Synthesis of Kisspeptin-Mimicking Fragments and Investigation of their Skin Anti-Aging Effects

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
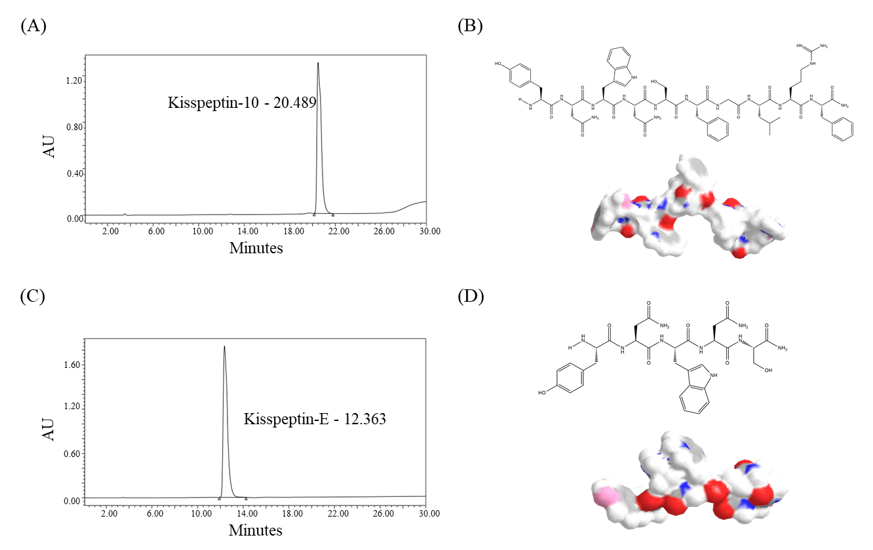
2.1. Characterization of Kisspeptin-Mimicking Peptides
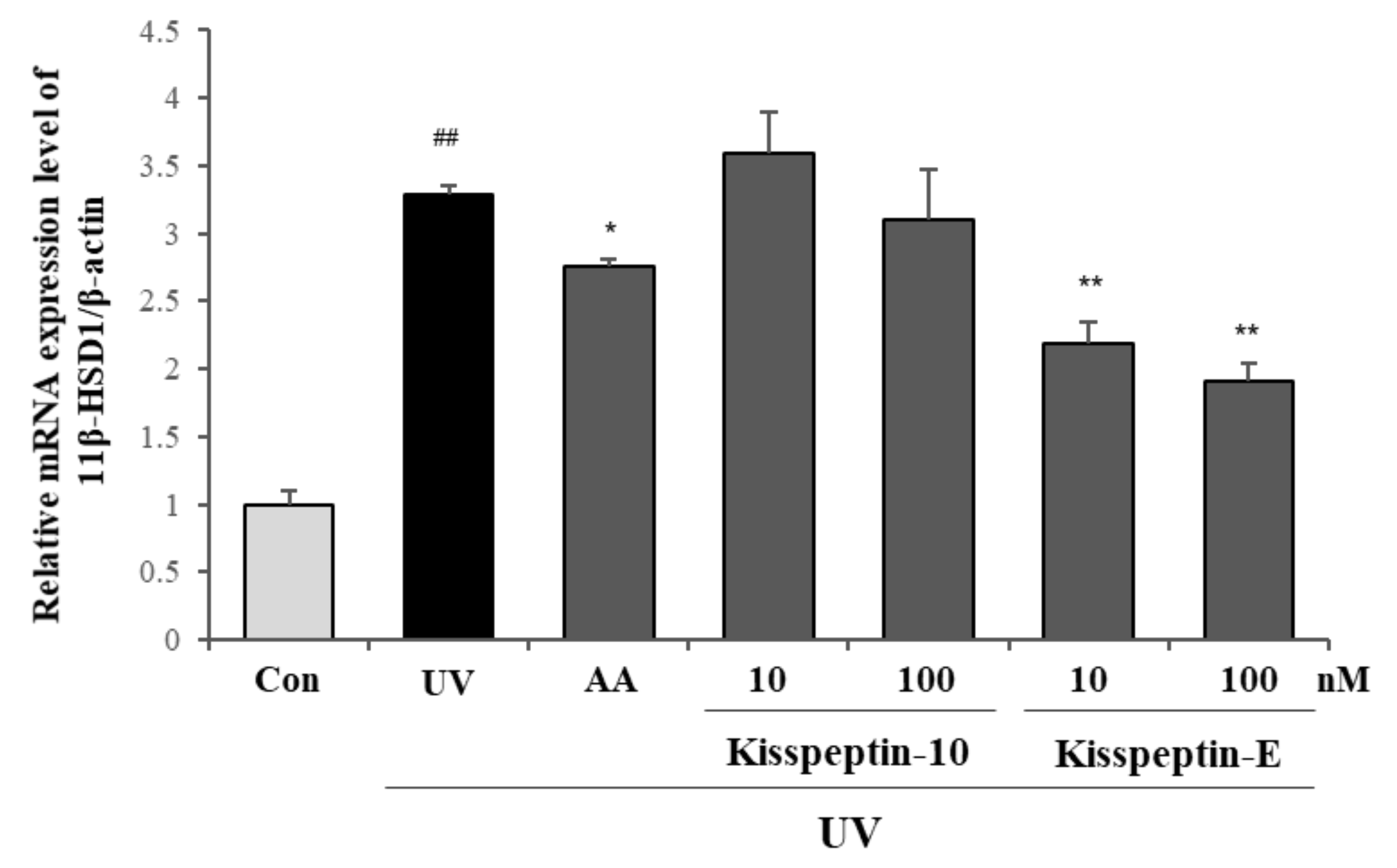
2.2. Inhibitory Effects of Kisspeptin-Mimicking Peptides on 11β-HSD1 Expression
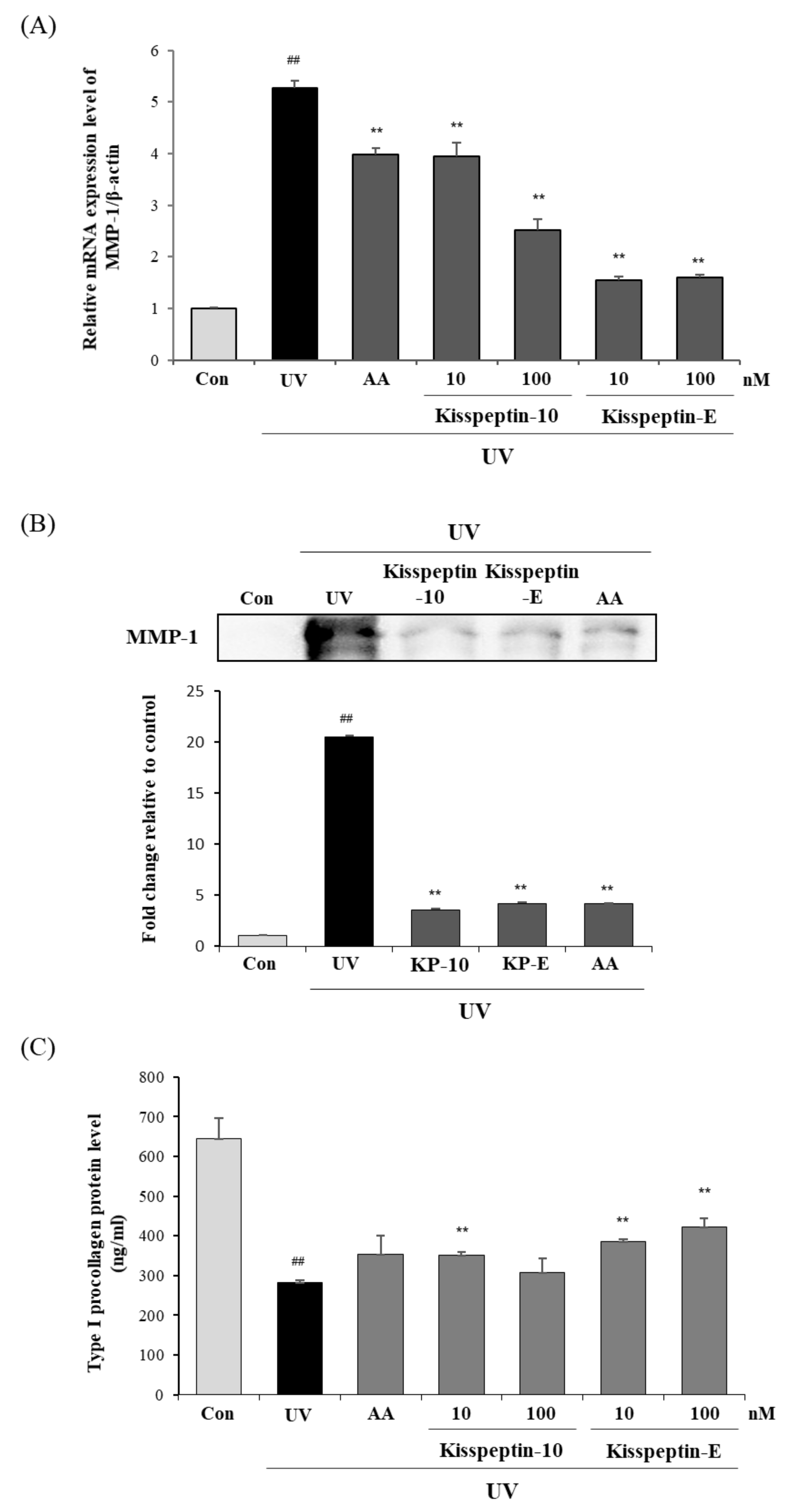
2.3. The Effects of Kisspeptin-Mimicking Peptides on MMP-1 and Type I Procollagen Expression
2.4. Inhibitory Effects of Kisspeptin-Mimicking Peptides on IL-6 and IL-8 Expression
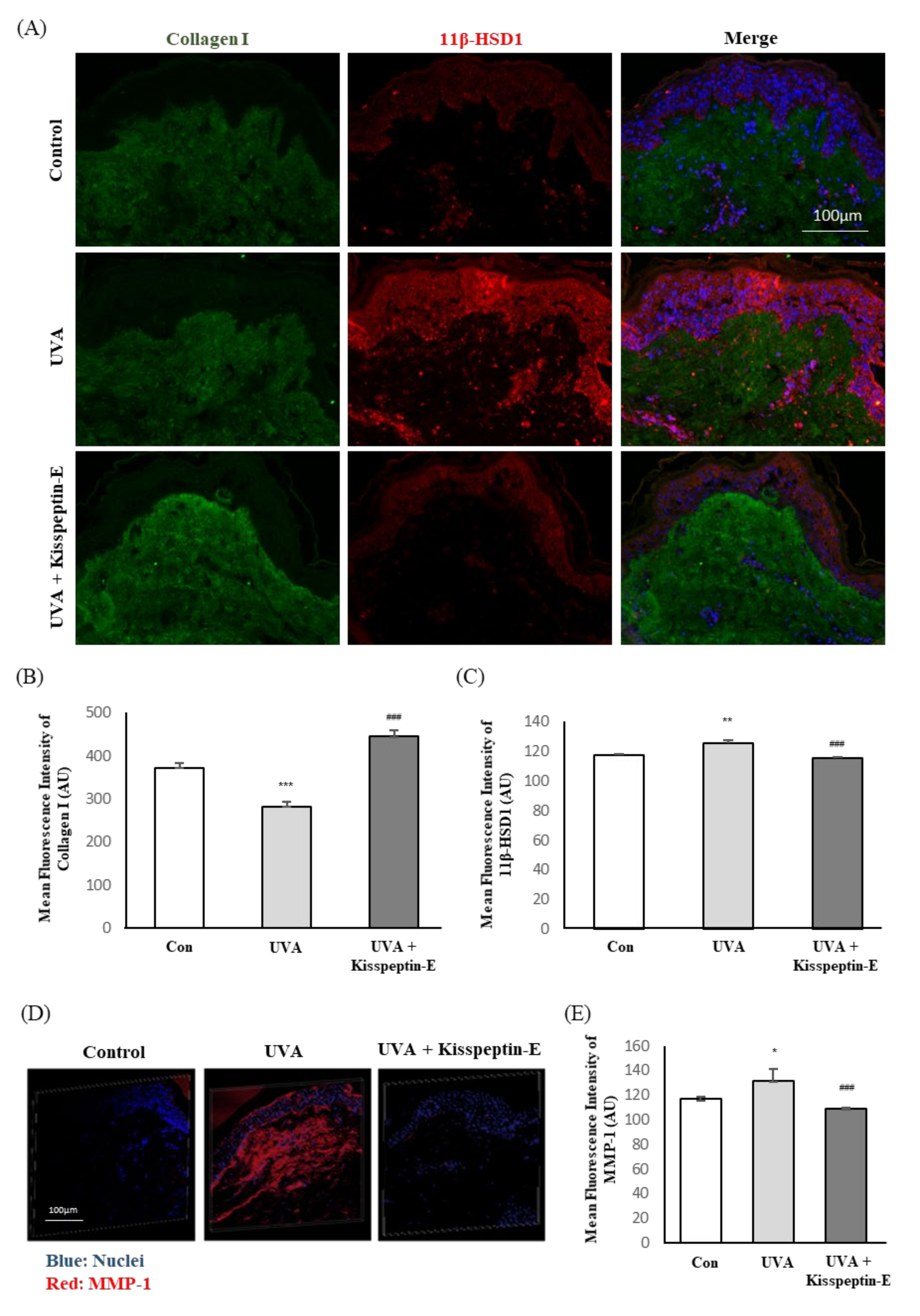
2.5. Regulatory Effects of Kisspeptin-E on Dermis Proteins in Human Skin
2.6. Recovery Effects of Kisspeptin-E on Epidermis Thickness in Human Skin
3. Discussion
4. Materials and Methods
4.1. Synthesis of Kisspeptin-Mimicking Peptides
4.2. Characterization of Kisspeptin-Mimicking Peptides
4.3. Cell Culture and UVB Irradiation
4.4. RT-qPCR
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Immunoblotting Analysis
4.7. Human Skin Explants and UVA Irradiation
4.8. Immunostaining in Human Skin
4.9. Measurements of Epidermis Thickness
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Ascorbic acid |
| ATCC | American Type Culture Collection |
| DCM | Dichloromethane |
| DIC | N, N’-diisopropyl carbodiimide |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMF | Dimethylformamide |
| DMSO | Dimethylsulfoxide |
| DPBS | Dulbecco’s phosphate buffered saline |
| ECM | Extracellular matrix |
| ELISA | Enzyme-linked immunosorbent assay |
| FMOC | 9-fluorenylmethoxycarbonyl |
| GC | Glucocorticoids |
| GnRH | Gonadotropin-releasing hormone |
| GR | Glucocorticoid receptor |
| HOBt | N-hydroxybenzotriazole |
| HPA | Hypothalamic-pituitary-adrenal |
| HPG | Hypothalamic-pituitary-gonadal |
| HPLC | High performance liquid chromatograph |
| IL-6 | Interleukin-6 |
| MALDI-TOF | Matrix Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry |
| MMP-1 | Matrix metalloproteinase-1 |
| MW | Molecular weight |
| PBS | Phosphate buffered saline |
| RIPA | Radioimmunoprecipitation assay |
| RF | Arginine phenylalanine |
| RT-qPCR | Quantitative real-time polymerase chain reaction |
| TBST | Tris-Buffered Saline with Tween 20 |
| UV | Ultraviolet |
| 11β-HSD1 | 11 beta-hydroxysteroid dehydrogenase type 1 |
References
- El-Domyati, M.; Attia, S.; Saleh, F.; Brown, D.; Birk, D.E.; Gasparro, F.; Ahmad, H.; Uitto, J. Intrinsic aging vs. photoaging: A comparative histopathological, immunohistochemical, and ultrastructural study of skin. Exp. Dermatol. 2002, 11, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lyga, J. Brain-skin connection: Stress, inflammation and skin aging. Inflamm. Allergy Drug Targets 2014, 13, 177–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.T.; Manna, P.R.; Tuckey, R.C. On the role of skin in the regulation of local and systemic steroidogenic activities. Steroids 2015, 103, 72–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skobowiat, C.; Dowdy, J.C.; Sayre, R.M.; Tuckey, R.C.; Slominski, A. Cutaneous hypothalamic-pituitary-adrenal axis homolog: Regulation by ultraviolet radiation. Am. J. Physiol. Metab. 2011, 301, E484–E493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terao, M.; Tani, M.; Itoi, S.; Yoshimura, T.; Hamasaki, T. 11β-Hydroxysteroiddehydrogenase 1 specific inhibitor increased dermal collagen content and promotes fibroblast proliferation. PLoS ONE 2014, 9, e93051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiganescu, A.; Tahrani, A.A.; Morgan, S.A.; Otranto, M.; Desmoulière, A.; Abrahams, L.; Hassan-Smith, Z.; Walker, E.A.; Rabbitt, E.H.; Cooper, M.S.; et al. 11β-Hydroxysteroid dehydrogenase blockade prevents age-induced skin structure and function defects. J. Clin. Investig. 2013, 123, 3051–3060. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.-J.; Min, J.-E.; Son, M.-H.; Oh, J.-H.; Kang, S. Ultraviolet- and infrared-induced 11 beta-hydroxysteroid dehydrogenase type 1 activating skin photoaging is inhibited by red ginseng extract containing high concentration of ginsenoside Rg3(S). Photodermatol. Photoimmunol. Photomed. 2017, 33, 311–320. [Google Scholar] [CrossRef]
- Zhang, L.; Falla, T.J. Cosmeceuticals and peptides. Clin. Dermatol. 2009, 27, 485–494. [Google Scholar] [CrossRef]
- Katayama, K.; Armendariz-Borunda, J.; Raghow, R.; Kang, A.H.; Seyer, J.M. A pentapeptide from type I procollagen promotes extracellular matrix production. J. Biol. Chem. 1993, 268, 9941–9944. [Google Scholar]
- Kim, D.J.; Chang, S.S.; Lee, J. Anti-aging potential of substance P-based hydrogel for human skin longevity. Int. J. Mol. Sci. 2019, 20, 4453. [Google Scholar] [CrossRef] [Green Version]
- Skorupskaite, K.; George, J.T.; Anderson, R.A. The kisspeptin-GnRH pathway in human reproductive health and disease. Hum. Reprod. Updat. 2014, 20, 485–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinilla, L.; Aguilar, E.; Dieguez, C.; Millar, R.P.; Tena-Sempere, M. Kisspeptins and reproduction: Physiological roles and regulatory mechanisms. Physiol. Rev. 2012, 92, 1235–1316. [Google Scholar] [CrossRef] [PubMed]
- Roa, J.; Aguilar, E.; Dieguez, C.; Pinilla, L.; Tena-Sempere, M. New frontiers in kisspeptin/GPR54 physiology as fundamental gatekeepers of reproductive function. Front. Neuroendocr. 2008, 29, 48–69. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.-M.; Butler, J.P.; Pinnell, N.E.; Pralong, F.P.; Crowley, W.F.; Ren, C.; Chan, K.K.; Seminara, S.B. Kisspeptin resets the hypothalamic GnRH clock in men. J. Clin. Endocrinol. Metab. 2011, 96, E908–E915. [Google Scholar] [CrossRef] [Green Version]
- Dhillo, W.S.; Chaudhri, O.B.; Patterson, M.; Thompson, E.L.; Murphy, K.G.; Badman, M.K.; McGowan, B.M.; Amber, V.; Patel, S.; Ghatei, M.A.; et al. Kisspeptin-54 stimulates the hypothalamic-pituitary gonadal axis in human males. J. Clin. Endocrinol. Metab. 2005, 90, 6609–6615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calley, J.L.; Dhillo, W.S. Effects of the Hormone Kisspeptin on Reproductive Hormone Release in Humans. Adv. Biol. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Kotani, M.; Detheux, M.; Vandenbogaerde, A.; Communi, D.; Vanderwinden, J.-M.; Le Poul, E.; Brézillon, S.; Tyldesley, R.; Suarez-Huerta, N.; Vandeput, F.; et al. The metastasis suppressor gene KiSS-1 encodes Kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. J. Biol. Chem. 2001, 276, 34631–34636. [Google Scholar] [CrossRef] [Green Version]
- Milton, N.G.; Chilumuri, A.; Rocha-Ferreira, E.; Nercessian, A.N.; Ashioti, M. Kisspeptin prevention of amyloid-beta peptide neurotoxicity in vitro. ACS Chem. Neurosci. 2012, 3, 706–718. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Miele, M.E.; Hicks, D.J.; Phillips, K.K.; Trent, J.M.; Weissman, B.E.; Welch, D.R. KiSS-1, a Novel human malignant melanoma metastasis-suppressor gene. J. Natl. Cancer Inst. 1996, 88, 1731–1737. [Google Scholar] [CrossRef]
- Ohtaki, T.; Shintani, Y.; Honda, S.; Matsumoto, H.; Hori, A.; Kanehashi, K.; Terao, Y.; Kumano, S.; Takatsu, Y.; Masuda, Y.; et al. Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nat. Cell Biol. 2001, 411, 613–617. [Google Scholar] [CrossRef]
- Comninos, A.N.; Wall, M.B.; Demetriou, L.; Shah, A.J.; Clarke, S.A.; Narayanaswamy, S.; Nesbitt, A.; Izzi-Engbeaya, C.; Prague, J.K.; Abbara, A.; et al. Kisspeptin modulates sexual and emotional brain processing in humans. J. Clin. Investig. 2017, 127, 709–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, C.; Wang, H.; Boyd, D.D. Kiss-1 repressses 92-kDa Type IV collagenase expression by down-regulating NF-kB binding to the promoter as a consequence of IkB-induced block of p65/p50 nuclear translocation. J. Biol. Chem. 2001, 276, 1164–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: Roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation—A review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Abadie, S.; Bedos, P.; Rouquette, J. A human skin model to evaluate the protective effect of compounds against UVA damage. Int. J. Cosmet. Sci. 2019, 41, 594–603. [Google Scholar] [CrossRef]
- Harter, C.J.L.; Kavanagh, G.S.; Smith, J.T. The role of kisspeptin neurons in reproduction and metabolism. J. Endocrinol. 2018, 238, R173–R183. [Google Scholar] [CrossRef] [Green Version]
- De Roux, N.; Genin, E.; Carel, J.C.; Matsuda, F.; Chaussain, J.L.; Milgrom, E. Hypogonadotroic hypogonadism due to loss of function of the KISS-1 dericed peptide receptor GPR54. Proc. Natl. Acad. Sci. USA 2003, 100, 10972–10976. [Google Scholar] [CrossRef] [Green Version]
- Seminara, S.B.; Messager, S.; Chatzidaki, E.E.; Thresher, R.R.; Acierno, J.S.; Shagoury, J.K.; Bo-Abbas, Y.; Kuohung, W.; Schwinof, K.M.; Hendrick, A.G.; et al. The GPR54 gene as a regulator of puberty. N. Engl. J. Med. 2003, 349, 1614–1627. [Google Scholar] [CrossRef] [Green Version]
- Heinrichs, M.; Domes, G. Neuropeptides and social behavior: Effects of oxytocin and vasopressin in humans. Prog. Brain Res. 2008, 170, 337–350. [Google Scholar]
- Deing, V.; Roggenkamp, D.; Kühnl, J.; Gruschka, A.; Stäb, F.; Wenck, H.; Bürkle, A.; Neufang, G. Oxytocin modulates proliferation and stress responses of human skin cells: Implications for atopic dermatitis. Exp. Dermatol. 2013, 22, 399–405. [Google Scholar] [CrossRef] [Green Version]
- Roh, K.B.; Park, D.; Jung, E. Inhibitory effects of prunella vulgaris L. extract on 11𝛽-HSD1 in human skin cells. Evid. Based Complement. Altern. Med. 2018, 2018, 1–8. [Google Scholar]
- Wlaschek, M.; Bolsen, K.; Herrmann, G.; Schwarz, A.; Wilmroth, F.; Heinrich, P.C.; Goerz, G.; Scharffetter-Kochanek, K. UVA-induced autocrine stimulation of fibroblast-derived-collagenase by IL-6: A possible mechanism in dermal photodamage? J. Investig. Dermatol. 1993, 101, 164–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoepe, S.; Schacke, H.; May, E.; Asadullah, K. Glucocorticoid therapy-induced skin atrophy. Exp. Dermatol. 2006, 15, 406–420. [Google Scholar] [CrossRef] [PubMed]
- Kolbe, L.; Kligman, A.M.; Schreiner, V.; Stoudemayer, T. Corticosteroid-induced atrophy and barrier impairment measured by non-invasive methods in human skin. Skin Res. Technol. 2001, 7, 73–77. [Google Scholar] [CrossRef] [PubMed]
- King, D.; Fields, C.G.; Fields, G.B. A cleavage method which minimizes side reactions following Fmoc solid phase peptide synthesis. Int. J. Pept. Protein Res. 1990, 36, 255–266. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.-E.; Jeong, S.; Yun, S.K.; Kyung, S.; Sophie, A.; Moh, S.H.; Seo, H.H.; Park, M.S.; Kang, S.; Yeo, H. Synthesis of Kisspeptin-Mimicking Fragments and Investigation of their Skin Anti-Aging Effects. Int. J. Mol. Sci. 2020, 21, 8439. https://doi.org/10.3390/ijms21228439
Lee K-E, Jeong S, Yun SK, Kyung S, Sophie A, Moh SH, Seo HH, Park MS, Kang S, Yeo H. Synthesis of Kisspeptin-Mimicking Fragments and Investigation of their Skin Anti-Aging Effects. International Journal of Molecular Sciences. 2020; 21(22):8439. https://doi.org/10.3390/ijms21228439
Chicago/Turabian StyleLee, Kyung-Eun, Sugyeong Jeong, Seok Kyun Yun, Seoyeon Kyung, Abadie Sophie, Sang Hyun Moh, Hyo Hyun Seo, Myeong Sam Park, Seunghyun Kang, and Hyeonju Yeo. 2020. "Synthesis of Kisspeptin-Mimicking Fragments and Investigation of their Skin Anti-Aging Effects" International Journal of Molecular Sciences 21, no. 22: 8439. https://doi.org/10.3390/ijms21228439
APA StyleLee, K. -E., Jeong, S., Yun, S. K., Kyung, S., Sophie, A., Moh, S. H., Seo, H. H., Park, M. S., Kang, S., & Yeo, H. (2020). Synthesis of Kisspeptin-Mimicking Fragments and Investigation of their Skin Anti-Aging Effects. International Journal of Molecular Sciences, 21(22), 8439. https://doi.org/10.3390/ijms21228439