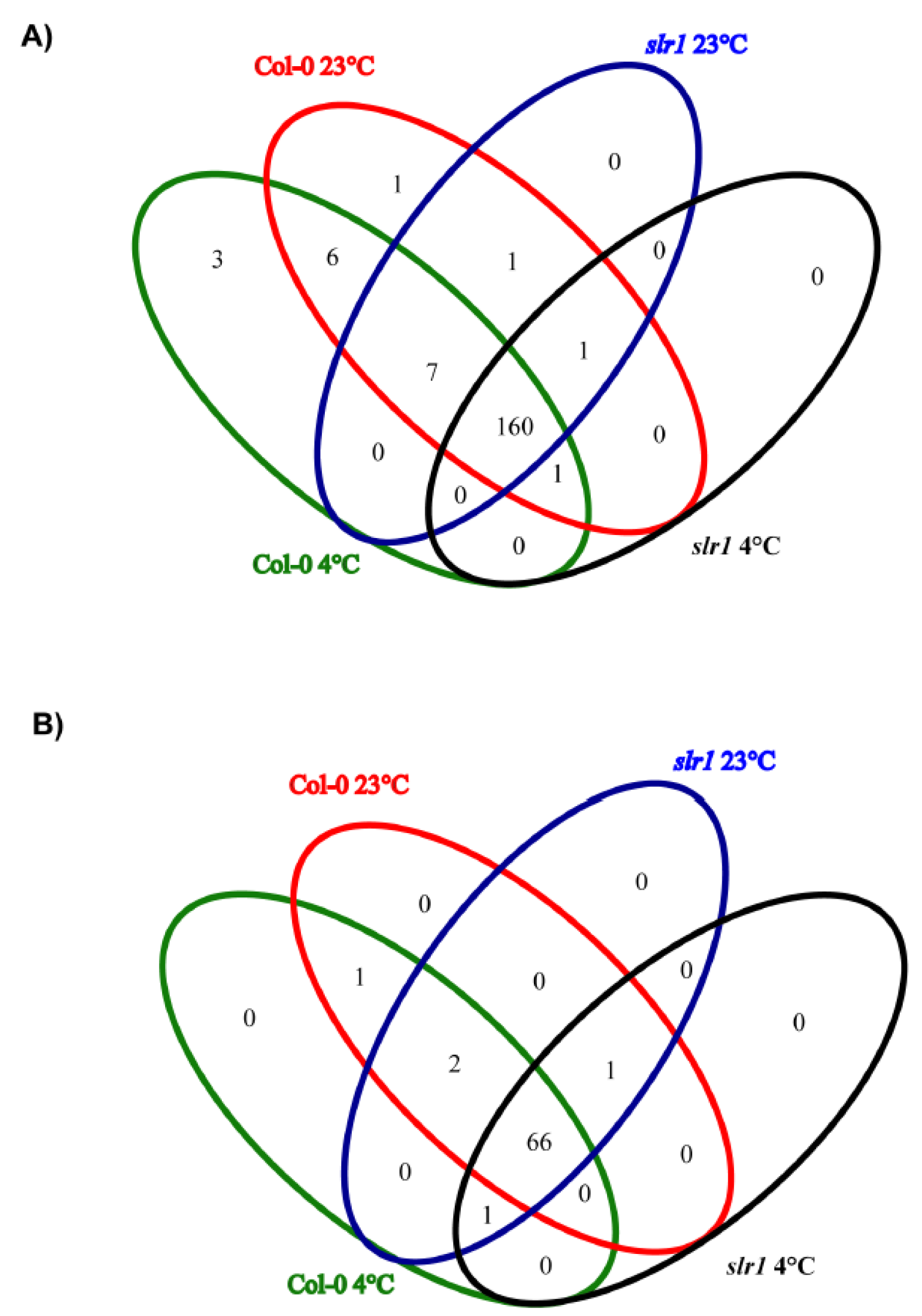
2.2. High-Throughput Sequencing of Small RNA Libraries
2.3. Identification of Known and Novel miRs
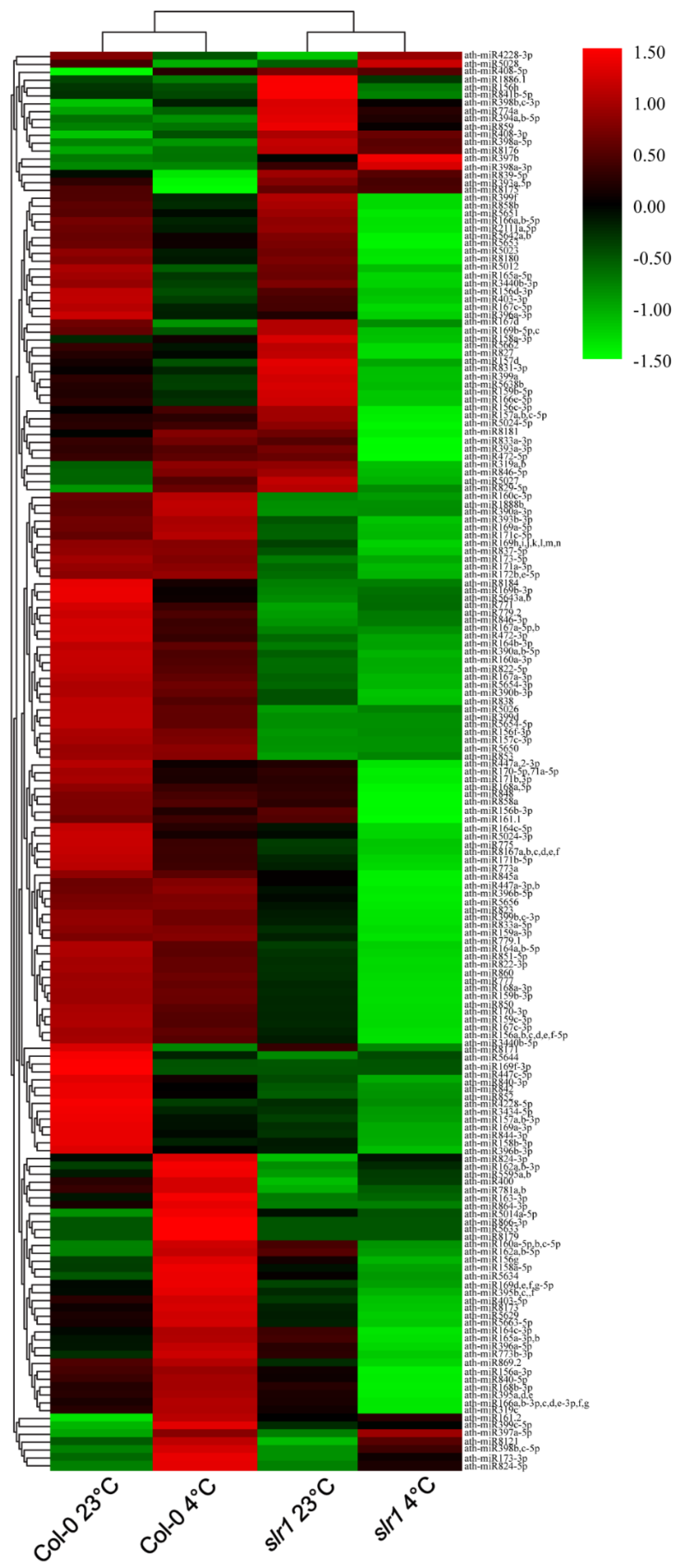
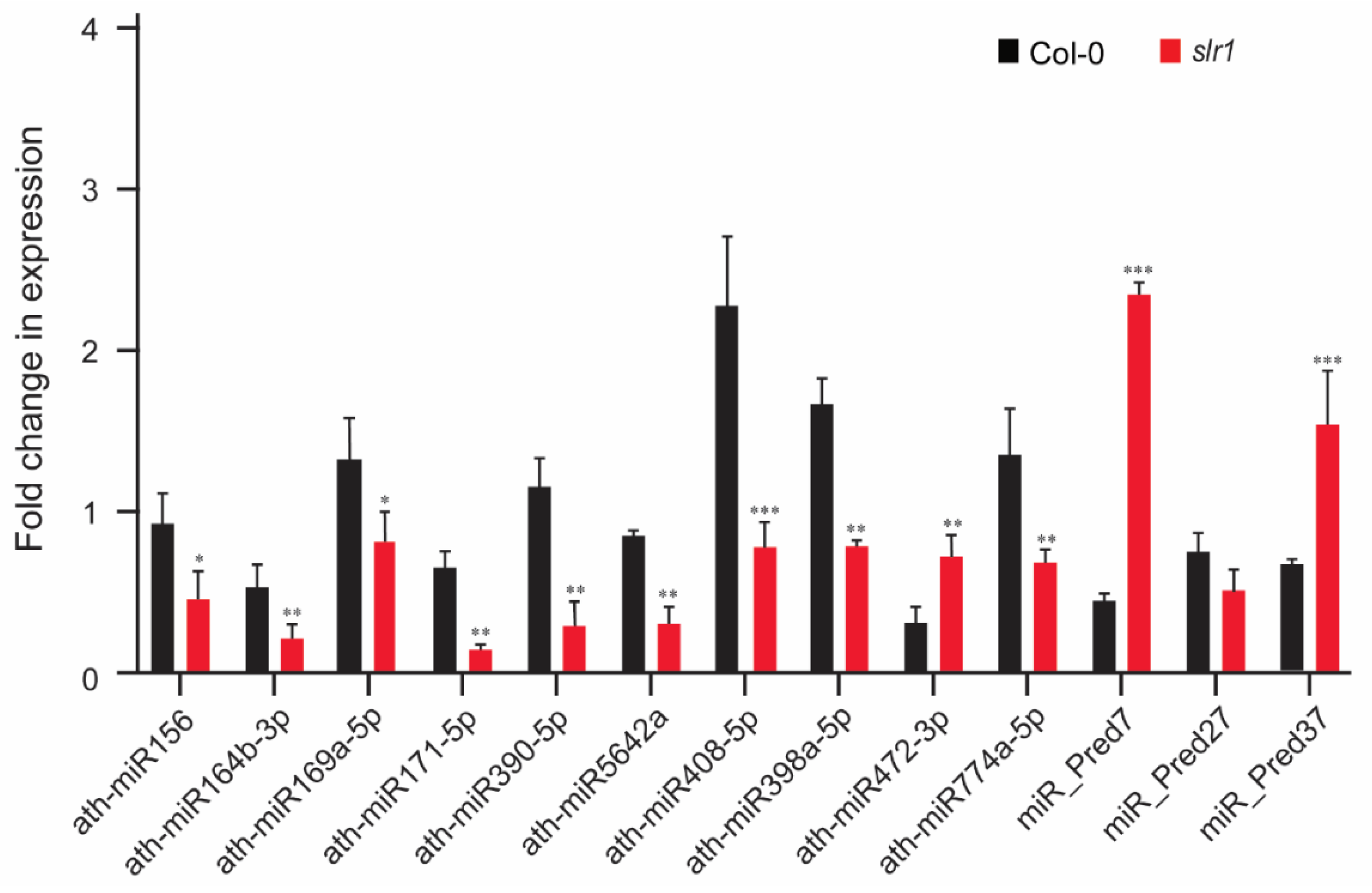
2.4. Validation of miR Expression Patterns
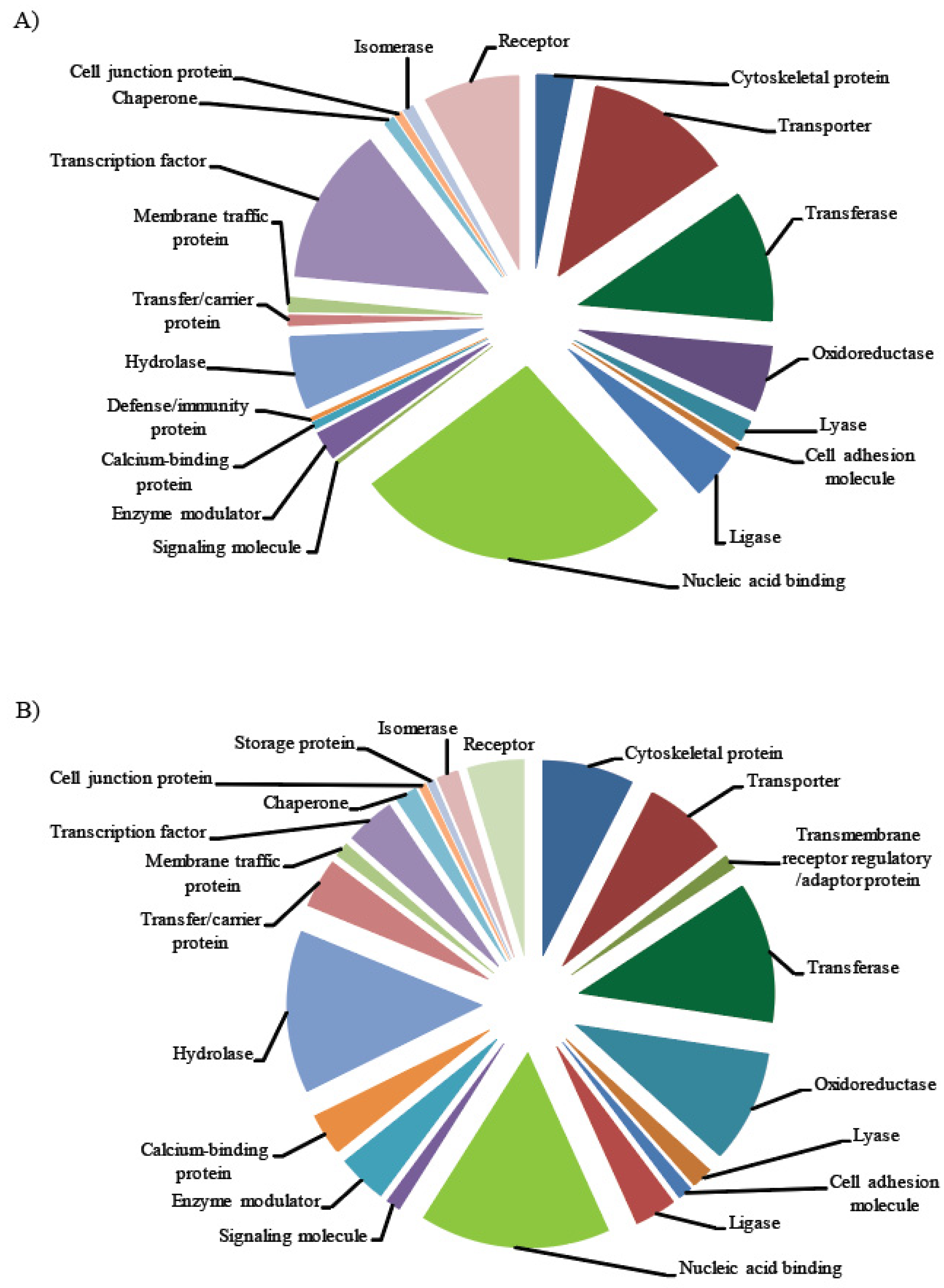
2.5. miR Target Prediction
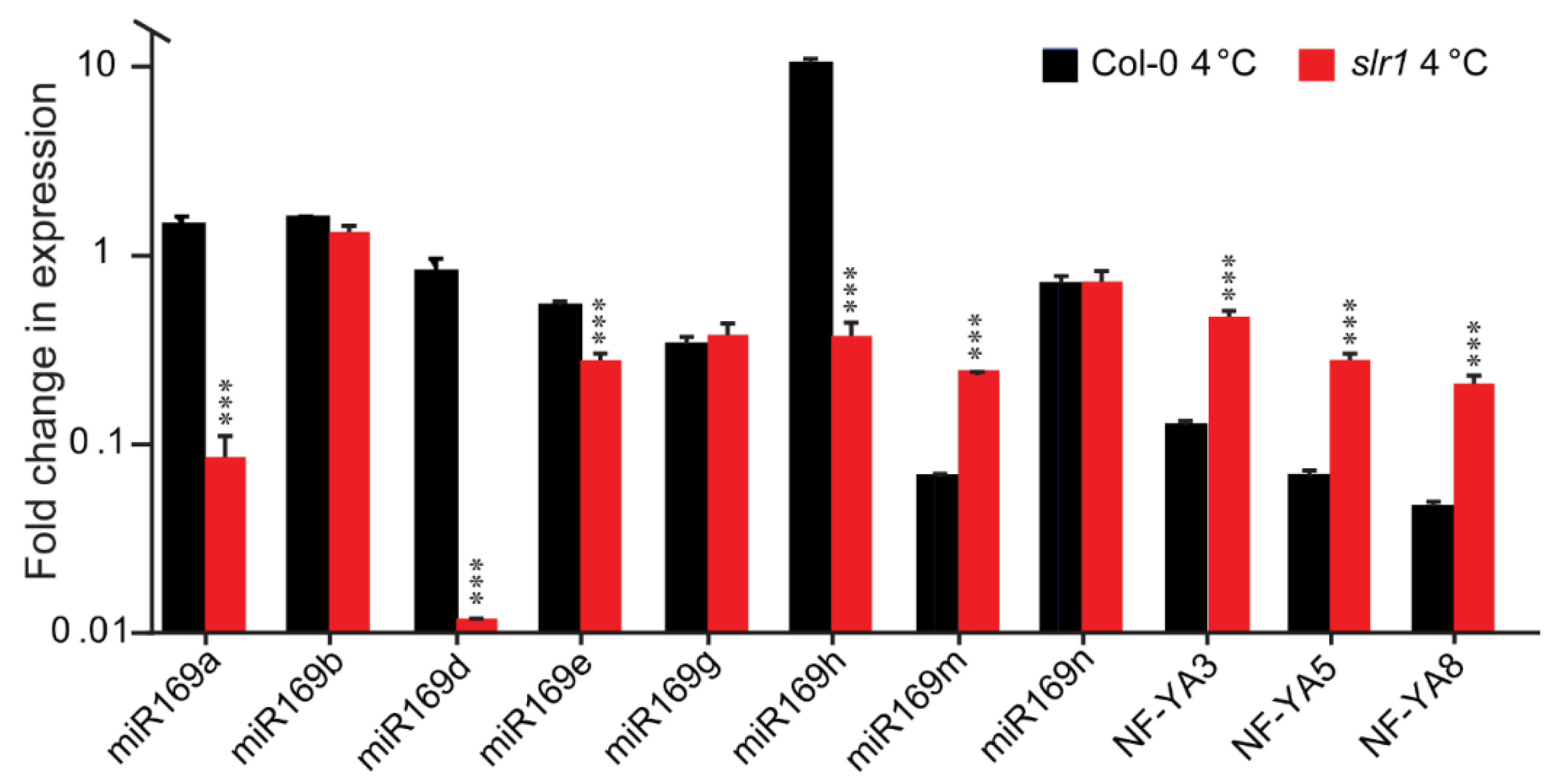
2.6. miR169/NF-YA Module Is Altered in slr1 under Low-Temperature Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Condition
4.2. Cold Stress Treatment and Analysis of Root Growth Recovery
4.3. Chemicals
4.4. Small RNA Isolation and Sequencing
4.5. Data Deposition Information
4.6. Bioinformatic Analysis of the sRNA Sequencing Data
4.7. Prediction of miR Targets
4.8. Quantitative RT-PCR Validation of Selected Differentially Expressed miRs
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| miRs | MicroRNAs |
| qRT-PCR | Quantitative Real-Time PCR |
| EF1α | Elongation Factor 1-α |
| TEs | Transposable Elements |
| nt | Nucleotide |
| IAA | Indole-3-aCetic Acid |
| ARF | Auxin Response Factor |
| QC | Quiescent Center |
| SIZ1 | SAP and MIZ1 domain-containing ligase1 |
| PIN | Pin-formed |
| SAUR | Small Auxin Upregulated RNA |
| GH | Gretchen Hagen |
References
- Rahman, A. Auxin: A regulator of cold stress response. Physiol. Plant 2013, 147, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Sanghera, G.S.; Wani, S.H.; Hussain, W.; Singh, N.B. Engineering cold stress tolerance in crop plants. Curr. Genom. 2011, 12, 30–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, S.K. Cold stress tolerance mechanisms in plants. A review. Agron. Sustain. Dev. 2010, 30, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Thakur, P.; Kumar, S.; Malik, J.A.; Berger, J.D.; Nayyar, H. Cold stress effects on reproductive development in grain crops: An overview. Environ. Exp. Bot. 2010, 67, 429–443. [Google Scholar] [CrossRef]
- Chandler, J.W. Auxin as compère in plant hormone crosstalk. Planta 2009, 231, 1–12. [Google Scholar] [CrossRef]
- Davies, P.J. The plant hormones: Their nature, occurrence, and functions. In Plant Hormones: Biosynthesis, Signal Transduction, Action! Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 1–15. [Google Scholar] [CrossRef]
- Gray, W.M.; Ostin, A.; Sandberg, G.; Romano, C.P.; Estelle, M. High temperature promotes auxin-mediated hypocotyl elongation in Arabidopsis. Proc. Natl. Acad. Sci. USA 1998, 95, 7197–7202. [Google Scholar] [CrossRef] [Green Version]
- Koini, M.A.; Alvey, L.; Allen, T.; Tilley, C.A.; Harberd, N.P.; Whitelam, G.C.; Franklin, K.A. High temperature-mediated adaptations in plant architecture require the bHLH transcription factor PIF4. Curr. Biol. 2009, 19, 408–413. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, K.; Uemura, M.; Tsurumi, S.; Rahman, A. Auxin response in Arabidopsis under cold stress: Underlying molecular mechanisms. Plant Cell 2009, 21, 3823–3838. [Google Scholar] [CrossRef] [Green Version]
- Hanzawa, T.; Shibasaki, K.; Numata, T.; Kawamura, Y.; Gaude, T.; Rahman, A. Cellular auxin homeostasis under high temperature is regulated through a sorting NEXIN1-dependent endosomal trafficking pathway. Plant Cell 2013, 25, 3424–3433. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.A.; Rahman, A. Hormonal regulation of cold stress response. In Cold Tolerance in Plants: Physiological, Molecular and Genetic Perspectives; Wani, S.H., Herath, V., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 65–88. [Google Scholar] [CrossRef]
- Hannah, M.A.; Heyer, A.G.; Hincha, D.K. A global survey of gene regulation during cold acclimation in Arabidopsis thaliana. PLoS Genet. 2005, 1, e26. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Rahman, A. Cold stress response in Arabidopsis thaliana is mediated by GNOM ARF-GEF. Plant J. 2019, 97, 500–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.H.; Savina, M.; Du, J.; Devendran, A.; Kannivadi Ramakanth, K.; Tian, X.; Sim, W.S.; Mironova, V.V.; Xu, J. A sacrifice-for-survival mechanism protects root stem cell niche from chilling stress. Cell 2017, 170, 102–113.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, H.; Liu, H.; Xiong, L. Endogenous auxin and jasmonic acid levels are differentially modulated by abiotic stresses in rice. Front. Plant Sci. 2013, 4, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Zhang, S.; Yang, T.; Zeng, Z.; Huang, Z.; Liu, Q.; Wang, X.; Leach, J.; Leung, H.; Liu, B. Global transcriptional profiling of a cold-tolerant rice variety under moderate cold stress reveals different cold stress response mechanisms. Physiol Plant 2015, 154, 381–394. [Google Scholar] [CrossRef]
- Jain, M.; Khurana, J.P. Transcript profiling reveals diverse roles of auxin-responsive genes during reproductive development and abiotic stress in rice. FEBS J. 2009, 276, 3148–3162. [Google Scholar] [CrossRef]
- Du, H.; Wu, N.; Fu, J.; Wang, S.; Li, X.; Xiao, J.; Xiong, L. A GH3 family member, OsGH3-2, modulates auxin and abscisic acid levels and differentially affects drought and cold tolerance in rice. J. Exp. Bot. 2012, 63, 6467–6480. [Google Scholar] [CrossRef] [Green Version]
- Miura, K.; Jin, J.B.; Lee, J.; Yoo, C.Y.; Stirm, V.; Miura, T.; Ashworth, E.N.; Bressan, R.A.; Yun, D.J.; Hasegawa, P.M. SIZ1-mediated sumoylation of ICE1 controls CBF3/DREB1A expression and freezing tolerance in Arabidopsis. Plant Cell 2007, 19, 1403–1414. [Google Scholar] [CrossRef] [Green Version]
- Miura, K.; Lee, J.; Gong, Q.; Ma, S.; Jin, J.B.; Yoo, C.Y.; Miura, T.; Sato, A.; Bohnert, H.J.; Hasegawa, P.M. SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation. Plant Physiol. 2011, 155, 1000–1012. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.H.; Hu, X.; Tang, W.; Zheng, X.; Kim, Y.S.; Lee, B.H.; Zhu, J.K. A putative Arabidopsis nucleoporin, AtNUP160, is critical for RNA export and required for plant tolerance to cold stress. Mol. Cell Biol. 2006, 26, 9533–9543. [Google Scholar] [CrossRef] [Green Version]
- Parry, G.; Ward, S.; Cernac, A.; Dharmasiri, S.; Estelle, M. The Arabidopsis SUPPRESSOR OF AUXIN RESISTANCE proteins are nucleoporins with an important role in hormone signaling and development. Plant Cell 2006, 18, 1590–1603. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, M.; Ghazanfari, F.; Fadaei, A.; Ahmadi, L.; Shiran, B.; Rabei, M.; Fallahi, H. The small-RNA profiles of almond (Prunus dulcis Mill.) reproductive tissues in response to cold stress. PLoS ONE 2016, 11, e0156519. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Liu, Y.; Li, W.; Song, L.; Zhang, J.; Guo, C. Genome-wide investigation of microRNAs and their targets in response to freezing stress in Medicago sativa L., based on high-throughput sequencing. G3 (Bethesda) 2016, 6, 755–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, G.; Zhang, R.; Zhang, S.; Li, Y.; Gao, J.; Han, X.; Chen, M.; Wang, J.; Li, W.; Li, G. Response of microRNAs to cold treatment in the young spikes of common wheat. BMC Genom. 2017, 18, 212. [Google Scholar] [CrossRef] [Green Version]
- Tyczewska, A.; Gracz, J.; Kuczyński, J.; Twardowski, T. Deciphering the soybean molecular stress response via high-throughput approaches. Acta Biochim. Pol. 2016, 63, 631–643. [Google Scholar] [CrossRef]
- Bej, S.; Basak, J. MicroRNAs: The potential biomarkers in plant stress response. Am. J. Plant Sci. 2014, 5, 748–759. [Google Scholar] [CrossRef] [Green Version]
- Megha, S.; Basu, U.; Kav, N.N.V. Regulation of low temperature stress in plants by microRNAs. Plant Cell Environ. 2018, 41, 1–15. [Google Scholar] [CrossRef]
- Sunkar, R.; Li, Y.F.; Jagadeeswaran, G. Functions of microRNAs in plant stress responses. Trends Plant Sci. 2012, 17, 196–203. [Google Scholar] [CrossRef]
- Lv, D.K.; Bai, X.; Li, Y.; Ding, X.D.; Ge, Y.; Cai, H.; Ji, W.; Wu, N.; Zhu, Y.M. Profiling of cold-stress-responsive miRNAs in rice by microarrays. Gene 2010, 459, 39–47. [Google Scholar] [CrossRef]
- Abdel-Ghany, S.E.; Pilon, M. MicroRNA-mediated systemic down-regulation of copper protein expression in response to low copper availability in Arabidopsis. J. Biol. Chem. 2008, 283, 15932–15945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Burd, S.; Lers, A. miR408 is involved in abiotic stress responses in Arabidopsis. Plant J. 2015, 84, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.-H.; Pei, H. Over-expression of miR397 improves plant tolerance to cold stress in Arabidopsis thaliana. J. Plant Biol. 2014, 57, 209–217. [Google Scholar] [CrossRef]
- Song, J.B.; Gao, S.; Wang, Y.; Li, B.W.; Zhang, Y.L.; Yang, Z.M. miR394 and its target gene LCR are involved in cold stress response in Arabidopsis. Plant Gene 2016, 5, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.-T.; Sun, X.-L.; Hoshino, Y.; Yu, Y.; Jia, B.; Sun, Z.-W.; Sun, M.-Z.; Duan, X.-B.; Zhu, Y.-M. MicroRNA319 positively regulates cold tolerance by targeting OsPCF6 and OsTCP21 in rice (Oryza sativa L.). PLoS ONE 2014, 9, e91357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Li, D.; Mao, D.; Liu, X.; Ji, C.; Li, X.; Zhao, X.; Cheng, Z.; Chen, C.; Zhu, L. Overexpression of microRNA319 impacts leaf morphogenesis and leads to enhanced cold tolerance in rice (Oryza sativa L.). Plant Cell Environ. 2013, 36, 2207–2218. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Wang, M.; Zhang, H.Y.; Liu, J.H. The miR396b of Poncirus trifoliata functions in cold tolerance by regulating ACC oxidase gene expression and modulating ethylene-polyamine homeostasis. Plant Cell Physiol. 2016, 57, 1865–1878. [Google Scholar] [CrossRef] [Green Version]
- Bai, B.; Bian, H.; Zeng, Z.; Hou, N.; Shi, B.; Wang, J.; Zhu, M.; Han, N. miR393-mediated auxin signaling regulation is involved in root elongation inhibition in response to toxic aluminum stress in Barley. Plant Cell Physiol. 2017, 58, 426–439. [Google Scholar] [CrossRef]
- Gray, W.M.; Kepinski, S.; Rouse, D.; Leyser, O.; Estelle, M. Auxin regulates SCF(TIR1)-dependent degradation of AUX/IAA proteins. Nature 2001, 414, 271–276. [Google Scholar] [CrossRef]
- Jones-Rhoades, M.W.; Bartel, D.P. Computational identification of plant microRNAs and their targets, including a stress-induced miRNA. Mol. Cell 2004, 14, 787–799. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, K.; Li, D.; Yan, J.; Zhang, W. Enhanced cold tolerance and tillering in Switchgrass (Panicum virgatum L.) by heterologous expression of Osa-miR393a. Plant Cell Physiol. 2017, 58, 2226–2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Chen, D.; Ma, X.; Mao, C.; Cao, J.; Wu, P.; Chen, M. Mechanisms of microRNA-mediated auxin signaling inferred from the rice mutant osaxr. Plant Signal. Behav. 2010, 5, 252–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, K.; Chen, L.; Zou, Y.; Liu, H.; Tian, Y.; Li, D.; Wang, R.; Zhao, F.; Ferguson, B.J.; et al. MicroRNA167-directed regulation of the auxin response factors GmARF8a and GmARF8b is required for soybean nodulation and lateral root development. Plant Physiol. 2015, 168, 984–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Lai, Z.; Tian, Q.; Lin, L.; Lai, R.; Yang, M.; Zhang, D.; Chen, Y.; Zhang, Z. Endogenous target mimics down-regulate miR160 mediation of ARF10, -16, and -17 cleavage during somatic embryogenesis in Dimocarpus longan Lour. Front. Plant Sci. 2015, 6, 956. [Google Scholar] [CrossRef]
- Rhoades, M.W.; Reinhart, B.J.; Lim, L.P.; Burge, C.B.; Bartel, B.; Bartel, D.P. Prediction of plant microRNA targets. Cell 2002, 110, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Teotia, P.S.; Mukherjee, S.K.; Mishra, N.S. Fine tuning of auxin signaling by miRNAs. Physiol. Mol. Biol. Plants 2008, 14, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Mallory, A.C.; Bartel, D.P.; Bartel, B. MicroRNA-directed regulation of Arabidopsis AUXIN RESPONSE FACTOR17 is essential for proper development and modulates expression of early auxin response genes. Plant Cell 2005, 17, 1360–1375. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zeng, Z.; Chen, C.; Li, C.; Xia, R.; Li, J. Genome-wide characterization of the auxin response factor (ARF) gene family of Litchi (Litchi chinensis Sonn.): Phylogenetic analysis, miRNA regulation and expression changes during fruit abscission. PeerJ 2019, 7, e6677. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Jia, S.; Shen, D.; Liu, J.; Li, J.; Zhao, H.; Han, S.; Wang, Y. Four AUXIN RESPONSE FACTOR genes downregulated by microRNA167 are associated with growth and development in Oryza sativa. Funct. Plant Biol. 2012, 39, 736–744. [Google Scholar] [CrossRef]
- Liu, N.; Wu, S.; Van Houten, J.; Wang, Y.; Ding, B.; Fei, Z.; Clarke, T.H.; Reed, J.W.; van der Knaap, E. Down-regulation of AUXIN RESPONSE FACTORS 6 and 8 by microRNA 167 leads to floral development defects and female sterility in tomato. J. Exp. Bot. 2014, 65, 2507–2520. [Google Scholar] [CrossRef] [Green Version]
- Sanan-Mishra, N.; Varanasi, S.P.; Mukherjee, S.K. Micro-regulators of auxin action. Plant Cell Rep. 2013, 32, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Yang, C.; Liu, S.; Qi, H.; Wu, L.; Xu, L.A.; Xu, M. MiRNA-target pairs regulate adventitious rooting in Populus: A functional role for miR167a and its target Auxin response factor 8. Tree Physiol. 2019, 39, 1922–1936. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.S.; Xie, Q.; Fei, J.F.; Chua, N.H. MicroRNA directs mRNA cleavage of the transcription factor NAC1 to downregulate auxin signals for Arabidopsis lateral root development. Plant Cell 2005, 17, 1376–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukaki, H.; Tameda, S.; Masuda, H.; Tasaka, M. Lateral root formation is blocked by a gain-of-function mutation in the SOLITARY-ROOT/IAA14 gene of Arabidopsis. Plant J. 2002, 29, 153–168. [Google Scholar] [CrossRef]
- Dharmasiri, N.; Dharmasiri, S.; Estelle, M. The F-box protein TIR1 is an auxin receptor. Nature 2005, 435, 441–445. [Google Scholar] [CrossRef]
- Leyser, H.M.; Lincoln, C.A.; Timpte, C.; Lammer, D.; Turner, J.; Estelle, M. Arabidopsis auxin-resistance gene AXR1 encodes a protein related to ubiquitin-activating enzyme E1. Nature 1993, 364, 161–164. [Google Scholar] [CrossRef]
- Dharmasiri, N.; Dharmasiri, S.; Weijers, D.; Lechner, E.; Yamada, M.; Hobbie, L.; Ehrismann, J.S.; Jurgens, G.; Estelle, M. Plant development is regulated by a family of auxin receptor F box proteins. Dev. Cell 2005, 9, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Pickett, F.B.; Wilson, A.K.; Estelle, M. The aux1 mutation of Arabidopsis confers both auxin and ethylene resistance. Plant Physiol. 1990, 94, 1462–1466. [Google Scholar] [CrossRef] [Green Version]
- Luschnig, C.; Gaxiola, R.A.; Grisafi, P.; Fink, G.R. EIR1, a root-specific protein involved in auxin transport, is required for gravitropism in Arabidopsis thaliana. Genes Dev. 1998, 12, 2175–2187. [Google Scholar] [CrossRef] [Green Version]
- Friml, J.; Wisniewska, J.; Benkova, E.; Mendgen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 2002, 415, 806–809. [Google Scholar] [CrossRef] [Green Version]
- Friml, J.; Benkova, E.; Blilou, I.; Wisniewska, J.; Hamann, T.; Ljung, K.; Woody, S.; Sandberg, G.; Scheres, B.; Jurgens, G.; et al. AtPIN4 mediates sink-driven auxin gradients and root patterning in Arabidopsis. Cell 2002, 108, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, F.; Axtell, M.J. Analysis of complementarity requirements for plant microRNA targeting using a Nicotiana benthamiana quantitative transient assay. Plant Cell 2014, 26, 741–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallory, A.C.; Reinhart, B.J.; Jones-Rhoades, M.W.; Tang, G.; Zamore, P.D.; Barton, M.K.; Bartel, D.P. MicroRNA control of PHABULOSA in leaf development: Importance of pairing to the microRNA 5’ region. EMBO J. 2004, 23, 3356–3364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwakawa, H.O.; Tomari, Y. Molecular insights into microRNA-mediated translational repression in plants. Mol. Cell 2013, 52, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwakawa, H.O.; Tomari, Y. The functions of microRNAs: mRNA decay and translational repression. Trends Cell Biol. 2015, 25, 651–665. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Kuang, Z.; Wang, Y.; Zhao, Y.; Tao, Y.; Cheng, C.; Yang, J.; Lu, X.; Hao, C.; Wang, T.; et al. PmiREN: A comprehensive encyclopedia of plant miRNAs. Nucleic Acids Res. 2020, 48, D1114–D1121. [Google Scholar] [CrossRef] [Green Version]
- Sunkar, R.; Jagadeeswaran, G. In silico identification of conserved microRNAs in large number of diverse plant species. BMC Plant Biol. 2008, 8, 37. [Google Scholar] [CrossRef] [Green Version]
- Axtell, M.J.; Bartel, D.P. Antiquity of microRNAs and their targets in land plants. Plant Cell 2005, 17, 1658–1673. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.; Fu, Y.R.; Ji, L.S.; Wu, C.A.; Zheng, C.C. Characterization and expression analysis of the Arabidopsis miR169 family. Plant Sci. 2010, 178, 271–280. [Google Scholar] [CrossRef]
- Rao, S.; Balyan, S.; Jha, S.; Mathur, S. Novel insights into expansion and functional diversification of MIR169 family in tomato. Planta 2020, 251, 55. [Google Scholar] [CrossRef]
- Petroni, K.; Kumimoto, R.W.; Gnesutta, N.; Calvenzani, V.; Fornari, M.; Tonelli, C.; Holt, B.F., 3rd; Mantovani, R. The promiscuous life of plant NUCLEAR FACTOR Y transcription factors. Plant Cell 2012, 24, 4777–4792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.E.; Repetti, P.P.; Adams, T.R.; Creelman, R.A.; Wu, J.; Warner, D.C.; Anstrom, D.C.; Bensen, R.J.; Castiglioni, P.P.; Donnarummo, M.G.; et al. Plant nuclear factor Y (NF-Y) B subunits confer drought tolerance and lead to improved corn yields on water-limited acres. Proc. Natl. Acad. Sci. USA 2007, 104, 16450–16455. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.J.; Yu, T.F.; Li, X.H.; Cao, X.Y.; Ma, J.; Chen, J.; Zhou, Y.B.; Chen, M.; Ma, Y.Z.; Zhang, J.H.; et al. Overexpression of GmNFYA5 confers drought tolerance to transgenic Arabidopsis and soybean plants. BMC Plant Biol. 2020, 20, 123. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.K.; Kim, H.I.; Jang, G.; Chung, P.J.; Jeong, J.S.; Kim, Y.S.; Bang, S.W.; Jung, H.; Choi, Y.D.; Kim, J.K. The NF-YA transcription factor OsNF-YA7 confers drought stress tolerance of rice in an abscisic acid independent manner. Plant Sci. 2015, 241, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Hu, Z.; Jiang, Q.; Zhang, H. GmNFYA3, a target gene of miR169, is a positive regulator of plant tolerance to drought stress. Plant Mol. Biol. 2013, 82, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.T.; Ge, L.F.; Liang, R.Q.; Li, W.; Ruan, K.C.; Lin, H.X.; Jin, Y.X. Members of miR-169 family are induced by high salinity and transiently inhibit the NF-YA transcription factor. BMC Mol. Biol. 2009, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, M.; Xu, M.; Lu, Y.; Zhang, L.; Fan, Y.; Wang, L. Expression of Zma-miR169 miRNAs and their target ZmNF-YA genes in response to abiotic stress in maize leaves. Gene 2015, 555, 178–185. [Google Scholar] [CrossRef]
- Du, Q.; Zhao, M.; Gao, W.; Sun, S.; Li, W.X. microRNA/microRNA* complementarity is important for the regulation pattern of NFYA5 by miR169 under dehydration shock in Arabidopsis. Plant J. 2017, 91, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Li, W.X.; Oono, Y.; Zhu, J.; He, X.J.; Wu, J.M.; Iida, K.; Lu, X.Y.; Cui, X.; Jin, H.; Zhu, J.K. The Arabidopsis NFYA5 transcription factor is regulated transcriptionally and posttranscriptionally to promote drought resistance. Plant Cell 2008, 20, 2238–2251. [Google Scholar] [CrossRef] [Green Version]
- Gyula, P.; Baksa, I.; Toth, T.; Mohorianu, I.; Dalmay, T.; Szittya, G. Ambient temperature regulates the expression of a small set of sRNAs influencing plant development through NF-YA2 and YUC2. Plant Cell Environ. 2018, 41, 2404–2417. [Google Scholar] [CrossRef]
- Eremina, M.; Rozhon, W.; Poppenberger, B. Hormonal control of cold stress responses in plants. Cell Mol. Life Sci. 2016, 73, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Zhang, L.; Xu, C.; Yuan, S.; Zhang, F.; Zheng, Y.; Zhao, C. Uncovering small RNA-mediated responses to cold stress in a wheat thermosensitive genic male-sterile line by deep sequencing. Plant Physiol. 2012, 159, 721–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Wang, G.; Sutoh, K.; Zhu, J.K.; Zhang, W. Identification of cold-inducible microRNAs in plants by transcriptome analysis. Biochim. Biophys Acta 2008, 1779, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Qi, L.; Li, Y.; Chu, J.; Li, C. PIF4-mediated activation of YUCCA8 expression integrates temperature into the auxin pathway in regulating Arabidopsis hypocotyl growth. PLoS Genet. 2012, 8, e1002594. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.H.; Domijan, M.; Klose, C.; Biswas, S.; Ezer, D.; Gao, M.; Khattak, A.K.; Box, M.S.; Charoensawan, V.; Cortijo, S.; et al. Phytochromes function as thermosensors in Arabidopsis. Science 2016, 354, 886–889. [Google Scholar] [CrossRef] [Green Version]
- Fiorucci, A.-S.; Galvão, V.C.; Ince, Y.Ç.; Boccaccini, A.; Goyal, A.; Allenbach Petrolati, L.; Trevisan, M.; Fankhauser, C. PHYTOCHROME INTERACTING FACTOR 7 is important for early responses to elevated temperature in Arabidopsis seedlings. New Phytol. 2020, 226, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhang, Y.; Kieffer, M.; Yu, H.; Kepinski, S.; Estelle, M. Corrigendum: HSP90 regulates temperature-dependent seedling growth in Arabidopsis by stabilizing the auxin co-receptor F-box protein TIR1. Nat. Commun. 2016, 7, 11677. [Google Scholar] [CrossRef]
- Bawa, G.; Feng, L.; Chen, G.; Chen, H.; Hu, Y.; Pu, T.; Cheng, Y.; Shi, J.; Xiao, T.; Zhou, W.; et al. Gibberellins and auxin regulate soybean hypocotyl elongation under low light and high-temperature interaction. Physiol. Plant 2020. [Google Scholar] [CrossRef]
- Nadella, V.; Shipp, M.J.; Muday, G.K.; Wyatt, S.E. Evidence for altered polar and lateral auxin transport in the gravity persistent signal (gps) mutants of Arabidopsis. Plant Cell Environ. 2006, 29, 682–690. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yan, S.; Yang, T.; Zhang, S.; Chen, Y.Q.; Liu, B. Small RNAs in regulating temperature stress response in plants. J. Integr. Plant Biol. 2017, 59, 774–791. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhang, X.; Su, Y.; Zou, J.; Wang, Z.; Xu, L.; Que, Y. miRNA alteration is an important mechanism in sugarcane response to low-temperature environment. BMC Genom. 2017, 18, 833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Wu, Z.; Jiang, F.; Zhou, R.; Yang, Z. Identification of chilling stress-responsive tomato microRNAs and their target genes by high-throughput sequencing and degradome analysis. BMC Genom. 2014, 15, 1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, X.; Zhang, L.; Li, G.; Wang, X.; Cao, X.; Fang, X.; Chen, F. Identification of novel stress-regulated microRNAs from Oryza sativa L. Genomics 2010, 95, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, A.; Bannigan, A.; Sulaman, W.; Pechter, P.; Blancaflor, E.B.; Baskin, T.I. Auxin, actin and growth of the Arabidopsis thaliana primary root. Plant J. 2007, 50, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Bussell, J.D.; Pacurar, D.I.; Schwambach, J.; Pacurar, M.; Bellini, C. Phenotypic plasticity of adventitious rooting in Arabidopsis is controlled by complex regulation of AUXIN RESPONSE FACTOR transcripts and microRNA abundance. Plant Cell 2009, 21, 3119–3132. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhang, B. MicroRNAs in control of plant development. J. Cell Physiol. 2016, 231, 303–313. [Google Scholar] [CrossRef]
- Fukaki, H.; Nakao, Y.; Okushima, Y.; Theologis, A.; Tasaka, M. Tissue-specific expression of stabilized SOLITARY-ROOT/IAA14 alters lateral root development in Arabidopsis. Plant J. 2005, 44, 382–395. [Google Scholar] [CrossRef]
- Kumar, R. Role of microRNAs in biotic and abiotic stress responses in crop plants. Appl. Biochem. Biotechnol. 2014, 174, 93–115. [Google Scholar] [CrossRef]
- Sunkar, R.; Zhu, J.K. Novel and stress-regulated microRNAs and other small RNAs from Arabidopsis. Plant Cell 2004, 16, 2001–2019. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Lin, L.; Sui, N. Regulation mechanism of microRNA in plant response to abiotic stress and breeding. Mol. Biol. Rep. 2019, 46, 1447–1457. [Google Scholar] [CrossRef]
- Sunkar, R.; Chinnusamy, V.; Zhu, J.; Zhu, J.K. Small RNAs as big players in plant abiotic stress responses and nutrient deprivation. Trends Plant Sci. 2007, 12, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Upadhyay, S.; Bhat, B.; Singh, G.; Bhattacharya, S.; Singh, A. Abiotic stress induced miRNA-TF-gene regulatory network: A structural perspective. Genomics 2020, 112, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.C.; Sieber, P.; Wellmer, F.; Meyerowitz, E.M. The early extra petals1 mutant uncovers a role for microRNA miR164c in regulating petal number in Arabidopsis. Curr. Biol. 2005, 15, 303–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Chan, Z.L. AtHAP5A modulates freezing stress resistance in Arabidopsis independent of the CBF pathway. Plant Signal. Behav. 2014, 9, e29109. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Ye, T.; Zhong, B.; Liu, X.; Jin, R.; Chan, Z. AtHAP5A modulates freezing stress resistance in Arabidopsis through binding to CCAAT motif of AtXTH21. New Phytol. 2014, 203, 554–567. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, L.; Lai, R.; Liu, W.; Chen, Y.; Zhang, Z.; XuHan, X.; Lai, Z. MicroRNA390-directed TAS3 cleavage leads to the production of tasiRNA-ARF3/4 during somatic embryogenesis in Dimocarpus longan Lour. Front. Plant Sci. 2015, 6, 1119. [Google Scholar] [CrossRef]
- He, F.; Xu, C.; Fu, X.; Shen, Y.; Guo, L.; Leng, M.; Luo, K. The MicroRNA390/TRANS-ACTING SHORT INTERFERING RNA3 module mediates lateral root growth under salt stress via the auxin pathway. Plant Physiol. 2018, 177, 775–791. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Wei, J.; Li, B.; Wang, M.; Bao, Y. AtVps11 is essential for vacuole biogenesis in embryo and participates in pollen tube growth in Arabidopsis. Biochem. Biophys. Res. Commun. 2017, 491, 794–799. [Google Scholar] [CrossRef]
- Kitakura, S.; Vanneste, S.; Robert, S.; Lofke, C.; Teichmann, T.; Tanaka, H.; Friml, J. Clathrin mediates endocytosis and polar distribution of PIN auxin transporters in Arabidopsis. Plant Cell 2011, 23, 1920–1931. [Google Scholar] [CrossRef] [Green Version]
- Jing, Y.; Cui, D.; Bao, F.; Hu, Z.; Qin, Z.; Hu, Y. Tryptophan deficiency affects organ growth by retarding cell expansion in Arabidopsis. Plant J. 2009, 57, 511–521. [Google Scholar] [CrossRef]
- Hermand, D. F-box proteins: More than baits for the SCF? Cell Div. 2006, 1, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feschotte, C. Transposable elements and the evolution of regulatory networks. Nat. Rev. Genet. 2008, 9, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogvadze, E.; Buzdin, A. Retroelements and their impact on genome evolution and functioning. Cell Mol. Life Sci. 2009, 66, 3727–3742. [Google Scholar] [CrossRef] [PubMed]
- Goodier, J.L.; Kazazian, H.H., Jr. Retrotransposons revisited: The restraint and rehabilitation of parasites. Cell 2008, 135, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Casacuberta, E.; Gonzalez, J. The impact of transposable elements in environmental adaptation. Mol. Ecol. 2013, 22, 1503–1517. [Google Scholar] [CrossRef]
- Chu, C.G.; Tan, C.T.; Yu, G.T.; Zhong, S.; Xu, S.S.; Yan, L. A novel retrotransposon inserted in the Dominant Vrn-B1 allele confers spring growth habit in tetraploid wheat (Triticum turgidum L.). G3 (Bethesda) 2011, 1, 637–645. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, A.; Liu, B.; Kong, F.; Arase, S.; Abe, J. Adaptive evolution involving gene duplication and insertion of a novel Ty1/copia-like retrotransposon in soybean. J. Mol. Evol. 2009, 69, 164–175. [Google Scholar] [CrossRef]
- Lin, R.; Ding, L.; Casola, C.; Ripoll, D.R.; Feschotte, C.; Wang, H. Transposase-derived transcription factors regulate light signaling in Arabidopsis. Science 2007, 318, 1302–1305. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Fu, D.; Li, C.; Blechl, A.; Tranquilli, G.; Bonafede, M.; Sanchez, A.; Valarik, M.; Yasuda, S.; Dubcovsky, J. The wheat and barley vernalization gene VRN3 is an orthologue of FT. Proc. Natl. Acad. Sci. USA 2006, 103, 19581–19586. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Jain, M. Genome-wide survey and comprehensive expression profiling of Aux/IAA gene family in chickpea and soybean. Front. Plant Sci. 2015, 6, 918. [Google Scholar] [CrossRef] [Green Version]
- Baskin, T.I.; Wilson, J.E. Inhibitors of protein kinases and phosphatases alter root morphology and disorganize cortical microtubules. Plant Physiol. 1997, 113, 493–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Li, L. miRDeep-P: A computational tool for analyzing the microRNA transcriptome in plants. Bioinformatics 2011, 27, 2614–2615. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Serivichyaswat, P.T.; Susila, H.; Ahn, J.H. Elongated hypocotyl 5-homolog (HYH) negatively regulates expression of the ambient temperature-responsive microRNA gene MIR169. Front. Plant Sci. 2017, 8, 2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No | Sample Name | Number of Reads | Average Length | Number of Reads after Trim | Percentage Trim | Average Length after Trim |
|---|---|---|---|---|---|---|
| 1 | Col-0 23 °C_Rep1 | 47,318,921 | 21.6 | 46,240,335 | 97.72% | 21.6 |
| 2 | Col-0 23 °C_Rep2 | 38,919,856 | 22.4 | 38,635,669 | 99.27 | 22.4 |
| 3 | Col-0 4 °C_Rep1 | 30,230,648 | 23.8 | 30,004,070 | 99.25% | 23.8 |
| 4 | Col-0 4 °C_Rep2 | 23,124,483 | 21.9 | 22,863,639 | 98.87% | 21.9 |
| 5 | slr1 23 °C_Rep1 | 22,247,497 | 21.4 | 21,471,561 | 95.51% | 21.4 |
| 6 | slr1 23 °C_Rep2 | 43,063,180 | 22.1 | 42,081,401 | 97.72% | 22.1 |
| 7 | slr1 4 °C_Rep1 | 25,807,408 | 22.9 | 24,711,139 | 95.75 | 22.4 |
| 8 | slr1 4 °C_Rep2 | 43,591,146 | 23.0 | 43,069,325 | 98.8% | 23.0 |
| Sample Name | Frequency Percentage of nt Length | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 21 nt | 24 nt | 26 nt | 27 nt | 28 nt | 29 nt | 30 nt | 31 nt | ||
| 1 | Col-0 23 °C | 32.76 | 13.50 | 3.46 | 2.25 | 1.37 | 0.71 | 0.56 | 0.41 |
| 2 | Col-0 4 °C | 42.49 | 13.95 | 3.02 | 1.31 | 0.69 | 0.30 | 0.16 | 0.06 |
| 3 | slr1 23 °C | 30.52 | 12.14 | 3.75 | 2.06 | 1.08 | 0.48 | 0.26 | 0.16 |
| 4 | slr1 4 °C | 20.12 | 10.31 | 6.72 | 4.42 | 4.00 | 2.97 | 2.68 | 2.43 |
| miR ID | Col-0 | slr1 | p-Value Col-0 vs. slr1 | |||||
|---|---|---|---|---|---|---|---|---|
| 23 °C | 4 °C | p-Value | 23 °C | 4 °C | p-Value | |||
| 23 °C vs. 4 °C | 23 °C vs. 4 °C | 23 °C | 4 °C | |||||
| miR156a,b,c,d,e,f, | 114.53 | 100.9 | 0.809 | 78.61 | 54.51 | 0.654 | 0.648 | 0.002 a |
| miR164b-3p | 22.5 | 13.47 | 0.611 | 3.64 | 2.79 | 0.447 | 0.338 | 0.006 a |
| miR169a-5p | 26.49 | 30.81 | 0.613 | 15.9 | 12.7 | 0.587 | 0.296 | 0.059 a |
| miR170,miR171a-5p | 4.7 | 2.96 | 0.638 | 3.17 | 0.92 | 0.051 | 0.652 | 0.27 |
| miR390a,b-5p | 1385 | 812.14 | 0.423 | 383.72 | 263.68 | 0.341 | 0.223 | 0.029 a |
| miR390b-3p | 65.18 | 49.73 | 0.583 | 25.47 | 16.48 | 0.254 | 0.237 | 0.029 a |
| miR396b-3p | 6.34 | 2.97 | 0.389 | 2.72 | 1.38 | 0.009 | 0.36 | 0.018 a |
| miR398a-5p | 0.56 | 0.48 | 0.854 | 2.87 | 1.88 | 0.603 | 0.297 | 0.013 a |
| miR399a | 4 | 3.15 | 0.165 | 5.65 | 2.31 | 0.049 | 0.148 | 0.224 |
| miR399b,c-3p | 25.8 | 23.93 | 0.845 | 17.41 | 10.81 | 0.027 | 0.413 | 0.029 a |
| miR408-5p | 14.55 | 33.15 | 0.249 | 42.01 | 37.83 | 0.684 | 0.034 | 0.764 |
| miR447a-3p,miR447b | 21.39 | 23 | 0.864 | 15.29 | 6.72 | 0.036 | 0.43 | 0.105 |
| miR472-3p | 379.33 | 154.02 | 0.539 | 63.04 | 45.1 | 0.241 | 0.41 | 0.04 a |
| miR5642a,b | 77.64 | 62.74 | 0.059 | 82.11 | 34 | 0.239 | 0.891 | 0.028 a |
| miR5656 | 1.31 | 1.33 | 0.982 | 0.81 | 0.18 | 0.044 | 0.235 | 0.257 |
| miR773a | 419.47 | 248.54 | 0.605 | 167.81 | 77.04 | 0.032 | 0.465 | 0.019 a |
| miR774a | 0.21 | 0.3 | 0.496 | 0.85 | 0.51 | 0.327 | 0.028 | 0.495 |
| miR775 | 26.11 | 15.41 | 0.621 | 9.78 | 5.68 | 0.266 | 0.472 | 0.04 a |
| miR8180 | 0.85 | 0.5 | 0.257 | 0.83 | 0.13 | 0.048 | 0.914 | 0.257 |
| miR8181 | 1.49 | 2.08 | 0.567 | 1.98 | 0.73 | 0.018 | 0.25 | 0.243 |
| miR824-3p | 177.08 | 365.65 | 0.267 | 99.59 | 171 | 0.037 | 0.484 | 0.149 |
| miR850 | 8.16 | 6.72 | 0.685 | 3.71 | 1.52 | 0.103 | 0.017 | 0.238 |
| miR852 | 3.7 | 1.72 | 0.468 | 1.15 | 0.9 | 0.384 | 0.372 | 0.058 a |
| miR ID | Col-0 | slr1 | p-Value Col-0 vs. slr1 | |||||
|---|---|---|---|---|---|---|---|---|
| 23 °C | 4 °C | p-Value | 23 °C | 4 °C | p-Value | |||
| 23 °C vs. 4 °C | 23 °C vs. 4 °C | 23 °C | 4 °C | |||||
| Pred_7 | 30.68 | 16.18 | 0.521 | 36.20 | 69.86 | 0.033 | 0.84 | 0.015 a |
| Pred_9 | 27.75 | 21.95 | 0.483 | 17.38 | 12.98 | 0.274 | 0.294 | 0.007 a |
| Pred_11 | 10.06 | 6.75 | 0.637 | 15.59 | 21.14 | 0.289 | 0.506 | 0.018 a |
| Pred_19 | 10.24 | 6.64 | 0.271 | 6.71 | 4.67 | 0.434 | 0.383 | 0.0031 a |
| Pred_25 | 3.38 | 1.59 | 0.185 | 2.15 | 0.98 | 0.002 | 0.305 | 0.011 a |
| Pred_26 | 6.32 | 5.14 | 0.647 | 2.90 | 1.74 | 0.503 | 0.304 | 0.056 a |
| Pred_27 | 4.01 | 4.49 | 0.774 | 3.03 | 2.17 | 0.302 | 0.040 | 0.280 |
| Pred_35 | 2.80 | 1.26 | 0.099 | 1.49 | 0.48 | 0.071 | 0.029 | 0.294 |
| Pred_36 | 2.19 | 2.92 | 0.755 | 3.08 | 1.81 | 0.039 | 0.107 | 0.638 |
| Pred_37 | 2.30 | 1.41 | 0.007 | 3.57 | 2.38 | 0.247 | 0.010 | 0.313 |
| Pred_44 | 1.47 | 0.78 | 0.015 | 1.01 | 0.43 | 0.466 | 0.551 | 0.076 |
| Pred_47 | 0.99 | 0.66 | 0.387 | 0.30 | 0.33 | 0.827 | 0.012 | 0.411 |
| Pred_53 | 0.64 | 1.03 | 0.743 | 1.20 | 1.27 | 0.941 | 0.058 | 0.868 |
| Pred_60 | 0.31 | 0.33 | 0.835 | 0 | 0 | 0 | 0.000 | 0.040 a |
| Pred_62 | 0.22 | 0.14 | 0.060 | 0.001 | 0.147 | 0.116 | 0.001 | 0.983 |
| Pred_65 | 0.26 | 0.28 | 0.966 | 1.21 | 0.647 | 0.475 | 0.015 | 0.658 |
| miR ID | Target Locus |
|---|---|
| miR156a,b,c,d,e,f, | SQUAMOSA promoter-binding protein-like 15 (AT3G57920), SQUAMOSA promoter-like 11 (AT1G27360), SPL13B (AT5G50670), SPL9 (AT2G42200), SPL2 (AT5G43270), SPL10 (AT1G27370), SPL13A (AT5G50570), SPL10 (AT1G27370), SBP domain transcription factor (AT1G69170), SPL3 (AT2G33810), protein kinase superfamily protein (AT3G28690), SPL4 (AT1G53160), SPL5 (AT3G15270), transposable element gene (AT1G16660), cysteine/histidine-rich C1 domain family protein (AT2G21840) |
| miR164b-3p | PPR repeat protein (AT5G14770) |
| miR169a-5p | Nuclear factor Y, subunit A8 (AT1G17590, AT1G54160) |
| miR170, miR171a-5p | Transposable element gene (AT2G06790, AT3G30393, AT1G36470, AT1G50860, AT3G29732, AT2G12305), MATE efflux family protein (AT1G15180), F-box family protein (AT5G3946) |
| miR390a,b-5p | TASIR-ARF (AT5G57735), transmembrane kinase-like 1 (AT3G24660) |
| miR390b-3p | Vacuolar protein sorting 11 (AT2G05170), RNA-binding (RRM/RBD/RNP motifs) family protein (AT3G07810), transposable element gene (AT2G41570, AT1G35990), clathrin heavy chain (AT3G11130) |
| miR396b-3p | MYB76 (AT5G07700), ATBTAF1 (AT3G54280), RNA helicase family protein (AT1G58060) |
| miR398a-5p | SETH2, UDP-glycosyltransferase superfamily protein (AT3G45100); alpha/beta-hydrolases superfamily protein (AT3G48080); ARM repeat superfamily protein (AT5G06120) |
| miR399a | PHO2/UBC24 (AT2G33770), CYP705A30 (AT3G20940), terpenoid cyclases family protein (AT1G78500), sodium bile acid symporter family (AT2G26900), transposable element gene (AT3G43867), nucleic acid-binding (AT1G27750) |
| miR399b,c-3p | PHO2 (AT2G33770), wall-associated kinase 2 (AT1G21270) |
| miR408-5p | Oxidoreductase, 2OG-Fe(II) oxygenase (AT4G02940, AT4G25310), nucleotide/sugar transporter family (AT4G03950), glutathione S-transferase TAU 25 (AT1G17180), leucine-rich repeat protein kinase family protein (AT1G04210), unknown protein (AT4G37030), pseudogene (AT2G31860), calmodulin-binding protein-related (AT5G10660), early-responsive to dehydration stress protein (AT4G35870) |
| miR447a-3p, miR447b | P-loop containing nucleoside triphosphate (AT5G60760), FAD-dependent oxidoreductase family protein (AT2G22650), lncRNA (AT5G05905) |
| miR472-3p | Disease resistance protein (CC-NBS-LRR class) family (AT5G43740, AT1G12290, AT1G15890); RPS5, disease resistance protein (CC-NBS-LRR class) family (AT1G12220) |
| miR5642a,b | Tryptophan synthase beta-subunit 1 (AT5G54810); VHA-E3, vacuolar H+-ATPase subunit E isoform 3 (AT1G64200) |
| miR5656 | Mitochondrial editing factor 9 (AT1G62260), zinc finger (C3HC4-type RING finger) family protein (AT5G60710), transposable element gene (AT3G29787) |
| miR773a | Remorin family protein (AT3G57540), root FNR 1 (AT4G05390), Acyl-CoA N-acyltransferase with RING/FYVE/PHD-type zinc finger protein (AT4G14920), transposable element gene (transposable element gene), RAD3-like DNA-binding helicase protein (AT1G79950), oleosin3 (AT5G51210) |
| miR774a | F-box and associated interaction domains-containing protein (AT3G17490), F-box family protein (AT3G19890), transposable element gene (AT1G34405, AT2G01024, AT4G16910, AT3G42996, AT2G07660) |
| miR775 | Dicer-like 1 (AT1G01040), galactosyltransferase family protein (AT1G53290), galactosyltransferase family protein (AT1G53290) |
| miR8180 | Alpha/beta-hydrolases superfamily protein (AT3G55190), plastid division2 (AT2G16070), ATP-dependent helicase family protein (AT2G28240), lncRNA (AT3G08825), fatty acid reductase 1 (AT5G22500), |
| miR8181 | Ovate family protein (AT2G36026), cysteine/histidine-rich C1 domain family protein (AT1G55430), LOB domain-containing protein 39 (AT4G37540), transposable element gene (AT3G30713) |
| miR824-3p | Concanavalin A-like lectin protein kinase family protein (AT3G08870), pentatricopeptide repeat (PPR)-containing protein (AT5G27300) |
| miR850 | Chloroplast RNA binding (AT1G09340), threonyl-tRNA synthetase (AT2G04842), lncRNA (AT2G08250), AtSWEET4 (AT3G28007) |
| miR852 | IAA-leucine resistant (ILR)-like 1(AT5G56650); IAA-leucine resistant (ILR)-like 2 (AT5G56660); TIR1, F-box/RNI-like superfamily protein (AT3G62980); K+ transporter 1 (AT2G26650); H(+)-ATPase 11 (AT5G62670) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslam, M.; Sugita, K.; Qin, Y.; Rahman, A. Aux/IAA14 Regulates microRNA-Mediated Cold Stress Response in Arabidopsis Roots. Int. J. Mol. Sci. 2020, 21, 8441. https://doi.org/10.3390/ijms21228441
Aslam M, Sugita K, Qin Y, Rahman A. Aux/IAA14 Regulates microRNA-Mediated Cold Stress Response in Arabidopsis Roots. International Journal of Molecular Sciences. 2020; 21(22):8441. https://doi.org/10.3390/ijms21228441
Chicago/Turabian StyleAslam, Mohammad, Kenji Sugita, Yuan Qin, and Abidur Rahman. 2020. "Aux/IAA14 Regulates microRNA-Mediated Cold Stress Response in Arabidopsis Roots" International Journal of Molecular Sciences 21, no. 22: 8441. https://doi.org/10.3390/ijms21228441
APA StyleAslam, M., Sugita, K., Qin, Y., & Rahman, A. (2020). Aux/IAA14 Regulates microRNA-Mediated Cold Stress Response in Arabidopsis Roots. International Journal of Molecular Sciences, 21(22), 8441. https://doi.org/10.3390/ijms21228441